一种水稻分生组织强表达启动子OsNPY4及其应用的制作方法
一种水稻分生组织强表达启动子osnpy4及其应用
技术领域
1.本发明涉及生物技术和基因工程技术领域,具体而言,本发明涉及一种水稻分生组织强表达启动子及其应用,该启动子能够在水稻转基因体系中驱动目的基因在侧根和分蘖起始部分强烈表达部位。
背景技术:
2.启动子是指基因组中一段能准确起始转录的dna序列,通常位于基因的上游。启动子作为转录水平上一个重要调控元件,调控基因在不同发育阶段和组织的时空特异性表达(levine and tjian, 2003;smale and kadonaga, 2003)。根据启动子的功能和模式,可将它们分成三类:组成型启动子、特异性启动子和诱导型启动子。深入研究这些启动子的表达模式,不仅有助于我们理解植物的形态建成、生理代谢、逆境响应等基础理论,而且具有广泛的应用价值。
3.水稻作为最重要的粮食作物之一,同时也是一种优良的模式植物,一直是人们研究的重点。所以水稻分蘖数、穗的结构、根的结构(不定根和侧根的形成)等重要农艺形状一直受到重点关注,而水稻分蘖数、穗和根的结构都受到分生组织的影响(meng et al., 2009;wang et al., 2018)。
4.分生组织(meristem)是在植物体的一定部位,具有持续或周期性分裂能力的细胞群。分裂所产生的细胞排列紧密,无细胞间隙;细胞壁薄,细胞质浓厚,细胞体积较小,一般呈等径多面体,细胞核大,一小部分仍保持高度分裂的能力,大部分则陆续长大并分化为具有一定形态特征和生理功能的细胞,构成植物体的其他各种组织,使器官得以生长或新生(chongloi et al., 2019)。
5.生长素作为最早被发现的调控植物生长素发育的植物激素,它在分生组织的形成和维持过程中发挥重要作用,生长素相关基因几乎参与了植物所有的生长发育过程(wang et al., 2017)。我们通过对生长素相关基因表达模式的研究,不仅有助于阐述它们的调控机理,而且分生组织特异型表达启动子的研究十分有应用潜力。通过这些特异型启动子驱动外源基因表达,我们可以直接改良水稻的分蘖、侧根和不定根的形成还有穗型(如枝梗数目),这些性状都是培育水稻理想株型的重要指标(liang et al., 2014)。
技术实现要素:
6.本发明的目的在于提供一种在分生组织中强表达的启动子、含有该启动子序列的转化子以及启动子的应用。其中,本文中涉及“作物”是指禾本类作物,例如水稻、小麦、大麦、黑麦、燕麦、高粱、玉米等,优选是水稻。
7.为实现上述目的,本发明采用的技术方案如下:一方面,本发明提供一种植物分生组织强表达启动子,所述启动子包含seq id no:1所示的dna序列。序列表中seq id no:1所示的dna序列来源于日本晴水稻(oryza sativa l. japonica. cv. nipponbare)的分生组织强表达启动子,本文中称为osnpy4或
者启动子osnpy4。
8.优选地,本发明提供的水稻分生组织强启动子以中花11基因组dna作为模板pcr扩增,即osnpy4或者启动子osnpy4,其序列如seq id no:1所示。
9.另一方面,本发明提供一组用于扩增权利要求1所述启动子的引物序列,其特征在于,所述引物序列包括正向引物和反向引物,所述正向引物核苷酸序列:acggtatcgataagcttgatatcgaattcaaaacctgtctctctcccgc,所述反向引物核苷酸序列:tgggttaacccattccaactagaattctgtctggaagaggtctggct。
10.另一方面,本发明提供一种重组表达载体,所述表达载体包含上述的水稻分生组织强表达启动子。在所述重组表达中,所述的水稻分生组织强表达启动子连接于待表达基因上游。优选地,所述待表达基因是gus基因,所述重组表达载体为pplv15-osnpy4,该重组表达载体是将seq id no:1所示dna序列即osnpy4或者启动子osnpy4构建于pplv15得到的重组载体,本文中称为pplv15-osnpy4。
11.或者待表达基因是对作物的分蘖或者根性状具有改良能力的基因。通过本发明的启动子驱动该基因在分生组织中强烈表达,从而实现理想株型的改良。
12.再一方面,本发明提供上述水稻分生组织强表达启动子在培育转基因作物中的应用。所述应用包括将本发明提供的上述水稻分生组织强表达启动子连接于待表达基因的上游,从而构建重组表达载体,将所述重组表达载体转化到细胞、组织或者器官中进行培育。
13.另一方面,本发明提供一种在分生组织中驱动特定待表达基因表达的方法,其特征在于,所述方法包括:1)将权利要求1中所述的水稻分生组织强表达启动子osnpy4连接与待表达基因上游;2)将所述水稻分生组织强表达启动子osnpy4和待表达基因的连接产物转入根癌农杆菌中;3)利用转入所述连接产物的根癌农杆菌对目标植物的组织例如叶片或者种子进行转化;4)利用转化后的组织培育相应的转基因植株,在所述转基因植株中,权利要求1所述的水稻分生组织强表达启动子osnpy4驱动待表达基因表达。
14.优选地在所述方法和所述应用中,所述待表达基因为结构基因、调节基因、结构基因的反义基因或者能够干扰内源基因表达的小rna,优选为具有改良分蘖和根性状的基因。
