杜仲漆酶EuLAC1基因及其用途
杜仲漆酶eulac1基因及其用途
技术领域
1.本发明属于植物生物技术领域,具体涉及一种新基因(eulac1)及其在抗病方面的应用。
背景技术:
2.在农业生产中,植物病害尤其是真菌病害一直是限制农作物产量提高的重要因素。目前主要采用化学农药作为防治病害的主要手段,但长期使用化学农药不仅导致病原菌产生抗药性,同时还造成生态环境的破坏。因此,抗病品种选育越来越受关注,成为植物病害的有效防治措施之一。然而,由于常规育种周期长、育种资源有限、病原菌变异迅速,因此不易获得抗性植株,难以满足生产的需要。随着生物工程技术发展,研究者开始利用转基因技术,将外源抗病基因转入植物以提高植株的抗病性;这种方法具有选育速度快,易于获得抗性植株,且具有较好的遗传稳定性等优点,已成为获得抗病性植株的主要手段。
3.杜仲(eucommia ulmoides oliv.)属于单科单属落叶乔木,别名为胶木,具有广泛的胶用和药用价值,是我国的一种特有经济树种。漆酶(laccase,lac)是属于铜蓝氧化酶蛋白家族的一种含四个铜离子的糖蛋白氧化酶,在高等植物中主要影响木质素的合成、植物体内酚类物质含量的调控以及参与到植物对病虫害的抵御等。目前,漆酶在许多高等植物中均有报道,如拟南芥(arabidopsis thaliana)、棉花(gossypium spp)、毛果杨(populus trichocarpa)、烟草(nicotiana tabacum)以及玉米(zea mays)等物种。植物漆酶家族成员众多,且不同成员的功能不尽相同,但多数的功能研究还是集中在木质素的合成上。但有关杜仲lac基因的研究未见报道,其功能机制存在空白。
技术实现要素:
4.本发明的目的是提供一种杜仲漆酶eulac1基因,为植物的基因改造提供了一个新的高效抗病基因,为植物抗病基因工程提供了更多的选择,同时也为通过分子手段改良植物抵抗真菌病害能力奠定了基础。
5.为实现上述目的,本发明采用如下方案:
6.杜仲漆酶eulac1蛋白,其氨基酸序列如seq id no.1所示。
7.编码上述蛋白的杜仲漆酶eulac1基因。
8.优选其核苷酸序列如seq id no.2所示。
9.一种重组载体,包括上述eulac1基因的核苷酸序列。
10.一种遗传工程化的宿主细胞,含有上述重组载体。
11.上述杜仲漆酶eulac1蛋白在植物抵抗疾病中的应用。
12.进一步,所述疾病为由灰葡萄孢(botrytis cinerea,b.cinera)引发的灰霉病。
13.一种提高植物对灰霉病抗性的方法,包括以下步骤:将制备的或提供的含有所述的eulac1基因的表达载体转化农杆菌,得到农杆菌工程菌,再将所述农杆菌工程菌侵染植物,使eulac1基因过表达,即得到转基因植物,以增强抗灰霉病的性能。
14.进一步,所述转基因植物漆酶活性增加、木质素含量增加和木质部附近的细胞壁增厚且轮廓清晰。
15.本发明从杜仲中克隆并获得了杜仲漆酶基因eulac1及其序列,对其进行生物信息学分析。通过构建植物过表达载体,遗传转化烟草,从而验证分析eulac1基因抗病方面的功能。
16.相比现有技术,本发明具有如下有益效果:
17.本发明首次克隆了杜仲漆酶eulac1基因,获得其编码的蛋白序列,并将该基因在烟草中表达后,用离体叶片法进行接菌实验,实验结果表明该基因能够明显提高烟草对灰霉病的抗性,为植物抗病基因工程提供了一个高效的抗病基因,并为进一步培育具有真菌抗性的转基因新材料或新品种奠定了基础。利用该方法可以减少农药使用,降低环境污染。本发明为今后利用该基因进行植物分子遗传改良提供理论和技术支持,因此,具有重要的推广价值。
