增强羊栖菜多糖生物活性的方法与流程
本发明属于化学化工技术领域,涉及一种通过对羊栖菜多糖进行酶解从而增强其生物活性的方法。
背景技术:
羊栖菜中含有各种具有健康益处的生物活性成分,尤其是其中的酸性多糖,具有抗病毒,抗真菌,抗菌,抗氧化,抗炎,降血脂和抗肿瘤特性。由于羊栖菜价格低廉,分布广泛,因此将羊栖菜加工为功能性食品或开发成低副作用的药物具有良好的研究前景。羊栖菜中碳水化合物的含量较高,但其中的粗多糖的生物活性相对偏低。多糖的生物活性与其分子结构密切相关,包括糖苷键、单糖组成、硫酸基含量和分子量等。高分子量多糖会由于水溶性较低,从而阻碍它们渗透到细胞中以执行给定的功能,而较低分子量的硫酸多糖则显示出更高的生物活性。由于活性多糖经不同方法降解得到的多糖的功能特性也不同,因此选取恰当的提取与降解方法以更大程度地提高多糖的各种生物活性是非常重要的。目前现有的羊栖菜多糖的降解方法多为氧化降解法、酸降解法,采用酶法降解羊栖菜多糖则鲜见报道。
技术实现要素:
本发明要解决的技术问题是提供一种新的提高羊栖菜降解多糖生物活性方法。
为了解决上述技术问题,提供一种增强羊栖菜多糖生物活性的方法(对羊栖菜粗多糖进行复合酶降解的方法),包括以下步骤:
1)、降解:
先将羊栖菜粗多糖(sfp)溶于ph为4.5~7.0的缓冲溶液,接着加入由果胶酶和糖化酶组成的复合酶,再加入缓冲溶液进行稀释,所得的体系中羊栖菜粗多糖(sfp)的浓度为2~5mg/ml、复合酶总浓度为20~80u/ml;所述果胶酶:糖化酶的活力比=6:0~0:6;
将所述体系于40~65℃的温度下振荡反应1~5h,从而实现对羊栖菜粗多糖的降解;
2)、将步骤1)所得的反应液除酶(高温除酶)后过滤,所得滤液浓缩至原体积的20~30%(即,约1/4);用截留分子量为3500da的透析袋透析24~48h,将所得的透析液(留在透析袋中的溶液)浓缩,冷冻干燥(-50~-60℃浓缩干燥至恒重),得到羊栖菜复合酶解多糖(esfp)。
作为本发明的增强羊栖菜多糖生物活性的方法的改进:
所得的体系中羊栖菜粗多糖(sfp)的浓度为4.0mg/ml、复合酶总浓度为68.4u/ml;所述果胶酶:糖化酶的活力比=3.3:1,缓冲溶液的ph为6.0;
于54.2℃的温度下振荡反应3h;得到羊栖菜复合酶解多糖esfp1-1。
esfp1-1的分子量为86.8kda。其与未降解的羊栖菜多糖(sfp)相比,其氧化活性、胆酸结合活性及免疫调节作用得到显著提高。
作为本发明的增强羊栖菜多糖生物活性的方法的进一步改进:
所述ph为4.5~7.0的缓冲溶液是ph为4.5~7.0的乙酸-乙酸钠缓冲溶液、柠檬酸–柠檬酸钠缓冲液。
作为本发明的增强羊栖菜多糖生物活性的方法的进一步改进:将羊栖菜复合酶解多糖esfp1-1进行分离纯化;包括以下步骤:
①、粗分:
选用cellulosedeae-52层析柱(规格为60cm×φ2.6cm,有效长度为50cm);
称取400±40mg羊栖菜复合酶解多糖esfp1-1,溶于7ml的去离子水中离心取上清液,将上清液均匀的加入到层析柱填料上表面,依次用去离子水、以及浓度分别为0.1mol/l、0.3mol/l、0.5mol/l、0.8mol/l的nacl溶液洗脱(当检测到洗脱剂对应的洗脱液中不含有多糖时,则换下一种洗脱剂进行洗脱;可采用硫酸-苯酚法隔管检测其在490nm处的紫外吸收值),流速为1ml/min,
分别收集0.1mol/l、0.3mol/l、0.5mol/l、0.8mol/l的nacl溶液这4种洗脱剂各自对应的洗脱液,减压浓缩、透析脱盐、冷冻干燥,从而对应得到羊栖菜复合酶解多糖的deae-52的四种初步纯化的多糖组分;
