一种高活性高产的小牛皱胃凝乳酶及基因与菌株和应用的制作方法
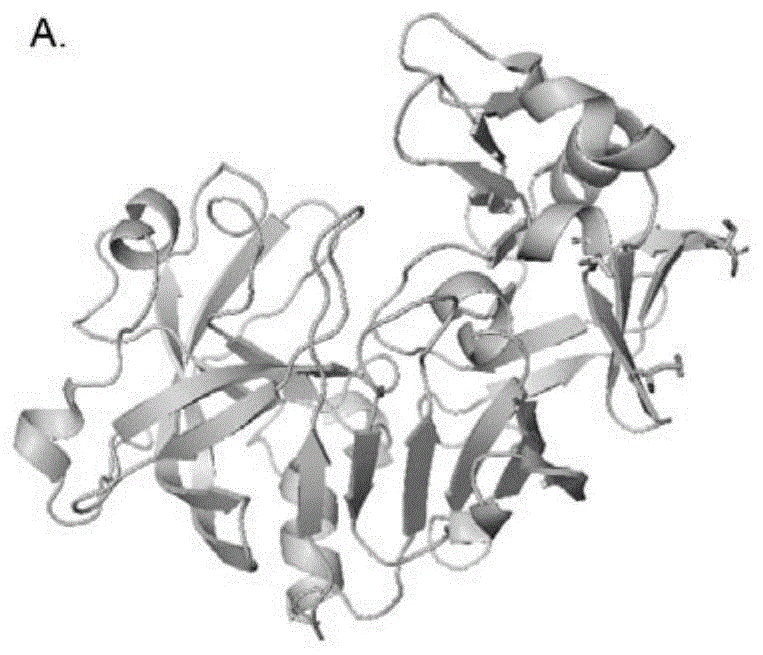
本发明属于基因工程技术领域,更具体地,涉及一种高活性高产的小牛皱胃凝乳酶及编码基因、重组表达载体、重组表达菌株,以及它们的制备方法和应用。
背景技术:
凝乳酶是可使乳液凝结的一类酶的总称。主要应用于乳酪的制作中。完全意义上的凝乳酶最早在小牛皱胃中发现。它是一种天冬氨酸蛋白酶,可专一地切割乳中κ-酪蛋白的phe105-met106之间的肽键,破坏酪蛋白胶束结构并形成稳定的副κ-酪蛋白及亲水性糖肽。当κ-酪蛋白被水解后,在钙离子存在下通过κ-酪蛋白分子胶束表面亚基之间以疏水键和胶体磷酸钙相互作用的方式连接在一起而形成乳液的凝固体。凝乳酶是乳酪生产必须的酶种,被广泛地应用于奶酪和酸奶的制作,在乳制品用酶中占首要地位,其产值约占整个酶制剂总产值的15.0%。
传统上,凝乳酶专指来源于未断奶小牛皱胃粘膜的一种蛋白酶。但由于乳制品行业对凝乳酶的需求量巨大,而资源又极为缺乏,因此,其它来源的且能使乳液凝结的蛋白酶均可称为凝乳酶。目前,凝乳酶的主要来源有:①动物源凝乳酶。凝乳酶最早来源于小牛的第四胃黏膜;②植物源凝乳酶。多种植物中均含有能使乳液凝固的蛋白酶。常见的有:木瓜蛋白酶、无花果蛋白酶、菠萝蛋白酶等;③微生物源凝乳酶。目前已发现有40余种微生物可生产一定活力的凝乳蛋白酶,主要来自真菌、放线菌、细菌等。
尽管不同来源的凝乳酶都具有使乳液凝固的作用,但在实际的乳酪生产应用过程,经典意义的小牛皱胃凝乳酶仍然是首选品种。它具有专一性强的特征,酪蛋白水解产物中没有呈苦味的氨基酸产生。其它来源的蛋白酶对酪蛋白的水解产物专一性不强,水解产生的苦味氨基酸含量高,影响乳酪的风味。因此,小牛胃凝乳酶仍是首选的凝乳酶品种。
采用基因工程手段,获得高活性、高产的凝乳酶生产菌株是获得可满足应用需求的、经济的小牛皱胃凝乳酶的主要方式。国际上,小牛皱胃凝乳酶基因已经在大肠杆菌(escherichiacoli)和乳酸克鲁维酵母菌(kluyveromyceslactis)中高效表达,并已经实现了产业化。但由于e.coli潜在的生物安全性,k.lactis等产量相对较低等原因,还需要从新的途径获得更高效、更高产的小牛皱胃凝乳酶及生产菌株。
毕赤酵母真核表达系统是目前广泛使用的真核生物表达系统,它具有生物安全性高、高密度发酵工艺成熟、生产技术配套等优点,是凝乳酶工业化生产的理想菌株。然而,由于受到毕赤酵母对异源基因密码子使用偏好性等因素的影响,来源于小牛皱胃的凝乳酶往往不会高效表达。因此,对小牛皱胃凝乳酶进行设计和改造,提高它的活性和表达量;通过异源基因的密码子依据毕赤酵母的偏好性进行优化等方式进一步提高其产量,获取高活性、高产菌株具有非常重要的意义。
技术实现要素:
本发明的主要目的是提出一种高活性、高产的小牛皱胃凝乳酶及基因、重组表达载体、高产菌株,以及它们的制备方法,旨在为小牛皱胃凝乳酶的产业化及应用奠定基础。
为了实现上述目的,本发明提供一种高活性高产的小牛皱胃凝乳酶bochy,所述凝乳酶bochy的氨基酸序列如seqidno:1所示。
根据本发明,所述小牛皱胃凝乳酶bochy的原始凝乳酶chy为来自小牛的皱胃(bostaurus)的凝乳酶(chymosin)(genbank登录号:nm_180994)。所述原始凝乳酶chy的氨基酸序列如seqidno:3所示,原始凝乳酶基因chy的核苷酸序列如seqidno:4所示。为提高其活性、蛋白质结构的稳定性和表达量。通过蛋白质理性设计的方式对其中的三个氨基酸进行了突变。获得具有新的氨基酸序列的小牛皱胃凝乳酶。所述小牛皱胃凝乳酶bochy的氨基酸序列如seqidno:1所示。具体的设计包括:通过定点突变的方法将原始凝乳酶chy序列中第201位的丝氨酸突变为天冬酰胺(s201n),第267位的甲硫氨酸突变为赖氨酸(m267k),第271位的苏氨酸突变为脯氨酸(t271p)。所述定点突变技术为本领域常规的方法,其具体操作步骤为本领域技术人员公知,在此不再赘述。
