一种水稻氨基酸转运基因及其应用以及水稻育种方法
本发明涉及水稻分子育种技术领域,具体涉及一种水稻氨基酸转运基因及其应用以及水稻育种方法。
背景技术:
水稻(oryzasatival.)是我国最主要的粮食作物之一,如何提高水稻产量,增加稳定供给,是水稻育种长期以来一直致力于解决的问题。传统育种只能依靠品种或者种之间的杂交实现重组,选育出具有优良性状的品种。为了缩短育种周期,且能快速培育出高产、综合抗性强、结实率高的目标品系,逐渐产生了转基因育种和基于基因编辑技术的育种方式。转基因育种是指通过转基因的方法,导入外源的基因,达到性状改良的目标,从而培育出新品种。而转基因育种可以实现跨物种的基因交流,对目标性状改良的针对性强,提高育种效率。近年来,基因编辑技术的突飞猛进,特别是crispr/cas9技术的应用,基因敲除技术已经成为常规技术,基因敲入技术也产生了突破。因此,定向敲除不良目标基因和定向整合优良目标基因,将大幅提高水稻定向遗传改良效率。
高效利用氮素的作物对于全球粮食供应和环境可持续发展至关重要(oldroydged,leysero.aplant'sdiet,survivinginavariablenutrientenvironment.science,2020,368(6486):eaba0196.)。氮素中的氨基酸是水稻氮素营养吸收、转运、储存和利用的重要形式,能被各种氨基酸转运蛋白进行运输。有研究指出,氮转运蛋白是提高作物生产力和氮利用效率的有效靶点(tegederm,masclaux-daubressec.sourceandsinkmechanismsofnitrogentransportanduse.newphytologist.2018,217(1):35-53.)。水稻中氨基酸转运蛋白(aat)成员有85个(zhaoh,mah,yul,wangx,zhaoj.genome-widesurveyandexpressionanalysisofaminoacidtransportergenefamilyinrice(oryzasatival.).plosone,2012,7(11):e49210.),在水稻生长发育的各个时期,及各个组织部位,分别扮演不同的作用。不同氨基酸转运蛋白对水稻生长发育影响差异较大,有些可能是正向调控作用,有些可能是负向调控作用,且调控生长发育的表型也不尽相同。并且每一个氨基酸转运蛋白响应的氨基酸也不尽相同。因此,发现新的水稻氨基酸转运蛋白,并利用其特殊的功效作用进行转基因或者基因敲除育种,对于水稻适应不同种植环境及利用氨基酸类有机氮肥进一步改善水稻株型和稻米品质,以满足消费者需求具有非常重要的意义。
技术实现要素:
本发明意在提供一种水稻氨基酸转运基因,以解决提高水稻生物量以及分蘖芽生长速度的技术问题。
为达到上述目的,本发明采用如下技术方案:
一种水稻氨基酸转运基因,其蛋白的氨基酸序列如seqidno.1所示,或者其蛋白为如seqidno.1所示的氨基酸序列经取代、缺失和/或增加若干个氨基酸而获得的具有同等功能的蛋白质。
本方案的原理及优点是:发明人通过分蘖和单株产量等表型在533份自然变异的水稻中进行全基因组关联分析,再通过对大量候选序列的功能验证实验,发现氨基酸序列如seqidno.1所示的蛋白具有氨基酸转运蛋白活性,并将其蛋白命名为osaap12蛋白,将其基因命名为osaap12基因。该蛋白的功能为发明人首次发现,并将该蛋白应用于水稻的遗传育种的实践当中。在不影响osaap12蛋白活性的前提下(即不在蛋白的活性中心),本领域技术人员可对seqidno.1所示的氨基酸序列进行各种取代、添加和/或缺失若干个氨基酸获得具有同等功能的氨基酸序列。因此,osaap12蛋白还包括seqidno.1所示氨基酸序列经取代、替换和/或增加若干个氨基酸获得的具有同等活性的蛋白质。