②、细分:
选用sephadexg-100凝胶色谱柱(层析柱规格为60cm×φ1.6cm,柱子的有效体积为50cm×φ1.6cm);
将以上经过cellulosedeae-52粗分的四种初步纯化的多糖组分分别进行以下操作:
称取100±10mg初步纯化的多糖组分溶于3ml去离子水中,离心取上清液,加样于sephadexg-100凝胶色谱柱中,用去离子水洗脱,流速为0.20ml/min,合并多糖洗脱液(可先用硫酸-苯酚法逐管检测在490nm处紫外吸收值,绘制sephadexg-100色谱柱洗脱曲线,一般每管20min),减压浓缩、冷冻干燥得到羊栖菜复合酶解多糖的四个纯化组分(纯化多糖),分别命名为esfp1、esfp2、esfp3和esfp4。
它们的分子量分别为75.5、86.3、76.2和86.5kda。
本发明还同时提供了依照上述方法所得的降解羊栖菜多糖、纯化多糖在制备功能性食品基料中的应用。
本发明采用cellulosedeae-52柱层析法和sephadexg-100凝胶色谱法对羊栖菜酶解多糖(esfp)进行分离纯化,以得到活性更好的纯化多糖组分。
本发明所得到的羊栖菜酶解多糖与作为原料的羊栖菜粗多糖(sfp)的组成和分子量数据对比如表1所述。
本发明具有如下技术效果:
1.与未降解的羊栖菜粗多糖(sfp)相比,本发明得到的羊栖菜酶解多糖(esfp)的体外抗氧化活性、胆酸结合活性和raw264.7巨噬细胞的免疫活性(对巨噬细胞的增殖、吞噬与释放no分子起到显著促进作用)显著提高。
本发明首次采用果胶酶和糖化酶复合酶法降解羊栖菜多糖;酶法降解具有作用条件温和、对多糖上的硫酸基等取代基影响小等优点。
2.对羊栖菜降解多糖(espy)进行分离纯化后得到的四个分离纯化组分(esfp1、esfp2、esfp3、esfp4),都具有更好的抗氧化活性和raw264.7巨噬细胞的免疫活性。
综上所述,采用本发明的方法不但能显著提高羊栖菜多糖的抗氧化活性与胆酸结合能力(与胆汁酸相结合,能加快体内胆固醇分解,有效降低人体血清以及肝中胆固醇的含量从而达到降血脂的作用效果),还能增强其对巨噬细胞的免疫调节活性。
附图说明
下面结合附图对本发明的具体实施方式作进一步详细说明。
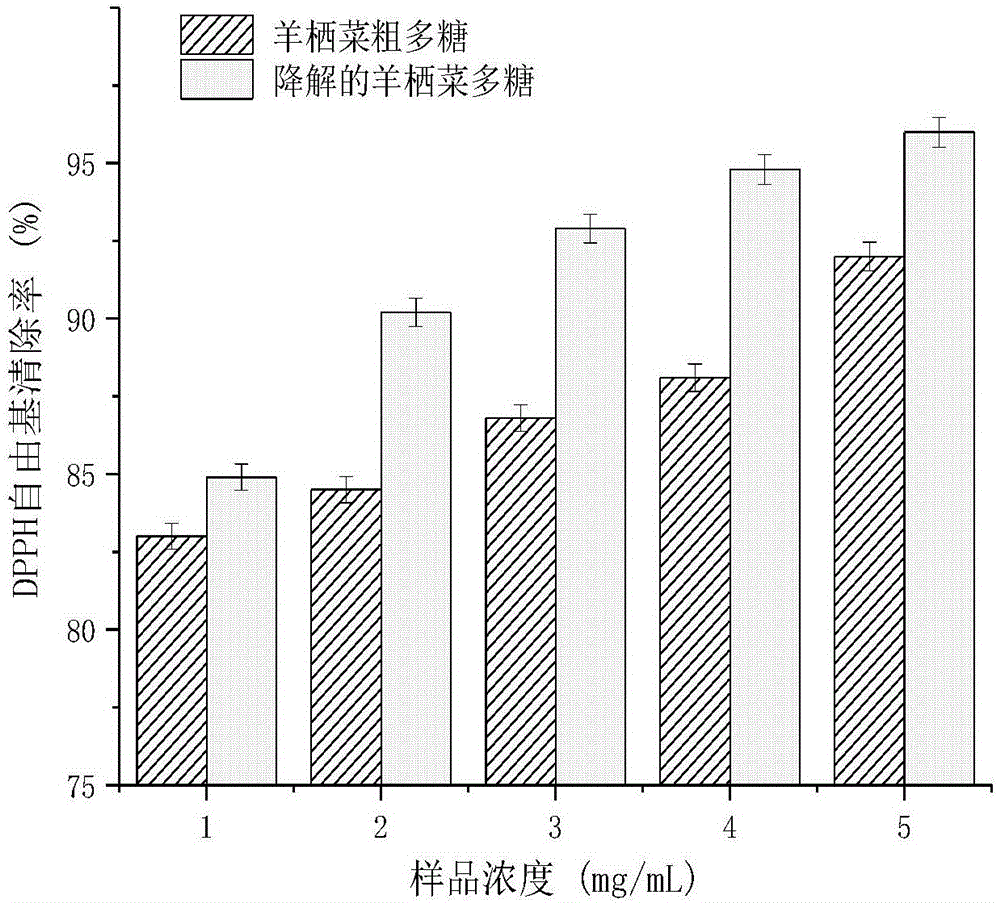
图1是由本发明实施例1-1得到的复合酶解羊栖菜多糖(esfp)与羊栖菜粗多糖(sfp)的dpph自由基清除能力的测定结果。
图2是由本发明实施例1-1得到的复合酶解羊栖菜多糖(esfp)与羊栖菜粗多糖(sfp)的超氧阴离子自由基清除能力的测定结果。
图3是由本发明实施例1-1得到的复合酶解羊栖菜多糖(esfp)与羊栖菜粗多糖(sfp)的羟基自由基清除能力的测定结果。
图4是由本发明实施例1-1得到的复合酶解羊栖菜多糖(esfp)与羊栖菜粗多糖(sfp)的还原力的测定结果。
图5是由本发明实施例2-1得到的复合酶解羊栖菜多糖(esfp)的各分离纯化组分esfp1、esfp2、esfp3、esfp4与羊栖菜粗多糖(sfp)对raw264.7巨噬细胞增殖的影响。
图6是由本发明实施例2-1得到的复合酶解羊栖菜多糖(esfp)的各分离纯化组分esfp1、esfp2、esfp3、esfp4与羊栖菜粗多糖(sfp)对raw264.7细胞吞噬活性的影响。
图7是由本发明实施例2-1得到的复合酶解羊栖菜多糖(esfp)的各分离纯化组分esfp1、esfp2、esfp3、esfp4与羊栖菜粗多糖(sfp)对raw264.7细胞释放no活性的影响。
