一种编码双特异性T细胞衔接子的溶瘤痘苗病毒及其制备方法和应用与流程
一种编码双特异性t细胞衔接子的溶瘤痘苗病毒及其制备方法和应用
技术领域
1.本发明属于基因治疗和生物医药技术领域,具体涉及一种表达双特异性t 细胞衔接子的溶瘤痘苗病毒及其制备方法和应用。
背景技术:
2.传统的肿瘤治疗方法包括手术、放疗、化疗等在晚期肿瘤治疗中治疗效果有限且毒副作用严重,近年来,随着细胞生物学、分子生物学及生物工程技术的迅速发展,癌症的免疫疗法取得了重大突破,其中溶瘤病毒治疗以其突出的效果引起了广泛关注。
3.溶瘤痘苗病毒是双链dna病毒,具有一些独特的优点:(1)痘苗病毒能够在肿瘤细胞中快速复制并裂解细胞;(2)通过膜融合的方式感染肿瘤细胞,不需要特定的细胞表面受体,因此对肿瘤细胞的靶向性较广泛,能感染多种肿瘤细胞; (3)痘苗病毒始终存在于细胞质中,不会整合到宿主基因组中;(4)病毒感染细胞后的产物能自我包裹,病毒能通过血流屏障远距离传播,因此具有很好的全身抗肿瘤作用;(5)病毒能容纳较大的外源性基因插入;(6)即使溶瘤痘苗病毒治疗过程中发生副作用,可用临床上常用的抗病毒药物控制;(7)痘苗病毒颗粒比较稳定,可以冻干,以干粉形式储存,因而易于运输和临床应用。
4.早期研究表明,虽然溶瘤痘苗病毒具有很高的安全性,临床研究中也取得了一定的疗效,但是溶瘤痘苗病毒的抗肿瘤效果仍不够理想,可能归因于肿瘤细胞的各种免疫逃避机制限制了肿瘤特异性t细胞活化及保持,因此需要对其改造增强其抗癌效果。
5.双特异性t细胞衔接子(bispecific t cell engager,bite)提供了一种将t细胞接合至肿瘤细胞的方式且临床上显示出有效治疗成果,比如治疗非霍奇金氏淋巴瘤(non-hodgkin's lymphomas)和前驱b细胞急性淋巴母细胞白血病,然而双特异性t细胞衔接子半衰期短,临床上需要连续输注,导致全身性毒副作用。
6.本发明以溶瘤痘苗病毒wr株为研究模式,发现搭载靶向肿瘤抗原和效应细胞的双特异性t细胞衔接子可显著增强溶瘤痘苗病毒的抗肿瘤效果。本发明将着重阐述表达双特异性t细胞衔接子的溶瘤痘苗病毒制备方法及其应用。
技术实现要素:
7.针对现有技术中溶瘤痘苗病毒虽然可以诱导t细胞在肿瘤组织内有效浸润,但肿瘤细胞的各种免疫逃避机制限制了肿瘤特异性t细胞活化,降低了溶瘤痘苗病毒的抗肿瘤效果等问题,本发明的第一个目的是提供一种编码双特异性t 细胞衔接子的溶瘤痘苗病毒;本发明的第二个目的是提供一种编码双特异性t 细胞衔接子的溶瘤痘苗病毒的制备方法;本发明的第三个目的是提供一种编码双特异性t细胞衔接子的溶瘤痘苗病毒的应用。
8.本技术具有如下技术方案:
9.一种编码双特异性t细胞衔接子的溶瘤痘苗病毒,所述病毒基因组的胸腺嘧啶核苷激酶(tk)区包含编码双特异性t细胞衔接子基因序列,并且能够感染肿瘤细胞使其表达
双特异性t细胞衔接子。
10.其中,该双特异性t细胞衔接子一端特异性结合肿瘤细胞表面抗原epcam,另一端特异性结合t细胞表面受体cd3,所述溶瘤痘苗病毒能够感染肿瘤细胞使其表达双特异性t细胞衔接子。
11.在溶瘤痘苗病毒中搭载双特异性t细胞衔接子,病毒能够在肿瘤局部大量表达双特异性t细胞衔接子,介导t细胞靶向杀伤肿瘤,且该杀伤作用不依赖 mhc分子,因此可以克服肿瘤细胞通过下调mhc分子形成的免疫逃逸,增强t 细胞对肿瘤细胞的免疫应答,抑制活化t细胞的耗竭,从而发挥多重的抗肿瘤效应。
12.作为本技术的优选技术方案,所述的双特异性t细胞衔接子是由特异性识别肿瘤相关抗原epcam的单链抗体(scfv)、接头多肽、特异性结合t细胞表面受体cd3的单链抗体(scfv)组成,其核苷酸序列如seq id no.1所示。
13.作为本技术的优选技术方案,所述特异性识别肿瘤相关抗原epcam的单链抗体的轻链可变区氨基酸序列如seq id no.2所示。