附图说明
18.图1为杜仲漆酶eulac1基因的pcr扩增电泳图;
19.图2为植物过表达载体pcambia1300-35s-eulac1的图谱示意图;
20.图3为烟草遗传转化过程图,a.共培养,b.筛选培养,c.愈伤组织形成,d.不定芽分化,e.抗性苗生根,f.抗性植株移栽;
21.图4为转基因烟草基因组dna的pcr检测结果电泳图,m:2000maker,1:阳性质粒对照,2-7:抗性植株,8:野生型烟草,9:转pcambia300-35s-gus空载烟草;
22.图5为eulac1在杜仲中的相对表达量,“**”表示差异极显著(p《0.01);
23.图6为阳性转基因烟草中eulac1基因的表达量,转pcambia300-35s-gus空载烟草,
24.tp1-tp20为转基因烟草阳性株系,“**”表示差异极显著(p《0.01),“*”表示差异显著(p《0.05);
25.图7为转基因烟草的漆酶活性,wt:野生型植株,转pcambia300-35s-gus空载烟草,tp:转基因植株,“**”表示具有极显著差异p《0.01;
26.图8为转基因烟草灰霉病抗性实验,a为叶片拍照图,(a)接菌前,(b)接菌3d后,wt:野生型植株,b为病斑直径测量结果图,wt:野生型植株,tp:转基因植株,“**”表示具有极显著差异p《0.01;
27.图9为转基因烟草的木质素含量,wt:野生型植株,转pcambia300-35s-gus空载烟草,tp:转基因植株;a为转基因烟草茎中木质素含量,b为转基因烟草叶中木质素含量,根据最小显著性差异检验,具有相同字母的平均值不具有统计学意义(p《0.01);
28.图10为烟草茎的横截面电镜观察图,wt:野生型植株,tp:转基因植株,放大倍数1000倍;图中箭头所指为:左图为右图圆圈内的放大图。
具体实施方式
29.下面通过具体实施方式结合附图对本发明作进一步详细说明。
30.下述所用生物材料均为市售,本技术人实验室有保存,可以对外公开发放。
31.实施例1杜仲漆酶eulac1基因克隆及分析
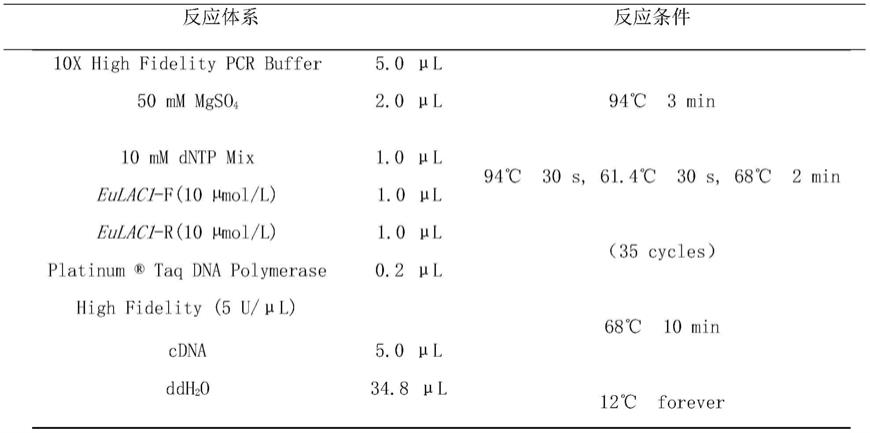
32.采用康为世纪公司的rna提取试剂盒rna pure kit,按说明书步骤提取杜仲rna,用分光光度计测定rna的浓度,选取数值在2.0左右的rna,按照takara公司的反转录试剂盒说明书合成cdna,然后进行pcr扩增目的片段。pcr体系如表1所示。
33.表1 pcr扩增体系
[0034][0035]
其中,正向引物和反向引物为:
[0036]
eulac1-f:5
’-
atgggttcttgtattcttaaggcat-3’,
[0037]
eulac1-r:5
’-
ctaacctggattgtctgccc-3’。
[0038]
获得pcr产物通过琼脂糖凝胶电泳分析,如图1所示,可以观察到一条特异性的扩增条带。按照omega公司胶回收试剂盒纯化目的条带,按照表2连接体系将其与ptopo-blunt载体相连,连接产物转化大肠杆菌dh5α菌株,用碱裂解法提取经过amp筛选的阳性菌落的质粒,经pcr实验验证后,送交华大基因测序,经过测序拼接获得全长为1617bp的基因序列,如seq id no.2所示。
[0039]
表2连接体系
[0040][0041]