水稻中有85个氨基酸转运蛋白成员,其中aap亚家族(osaap,氨基酸通透酶基因)也有多个成员。从目前的研究上看,已经发现的每一个osaap基因均有其独特的功能,对于水稻的生长发育有着不同的作用方式,因此,发现新的osaap基因并理清新其调控机制,对于水稻的遗传育种改良具有十分重大的意义。目前已发现,水稻氨基酸转运蛋白基因家族中氨基酸通透酶基因osaap6在种子中表达,与籽粒蛋白含量正关联,影响稻米品质(pengb,kongh,liy,wangl,zhongm,sunl,gaog,zhangq,luol,wangg,xiew,chenj,yaow,pengy,leil,lianx,xiaoj,xuc,lix,hey.osaap6functionsasanimportantregulatorofgrainproteincontentandnutritionalqualityinrice.naturecommunication,2014,5(1),4847.)。osaap1蛋白主要运输中性氨基酸,基因超表达后增加了水稻分蘖并增加水稻单株实粒数,而干扰和突变体植株则表现出相反的表型(jiyy,huangwt,wubw,fangzm,wangxl.theaminoacidtransporterosaap1mediatesgrowthandgrainyieldbyregulatingneutralaminoacidsuptakeandreallocationinoryzasativa.journalofexperimentalbotany,2020,71(16):4763-4777.)。可见,不同氨基酸转运蛋白调控生长发育的表型也不尽相同,有些影响水稻分蘖,有些影响水稻品质。同时,由于每一个氨基酸转运蛋白转运的氨基酸不尽相同,其可能受特定几种氨基酸诱导表达,并在不同浓度的某一氨基酸处理下其生长受到促进或抑制效果。这表明每一个氨基酸转运蛋白可能均与一种或多种氨基酸产生密切联系,将氨基酸转运蛋白和相应氨基酸处理联系起来可能有助于水稻选育中的应用,但这需要实验研究才能揭示其可能的内在联系。
本方案发现的osaap12蛋白与已发现的osaap蛋白在序列和功能上均存在较大差异,发明人构建了osaap12基因的超表达植株和基因敲除植株(突变体植株),并经研究发现:将水稻osaap12基因超表达植株、osaap12基因突变体植株与对照中花11放置在普通水稻培养液及分别添加了低中高浓度的某一氨基酸单一处理的水稻培养液中进行培养。与在添加了不同浓度丝氨酸的水稻培养液处理下,中花11、osaap12基因的超表达植株及突变体植株之间的分蘖芽伸长及生物量没有差异。在普通水稻培养液及添加1mm或2mm天冬酰胺、2mm或3mm苏氨酸的普通水稻营养液中,对比中花11,osaap12基因的超表达植株水稻第一分蘖芽和第二分蘖芽的伸长和生物量均受到抑制,osaap12基因突变体植株则受到促进效果。在添加0.5mm天冬酰胺或1mm苏氨酸的水稻普通营养液中,对比中花11,osaap12基因的超表达植株水稻第一分蘖芽和第二分蘖芽的伸长和生物量均受到促进,osaap12基因突变体植株则受到抑制效果。综上,在不同浓度的单一氨基酸处理下,osaap12基因的超表达植株及突变体植株在水稻生物量及分蘖芽伸长能受到不同浓度的天冬酰胺及苏氨酸处理而产生明显促进或抑制效果。提高或敲除osaap12基因表达并放置在对应的氨基酸水稻培养液中,可以促进水稻第一分蘖芽和第二分蘖芽的生长,促进了地上部分的生长,从而提高水稻单株产量。osaap12基因的超表达植株及突变体植株可以响应于特定的氨基酸(天冬酰胺及苏氨酸),可以利用上述现象,将osaap12基因的表达量改变于结合氨基酸处理结合,应用于水稻株型和产量的遗传改良的实践操作中。
进一步,一种水稻氨基酸转运基因的cdna序列如seqidno.2所示。cdna序列如seqidno.2所示的基因可以编码获得如seqidno.1所示的蛋白。在天冬酰胺或苏氨酸低浓度水平的环境下,过表达该基因,可以提高水稻生物量和分蘖芽长度;在天冬酰胺或苏氨酸高浓度水平或者不含氨基酸的环境下,敲除该基因,可以提高水稻生物量和分蘖芽长度。由针对该基因的转基因或者基因编辑技术获得的水稻植株可以适应于不同生长环境,为增加水稻品种的品质以及多样性创造了基础。