本发明提供了一种高活性的小牛皱胃凝乳酶基因bochy,用于编码小牛皱胃凝乳酶bochy。具体地,所述小牛皱胃凝乳酶基因bochy的核苷酸序列与seqidno:2所示的碱基序列具有95%以上的同一性。优选地,根据本发明,所述的小牛皱胃凝乳酶基因bochy含有seqidno:2所示的碱基序列即可实现本发明的效果。在最优情况下,本发明所述的小牛皱胃凝乳酶bochy的碱基序列为seqidno:2所示的碱基序列。
本发明提供的小牛皱胃凝乳酶基因bochy,系基于bochy氨基酸序列人工设计合成而来。在设计小牛皱胃凝乳酶基因bochy时,以bochy的氨基酸序列为模板,选择简并密码子中在毕赤酵母中高频使用的密码子来翻译所对应的氨基酸,显著提高了人工设计的小牛皱胃凝乳酶基因bochy的密码子使用频率,有助于提高bochy在毕赤酵母细胞中的表达量。所设计的bochy通过人工合成的方法获得,其碱基序列如seqidno:2所示,命名为bochy。
本发明提供了一种重组表达载体,其包括上述小牛皱胃凝乳酶基因bochy,还可包括其他功能单元。在小牛皱胃凝乳酶bochy的氨基酸序列和小牛皱胃凝乳酶基因bochy的碱基序列确定的情况下,本领域技术人员能够选择合适的重组表达载体以及其他功能单元,例如,毕赤酵母表达载体。
根据本发明一种具体实施方式,所述重组表达载体的制备方法包括以下步骤:将小牛皱胃凝乳酶基因bochy分别用限制性内切酶ecori和noti双酶切,得到具有粘性末端的凝乳酶基因bochy片段;将毕赤酵母表达载体ppiczαa分别用限制性内切酶ecori和noti双酶切,得到具有粘性末端的载体ppiczαa片段;将所述具有粘性末端的凝乳酶基因bochy片段和所述具有粘性末端的载体ppiczαa片段通过t4dna连接酶连接,获得凝乳酶重组表达载体ppic-bochy。除ppic-bochy外,在本发明其他实施例中,也可选用其他毕赤酵母表达载体。
本发明提供一种重组表达菌株,该菌株含有bochy基因,其表达产物为上述小牛皱胃凝乳酶bochy。通常来说,所述宿主细胞可以是大肠杆菌、芽孢杆菌、曲霉,也可以是酵母等细胞,或者是其他类型的细胞,例如动物细胞等。优选地,所述重组表达菌株的宿主细胞为毕赤酵母。
根据本发明一种具体实施方式,所述重组表达菌株的制备方法包括如下步骤:将ppic-bochy重组表达质粒通过限制性内切酶(如bamhi限制性内切酶)线性化;将线性化的ppic-bochy导入毕赤酵母宿主细胞中,获得重组表达菌株。可选地,通过电转化将重组表达载体导入毕赤酵母宿主细胞中,获得重组表达菌株。在本发明的一些实施例中,所述毕赤酵母为巴斯德毕赤酵母x-33,当然,在本发明其他实施例中,也可选用其他毕赤酵母菌株。
本发明对所述重组表达菌株的制备方法没有特别限定,可根据本领域常规技术手段确定。在小牛皱胃凝乳酶bochy的氨基酸序列确定的情况下,本领域技术人员能够获得合适的重组表达菌株。
本发明中,重组表达菌株制备小牛皱胃凝乳酶的方法可以采用本领域常规的方法。根据本发明一种具体实施方式,生产小牛皱胃凝乳酶的方法包括:挑取重组表达菌株至培养基中培养约24小时,并收获菌体;将菌体转入新鲜的培养基中,每24小时加入约1%的甲醇,诱导小牛皱胃凝乳酶bochy的表达。诱导表达约96小时后,离心收集上清液,获得含bochy的溶液。在一些实施方式中,也可以通过其它方式进行培养,并将发酵上清液视为凝乳酶产物。
本发明提供一种采用小牛皱胃凝乳酶bochy凝聚乳和乳制品的方法,该方法包括:将上述小牛皱胃凝乳酶bochy与含乳物料接触。所述含乳物料包括牛乳和各种类型的乳制品,也包括含牛乳类成分的其它物质。
本发明提供的凝乳酶bochy通过人工定向改造,显著提高了其活性和表达水平。本发明提供的凝乳酶基因bochy以凝乳酶bochy的氨基酸序列为参照,以酵母密码子使用频率为标准,采用氨基酸的简并密码子中的高频密码子来翻译对应的氨基酸,从而替换原有的低频密码子,进一步提高了人工定向改造后的凝乳酶bochy在酵母细胞中的表达量。凝乳酶在发酵罐中的表达量达到75,600su/ml发酵液,其制备工艺简单且产量高,极大地降低了凝乳酶的生产成本。