根据seq id no.2,通过dnaman软件翻译,获得杜仲漆酶eulac1的蛋白质序列如seq id no.1所示,含有538个氨基酸。通过expasy在线分析表明,eulac1基因分子量为59.4kd,理论等电点为9.83。磷酸化位点预测分析发现eulac1蛋白具有15个丝氨酸磷酸化位点、19个苏氨酸磷酸化位点和7个酪氨酸磷酸化位点,对eulac1基因编码的蛋白结构域进行分析,发现该序列中含有三个铜氧还蛋白结构域,属于铜蓝氧化酶蛋白家族的一员,符合
漆酶的基本特征。
[0042]
实施例2表达载体构建及农杆菌介导的遗传转化
[0043]
采用冻融法将上述重组质粒转入农杆菌lba4404感受态细胞中,使用质粒提取试剂盒提取阳性农杆菌菌落的质粒并进行kpni、xbai双酶切验证。用琼脂糖凝胶进行检测,电泳结果显示得到预期大小为1617bp的目的条带,表明农杆菌中已含有重组质粒。eulac1基因过表达载体pcambia1300-35s-eulac1,经本项目组设计后由转导精进(武汉)生物技术有限公司构建(图2)。
[0044]
通过农杆菌介导的叶盘法转入三星烟草中,其步骤如下:准备100ml验证成功的携带pcambia1300-35s-eulac1质粒的农杆菌菌株,培养至浓度为od600=0.6-0.8,室温下4500rpm离心5min,然后倒掉上层培养液,用100ml重悬液重悬菌体;采取较嫩的三星烟草叶片,洗净后用自来水水流冲洗20-30min;在超净工作台中用75%的医用酒精浸泡叶片30-60s;再用0.01%的升汞浸泡摇晃8min,然后再用无菌水清洗3次;用吸水纸把叶片表面的水分吸干,于事先准备好的无菌皿中用无菌刀片切去叶片的叶柄和叶边缘,切成1cm2左右的方形小块;将切好的叶片放入重悬好的菌液中,轻轻摇晃,侵染8min左右,取准备好的无菌滤纸,小心吸干叶片表面的重悬菌液,注意不要触碰到台面;将叶片倒置并紧密铺于烟草共培培养基(ms粉4.43g/l,蔗糖30g/l,6-ba1.0mg/l,naa0.1mg/l,琼脂粉7g/l;ph 5.8-6.0)上,让叶片边缘紧密接触,然后在28℃黑暗条件下共培养2-3d;然后将共培后的烟草叶块转移至烟草筛选培养基(ms粉4.43g/l,蔗糖30g/l,6-ba1.0mg/l,naa0.1mg/l,琼脂粉7g/l,hyg 20mg/l,tim 100mg/l;ph 5.8-6.0)上,叶片正面朝上,并且使叶缘充分接触培养基,于24℃下,光周期为16h光照/8h黑暗条件下培养,每隔14d继代培养一次,诱导抗性芽的分化;抗性芽生长至约2cm时,小心切下,去除多余的培养基并转移到烟草生根培养基(ms粉4.43g/l,蔗糖30g/l,naa0.1mg/l,琼脂粉7g/l,hyg 20mg/l,tim 100mg/l;ph 5.8-6.0)进行生根培养;当抗性苗长至约5cm,进行炼苗并移植于实验室温室生长(图3)。使用ctab法提取烟草基因组dna,经pcr检测,结果显示,在野生型植株(wt)中检测不到目标片段存在,只有在转基因植株及阳性对照中扩增获得目标片段,表明外源基因eulac1已经导入烟草基因组(图4)。
[0045]
实施例3 eulac1基因组织表达特征
[0046]
选用primer 5.0软件,以克隆得到的eulac1基因序列为模板,依据荧光定量pcr引物的设计原则分析选用适合于eulac1的引物,并将其命名为eulac1-qtf和eulac1-qtr送至华大基因科技有限公司合成。eulac1-qtf:5
’-
gccctattcagaaagaccagac-3’;eulac1-qtr:5
’-