此外,应理解,考虑到密码子的简并性以及不同物种密码子的偏爱性,本领域技术人员可以根据需要使用适合特定物种表达的密码子。
本发明还采用如下技术方案:一种水稻氨基酸转运基因在水稻育种上的应用。
本方案的原理及优点是:基于本发明发现新基因osaap12基因,其可用于水稻选育中,通过水稻选育可获得促进水稻第一分蘖芽和第二分蘖芽的伸长以及生物量的增加的效果。构建osaap12基因的超表达载体提高osaap12基因表达并结合0.5mm天冬酰胺或1mm苏氨酸的水稻培养液中进行培养,或基因敲除技术降低osaap12基因的表达并结合添加1mm-2mm天冬酰胺、2mm-3mm苏氨酸的任一普通水稻营养液进行培养,可以促进水稻生物量的增加,从而达到提高水稻产量的目的。
进一步,敲除水稻的cdna序列如seqidno.2所示的基因。通过构建osaap12基因的敲除载体,将敲除载体导入中花11中,得到osaap12基因的基因敲除植株,在不含氨基酸的普通水稻培养液中,基因敲除植株的第一分蘖芽和第二分蘖芽的长度增加,生物量增加,促进了地上部分的生长。
进一步,在含有1mm-2mm天冬酰胺的培养环境中,对水稻的cdna序列如seqidno.2所示的基因进行基因敲除处理。通过敲除osaap12基因并放置在添加1mm-2mm天冬酰胺的水稻营养液中,可促进水稻第一分蘖芽和第二分蘖芽的伸长以及生物量的增加。
进一步,在含有2mm-3mm苏氨酸的培养环境中,对水稻的cdna序列如seqidno.2所示的基因进行基因敲除处理。通过敲除osaap12基因并放置在添加2mm-3mm苏氨酸的水稻营养液中,可促进水稻第一分蘖芽和第二分蘖芽的伸长以及生物量的增加。
进一步,在含有0.5mm天冬酰胺的培养环境中,将水稻的cdna序列如seqidno.2所示的基因的表达水平上调。将osaap12基因超表达植株放置在添加0.5mm天冬酰胺的水稻培养液中,其第一分蘖芽和第二分蘖芽的伸长受到促进,其生物量增加。
进一步,在含有1mm苏氨酸的培养环境中,将水稻的cdna序列如seqidno.2所示的基因的表达水平上调。将osaap12基因超表达植株放置在添加1mm苏氨酸的水稻培养液中,其第一分蘖芽和第二分蘖芽的伸长受到促进,其生物量增加。
进一步,敲除或者过表达cdna序列如seqidno.2所示的基因用于促进第一分蘖芽和第二分蘖芽的伸长以及水稻生物量的增加。调节osaap12基因的表达量以及调节水稻植株的培养环境中的氨基酸含量,可以促进第一分蘖芽和第二分蘖芽的伸长以及水稻生物量的增加。
本发明还采用如下技术方案:一种水稻育种的方法,当培养环境中不含氨基酸或者含有1mm-2mm天冬酰胺或者含有2mm-3mm苏氨酸时,对水稻的cdna序列如seqidno.2所示的基因进行基因敲除处理;当培养环境中含有0.5mm天冬酰胺或者1mm苏氨酸时,将水稻的cdna序列如seqidno.2所示的基因的表达水平上调。
本方案的原理及优点是:通过构建osaap12基因超表达载体,将超表达载体导入中花11中,得到osaap12基因超表达植株,将其放置在添加0.5mm天冬酰胺或1mm苏氨酸的水稻培养液中,其第一分蘖芽和第二分蘖芽的伸长受到促进,增加生物量。通过构建osaap12基因的敲除载体,将敲除载体导入中花11中,得到osaap12基因的基因敲除植株,并放置在添加1mm-2mm天冬酰胺、2mm-3mm苏氨酸的任一普通水稻营养液或者不含有氨基酸的普通水稻培养基中,能促进其第一分蘖芽和第二分蘖芽的伸长,促进了地上部分的生长。
综上所述,本发明的技术方案的优点和有益效果如下:
(1)本发明可以通过基因敲除技术降低osaap12基因的表达,并放置在添加1mm-2mm天冬酰胺或2mm-3mm苏氨酸的普通水稻营养液或者无氨基酸添加的水稻营养液中,可以促进osaap12基因突变体第一分蘖芽和第二分蘖芽的伸长以及生物量的增加,因此,可以通过结合基因编辑技术和分子育种进行植物的品种改良。