具体实施方式
下面结合具体实施例对本发明进行进一步描述,但本发明的保护范围并不仅限于此:
实施例1-1:
称取1g羊栖菜粗多糖(sfp)溶解于100ml柠檬酸–柠檬酸钠缓冲液(ph6.0)中,加入果胶酶(3万u/g,437.4mg)和糖化酶(10万u/ml,39.8μl)(活力比为3.3:1),然后用相同乙酸-乙酸钠缓冲溶液稀释至250ml,使所得的体系中多糖终浓度为4mg/ml,酶终浓度为68.4u/ml,该体系在54.2℃水浴,振荡(频率为220r/min)反应3h。然后90℃以上水浴加热15分钟灭酶,过滤,将清液(滤液)旋蒸(60~70℃)浓缩至为原体积的1/4,用截留分子量为3500da的透析袋透析48h,将透析液(留在透析袋中的溶液)旋蒸浓缩(60~70℃浓缩至为透析液体积的25%)后冷冻(-50~-60℃)干燥24h,得到片状的羊栖菜酶解多糖(esfp1-1)921mg,此条件下得到的多糖的降解率为18.6%。
降解率=(降解多糖中的还原糖含量-未降解的粗多糖中的还原糖含量)/(总糖含量-未降解的粗多糖中的还原糖含量)×100%。
esfp1-1的组成及分子量见表1。
表1、降解多糖esfp1-1和粗多糖的组成和分子量比较
注:fuc,gal,man,glc,xyl和分别代表岩藻糖、半乳糖、甘露糖、葡糖糖和木糖。
实施例1-2:
将振荡反应时间改为4h,其余等同于实施例1-1,得到羊栖菜酶解多糖(esfp1-2)882mg,此条件下得到的多糖的降解率为18.7%。
实施例1-3:
将振荡反应时间改为1h,果胶酶和糖化酶活力比为3:1,其余等同于实施例1-1,得到羊栖菜酶解多糖(esfp1-3)913mg,此条件下得到的多糖的降解率为13.8%。
实施例1-4:
将振荡反应时间改为1h,反应温度为50℃,复合酶终浓度为70u/ml,果胶酶和糖化酶活力比为2:1,其余等同于实施例1-1,得到羊栖菜酶解多糖(esfp1-4)904mg,此条件下得到的多糖的降解率为9.4%。
实施例1-5:
将振荡反应时间改为1h,反应温度为60℃,复合酶终浓度为80u/ml,其余等同于实施例1-1相同,得到羊栖菜酶解多糖(esfp1-5)896mg,此条件下得到的多糖的降解率为8.9%。
对比例1、将实施例1-1中的果胶酶与糖化酶的活力比由3:1改成4:1;酶终浓度不变;其余等同于实施例1-1。得到羊栖菜酶解多糖(esfp-1d)878mg,此条件下得到的多糖的降解率为17.5%。
对比例2、将实施例1-1中的果胶酶与糖化酶的活力比由3:1改成2:1;酶终浓度不变;其余等同于实施例1-1。得到羊栖菜酶解多糖(esfp-2d)892mg,此条件下得到的多糖的降解率为15.9%。
实施例2-1、
将实施例1-1得到的羊栖菜复合酶解多糖(esfp1-1)400mg溶于7ml的去离子水中离心(6000r/min的转速下离心10分钟)取上清液,将上清液均匀的加入到层析柱填料上表面,经deaecellulose-52(柱子规格为60cm×φ2.6cm,有效长度为50cm)分别用去离子水、0.1mol/l、0.3mol/l、0.5mol/l、0.8mol/l的nacl溶液这5种洗脱剂进行洗脱,控制流速1ml/min;每管11min,用自动收集器收集;以硫酸-苯酚法隔管检测洗脱液在490nm处的紫外吸收值从而判定洗脱液中是否含有多糖,当检测到洗脱剂对应的洗脱液中不含有多糖时,则换下一种洗脱剂进行洗脱;分别收集0.1mol/l、0.3mol/l、0.5mol/l、0.8mol/l的nacl溶液这4种洗脱剂各自对应的洗脱液(为含糖洗脱液,体积分别为770、440、330、264ml)。对上述4种洗脱液分别进行如下处理:用旋转蒸发仪(60~70℃)浓缩至为原体积的25%、透析脱盐(用截留分子量为1000da的透析袋透析36h)、将透析液(留在透析袋中的溶液)冷冻干燥(-60℃干燥至恒重),从而对应得到羊栖菜酶解多糖的四个分离组分(四种初步纯化的多糖组分)。
