治疗或预防新冠病毒的靶点的制作方法
1.本发明涉及生物医药领域,具体涉及一种治疗或预防新冠病毒的靶点。
背景技术:
2.由新冠病毒(sars
‑
cov
‑
2) 引起的 covid
‑
19大流行已造成全球公共卫生危机。为了对抗 sars
‑
cov
‑
2,大多数治疗策略都集中在针对病毒棘突(spike, s)蛋白的疫苗或抗病毒药物上。然而,rna 病毒的病毒进化是由高突变率驱动的,可使病毒逃避宿主免疫攻击。尽管人类为对抗 sars
‑
cov
‑
2 感染做出了许多努力,许多国家目前仍面临新冠病毒突变株引起的新一波疫情,发病率依然很高。因此,针对s蛋白的抗病毒药物或疫苗的开发可能会遇到耐药突变的问题。实际情况也如此,针对 b.1.351 突变株测试的三种疫苗(janssen、novavax 和 astrazeneca)对预防症状性感染的效果降低,其有效性估计值分别为 57%、49% 和统计学上甚至不显著的百分比。对于mrna疫苗 bnt162b2 (pfizer) 和 mrna
‑
1273 (moderna),接种者中针对b.1.351 突变株的血清中和活性降低了 6.5 倍和 8.6 倍。因此,迫切需要开发针对sars
‑
cov
‑
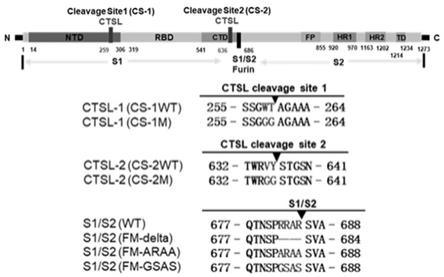
2的广谱治疗措施。
3.在病毒感染期间,s蛋白需要被宿主细胞(例如人体)蛋白酶激活,例如弗林蛋白酶、tmprss2 和组织蛋白酶 l (ctsl)。ctsl是人体内产生的溶酶体半胱氨酸蛋白酶的成员,其主要功能是对病原体产生的抗原进行蛋白水解。
4.据报道,在 sars
‑
cov
‑
2感染患者中,血液样本中 ctsl 的水平与 covid
‑
19 的严重程度和病程高度相关。ctsl通过切割 s 和增强病毒进入细胞来促进 sars
‑
cov
‑
2感染。
技术实现要素:
5.第一方面,本发明提供一种蛋白片段,所述蛋白片段是如下a1)
‑
a4)任意所述的蛋白片段:a1)氨基酸酸序列如seq id no.1所示;a2)氨基酸序列如seq id no.1第12位
‑
34位所示;a3)将a1)的蛋白片段的第18、19、28和29位中的任意一个或几个氨基酸残基经过一个或几个氨基酸残基的取代和/或缺失和/或添加得到的与a1)所示的蛋白片段具有90%以上的同一性的蛋白片段;a4)氨基酸酸序列如seq id no.2所示;a5)氨基酸序列如seq id no.2第32
‑
41位所示;a6)将a4)的蛋白片段的第35和36位中的任意1个或2个氨基酸残基经过一个或几个氨基酸残基的取代和/或缺失和/或添加得到的与a4)所示的蛋白片段具有90%以上的同一性的蛋白片段。
6.第二方面,使冠状病毒的s蛋白片段1和/或片段2不被蛋白酶切割或被蛋白酶切割效率降低的物质在制备的产品中的应用;所述冠状病毒为sars
‑
cov
‑
2或其突变株;所述产品的功能为如下任意一种或几种:
(1)抑制细胞融合;(2)预防sars
‑
cov
‑
2或其突变株感染;(3)治疗sars
‑
cov
‑
2或其突变株感染引起的疾病;所述s蛋白片段1为如下任意一种:a1)氨基酸酸序列如seq id no.1所示;a2)氨基酸序列如seq id no.1第12位
‑
34位所示;a3)将a1)的蛋白片段的第18、19、28和29位中的任意一个或几个氨基酸残基经过一个或几个氨基酸残基的取代和/或缺失和/或添加得到的与a1)所示的蛋白片段具有90%以上的同一性的蛋白片段;所述s蛋白片段2为如下任意一种:a4)氨基酸酸序列如seq id no.2所示;a5)氨基酸序列如seq id no.2第32
‑
41位所示;a6)将a4)的蛋白片段的第35和36位中的任意1个或2个氨基酸残基经过一个或几个氨基酸残基的取代和/或缺失和/或添加得到的与a4)所示的蛋白片段具有90%以上的同一性的蛋白片段。