(2)本发明发现可以通过构建osaap12基因超表达载体提高osaap12基因表达并结合0.5mm天冬酰胺或1mm苏氨酸的水稻培养液中进行培养,可以促进水稻第一分蘖芽和第二分蘖芽的伸长,促进生物量的增加,因此,可以通过在水稻培养液中施加0.5mm天冬酰胺或1mm苏氨酸促进osaap12基因超表达水稻产量增加。
(3)osaap12基因的成功克隆,证实了氨基酸转运基因不仅在植物氨基酸含量改变和品质调控中发挥作用,还在不同有机氮下的植株生长和分蘖芽伸长中有重要的差异调控作用,可以丰富氨基酸转运蛋白的认识,对植株株型和单株产量的遗传改良有极大的推动作用。
附图说明
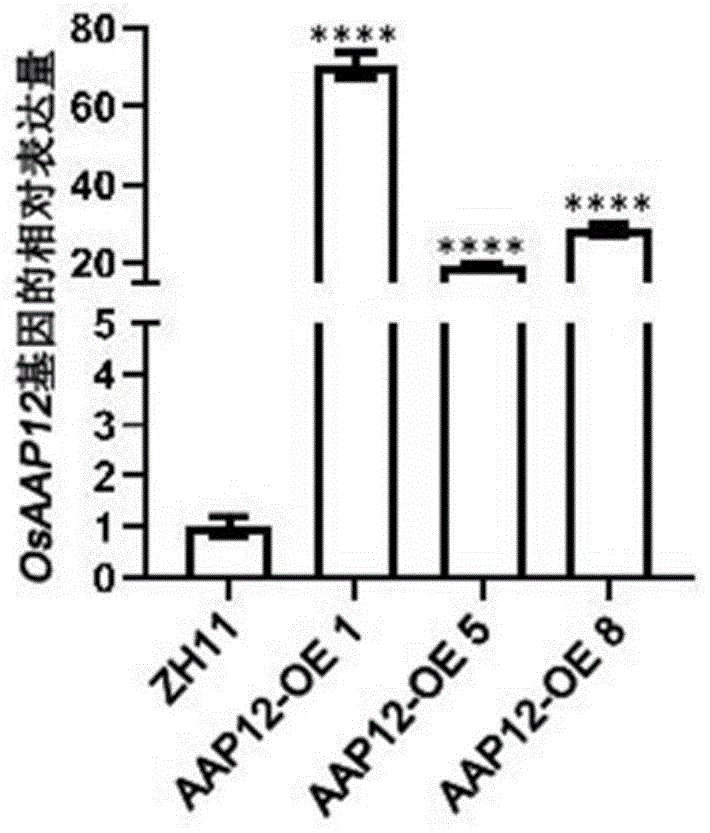
图1为本发明实施例1的三个超表达株系表达量鉴定结果。
图2为本发明实施例2的三个基因敲除株系的碱基缺失情况。
图3为本发明实验例1的不同株系的水稻在不含氨基酸的水稻营养液培养下的单株水稻表型图。
图4为本发明实验例1的不同株系的水稻在含0.5mm天冬酰胺的水稻营养液培养下的单株水稻表型图。
图5为本发明实验例1的不同株系的水稻在含1mm天冬酰胺的水稻营养液培养下的单株水稻表型图。
图6为本发明实验例1的不同株系的水稻在含2mm天冬酰胺的水稻营养液培养下的单株水稻表型图。
图7为本发明实验例1的不同株系的水稻在含1mm苏氨酸的水稻营养液培养下的单株水稻表型图。
图8为本发明实验例1的不同株系的水稻在含2mm苏氨酸的水稻营养液培养下的单株水稻表型图。
图9为本发明实验例1的不同株系的水稻在含3mm苏氨酸的水稻营养液培养下的单株水稻表型图。
图10为本发明实验例1的不同株系的水稻在含1mm丝氨酸的水稻营养液培养下的单株水稻表型图。
图11为本发明实验例1的不同株系的水稻在含2mm丝氨酸的水稻营养液培养下的单株水稻表型图。
图12为本发明实验例1的不同株系的水稻在普通水稻营养液培养下的单株水稻小苗第一分蘖芽和第二分蘖芽表型图。
图13为本发明实验例1的不同株系的水稻在含0.5mm天冬酰胺的水稻营养液培养下的单株水稻小苗第一分蘖芽和第二分蘖芽表型图。
图14为本发明实验例1的不同株系的水稻在含1mm天冬酰胺的水稻营养液培养下的单株水稻小苗第一分蘖芽和第二分蘖芽表型图。
图15为本发明实验例1的不同株系的水稻在含2mm天冬酰胺的水稻营养液培养下的单株水稻小苗第一分蘖芽和第二分蘖芽表型图。
图16为本发明实验例1的不同株系的水稻在含1mm苏氨酸的水稻营养液培养下的单株水稻小苗第一分蘖芽和第二分蘖芽表型图。
图17为本发明实验例1的不同株系的水稻在含2mm苏氨酸的水稻营养液培养下的单株水稻小苗第一分蘖芽和第二分蘖芽表型图。
图18为本发明实验例1的不同株系的水稻在含3mm苏氨酸的水稻营养液培养下的单株水稻小苗第一分蘖芽和第二分蘖芽表型图。
图19为本发明实验例1的不同株系的水稻在含1mm丝氨酸的水稻营养液培养下的单株水稻小苗第一分蘖芽和第二分蘖芽表型图。
图20为本发明实验例1的不同株系的水稻在含2mm丝氨酸的水稻营养液培养下的单株水稻小苗第一分蘖芽和第二分蘖芽表型图。
图21为本发明实验例1的不同株系的水稻在普通水稻营养液培养下的水稻小苗第一分蘖芽和第二分蘖芽长度柱状图(mean±sd,n=30)。
图22为本发明实验例1的不同株系的水稻在含有不同浓度的不同单一氨基酸的水稻营养液培养下的水稻小苗第一分蘖芽和第二分蘖芽长度柱状图(mean±sd,n=30)。
图23为本发明实验例1的不同株系的水稻在普通水稻营养液培养下的水稻小苗鲜重柱状图(mean±sd,n=30)。
图24为本发明实验例1的不同株系的水稻在含有不同浓度的不同单一氨基酸的水稻小苗鲜重柱状图(mean±sd,n=30)。
具体实施方式