14.作为本技术的优选技术方案,所述特异性识别肿瘤相关抗原epcam的单链抗体的重链可变区氨基酸序列如seq id no.3所示。
15.作为本技术的优选技术方案,所述的特异性结合t细胞表面受体cd3的单链抗体的轻链可变区氨基酸序列如seq id no.4所示。
16.作为本技术的优选技术方案,所述的特异性结合t细胞表面受体cd3的单链抗体的重链可变区氨基酸序列如seq id no.5所示。
17.作为本技术的优选技术方案,所述接头多肽为ggggs。
18.所述的溶瘤痘苗病毒能够裂解肿瘤细胞。
19.所述的溶瘤痘苗病毒表达的双特异性t细胞衔接子一端能够特异性结合肿瘤细胞表面抗原epcam,另一端能够特异性结合t细胞表面抗原cd3。
20.所述的编码双特异性t细胞衔接子的溶瘤痘苗病毒的制备方法包括以下步骤:
21.(1)在双特异性t细胞衔接子基因序列前后分别增加ecor1识别序列gaattc、 kozak序列gccacc和xba1识别序列tctaga,然后人工合成上述序列;
22.(2)采用ecor1和xba1双酶切上述合成的基因片段后将其亚克隆至穿梭质粒pvv-control对应的酶切位点,构建pvv-epcam-bite质粒;其中双特异性t 细胞衔接子的表达受痘苗病毒早晚期启动子pse/l控制;
23.(3)采用基因同源重组的方式,用pvv-epcam-bite质粒转染已经预先感染了野生痘苗病毒(wr株)的细胞,使质粒和野生痘苗病毒发生同源重组,产生重组溶瘤痘苗病毒vv-epcam-bite。
24.作为本技术的优选技术方案,所述双特异性t细胞衔接子基因序列为seqid no.1。
25.本发明还保护上述编码双特异性t细胞衔接子的溶瘤痘苗病毒在制备治疗或预防肿瘤的相关药物中的应用。
26.优选的,所述的肿瘤选自乳腺癌、大肠癌、肝癌、黑色素瘤、肺癌、胃癌、胰腺癌、胆囊癌、肾癌、膀胱癌、前列腺癌、卵巢癌、宫颈癌、子宫内膜癌、头颈癌、胶质瘤、癌性腹水、癌性胸水中的任一种;
27.更优选的,所述肿瘤为乳腺癌或结肠癌。
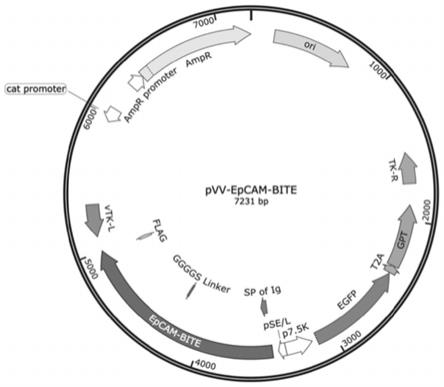
28.有益效果
29.与现有技术比较,本发明的溶瘤痘苗病毒可以发挥以下抗肿瘤效应:
30.(1)本发明将溶瘤痘苗病毒的溶瘤效应与双特异性t细胞衔接子的抗肿瘤效应有效结合,制备了一种可以表达双特异性t细胞衔接子的溶瘤痘苗病毒。一方面,病毒可以在肿瘤细胞中复制,发挥溶瘤效应;另外一方面,病毒可以有效诱导t细胞浸润,使“冷”肿瘤微环境转化为“热”肿瘤微环境。病毒可以感染肿瘤细胞并且使其表达双特异性t细胞衔接子,后者一端特异性结合肿瘤细胞表面抗原epcam,另一端特异性结合t细胞表面受体cd3,介导t细胞靶向杀伤肿瘤细胞,且该杀伤作用不依赖mhc分子,因此可以克服肿瘤细胞通过下调 mhc分子形成的免疫逃逸,增强t细胞对肿瘤细胞的免疫应答,从而发挥多重的抗肿瘤效应。
31.(2)在痘苗病毒基因组tk区插入双特异性t细胞衔接子基因导致病毒复制依赖的tk基因失活,使病毒选择性的在tk高活性的肿瘤细胞内复制,显著增强了溶瘤痘苗病毒的安全性。
附图说明
32.图1是溶瘤痘苗病毒vv-epcam-bite的模式图。
33.图2是痘苗病毒穿梭质粒pvv-epcam-bite的图谱。
34.图3是溶瘤痘苗病毒vv-epcam-bite感染hela-s3细胞产生的病毒空斑。
35.图4是western blot方法检测双特异性t细胞衔接子在溶瘤痘苗病毒 vv-epcam-bite感染的hela-s3细胞培养上清中的表达。从图中可知,双特异性t细胞衔接子的分子量大约为70kd。