备注:由于用去离子水洗脱得到的洗脱液(330ml)中多糖含量较低,没有进行进一步分离。
将上述四个分离组分再分别进行如下操作:用sephadexg-100层析柱(层析柱规格为60cm×φ1.6cm,有效长度为50cm)进行进一步纯化,用去离子水进行洗脱,纯化后分别得到esfp1、esfp2、esfp3和esfp4四个组分。
具体如下:
取100mg分离组分(初步纯化的多糖组分)溶于3ml去离子水中,离心(6000r/min的转速下离心12分钟)取上清液,加样于sephadexg-100凝胶色谱柱中,用200ml去离子水进行洗脱,流速为0.20ml/min,每20min收集一管。用硫酸-苯酚法逐管检测在490nm波长的吸收值,绘制sephadexg-100色谱柱洗脱曲线(用于确认收集的管数与样品的纯度,若为单峰则说明只有一个组分)。合并多糖洗脱液,旋蒸浓缩(浓缩至为原体积的25%)、冷冻干燥(-60℃干燥至恒重),从而分别得到羊栖菜复合酶解多糖的各纯化组分,命名为esfp1、esfp2、esfp3和esfp4。它们的分子量分别为75.5、86.3、76.2和86.5kda。其组成及分子量见表2。
表2、四个纯化多糖组分的组成和分子量
注:fuc,gal,man,glc,xyl和分别代表岩藻糖、半乳糖、甘露糖、葡糖糖和木糖。
实验1、
采用文献(卢茳虹.海带多糖的分离纯化、结构特性及在卷烟中应用研究[d].华南理工大学,2013.)报道方法,将实施例1-1得到的羊栖菜复合酶解多糖与羊栖菜粗多糖进行dpph自由基清除能力的测试,并进行比较。由图1可见,羊栖菜粗多糖经复合酶降解后,dpph自由基清除能力得到显著提高。sfp和esfp的ic50分别为0.768mg/ml和0.53mg/ml。esfp在浓度为5mg/ml时的清除能力相当于0.1mg/ml的vc的清除能力。但esfp的清除dpph自由基的活性仍小于维生素c。
实验2、
采用文献(marklunds,marklundg.involvementofthesuperoxideanionradicalintheautoxidationofpyrogallolandaconvenientassayforsuperoxidedismutase[j].europeanjournalofbiochemistery,1974,47:469-474.)报道方法,将实施例1-1得到的羊栖菜复合酶解多糖与羊栖菜粗多糖进行超氧阴离子自由基清除能力的测试,并进行比较。由图2可见,羊栖菜复合酶解多糖对超氧阴离子自由基的清除能力明显高于羊栖菜粗多糖。当浓度为5mg/ml时,sfp和esfp的清除率分别为73.9%、85.12%,sfp和esfp的ic50分别为0.723mg/ml和0.412mg/ml,说明酶解过程提高了羊栖菜多糖的超氧阴离子自由基清除能力。
实验3、