本发明的其它特征和优点将在随后具体实施方式部分予以详细说明。
附图说明
通过结合附图对本发明示例性实施方式进行更详细的描述,本发明的上述以及其它目的、特征和优势将变得更加明显。
图1a和图1b示出了本发明实施例1中凝乳酶chy用于突变的三个氨基酸第201位的丝氨酸,第267位的甲硫氨酸和第271位的苏氨酸在三维结构中所在的位置。
图2a和2b为本发明实施例2中原始凝乳酶基因chy和分子改造的凝乳酶基因bochy的密码子使用频率的对比图。
图3a和3b为本发明实施例3中将原始凝乳酶基因chy和重新设计的基因bochy克隆入重组表达载体ppiczαa的酶切检验结果。
图4a和4b为本发明实施例4中原始凝乳酶chy和重新设计的凝聚乳酶bochy在摇瓶条件下表达的结果。
图5和图6分别为本发明实施例5中重新设计的凝乳酶bochy在50升发酵罐中的产凝乳酶的酶活曲线和产酶图。
图7为本发明实施例6中凝乳酶bochy对牛乳的凝乳实验图。
具体实施方式
下面将更详细地描述本发明的优选实施方式。虽然以下描述了本发明的优选实施方式,然而应该理解,可以以各种形式实现本发明而不应被这里阐述的实施方式所限制。实施例中未注明具体条件者,按照常规条件或制造商建议的条件进行。所用试剂或仪器未注明生产厂商者,均为可以通过市售购买获得的常规产品。
实施例1
本实施例用于说明将原始凝乳酶chy中的三个氨基酸进行定点突变的过程。具体实施实施过程如下:
根据原始小牛皱胃凝乳酶chy(genbank登录号:nm_180994)的三维结构,通过蛋白质设计的方式对其中三个氨基酸进行突变。具体的设计包括:通过定点突变的方法将原始凝乳酶chy序列中第201位的丝氨酸突变为天冬酰胺(s201n),第267位的甲硫氨酸突变为赖氨酸(m267k),第271位的苏氨酸突变为脯氨酸(t271p)。突变后的凝乳酶bochy的氨基酸序列如seqidno:1所示,原始凝乳酶chy的氨基酸序列如seqidno:3所示。
图1a和图1b列出了凝乳酶chy中用于改造的氨基酸,即第201位的丝氨酸(s201n),第267位的甲硫氨酸(m267k),第271位的苏氨酸(t271p)在酶分子中的具体位置。
实施例2
本实施例用于说明通过密码子的优化获得凝乳酶基因bochy。
本实施例基于重新设计引入新的小牛皱胃凝乳酶bochy的氨基酸序列,人工重新设计了凝乳酶基因的核苷酸序列。具体包括:以凝乳酶bochy的氨基酸序列为基础,在每一个氨基酸对应的简并密码子中,采用在毕赤酵母中高频率使用的密码子替代低频率的密码子;同时在mfold软件的辅助下,降低新设计的基因bochy编码的mrna的二级结构的复杂度和最小自由能,提高凝乳酶在翻译过程中的速度和翻译效率,提高凝乳酶的表达量。完成凝乳酶基因bochy序列的人工设计后,通过化学法人工合成了凝乳酶基因bochy。所述人工合成基因的技术为本领域常规的方法,其具体操作步骤为本领域技术人员公知,在此不再赘述。
本实施例根据凝乳酶的氨基酸序列,经人工设计并优化后,获得了新型凝乳酶基因,其核苷酸序列如seqidno:2所示,命名为bochy,所述凝乳酶基因bochy编码的凝乳酶的氨基酸序列如seqidno:1所示,所述原始凝乳酶基因chy的核苷酶序列如seqidno:4如示,所编码的凝乳酶chy的氨基酸序列如seqidno:3所示。图2a为原始凝乳酶chy100个氨基酸密码子的使用频率,其使用频率普遍偏低。图2b为优化后凝乳酶bochy中前100个氨基酸密码子的使用频率,其使用频率大幅度提高。由此可见,本发明所获得的凝乳酶基因bochy的氨基酸密码子的使用频率有明显的优化。
实施例3
本实施例用于说明凝乳酶基因bochy重组表达菌株的构建。