下面结合实施例对本发明做进一步详细的说明,但本发明的实施方式不限于此。若未特别指明,下述实施例所用的技术手段为本领域技术人员所熟知的常规手段;所用的实验方法均为常规方法,并可按照已描述的重组技术(参见分子克隆,实验室手册,第2版,冷泉港实验室出版社,冷泉港,纽约;maxetal,arobustcrispr/cas9systemforconvenient,high-efficiencymultiplexgenomeeditinginmonocotanddicotplants.molplant.2015,8(8):1274-1284.)完成;所用的材料、试剂等,均可从商业途径得到。
实施例1:osaap12基因超表达植株的构建
提取水稻中花11的rna,并将其反转录成cdna,利用引物对f1和r1,通过pcr扩增osaap12基因的cdna(osaap12基因的蛋白序列参见seqidno.1,osaap12基因的cdna序列参见seqidno.2)。引物对f1和r1,分别为:
f1:5'-atggtaccatggttcagatcgagccgctcgag-3'(seqidno.3);
r1:5'-atggatcctccactgaagggacggaatttcat-3'(seqidno.4)。
通过pcr扩增osaap12基因的cdna后,通过(kpni和bamhi连入pcambia-1306载体(pcambia-1306载体购自cambia公司),构建出osaap12基因的超表达载体osaap12-p1306。采用农杆菌eha105介导的遗传转化方法,将超表达载体导入正常水稻品种中花11中。
将得到的所有转基因小苗移栽于带泥土的筐中,定期浇水,施肥,待小苗长高约10cm时,种于大田中,待苗长大后,提取基因组dna通过pcr对转基因植株进行检测,检测引物对为:
f2:5'-cttctacacagccatcggtccag-3'(seqidno.5);
r2:5'-cggaagtgcttgacattggggag-3'(seqidno.6)。
若扩增出700bp潮霉素抗性基因pcr的产物片段,证明载体质粒成功发挥作用,以潮霉素筛选成功获得osaap12的超表达转基因苗。阳性植株单株收种并种植,直至t2代鉴定出纯合的转基因植株,即得到osaap12基因超表达植株,获得osaap12基因超表达植株三个株系(aap12-oe1、aap12-oe5和aap12-oe8)。
取osaap12基因超表达植株叶片,提取rna并将其反转录成cdna,通过实时荧光定量pcr检测osaap12基因在超表达植株的表达量,超表达植株中osaap12基因的表达量比对照中花11提高,将对照的表达量定为1,超表达植株株系的表达量最高能提高70倍左右,三个株系表达量鉴定结果如图1所示。实时荧光定量pcr所用引物对:
f3:5'-ccaacgtcaccgccacgcaga-3'(seqidno.7);
r3:5'-cgccctgagcccgatccaca-3'(seqidno.8)。
实施例2:osaap12基因突变体植株的构建
利用单靶标序列,构建出osaap12基因的基因敲除载体osaap12-c(方法参考maxetal,arobustcrispr/cas9systemforconvenient,high-efficiencymultiplexgenomeeditinginmonocotanddicotplants.molplant.2015,8(8):1274-1284.)。采用农杆菌eha105介导的遗传转化方法,将基因敲除表达载体导入正常粳稻品种中花11中。突变体植株在t0代时进行测序,确定基因已经敲除3个株系,3各株系的碱基序列改变分别为缺失tgc和g、插入t、缺失gtgc(如图2所示),继续独立繁种到t1代,即得到osaap12基因的独立的突变体植株株系(aap12-c77、aap12-c78和aap12-c79)。单靶标序列的核苷酸序列为:5'-tgttctacgcgctgtgcgggtgg-3'(seqidno.9)。
实验例1:不同浓度氨基酸对水稻各株系的影响