ttgccctaagccacgatatg-3’。选取杜仲euactin作为内参基因,以此来检验杜仲漆酶基因eulac1在幼树期杜仲根、茎、叶中的表达情况。选用实时荧光定量染料技术测定分析杜仲植株中漆酶基因eulac1的表达情况,使用select master mix试剂盒,按照试剂盒说明书,分步进行real-time pcr,按照δδct法计算和分析基因的表达水平,不同时期不同部位的样品设置三个生物学重复,每个生物学重复设置三个机械重复。结果表明,在幼树期杜仲的根和叶中,eulac1的表达量很低,幼树期的杜仲茎中的表达量是同时期叶的33.18倍,根的3.56倍,差异极显著(图5)。这些结果证明杜仲漆酶基因eulac1在茎中的表达量最高,在叶中的表达量最低。
[0047]
实施例4转基因烟草中eulac1表达量
[0048]
以烟草β-actin基因作为参照,用引物n-actin-f:5
’-
tgagatgcaccacgaagctc-3’,n-actin-r:5
’-
ccaacattgtcaccaggaagtg-3’(杨玲玲.烟草nad1基因克隆及结构分析[d].贵州大学,2018.),eulac1-qtf:5
’-
gccctattcagaaagaccagac-3’,eulac1-qtr:5
’-
ttgccctaagccacgatatg-3’,进行荧光定量分析。如图6所示,在转基因株系tp8中eulac1基因的相对表达量最低,株系tp13中表达量最高。株系tp13、tp10、tp20、tp9中eulac1的表达量都显著高于tp8,并且是eulac1表达量最高的四个株系,分别是株系tp8的19.33倍、18.91倍、17.72倍和17.59倍。
[0049]
实施例5烟草的漆酶活性
[0050]
挑选生长位置相同,生理状态一致的四个转基因烟草株系、转空载体的烟草和野生型烟草叶片。参照王骥等(王骥,朱木兰,卫志明.棉花漆酶基因在转基因新疆杨中的表达及其对木质素合成的影响[j].分子细胞生物学报,2008(01):11-18.)的方法测定漆酶活性,将植物组织用液氮研磨成粉状,加入蛋白抽提缓冲液(25mmol/l mops,200mmol/l氯化钙,ph 7.0),4℃放置4h,期间不时摇动。然后4℃,16000g离心10min,上清即为蛋白粗提物,以牛血清白蛋白为标准品,按照考马斯亮兰法进行蛋白含量测定。取30μl蛋白粗提物,加入1.4ml检测缓冲液(100μmol/l氯化铜,50mmol/l醋酸钠,2mg/ml abts,ph 4.5),30℃反应30min后,加入70μl冰醋酸终止反应,空白对照吸取30μl蛋白抽提缓冲液代替蛋白粗提物,用分光光度计记录420nm处的吸光度值,每个样品设置三个机械重复。如图7所示,与野生型烟草相比,转入空载体的烟草的漆酶活性并没有产生显著性的变化,而转eulac1基因的烟草的漆酶活性确有显著性的增加,t检验,p<0.01存在显著差异。其中eulac1相对表达量最高的株系tp13的漆酶活性最高,是野生型烟草漆酶活性的3.05倍(烟草品种为nicotiana tabacum cv.xanthin,普通烟草,其本身含有烟草漆酶蛋白,无杜仲漆酶,杜仲漆酶基因过表达可以提高烟草的整体漆酶活性,转基因烟草中的漆酶活性高于原有的烟草的。)。转基因株系tp10、tp20、tp9的漆酶活性分别是野生型的1.88,1.55和1.39倍。
[0051]
实施例6转基因烟草灰霉病抗性分析
[0052]