采用文献(yangwf,wangy,lixp,etal.purificationandstructuralcharacterizationofchineseyampolysaccharidesanditsactivities[j].carbohydratepolymers,2015,117:1021-1027.)报道方法,将实施例1-1得到的羊栖菜复合酶解多糖与羊栖菜粗多糖进行羟基自由基清除能力的测试,并进行比较。由图3可见,羊栖菜复合酶解多糖对羟基自由基的清除能力高于羊栖菜粗多糖。sfp、esfp在浓度为5mg/ml时,羟基自由基清除率分别为:39.15%和50.1%。其中esfp清除能力相当于0.02mg/ml的vc的清除能力。
实验4、
采用文献方法(yeng,chenh.antioxidantactivityofvariousteaextractsinrelationtotheirantimutagenicity[j].journalofagriculturalandfoodchemistry,1995,43:27-32.),将实施例1-1得到的羊栖菜复合酶解多糖与羊栖菜粗多糖进行还原力的测试,并进行比较。由图4可见,当两种多糖的浓度相同时,esfp的吸光度明显高于sfp,这表明相比较于酶解前,酶解后的多糖esfp对三价铁还原能力显著提高且在以vc为阳性对照的前提下可以发现其还原能力属多糖中较强。
实验5、
采用文献方法(niu,y.g.,xie,z.h.,zhang,h.,sheng,y.,&yu,l.l.(2013).effectsofstructuralmodificationsonphysicochemicalandbileacid-bindingpropertiesofpsyllium.journalofagriculturalandfoodchemistry,61(3),596e601.),将实施例1-1得到的羊栖菜复合酶解多糖与羊栖菜粗多糖进行体外胆酸结合能力的测定,并进行比较。
结果表明,羊栖菜复合酶解多糖(esfp)对游离胆酸钠、鹅去氧胆酸钠的结合能力较强,分别相当于等浓度的消胆胺的结合能力的53%、47%;而羊栖菜粗多糖(sfp)的胆酸结合能力则有所逊色,分别为消胆胺结合能力的50%、38%。
实施例1-2~实施例1-5制备所得的羊栖菜复合酶解多糖(esfp1-2)~(esfp1-5)、对比例1~对比例2所得的酶解多糖按照上述实验1~实验5所述方法进行检测,所得结果与实施例1-1的对比,如下表3所示。
表3、实施例1-1~1-5以及对比例1和2得到的降解多糖的抗氧化活性比较
实验6-1、调节巨噬细胞增殖作用测定
向96孔板的每孔中加入细胞悬液95μl(raw264.7巨噬细胞浓度为2×105个/ml),加入不同浓度的lps溶液和esfp1、esfp2、esfp3、esfp4溶液各5μl(lps的最终浓度为1、5、10μg/ml;esfp1、esfp2、esfp3、esfp4的终浓度为10、50、250、500、1000μg/ml),混合液在细胞培养箱中以培养温度为37℃,5%co2的培养条件下分别培养24小时、48小时。实验平行6组。培养24h后,从细胞培养箱中取出96孔板,向各孔加入10μl的mtt溶液(5mg/ml),小心混匀后继续在细胞培养箱中孵育4小时。小心吸去各孔中的培养液,防止细胞层破裂。向每孔中加入100μl的mtt溶液(5mg/ml)后轻轻振荡,以使甲臜结晶全部溶解,测定各孔在570nm处的od值。48h为同样实验步骤进行操作。
本实验以不同浓度的脂多糖(lps)作为阳性对照,水作为空白对照(即,浓度为0时)。
所得结果如图5所述,根据图5,可得知:培养24h后,在浓度较低时,各多糖组分对raw264.7巨噬细胞都有显著的增殖活性,但达到一定浓度后随浓度增大,细胞增殖活性反而下降。培养48h后,细胞增殖活性随浓度的变化趋势相似,但四个纯化多糖(esfp1-4)的细胞增殖活性明细高于粗多糖sfp和降解多糖esfp。