7.可选的,蛋白酶为组织蛋白酶。
8.可选的,所述产品为药物、疫苗、中和抗体。
9.第三方面,本发明提供使冠状病毒的s蛋白片段1和/或片段2发生突变的物质在制备产品中的应用;所述冠状病毒为sars
‑
cov
‑
2或其突变株;所述产品的功能为如下任意一种或几种:(1)抑制细胞融合;(2)预防sars
‑
cov
‑
2或其突变株感染;(3)治疗sars
‑
cov
‑
2或其突变株感染引起的疾病;所述s蛋白片段1为如下任意一种:a1)氨基酸酸序列如seq id no.1所示;a2)氨基酸序列如seq id no.1第12位
‑
34位所示;a3)将a1)的蛋白片段的第18、19、28和29位中的任意一个或几个氨基酸残基经过一个或几个氨基酸残基的取代和/或缺失和/或添加得到的与a1)所示的蛋白片段具有90%以上的同一性的蛋白片段;所述s蛋白片段2为如下任意一种:a4)氨基酸酸序列如seq id no.2所示;a5)氨基酸序列如seq id no.2第32
‑
41位所示;a6)将a4)的蛋白片段的第35和36位中的任意1个或2个氨基酸残基经过一个或几个氨基酸残基的取代和/或缺失和/或添加得到的与a4)所示的蛋白片段具有90%以上的同一性的蛋白片段。
10.第四方面,本发明提供一种产品,其活性成分为使冠状病毒的s蛋白片段1和/或片段2不被蛋白酶切割或被蛋白酶切割效率降低的物质;所述产品的功能为如下任意一种或几种:(1)抑制细胞融合;
(2)预防sars
‑
cov
‑
2或其突变株感染;(3)治疗sars
‑
cov
‑
2或其突变株感染引起的疾病;所述s蛋白片段1为如下任意一种:a1)氨基酸酸序列如seq id no.1所示;a2)氨基酸序列如seq id no.1第12位
‑
34位所示;a3)将a1)的蛋白片段的第18、19、28和29位中的任意一个或几个氨基酸残基经过一个或几个氨基酸残基的取代和/或缺失和/或添加得到的与a1)所示的蛋白片段具有90%以上的同一性的蛋白片段;所述s蛋白片段2为如下任意一种:a4)氨基酸酸序列如seq id no.2所示;a5)氨基酸序列如seq id no.2第32
‑
41位所示;a6)将a4)的蛋白片段的第35和36位中的任意1个或2个氨基酸残基经过一个或几个氨基酸残基的取代和/或缺失和/或添加得到的与a4)所示的蛋白片段具有90%以上的同一性的蛋白片段。
11.可选的,蛋白酶为组织蛋白酶。
12.可选的,组织蛋白酶抑制剂为组织蛋白酶l抑制剂。
13.可选的,所述组织蛋白酶l抑制剂为z
‑
fy
‑
cho、oxocarbazate、mdl
‑
28170、k777、cathepsin inhibitor1和/或e64d。
14.第五方面:本发明提供冠状病毒的s蛋白片段1和/或片段2作为药物靶点在筛选产品中的应用;所述冠状病毒为sars
‑
cov
‑
2或其突变株;所述s蛋白片段1为如下任意一种:a1)氨基酸酸序列如seq id no.1所示;a2)氨基酸序列如seq id no.1第12位
‑
34位所示;a3)将a1)的蛋白片段的第18、19、28和29位中的任意一个或几个氨基酸残基经过一个或几个氨基酸残基的取代和/或缺失和/或添加得到的与a1)所示的蛋白片段具有90%以上的同一性的蛋白片段;所述s蛋白片段2为如下任意一种:a4)氨基酸酸序列如seq id no.2所示;a5)氨基酸序列如seq id no.2第32
‑
41位所示;a6)将a4)的蛋白片段的第35和36位中的任意1个或2个氨基酸残基经过一个或几个氨基酸残基的取代和/或缺失和/或添加得到的与a4)所示的蛋白片段具有90%以上的同一性的蛋白片段;所述产品的功能是如下任意一种或几种:1)使冠状病毒的s蛋白片段1和/或片段2不被蛋白酶切割或被蛋白酶切割效率降低;2)抑制细胞融合;3)预防sars