选取长势良好,木质素相对含量较高的转基因烟草和野生型烟草。配制0.7%的水琼脂培养基,取生长了四周的烟草植株相同位置的生理状态一致的叶片,用无菌刀片从叶柄处切下后,将叶柄用灭过菌的脱脂棉花包裹住,滴加适量的无菌水使棉花湿润保湿备用,每个株系选取三片叶子。配制新鲜的pda培养基,灭菌备用,将灰霉菌菌株从四摄氏度冰箱中取出,接种于配制好的pda培养基平板上,封口膜封装好后,放置在25℃的培养箱中暗培养,当菌斑直径长至8cm左右时,用打孔器取直径为5mm的菌饼(含有活性菌丝)。将取出的菌饼放置于备用叶片的同一位置,以接种pda固体培养基作为对照,将接种好的包含有菌丝菌饼的烟草叶片封闭于培养皿中于25℃暗培养。结果表明,在接种灰霉病病菌3d后,野生型烟草叶片出现明显的水浸状病斑,而转基因烟草叶片的病斑较小,扩散较慢。对病斑直径进行测量,发现转基因烟草叶片的病斑直径显著小于野生型(见图8)。
[0053]
实施例7烟草的木质素含量及烟草茎的横截面电镜观察
[0054]
选取生长周期相同的转基因烟草株系、转空载体的烟草和野生型烟草相同部位的叶片和茎段,木质素含量的测定采用乙酰溴法,实验步骤为:配制乙醇∶正己烷=1∶2(v/v)的混合溶液备用,将待测样品初步剪碎,准确称取1g碎裂的样品,加入适量的95%乙醇,将其研磨成匀浆,室温条件下在4500rpm转速下离心5min,倒掉上清,用移液枪吸干净;得到沉
淀,重新加入95%的乙醇,将其重悬,重复3次,再用准备好的乙醇正己烷混合溶液清洗2次,收集沉淀,将其放置于50℃烘箱干燥;配制25%溴乙酰冰醋酸(溴乙酰:冰醋酸=1:3,v/v)溶液,打开水浴锅调整至70℃,用移液枪将配好的溶液吸取2ml,加入到烘干的沉淀中,摇晃使沉淀溶解完全,置于准备好的水浴锅中,计时半小时;配制2mol/l的naoh溶液,配制7.5mol/l的羟胺盐酸,将0.9ml配制好的naoh溶液加入到上述溶液中,再取2ml冰醋酸和0.1ml配制好的羟胺盐酸,将其加到反应液中终止反应;用冰醋酸将上述溶液定容到5ml。室温下4500rpm离心5min,吸取上清液,用酶标仪分析记录在280nm处的吸光值,每个样品重复测三次。在280nm处测出的吸光值,表示了每克鲜样(fw)的木质素相对含量。结果表明,转基因烟草叶片和茎中木质素含量明显高于野生型烟草,而野生型和转基因烟草茎中木质素含量均明显高于叶片。且漆酶活性最高的株系tp13茎中的木质素含量最高(图9)。
[0055]
挑选移栽生长了1个月的野生型烟草和木质素含量较高的转基因烟草株系,用手术刀片取各植株相同部位茎的横截面样品(每株取两个样),加入2.5%的戊二醛固定剂浸没,放置于四摄氏度冰箱中过夜固定;固定结束后,将样品取出来,在超净工作台上,倒掉戊二醛固定液,用配置好的磷酸缓冲液,反复冲洗四至五次。然后用不同浓度的乙醇逐步清洗,浓度梯度如下所示:30%,50%,70%,80%,95%和100%,每次洗涤时间为15至20分钟。样品脱水洗涤结束之后,放入冷冻干燥机里面,干燥完毕后使用hitachi e-1010离子溅射设备,对样品镀金。最后,使用hitachi s3400扫描电子显微镜观察烟草茎部分的木质部细胞壁形态并拍照。如图10所示,转基因烟草木质部附近的细胞壁比野生型的厚且轮廓清晰,排列更加整齐。
[0056]
以上内容是结合具体的实施方式对本发明所作的进一步详细说明,不能认定本发明的具体实施只局限于这些说明。对于本发明所属技术领域的普通技术人员来说,在不脱离本发明构思的前提下,还可以做出若干简单推演或替换。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1