将中花11、osaap12基因超表达植株三个株系和突变体植株三个株系种子在培养皿上用蒸馏水浸种3天并培养7天后,转入水稻培养液(含有或者不含有氨基酸),分别培养35天,观察水稻表型,统计水稻分蘖芽数目。培养液的配制条件:配制水稻培养液(参照国际水稻研究所水稻营养液配方),并在水稻培养液的基础上分别添加0.5mm、1mm、2mm天冬酰胺,1mm、2mm、3mm苏氨酸,1mm、2mm丝氨酸。水稻培养液配方为nh4no3:11.43g,nah2po4·2h2o:5.04g,k2so4:8.93g,cacl2:3.164g,mgso4·7h2o:40.5g,微量元素液:100ml,铁盐螯合液:160ml,加清水80l,ph调为5.4。氨基酸处理即为在上述培养液的基础上个根据设置的氨基酸浓度加入相对应的氨基酸质量。由于天冬酰胺和苏氨酸难溶于水,先置于稀盐酸中完全溶解再施加到水稻培养液中。除了本方案的水稻培养液外,选用其他现有技术中的用于水稻培养的培养液也能达到相同效果。
随机选取不同处理的水稻小苗各一株放置在黑布上进行拍照,结果如图3-图11所示,其中,在图中,植株从左往右依次为:对照中花11(zh11)、超表达植株3个株系(oe1、oe5和oe8)、突变体植株3个株系(c77、c78和c79)。在未添加氨基酸的普通培养基中,超表达植株根长较短,整体植株比较矮小,突变体植株根长较长,整体植株比较高大(图3)。在0.5mm天冬酰胺或1mm苏氨酸水稻营养液处理下,其超表达植株根长较长,整体植株比较高大,而突变体植株则根长较短,整体植株比较矮小(图4和图7)。
再随机选取不同处理的水稻小苗各一株,用体式显微镜进行分蘖芽拍照,观察水稻小苗的第一分蘖芽和第二分蘖芽,实验结果参见图12-图20,其中,在图中,植株从左往右依次为:对照中花11(zh11)、超表达植株3个株系(oe1、oe5和oe8)、突变体植株3个株系(c77、c78和c79),右侧标尺为0.5cm。超表达植株第一分蘖芽和第二分蘖芽均短于中花11,突变体植株第一分蘖芽和第二分蘖芽均长于中花11(图12)。在0.5mm天冬酰胺或1mm苏氨酸水稻营养液处理下的超表达植株第一分蘖芽和第二分蘖芽长于中花11,突变体植株则相反(图13和图16)。
统计各株系的第一分蘖芽和第二分蘖芽平均长度,实验结果参见图21和图22。超表达植株第一分蘖芽和第二分蘖芽平均长度短于中花11,突变体植株第一分蘖芽和第二分蘖芽平均长度长于中花11(图21)。在0.5mm天冬酰胺或1mm苏氨酸水稻营养液处理下,超表达植株第一分蘖芽和第二分蘖芽平均长度长于中花11,突变体植株第一分蘖芽和第二分蘖芽平均长度短于中花11(图22)。而在丝氨酸存在的情况下,超表达植株、突变体植株和野生型植株的分蘖芽的长度不存在显著差异。继而统计各株系的平均鲜重,发现超表达植株的平均鲜重低于中花11,而突变体植株则相反(图23)。在0.5mm天冬酰胺或1mm苏氨酸水稻营养液处理下,超表达植株的平均鲜重高于中花11,而突变体植株的平均鲜重低于中花11(图24)。而在丝氨酸存在的情况下,超表达植株、突变体植株和野生型植株的鲜重不存在显著差异。在图21-24中,数据采用spss软件进行变量分析(anova),使用duncan’s在0.05水平上进行差异显著性分析,小写字母表示同一个表型中zh11、超表达三个株系、突变体三个株系这7种各材料之间相比具有差异显著。
上述结果表明,在普通水稻营养液培养条件下,osaap12基因的超表达植株第一分蘖芽和第二分蘖芽伸长受到抑制,水稻生物量偏低;osaap12基因的突变体植株第一分蘖芽和第二分蘖芽伸长受到促进,水稻生物量偏高。
在分别添加了低中高浓度的某一氨基酸单一处理的水稻培养液中:相比在施加低或高浓度丝氨酸的水稻营养液处理下,osaap12基因超表达植株、突变体植株和对照中花11在各项生理指标无明显差异(图10、11、19、20、22、24);在0.5mm天冬酰胺或1mm苏氨酸的水稻培养液中,对比野生型中花11,osaap12基因超表达植株水稻第一分蘖芽和第二分蘖芽的伸长,生物量的增加受到促进,而突变体植株则受到抑制;而在添加了1mm、2mm天冬酰胺,2mm、3mm苏氨酸的水稻培养液中,对比野生型中花11,osaap12基因突变体植株水稻第一分蘖芽和第二分蘖芽的伸长,生物量的增加均受到促进,而超表达植株则受到抑制。
以上所述的仅是本发明的实施例,方案中公知的具体技术方案和/或特性等常识在此未作过多描述。应当指出,对于本领域的技术人员来说,在不脱离本发明技术方案的前提下,还可以作出若干变形和改进,这些也应该视为本发明的保护范围,这些都不会影响本发明实施的效果和专利的实用性。本申请要求的保护范围应当以其权利要求的内容为准,说明书中的具体实施方式等记载可以用于解释权利要求的内容。
sequencelisting
<110>贵州大学
<120>一种水稻氨基酸转运基因及其应用以及水稻育种方法
<130>2021.04.08
<160>9
<170>patentinversion3.5
<210>1
<211>352
<212>prt
<213>oryzasatival.
<400>1
metvalilevalseralaglyasnalaaspseralaaspaspaspgly
151015
argargargthrglythrtrpthralaseralahisileilethrala
202530
valileglyserglyvalsertrpalathralaglytrpvalglygly
354045
alavalmetvalvalglyglyvalthrtyrthralathralacystyr
505560
argthrglyaspthrglyalaargasntyrthrtyrileglyalaval
65707580
argalaileglyglyalaasnalalyscysglyileiletyralaasn
859095
valglythralavalglytyrthrilealaalaserilesermetala
100105110
ilelysargalaglycyshisalaasnglyhisasnvalcyshisile
115120125
serserthrtyrmetileglyalailevalserileasphisiletrp
130135140
trpserilevalalaalavalmetserthrtyrserglyvalglygly
145150155160
glyilealathrvalalaaspglyglyargglythrilealaglyval
165170175
thrasnvalthralathrlysalatrpargseralaglyasnileala
180185190
alaalaserasnvaltyrthrileaspthrilelysalaseralalys
195200205
valmetlysalaserserilevalalathrservaltyralacysgly
210215220
trpmetglytyralaalaglyasnalaalaaspasnthrglyglytrp
225230235240
valaspalaalaasnvalalailealavalhisileglyalatyrval
245250255
tyrcysvalalavalarglysalaserargargtrpaspserglyval
260265270
asnserargvaltrpalaileseralaargalatrpargservalval
275280285
cysthrthrvalvalalametalaglyvalilevalglyglyalaile
290295300
sertrpthrvaltyrthrmettyrilealaargglyvalargarggly
305310315320
seralatrpileglyargalaalavalalaglyvalvalseralaala
325330335
alathrthrglyalavalalaasnvalglyaspmetlysargsergly
340345350
<210>2
<211>1407
<212>dna
<213>oryzasatival.