15.综上所述,本发明的发明人发现、鉴定了水稻日本晴(oryza sativa l. japonica. cv. nipponbare)一段具有转录调控活性的3661bp的dna序列,并将其命名为osnpy4(序列表中的seq id no:1)。具体而言,发明人发现该序列具有驱动基因在分生组织强表达的能力,扩增出该序列并对上述能力进行了鉴定。发明人扩增出目的片段后,利用gibson assembly方法将其连接到pplv15载体上,获得相应的重组质粒,阳性检测后将重组表达载体转化根癌农杆菌eha105。然后用农杆菌介导方法进行水稻转化,得到转基因植株。对获得的转基因植株进行组织染色检测,结果先osnpy4驱动待表达基因在分蘖、不定根、侧根的形成部位的分生组织中强烈表达,证明了权利要求1中的seq id no:1在分生组织中有强烈的表达活性。
16.本发明的所述的启动子序列可与作物的表达载体连接,用于取代组成型启动子。
该启动子序列可以与所需靶基因序列,构建重组表达载体,转化后靶基因可以在分生组织强烈表达。而且特异型启动子可以避免组成型启动子造成的外源靶标基因在其它组织的过量表达,影响作物农艺性状。
17.本发明的技术效果表明:利用本发明克隆的分生组织强表达启动子可以在水稻分蘖、不定根和侧根起始部位强烈表达,在实际运用中具有显著价值。通过该启动子对农作物进行基因改良,例如通过该启动子驱动生长素相关基因在分生组织中的表达,调控分蘖、侧根和不定根的产生,为理想株型的建立提供新的策略。并且,由于生长素路径在植物中的保守性,发明人认为上述启动子可以在作物中可以广泛的发挥功能。
附图说明
18.以下,结合附图来详细说明本发明的实施方案,其中:图1是将osnpy4启动子构建与pplv15载体质粒中的示意图。其中图a是pplv15示意图,图b是pplv15-osnpy4示意图,其中示出了利用osnpy4驱动下游gus基因表达;图2是osnpy4-gus转基因植株各组织gus染色示意图。(a)分蘖产生部位;(b)根尖;(c)侧根起始部位;(d)叶尖;(e)颖壳维管组织;(f)雌蕊柱头;(g)雄蕊。
具体实施方式
19.下面将结合本发明实施例中的附图,对本发明中的技术方案进行清楚、完整的说明。本领域技术人员能够理解,这些实施例仅用于说明本发明,而不以任何方式限制本发明的范围。
20.下述实施例中的实验方法,如无特殊说明,均为常规方法。下述实施例中所用的试剂材料,如无特殊说明,均为市售购买产品。
21.实施例1 启动子osnpy4的获得1)引物设计根据ncbi(www.ncbi.nlm.nih.gov)提供的水稻品种日本晴(oryza sativa l. japonica. cv. nipponbare)全基因组序列,依据osnpy4基因序列设计扩增引物,并根据选用的表达载体和酶切位点设计引物接头。
22.本发明中选用的是水稻表达载体pplv15(rybel bd et al., 2011),报告基因是gus。本发明中重组载体的构建采用gibson assembly方法,扩增引物上需要增加接头引物(下划线),具体引物序列如下:正向引物:acggtatcgataagcttgatatcgaattcaaaacctgtctctctcccgc反向引物:tgggttaacccattccaactagaattctgtctggaagaggtctggct由武汉擎科生物科技公司合成。
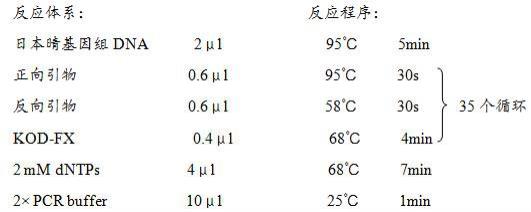
23.2)启动子osnpy4扩增提取水稻中花11(zh11,来自中国农业科学院作物科学研究所)叶片基因组dna作为模板,用正向引物和反向引物进行pcr扩增,具体pcr体系和条件如下:
补双蒸水至20μl目的片段大小3717bp,经凝胶电泳后挖胶回收(使用qiagen公司的回收柱)。
24.实施例2 植物表达载体的构建及转化1)重组表达载体构建并用gibson assembly一步法(gibson et al., 2009)将纯化过的osnpy4启动子的pcr产物和ecori切过的载体pplv15连接,50℃反应半小时,取1μl电转(1800v)大肠杆菌感受态top10,并涂布在含卡那霉素(50
µ
g/ml)抗性的lb培养基上。挑取单克隆菌落,抽提质粒并进行阳性检测,将质粒dna送测序公司测序核实,将阳性质粒命名为pplv15-osnpy4。
25.取1μl测序正确的osnpy4重组表达载体和1μl psoup 质粒混合电转(1800v)农杆菌感受态eha105,挑单克隆菌落培养1天并进行阳性检测,将阳性菌液与50%甘油1:1混合后-80℃保存。
26.2)重组表达载体转化水稻取osnpy4重组表达载体菌液侵染中花11愈伤,参考hiei y等人建立的农杆菌遗传转化方法(hiei y et al., 1994),具体转化步骤如下:1)将水稻品种中花11的成熟胚去壳,用75%乙醇泡1min,0.15%升汞消毒20min,无菌水洗3~4次;将所得的外植体接种到诱导培养基上,于26℃暗培养诱导愈伤组织;2)诱导培养35d后,取活力强、颗粒状的愈伤组织转入继代培养基进行继代培养;3)取继代培养20d的愈伤组织的颗粒,接入预培养培养基上,于26℃下暗培养4d;4)在预培养的第三天,用la(lb中加入1.5%琼脂)划线接种农杆菌菌株(pplv15-osnpy4),28℃静止培养2d;之后将农杆菌全部刮入悬浮培养基;于28℃,200r/m振荡培养0.5-1h;在分光光度计600nm波长光测定菌液浓度,调至od值=1.0;5)将预培养后的愈伤组织接入100ml锥形瓶(大约到40ml处),加入调制好的农杆菌菌液,浸泡30min,期间摇动数次。配制悬浮培养基:(500