在小牛皱胃凝乳酶基因bochy和原始凝乳酶基因chy两端分别加上酶切位点ecori和noti酶切位点,经ecori和noti酶切;同样载体ppiczαa经ecori和noti酶切。将经ecori和noti酶切的小牛皱胃凝乳酶基因bochy和原始凝乳酶基因chy分别与ppiczαa通过t4dna连接酶连接,获得ppic-bochy和ppic-chy重组表达载体。连接体系为:总体积为10μl,其中含有100ng的bochy(或chy)、50ng的ppiczαa和10u的t4dna连接酶;在16℃条件下连接10小时,获得重组表达载体ppic-bochy(或ppic-chy)。图3a所示为重组表达载体ppic-chy的检测结果(图中:m为dl5000dnamarker,1泳道为载体ppic-chy,2泳道为双酶切后的ppic-chy重组表达载体;图3b所示为重组表达载体ppic-bochy的检测结果(图中:m为dl5000dnamarker,1泳道为载体ppic-bochy,2泳道为双酶切后的ppic-bochy重组表达载体)。由图3a和图3b可知,优化后的凝乳酶基因bochy和原始凝乳酶基因chy成功连接至ppiczαa重组表达载体上。将重组表达载体ppic-chy和ppic-bochy转化入巴斯德毕赤酵母中,分别得到含凝乳酶基因bochy和chy的重组表达菌株。
实施例4
本实施例用于说明凝乳酶bochy在摇瓶中的发酵生产。
(1)将实施例3中含bochy基因(或者原始凝乳酶chy基因)的毕赤基因工程菌接种到5mlypd培养基中,过夜培养,获得种子液;
(2)将上述种子液接种至含25mlbmgy发酵培养基的摇瓶中进行发酵培养。bmgy发酵培养基的配方为:1%酵母粉、2%蛋白胨、100mm磷酸缓冲液(ph6.0)、1.34%酵母氮碱、0.004%生物素、1%甘油。培养过夜后,收集菌体并转移到25mlbmmy培养基中继续培养。bmmy培养基的配方为:1%酵母粉、2%蛋白胨、100mm磷酸缓冲液(ph6.0)、1.34%酵母氮碱、0.004%生物素、0.5%甲醇。每24h补充0.25ml甲醇,至96h时停止发酵。发酵液采用sds-page进行检测。图4a和4b所示为发酵液的检测结果(图4a中:m为蛋白质标准,1-8泳道为未优化凝乳酶基因chy表达的结果;图4b中:m为蛋白质标准,1-8泳道为优化后凝乳酶基因bochy表达的结果)。图4a和4b说明优化后的凝乳酶基因bochy的表达水平明显高于原始凝乳酶基因chy的表达水平。
实施例5
本实施例用于说明凝乳酶bochy在发酵罐中的发酵生产和活性测定。
(1)将实施例3中的凝乳酶bochy毕赤基因工程菌接种到500mlypd培养基中,培养24h,获得种子液;
(2)将上述种子液接种至含30-l无机盐发酵培养基的发酵罐中进行发酵培养,其中,发酵培养基包括:1050gkh2po4、25gcaso4、120g(nh4)2so4、150gmgso4、3600gk2so4和1700g甘油。
(3)在生长的前期阶段,培养温度为30℃,ph值为5.0,转速200rpm,通气量2.5l/min。本阶段持续约30h。生长阶段结束后即进入甲醇诱导表达阶段。在此阶段,溶氧维持在5-25%,温度28℃,ph值为5,甲醇流速为2.0-3.0ml/l/h,流加时间为120h。每隔24h取样,并测定发酵液的凝乳酶活性。
凝乳酶活性的测定采用arimak法。取5ml浓度为10%的脱脂乳,在35℃温度下保温10min,加入0.5ml适量稀释后的凝乳酶溶液,快速混合均匀,并在35℃温度下继续保温,倾斜试管,沿试管轴方向旋转并观察,以试管壁开始出现絮状凝集小颗粒为终点,准确记录样品加入到乳凝固时间t(s)。凝乳酶活性单位定义为:在40min内凝固1ml浓度为10%的脱脂乳的所需的酶量定义为一个索氏单位(su)。酶活力(su)=(供试乳量ml/凝乳酶量ml)*d*2400/t。式中:d为酶液稀释倍数;t为凝乳时间(s)。发酵液中蛋白质组份的检测采用sds-page法检测,检测结果如图5所示。随着发酵时间的延长,发酵液中bochy的含量逐渐增加。由图6可知,凝乳酶bochy生产水平随着培养时间的延长,发酵液中凝乳酶的活力逐渐增高,在培养时间为140h时,发酵液中凝乳酶的活力最高为75,600su/ml发酵液。
实施例6
本实施例用于说明采用凝乳酶aogal水解乳糖。