<400>2
atggttcagatcgagccgctcgaggtgtcactggaggccggcaaccaggcggactcggcg60
ttgctcgacgacgacgggcggccgcgccgcaccggcacgttctggacggcgagcgcgcac120
atcatcaccgccgtgatcgggtccggggtgctgtcgctgccgtgggcgacggcgcagctc180
gggtgggtcggcgggcccgccgtgatggtggtgttcggcggcgtcacctacttcaccgcc240
acgctccaggccgagtgctaccgcaccggcgacgaggagaccggcgcccgcaactacacc300
tacatcggcgccgtccgcgccatcctcggcggcgccaacgccaagctctgcggcatcatc360
cagtacgccaacctcgtcggcaccgccgtcggctacaccatcgccgcctccatcagcatg420
caggccatcaagagggcgggctgcttccacgccaatggccacaacgtcccgtgccacatc480
tcgagcaccccgtacatgctcatcttcggcgccttcgagatcgtcttctcgcagatcccc540
gacttccacgagatctggtggctgtccatcgtcgccgccgtcatgtccttcacctactcc600
ggcgtcggcctcggcctcggcatcgcccagaccgtcgccgacggcgggttccgcggcacg660
atcgccggcgtcaccaacgtcaccgccacgcagaaggcgtggcggtcgctgcaggcgctg720
ggcaacatcgccttcgcgttcgccttctccaacgtgtacaccgagatccaggacacgatc780
aaggcgccgccgccgtcggaggcgaaggtgatgaagcaggcgtcgctgctgagcatcgtg840
gcgacgtcggtgttctacgcgctgtgcgggtggatgggctacgcggcgttcggcaacgcg900
gcgccggacaacctcctcaccggattcggcttcttcgagccgttctggctcgtcgacgcc960
gccaacgtcgccatcgccgtgcacctcatcggcgcctaccaggtgtactgccagccggtc1020
ttcgccttcgtcgagcgcaaggcgtcgcgccggtggcccgacagcggcttcgtcaacagc1080
gagctccgggtgtggcccttcgccatcagcgcgttccggctggcgtggcgctcggtgttc1140
gtctgcttcaccaccgtggtcgccatggcgctgcccttcttcggcgtcatcgtcggcctc1200
ctcggtgccatctccttctggccgctcaccgtctacctccccacggagatgtacatcgcg1260
cagcgcggcgtgcggcgcggcagcgcactgtggatcgggctcagggcgctcgccgtcgcc1320
ggcttcgtcgtgtcggccgccgccaccacgggcgccgtcgccaacttcgtcggcgacttc1380
atgaaattccgtcccttcagtggatag1407
<210>3
<211>32
<212>dna
<213>人工序列
<400>3
atggtaccatggttcagatcgagccgctcgag32
<210>4
<211>32
<212>dna
<213>人工序列
<400>4
atggatcctccactgaagggacggaatttcat32
<210>5
<211>23
<212>dna
<213>人工序列
<400>5
cttctacacagccatcggtccag23
<210>6
<211>23
<212>dna
<213>人工序列
<400>6
cggaagtgcttgacattggggag23
<210>7
<211>21
<212>dna
<213>人工序列
<400>7
ccaacgtcaccgccacgcaga21
<210>8
<211>20
<212>dna
<213>人工序列
<400>8
cgccctgagcccgatccaca20
<210>9
<211>23
<212>dna
<213>人工序列
<400>9
tgttctacgcgctgtgcgggtgg23
- 还没有人留言评论。精彩留言会获得点赞!