µ
l乙酰丁香酮as(acetosringone)+5ml50%葡萄糖;6)倒去菌液,将愈伤组织置于灭菌滤纸上吸干表面菌液(一定要吸干菌液,使愈伤组织的表面发白),接入共培养培养基(250
µ
l乙酰丁香酮as+5ml50%葡萄糖),暗培养3d,再转入250ml共培养培养基上进行共培养;7)将共培养后的愈伤组织用无菌水先快速摇动清洗两次;然后加入无菌水浸泡10min,使愈伤组织内部的菌体游离出来;倒去洗液,再加入含400mg/l的羧苄青霉素(cn)无菌水浸泡15min;倒干洗液,将愈伤组织置于灭菌滤纸上吸干,接入筛选培养基;26℃暗培
养。每3周继代一次,共继代两次。每转化一个质粒,需灭菌单蒸水1~2瓶,其中第一次筛选时,在300ml筛选培养基中加入500ul羧苄青霉素(cn)和300ul潮霉素(hn);第二次继代筛选时,在筛选培养基中加入400ul羧苄青霉素(简称cn)和300ul潮霉素(简称hn)。
27.8)将筛选培养基培养的抗性愈伤组织接入预分化培养基,26℃暗培养一周。
28.9)将预分化培养一周的抗性愈伤组织转入分化培养基(50ml/瓶;改用三角瓶或平底试管做培养瓶);25℃,2000 lux光照培养,通过再生获得转基因植株。
29.10)待小植株3~5cm;转入生根培养基上促进生根。
30.11)将根系健壮的植株移入盆钵,凉棚过渡3~5d;然后移到自然条件下生长,直至成熟。
31.实施例3 利用启动子osnpy4驱动gus报告基因在水稻中表达1)转基因阳性植株鉴定取转基因植株叶片,经ctab法抽提dna样品。根据表达载体上的潮霉素抗性基因设计检测转基因阳性引物,引物序列如下:hyg-f:acggtgtcgtccatcacagtttgcchyg-r:ttccggaagtgcttgacattgggga取2μl转基因植株dna样品进行检测,具体体系和程序如下:补双蒸水至20μl。
32.将pcr产物电泳检测,目的片段是289bp。
33.2)转基因植株组织gus染色参照jefferson等提出的方法(jefferson, 1989),将转基因阳性植株的各部分组织放入gus染液中抽真空十分钟,然后37℃处理一小时。等代表gus活性的蓝色在组织上出现后用75%乙醇在37℃条件下脱色,中间更换75%乙醇脱色直至叶绿素完全脱去。结果见图2,在gus染液中处理12小时后在侧根形成部位还有根茎结合部位——分蘖产生部位有强烈蓝色信号出现,说明osnpy4在分生组织部位有强烈的表达。此外,作为一个生长素相关基因,osnpy4在受生长素调控的根尖、颖壳、雄蕊和雌蕊上也有表达,在这些组织中均观察到蓝色信号。
34.虽然本发明以水稻为例进行说明,但是申请人认为,本发明所获启动子的应用场景不限于水稻,可以用于其他禾本科作物,例如小麦、大麦、玉米、高粱或者燕麦。
35.以上对本发明具体实施方式的描述并不限制本发明,本技术领域人员可以根据本
发明做出各种改变或变形,只要不脱离本发明精神,均应属于本发明权利要求的范围。
36.参考文献:wang b., smith s.m. and li j.y., genetic regulation of shoot architecture,annu. rev. plant biol, 2018. 69:25.1
–
25.32levine m. and tjian r., transcription regulation and animal diversity,nature, 2003smale s.t. and kadonaga j.t., the rna polymerase ii core promoter, annu. rev. biochem. 2003. 72:449
–
79meng f.n., xiang d, zhu j.s., li y. and mao c.z., molecular mechanisms of root development in rice, rice, 2009. 12:1liang w.h., shang f., lin q.t., lou c. and zhang j., tillering and panicle branching genes in rice, gene, 2014. 537:1
–
5chongloi g.l. , prakash s. and vijayraghavan u., rice shoot and floral meristem development: an overview of developmental regulators of meristem maintenance and organ identity, j. exp. bot.,2019.rybel b.d., willy van den berg, lokerse a.s., liao c.y., mourik h., barbara mo