(1)凝乳酶对牛乳的凝聚实验采用如实施例5所述方法制得的凝乳酶进行。bochy凝乳过程为:取1ml浓度为10%的牛乳液体,在35℃温度下保温10min,加入0.1ml适量稀释后的凝乳酶bochy溶液,混合均匀并在35℃温度下继续保温,观察牛乳凝聚的情况。图7所示为不同稀释度的凝乳酶bochy对牛乳的凝聚状况。图7表明凝乳酶bochy可高效地对牛乳进行凝聚。
综上所述,本发明通过酶分子设计,向原始小牛皱胃凝乳酶中引入三个氨基酸突变,显著提高了其结构稳定性和表达水平,并根据毕赤酵母对同一氨基酸的简并密码子的偏好性对凝乳酶基因的核苷酸序列进行优化设计,合成出高效表达的小牛皱胃凝乳酶基因bochy。该基因与表达载体ppiczαa连接后,转入毕赤酵母中表达,获得了高效表达的、高产的小牛皱胃凝乳酶生产菌株;其在50l的发酵罐中培养表达140小时后,上清液中凝乳酶的酶活可达75,600su/ml。本发明获得的凝乳酶对牛乳具有高效的凝聚能力。
以上已经描述了本发明的各实施例,上述说明是示例性的,并非穷尽性的,并且也不限于所披露的各实施例。在不偏离所说明的各实施例的范围和精神的情况下,对于本技术领域的普通技术人员来说许多修改和变更都是显而易见的。
序列表
<110>武汉轻工大学
<120>一种高活性高产的小牛皱胃凝乳酶及基因与菌株和应用
<130>bji1901917whpu
<160>4
<170>siposequencelisting1.0
<210>1
<211>323
<212>prt
<213>人工序列(artificialsequence)
<400>1
glygluvalalaservalproleuthrasntyrleuaspserglntyr
151015
pheglylysiletyrleuglythrproproglngluphethrvalleu
202530
pheaspthrglyserseraspphetrpvalproseriletyrcyslys
354045
serasnalacyslysasnhisglnargpheaspproarglysserser
505560
thrpheglnasnleuglylysproleuserilehistyrglythrarg
65707580
sermetglnglyileleuglytyraspthrvalthrvalserasnile
859095
valaspileglnglnthrvalglyleuserthrglngluproglyasp
100105110
valphethrtyralaglupheaspglyileleuargmetalatyrpro
115120125
serleualaserglutyrserileprovalpheaspasnmetmetasn
130135140
arghisleuvalalaglnaspleupheservaltyrmetaspargasn
145150155160
glyglngluthrmetleuthrleuglyalaileaspprosertyrtyr
165170175
thrglyserleuhistrpvalprovalthrvalglnglntyrtrpgln
180185190
phethrvalaspservalthrileasnglyvalvalvalalacysglu
195200205
glyglycysglnalaileleuaspthrglythrserlysleuvalgly
210215220
proserseraspileleuasnileglnglnalaileglyalathrgln
225230235240
asnglntyrglyglupheaspileaspcysaspasnleusertyrmet
245250255
prothrvalvalphegluileasnglylyslystyrproleupropro
260265270
seralatyrthrserglnaspglnglyphecysthrseralaphegln
275280285
sergluasnhisserglnlystrpileleuglyaspvalpheilearg
290295300