36.图5是流式细胞术检测溶瘤痘苗病毒vv-epcam-bite感染肿瘤细胞后分泌的双特异性t细胞衔接子对乳腺癌细胞系4t1表面epcam的封闭作用。
37.图6是mtt试验检测溶瘤痘苗病毒vv-epcam-bite对乳腺癌细胞系4t1、结肠癌细胞系ct26和黑色素瘤细胞系b16/f10的体外溶瘤能力。从图中可知,随着感染复数(moi)的增加vv-epcam-bite的溶瘤能力逐渐增加。
38.图7是空斑法检测溶瘤痘苗病毒vv-epcam-bite在乳腺癌细胞系4t1、结肠癌细胞系ct26和黑色素瘤细胞系b16/f10的体外病毒复制。
39.图8是瘤内注射溶瘤痘苗病毒vv-epcam-bite对表达epcam的乳腺癌 4t1皮下移植瘤模型的治疗作用。从图中可知,与pbs以及对照病毒vv-control 比较,vv-epcam-bite可以显著抑制4t1皮下移植瘤的生长,延长小鼠生存时间。
40.图9是瘤内注射溶瘤痘苗病毒vv-epcam-bite对结肠癌mc38-epcam
+
皮下移植瘤模型的治疗作用。从图中可知,与pbs和对照病毒vv-control相比, vv-epcam-bite可以显著抑制mc38-epcam
+
皮下移植瘤的生长,延长小鼠生存时间。
具体实施方式
41.本发明结合以下实施例进一步说明,除特别说明外,本发明可采用本领域的常规技术,并且本发明的范围不受这些实施例的任何限制。
42.实施例1:穿梭质粒pvv-epcam-bite的构建
43.在双特异性t细胞衔接子基因序列(seq id no.1)前端分别增加核酸内切酶ecor1识别序列gaattc和kozak序列gccacc,末端增加核酸内切酶xba1 识别序列tctaga,然后采用人工合成的方法由南京金斯瑞公司合成上述序列,并且将其亚克隆至pvv-control质粒(专利cn202010790895.9公开的seq idno.4)对应的酶切位点ecor1和xba1。新合成的质粒命名为pvv-epcam-bite,其质粒图谱如图2所示。
44.实施例2:溶瘤痘苗病毒包装和筛选
45.用含有10%fbs的dmed高糖培养基常规培养hek293细胞,用0.5%的胰蛋白酶消化细胞,计数,以3
×
105cells/孔接种于6孔板中,置于37℃,5%co2培养箱中培养。待细胞生长到90%以上融合度时,加入野生痘苗病毒(wr株,购自atcc,保藏号:vr-1354)感染2小时(moi=1)。将质粒 pvv-epcam-bite采用转染试剂(polyplus-transfection公司)转染预先感染野生痘苗病毒的hek293细胞,转染后的hek293细胞置于37℃,5%co2培养箱中培养,使质粒和野生痘苗病毒发生同源重组,产生溶瘤痘苗病毒 vv-epcam-bite,其模式图如图1所示。溶瘤痘苗病毒vv-epcam-bite在野生病毒tk区插入鸟嘌呤-次黄嘌呤磷酸核糖转移酶(gpt)基因和荧光蛋白报告基因(egfp),两者采用2a肽进行连接,其表达受p7.5k启动子控制;插入的双特异性t细胞衔接子基因的表达受痘苗病毒早晚期启动子pse/l控制,其基因序列如seq id no.1所示。对照病毒vv-control的包装类似于 vv-epcam-bite,通过pvv-control质粒与野生痘苗病毒发生同源重组获得。
46.溶瘤痘苗病毒的筛选采用空斑纯化法,在转染后48小时,待溶瘤痘苗病毒产生空斑后(如图3所示),在含有25μg/ml霉酚酸(mpa),250μg/ml黄嘌呤(xanthine)和15μg/ml次黄嘌呤(hypoxanthine)的筛选培养基的条件下采用空斑纯化法筛选去除野生病毒,获得不含野生病毒的痘苗病毒vv-control和 vv-epcam-bite。然后采用病毒基因组提取试剂盒(康为世纪公司)提取溶瘤痘苗病毒的基因组,采用以下引物分别扩增目的基因和野生病毒胸苷激酶(tk) 基因,扩增条件为98℃3分钟,98℃10秒,55℃5秒,72℃20秒,30个循坏, 72℃5分钟。目的基因:5