¨
ller, llavata-peris c.i., and weijers d., a versatile set of ligation-independent cloning vectors for functional studies in plants, plant physiol., 2011. 156: 1292
–
1299.gibson, d.g., et al., enzymatic assembly of dna molecules up to several hundred kilobases. nat methods, 2009. 6(5): p. 343-5.jefferson r.a., the gus reporter gene system, nature, 1989. 342(6251):837-8.hiei y., ohta s., komari t. and kumashiro t., efficient transformation of rice (oryza sativa l.) mediated by agrobacterium and sequence analysis of the boundaries of the t-dna, plant j., 1994. 6(2), 271-282wang y, zhang t, wang rc and zhao y, recent advances in auxin research in rice and their implications for crop improvement, j exp bot., 2018. 69(2):255-263序列表seq id no:1是osnpy4基因的启动子核苷酸序列。序列长度为3661bp。
37.aaaacctgtctctctcccgctgataatgccaggaacattttgtccgtctattgactgctacactcatgtcgtgtccctcatggagaaaacaattagagattttttaaaggatatacaaggataaccttttctttttgagaacatatgcaaggataactacagttactttttgtggtacgttacactgaagaagattatggatcgagaaaattgctgaagaaatcgcgacagcatgttgacaagagatcccagctaacctatctttgtttgtaaaatatgtcatgccgtgcaattttaacgaccgttcgcgcattggctaagttaattactctactagagatatagccaaacatttctagttattttattatttttatatgtaaaaaaaattaactacatatcttattattatctatattatatatatgtatatattaaactttatatgtaattattcatttgttgtttataaattaaagaaaatatcattatccacactattatcgttaaatttaagctttataagataagataaaatatatgtatttatcttagttattttaaaaacacaatctctaaaccttttgaaaaaaaatcatc
ttcctctataaaaattgcatctccccccctaaaaaaagtacaatgaatctatattttggtttggcgcagaaaaaaattagtaacactaaatttcaaaatgatacaacacatattaattcttcgttttaactagatagtgaaaataaacttgttagcttacattttacatatagatatatagatgtatatggacgagctatatggctagagcaccaacgaaatataaagagacgataagagggccatgcatggtgagtactacgggaaggcagcaacaacgacgagcgtgtgttatgacacaggagggatacaagtatagtgtgtgtgatgagttttgaaaaaaaaatagtgatatgtgtgtccgtatttttcatgcatgtatttggaacaaaaatatgtttgtattctcactaaactttttgactttcatttactgtaattcattatttactttgttatcataaatttcagctaattactatcaacaacacgtaaacatattttaaatagaaagggtaatagtcgcgtgaacacgagacataggctgtgtttacatccaggggtgtaaagttttagtatgtcacatcgaatatacggacacatatttgaagtattaaacctagtctaataacaaaataaattacagaatccgcctgtaaaccgcgagacgaatttattaagcctaattaatccatcattagcaaatgtttactgtagcaccacattgtcaaatcatgatgcaactaggcttaaaagattcgtctcgcaatttacacgcgattctatgtaattagttattttttcgtttatatttaatatttcatgcatgtgtctaaatattcgatatgatagggtgtaaaattttgccagggaatttaaataggcccataatatgctagtagttaattgattaggcatacaccatacacaaaaccaaaatatttacatgtgaggatctcatggctccatatattagcttgtaacattgaaggcattatttaaagtagtacttgctccgtccccagttacagggcgctctttttttagaaaaattaaactcggtaatttttaactaataatcatactaatcatatatatgctagttatatcactatatttatatttttatattttaaagtactttcatattatgctaattttatatttattgaaaatataacatgaaataaattaatggttaaaatatattatattattggagatcgtataaaatataagccttataatttaataataactgagataaatatatactccctctgtcccaaaatataactatttttaacatcgtcacagtttttaagatgctactttgaccaacaacatctataaaaataaaatgttttaaataaaaagagttgcatattataatagtttgattaatgataaatctagtaagatcaattttacatgattgatcttttttattttattactattaatagttaattttaaaaatagttaatttgttattatgttaaaaaaaattatattttgagacacggagcgagttcattttatctcgagcgtgtaatagcgaaaaaaaatagtaggcgtgtaatagagaaaaaatcgagataaagacagtgtacttgagtgtctgcccccttcccttggcgtataaaagaaaatgtcccgtgtcgttctcctccgttcttgcgccatctcgcggctggacaggggaggggcgccgatctccatctggcgagcagagcagggcaggggaaggggatcctggtgagcatccacatcctctttctgattcatctctcactctcccaccggctgtgccttttgtctggaatttgctttccgttcgtccacccttgcttctcttctagatctggaagaagctcttgtcttaattccagagccttattcttaagacaagtaacagttttgtttgttcccccaaaagtttttggggttcaactgaacccccccccccatgtcccaagttggatccgctcctgctgtggcaatcccttctcgtgttacaacttacagaagctgattctaattcacattcgttgcaaaatagtataactgagtgtgttttgttgagatgtgtgtccttacatgtcgattctttaggcttgatctcacaaatgcataatgggtatcatcaagttagtttttttcctcataaattagttgtcttttttttttttggagggaatggtactgctagtttcatgttgcaatcttccgttgccatttactggtctttctgctcctatggtaaaattatcagaaatttatccgaacttccatatgcacaattcaatattgttgccagctaagatgggagaagaaaaaaaaaacataaactgtggactatgggtggcaattctttaatcccagtttgactgccttcatgtgcttacacccttaaaaaaaatcatcctgtatacactcttgttattcatgttgcttgcttgcaattttaaccaagataagattccctcccctcactgctgcttgttcccatggtctcccaacattgcttcaccaattattcccactgtcccctcacctcaagatgctgaccttgtgttctgcaggttctcagcgtgtgtgatgtgatccagcttgggggaggaagagcttgttggcctcttggcattgcggtctcctggacccagatcttgatgcaactgccgactactttttttttttttactgatagaggtggtctggttgggttcttgacctgattttgtgggggctttgccaccacgcaagggaggcagtttggcctggatctttatcactagctgttggtcatgtcctttgagctagcaagcattcacatggcagtggccgccggtttggtacattcttgatgggttaagtgctgtgggtttgttgtggagttctcttcctggttcccatttggggtggacggtatgtgttgaagggaaaggggcagagatagaaaga
gcaaaaagaagataggagttcttcggttaattgtttttttcagtacggcgcaatgaagcatatgaaacttggatcaaagccagacctcttccagaca。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1