glutyrtyrservalpheaspargalaasnasnleuvalglyleuala
305310315320
lysalaile
<210>2
<211>972
<212>dna
<213>人工序列(artificialsequence)
<400>2
ggtgaagttgcttctgttccattgactaactacttggattctcaatactttggtaagatt60
tacttgggtactccaccacaagaatttactgttttgtttgatactggttcttctgatttt120
tgggttccatctatttactgtaagtctaacgcttgtaagaaccatcaaagatttgatcca180
agaaagtcttctacttttcaaaacttgggtaagccattgtctattcattacggtactaga240
tctatgcaaggtattttgggttacgatactgttactgtttctaacattgttgatattcaa300
caaactgttggtttgtctactcaagaaccaggtgatgtttttacttacgctgaatttgat360
ggtattttgagaatggcttacccatctttggcttctgaatactctattccagtttttgat420
aacatgatgaacagacatttggttgctcaagatttgttttctgtttacatggatagaaac480
ggtcaagaaactatgttgactttgggtgctattgatccatcttactacactggttctttg540
cattgggttccagttactgttcaacaatactggcaatttactgttgattctgttactatt600
aacggtgttgttgttgcttgtgaaggtggttgtcaagctattttggatactggtacttct660
aagttggttggtccatcttctgatattttgaacattcaacaagctattggtgctactcaa720
aaccaatacggtgaatttgatattgattgtgataacttgtcttacatgccaactgttgtt780
tttgaaattaacggtaagaagtacccattgccaccatctgcttacacttctcaagatcaa840
ggtttttgtacttctgcttttcaatctgaaaaccattctcaaaagtggattttgggtgat900
gtttttattagagaatactactctgtttttgatagagctaacaacttggttggtttggct960
aaggctatttaa972
<210>3
<211>323
<212>prt
<213>bostaurus
<400>3
glygluvalalaservalproleuthrasntyrleuaspserglntyr
151015
pheglylysiletyrleuglythrproproglngluphethrvalleu
202530
pheaspthrglyserseraspphetrpvalproseriletyrcyslys
354045
serasnalacyslysasnhisglnargpheaspproarglysserser
505560
thrpheglnasnleuglylysproleuserilehistyrglythrarg
65707580
sermetglnglyileleuglytyraspthrvalthrvalserasnile
859095
valaspileglnglnthrvalglyleuserthrglngluproglyasp
100105110
valphethrtyralaglupheaspglyileleuargmetalatyrpro
115120125
serleualaserglutyrserileprovalpheaspasnmetmetasn
130135140
arghisleuvalalaglnaspleupheservaltyrmetaspargasn