’‑
caggtgatctgtttttattgtggag-3’,5
’‑
gatctacttccttaccgtgc-3’; tk:5
’‑
tgtgaagacgataaattaatgatc-3’,5
’‑
gtttgccatacgctcacag-3’。pcr成功扩增目的基因但是未同时扩增野生病毒tk基因的病毒为筛选成功的溶瘤痘苗病毒。筛选成功的溶瘤痘苗病毒进一步采用hela-s3细胞在细胞培养皿中进行扩增。采用 tcid50法测定病毒的滴度,计算公式如下:病毒滴度=0.7
×
10
×
10^(1+s (d-0.5)),其中s=log10(稀释度),d=各个稀释度egfp阳性比率之和。
47.实施例3:溶瘤痘苗病毒分泌双特异性t细胞衔接子及其功能鉴定
48.用含有10%fbs的dmed高糖培养基培养hela-s3细胞,用0.5%的胰蛋白酶消化细胞,计数,以5
×
105个细胞/孔接种于6孔板中,置于37℃,5%co2培养箱中培养。待细胞生长至90%以上融合度时,加入溶瘤痘苗病毒感染(moi =0.1)。48小时后收集细胞培养上清,以抗flag标签抗体作为第一抗体,westernblot检测细胞培养上清中双特异性t细胞衔接子的表达。如图4所示,溶瘤痘苗病毒vv-epcam-bite感染的hela-s3细胞可以在细胞培养上清中分泌双特异性t细胞衔接子,其分子量大约为70kd。
[0049]2×
105个乳腺癌4t1细胞分别与100μl感染了vv-epcam-bite的细胞培养上清或感染了vv-control的细胞培养上清室温共孵育,20分钟后,加入1ml
ꢀꢀ
pbs混匀,1000rpm/min,离心5min,去除上清,用100μl pbs重悬细胞,分别用isotype-pe、epcam-pe单克隆抗体
(ebioscience公司)标记4t1细胞。室温避光孵育15分钟,加入4%多聚甲醛对细胞进行固定。用facs calibur流式细胞仪(bd公司)进行检测,采用flowjo软件进行数据分析。如图5所示, 4t1细胞与含有分泌的双特异性t细胞衔接子细胞上清孵育后,epcam表达的峰值前移,提示vv-epcam-bite可以表达双特异性t细胞衔接子,并且结合和封闭4t1细胞表面的epcam分子。
[0050]
实施例4:溶瘤痘苗病毒对肿瘤细胞的体外溶瘤能力
[0051]
采用mtt试验定量分析溶瘤痘苗病毒对肿瘤细胞的溶瘤能力。用含有10% fbs的dmed高糖培养基培养乳腺癌细胞4t1、结肠癌细胞ct26和黑色素瘤细胞b16/f10,用0.5%的胰蛋白酶消化细胞,计数,以1
×
104细胞/孔接种于 96孔板中,置于37℃,5%co2培养箱中培养。待细胞生长到90%以上融合度时,分别加入不同感染复数(0、0.1、1、5、10)的溶瘤痘苗病毒。培养72小时后,每孔加入20μl噻唑蓝(mtt)继续培养。4小时后,去除上清液并加入 150μl异丙醇溶解甲瓒(formazan),酶标仪570nm处检测吸光度(od)值,根据以下公式计算细胞活率:细胞活率(%)=(实验孔-空白对照孔)/(对照孔
ꢀ‑
空白对照组)
×
100%。如图6所示,随着感染复数(moi)的增加, vv-epcam-bite对4t1、ct26和b16/f10细胞的溶瘤能力均逐渐增加。
[0052]
实施例5:溶瘤痘苗病毒在肿瘤细胞中的复制能力
[0053]
用含有10%fbs的dmed高糖培养基培养乳腺癌细胞4t1、结肠癌细胞 ct26和黑色素瘤细胞b16/f10,用0.5%的胰蛋白酶消化细胞,计数,以1
×