实验6-2、调节巨噬细胞吞噬作用测定
向96孔板的每孔中加入细胞悬液95μl(raw264.7巨噬细胞浓度为1×106个/ml),加入不同浓度的lps溶液和esfp1、esfp2、esfp3、esfp4溶液各5μl(lps的最终浓度为1、5、10μg/ml;esfp1、esfp2、esfp3、esfp4的终浓度为10、50、250、500、1000μg/ml),混合液在细胞培养箱中以培养温度为37℃,5%co2的培养条件下分别培养24小时、48小时。实验平行6组。培养24h后,从细胞培养箱中取出96孔板,将细胞培养液吸走,向各孔加入中性红溶液(0.075%)100μl,37℃,5%co2培养0.5h。弃去中性红溶液,用预温的pbs缓冲液洗去孔内残留的未结合中性红,每次100μl(用pbs清洗时要缓缓加入孔内防止将贴壁的细胞吹起)。将各孔pbs弃去,加入细胞裂解液150μl,37℃条件下反应1h。轻轻振荡混匀后,用酶标仪测定各孔在波长为570nm处的od值。48h为同样实验步骤进行操作。
本实验以不同浓度的lps作为阳性对照,水作为空白对照。
所得结果如图6所述,根据图6,可得知:培养24h后,6个样品中esfp1、esfp2和esfp的促进raw264.7细胞吞噬效果较好,其中esfp2在浓度为500μg/ml时有突出的效果,od值达到0.165,不仅远大于其他浓度的od值,还高于各个浓度的阳性对照。培养48h后的结果也表明,降解多糖esfp的巨噬细胞吞噬作用强于粗多糖sfp,巨噬细胞吞噬作用随多糖浓度的变化趋势与培养24h后的相似。
实验6-3、调节巨噬细胞释放no分子作用测定
向96孔板的每孔中加入细胞悬液95μl(为1×106浓度的raw264.7巨噬细胞的悬液),加入不同浓度(分别为1、5、10、50、250、500、1000μg/ml)的esfp1、esfp2、esfp3、esfp4溶液各5μl,在细胞培养箱中以培养温度为37℃,5%co2的培养条件下分别培养24小时、48小时。实验平行6组。培养24h后,从细胞培养箱中取出96孔板,将细胞培养液转移至另一新的96孔板,向各孔加入griess试剂(griessa:griessb=1:1)100μl,轻轻振荡混匀后,置于暗处静置10min,测定570nm处的od值。48h为同样实验步骤进行操作。
本实验以不同浓度的lps作为阳性对照,水作为空白对照。
所得结果如图7所述,根据图7,可得知:培养24h后,esfp的促进raw264.7细胞释放no因子的效果远好于粗多糖sfp。6个样品中esfp1、esfp2和esfp的促进raw264.7细胞释放no因子的效果较好。其中esfp2在浓度为1000μg/ml时有突出的效果,细胞液中no的浓度达到75.77μm,远远高于阳性对照lps的影响。培养48h后,对raw264.7细胞释放no的影响强弱依次为(按照峰值数值):esfp2>esfp1>esfp3>esfp4。esfp2的促进细胞分泌no的活性最强,在添加浓度为1000μg/ml时no浓度达到90.11μm。与培养24h的相比,培养48h后细胞液中的no的浓度显著增大。
最后,还需要注意的是,以上列举的仅是本发明的若干个具体实施例。显然,本发明不限于以上实施例,还可以有许多变形。本领域的普通技术人员能从本发明公开的内容直接导出或联想到的所有变形,均应认为是本发明的保护范围。。
- 还没有人留言评论。精彩留言会获得点赞!