145150155160
glyglngluthrmetleuthrleuglyalaileaspprosertyrtyr
165170175
thrglyserleuhistrpvalprovalthrvalglnglntyrtrpgln
180185190
phethrvalaspservalthrileserglyvalvalvalalacysglu
195200205
glyglycysglnalaileleuaspthrglythrserlysleuvalgly
210215220
proserseraspileleuasnileglnglnalaileglyalathrgln
225230235240
asnglntyrglyglupheaspileaspcysaspasnleusertyrmet
245250255
prothrvalvalphegluileasnglylysmettyrproleuthrpro
260265270
seralatyrthrserglnaspglnglyphecysthrseralaphegln
275280285
sergluasnhisserglnlystrpileleuglyaspvalpheilearg
290295300
glutyrtyrservalpheaspargalaasnasnleuvalglyleuala
305310315320
lysalaile
<210>4
<211>972
<212>dna
<213>bostaurus
<400>4
ggggaggtggccagcgtgcccctgaccaactacctggatagtcagtactttgggaagatc60
tacttggggaccccgccacaggagttcaccgtgctgtttgacactggctcctctgacttc120
tgggtaccctctatctactgcaagagcaatgcctgcaaaaaccaccagcgcttcgacccg180
agaaagtcgtccaccttccagaacctgggcaagcccctgtctatccactacgggacacgc240
agcatgcagggcatcctgggctatgacaccgtcactgtctccaacattgtggacatccag300
cagacagtaggcctgagcacccaggagccaggtgacgtcttcacctatgccgagttcgac360
gggatcctgcggatggcctacccctcgctggcctcagagtactcgatacccgtgtttgac420
aacatgatgaaccgtcacctggtggcccaagacctgttctcggtttacatggaccgtaat480
ggccaggagaccatgctgacgctgggtgccatcgacccgtcctactacacagggtccctg540
cattgggtgcccgtgacagtgcagcagtactggcagttcactgtggacagtgtcaccatc600
agcggtgtggttgtggcctgtgagggtggctgtcaggccatcctggacacgggcacctcc660
aagctggtcgggcccagcagcgacatcttgaacatccagcaggccattggagccacacag720
aaccagtacggtgagtttgacatcgactgcgacaacctgagctacatgcccactgtggtc780
tttgagatcaatggcaaaatgtacccactgaccccctccgcctataccagccaggaccag840
ggcttctgtaccagtgccttccagagtgaaaatcattcccagaaatggatcctgggggat900
gttttcatccgcgagtattacagcgtctttgaccgtgccaacaacttggtggggctggcc960
aaagccatctaa972
- 还没有人留言评论。精彩留言会获得点赞!