105细胞/孔接种于24孔板中,置于37℃、5%co2培养箱中培养,24h后分别加入感染复数为0.1的溶瘤痘苗病毒。细胞感染病毒24、48、72、96h后,采用tcid50 法检测病毒的复制能力。如图7所示,在所有这些细胞中,vv-epcam-bite溶瘤痘苗病毒均可以复制。在4t1和b16/f10细胞中,48小时的病毒滴度达到峰值。在ct26细胞中,72小时的病毒滴度达到峰值。
[0054]
实施例6:溶瘤痘苗病毒对乳腺癌皮下移植瘤的治疗作用
[0055]
在本实施例中,先用含有10%fbs的dmed高糖培养基培养4t1乳腺癌细胞,然后用0.5%的胰蛋白酶消化细胞并且进行细胞计数,取1
×
106个细胞分别用isotype-pe和epcam-pe单克隆抗体(ebioscience公司)标记细胞。室温避光孵育15分钟,加入4%多聚甲醛对细胞进行固定。用facs calibur流式细胞仪(bd公司)进行检测,采用flowjo软件进行数据分析。如图8所示,4t1 乳腺癌细胞系表面高表达epcam。
[0056]
用4t1细胞建立乳腺癌皮下瘤模型,选用6-8周龄雌性balbic小鼠(spf 级,购于南京大学模式动物研究所),每只小鼠皮下注射2
×
105个4t1细胞。在建模大约6天后,肿瘤直径大约长至5mm左右时,将小鼠随机分为3组,分别为pbs对照组,vv-control和vv-epcam-bite治疗组,分别瘤内多位点注射 pbs、vv-control和vv-epcam-bite进行治疗。病毒注射的剂量为每次1
×
10
7 pfu,每2天进行1次治疗,共治疗3次。在首次病毒治疗后,每2天测量肿瘤长、短径,称量小鼠体重。以以下公式计算肿瘤体积大小:肿瘤体积=1/2
×
长径(mm)
×
短径(mm)2计算肿瘤体积大小。当肿瘤体积大于2000mm3时,判定小鼠死亡。如图8所示,与pbs和vv-control比较,vv-epcam-bite可以显著抑制4t1皮下移植瘤的生长,延长小鼠的生存时间。三组之间比较,小鼠体重无显著性差异,说明瘤内注射溶瘤痘苗病毒vv-epcam-bite对balb/c 小鼠无明显的毒副作用。
[0057]
实施例7:溶瘤痘苗病毒对结肠癌皮下移植瘤的治疗作用
[0058]
在本实施例中,用过表达epcam的结肠癌细胞mc38-epcam
+
细胞建立结肠癌皮下瘤模型。首先用流式细胞术鉴定了mc38-epcam
+
细胞表面epcam的表达情况,如图9所示,epcam在mc38-epcam
+
细胞表面高表达。
[0059]
然后选用6-8周龄雄性c57bl/6小鼠(spf级,购于南京大学模式动物研究所),每只小鼠皮下注射2
×
106个mc38-epcam
+
细胞。在建模大约4天后,肿瘤直径大约长至5mm左右时,将小鼠随机分为3组,分别为pbs对照组、 vv-control对照组和vv-epcam-bite治疗组。治疗剂量、治疗方式以及检测指标同实施例7。如图9所示,与pbs和对照病毒vv-control比较,vv-epcam-bite 可以显著抑制mc38-epcam
+
皮下移植瘤的生长。同时,vv-epcam-bite治疗显著延长荷瘤小鼠的生存时间。pbs、vv-control和vv-epcam-bite治疗组之间比较,小鼠体重无显著性差异,说明瘤内注射溶瘤痘苗病毒vv-epcam-bite 治疗对c57bl/6小鼠无明显的毒副作用。
[0060]
本行业的技术人员应该了解,本发明不受上述实施例的限制,上述实施例和说明书中描述的只是说明本发明的原理,在不脱离本发明精神和范围的前提下,本发明还会有各种变化和改进,本发明要求保护范围由所附的权利要求书、说明书及其等效物界定。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1