‑
cov
‑
2或其突变株感染;4)治疗sars
‑
cov
‑
2或其突变株的感染引起的疾病。
15.第六方面,本发明提供冠状病毒的s蛋白片段1和/或s蛋白片段2在制备预防和/或
治疗冠状病毒引起的疾病的产品中的应用;所述冠状病毒为sars
‑
cov
‑
2或其突变株;所述s蛋白片段1为如下任意一种:a1)氨基酸酸序列如seq id no.1所示;a2)氨基酸序列如seq id no.1第12位
‑
34位所示;a3)将a1)的蛋白片段的第18、19、28和29位中的任意一个或几个氨基酸残基经过一个或几个氨基酸残基的取代和/或缺失和/或添加得到的与a1)所示的蛋白片段具有90%以上的同一性的蛋白片段;所述s蛋白片段2为如下任意一种:a4)氨基酸酸序列如seq id no.2所示;a5)氨基酸序列如seq id no.2第32
‑
41位所示;a6)将a4)的蛋白片段的第35和36位中的任意1个或2个氨基酸残基经过一个或几个氨基酸残基的取代和/或缺失和/或添加得到的与a4)所示的蛋白片段具有90%以上的同一性的蛋白片段。
16.所述产品为药物。
17.所述sars
‑
cov
‑
2或其突变株引起的疾病包括呼吸道系统疾病、消化道系统疾病或神经系统疾病。
18.可选的,所述sars
‑
cov
‑
2或其突变株引起的疾病包括但不限于感冒、发烧、额窦炎、中耳炎、咽炎、慢性支气管炎、肺炎、胸腔积液、呼吸综合征、急性肠胃炎、心肺疾病、免疫力低下、重复感染、肺损伤或器官衰竭;可选的,所述sars
‑
cov
‑
2引起的疾病为新型冠状病毒肺炎(covid
‑
19);可选的,所述sars
‑
cov
‑
2引起的疾病或感染为呼吸系统疾病;所述呼吸系统疾病为单纯性感染、轻症肺炎、重症肺炎、急性呼吸道感染、严重急性呼吸道感染、低氧性呼吸衰竭、急性呼吸窘迫综合症、脓毒症或脓毒症休克中的一种或几种;可选的,所述单纯性感染为发热、咳嗽、咽痛、鼻塞、乏力、头痛、肌肉疼痛或不适中的一种或几种;可选的,所述轻症肺炎为咳嗽、呼吸困难和/或呼吸急促;可选的,所述重症肺炎为呼吸频率增加,严重的呼吸衰竭或呼吸困难,中心型发绀、嗜睡、意识不清或惊厥、抽气;可选的,所述急性呼吸窘迫综合症为肺水肿;可选的,所述脓毒症为器官功能障碍。
19.核苷酸和氨基酸序列表计算机可读载体中seq id no.1用三联体表示ginitrfqtlclalhrsyltpgdsssgwtagaaayyvgylqprtfllk,核苷酸和氨基酸序列表计算机可读载体中seq id no.2用三联体表示gtntsnqvavlyqdvnctevpvaihadqltptwrvystgsnvfqtragcl。
20.本发明技术方案,具有如下优点:1、本发明在 sars
‑
cov
‑
2的 s蛋白中确定了 ctsl 的三个新的特异性切割位点,它们在所有流行的 sars
‑
cov
‑
2 突变株中都高度保守。结合功能实验,证明了 sars
‑
cov
‑
2的s蛋白上ctsl的新切割位点为克服病毒突变的药物开发提供了有希望的治疗靶点。
21.2、本发明通过实验证明ctsl抑制剂可预防及治疗sars
‑
cov
‑
2的感染。
附图说明
22.为了更清楚地说明本发明具体实施方式或现有技术中的技术方案,下面将对具体实施方式或现有技术描述中所需要使用的附图作简单地介绍,显而易见地,下面描述中的附图是本发明的一些实施方式,对于本领域普通技术人员来讲,在不付出创造性劳动的前提下,还可以根据这些附图获得其他的附图。
23.图1a是本发明实施例1中sars
‑
cov
‑
2 s蛋白的示意图,其中突出显示了功能域和切割位点,ctsl在ctsl切割位点1(cs
‑
1)和ctsl切割位点2(cs
‑
2)处切割, furin(弗林蛋白酶)在 s1/s2 位点切割;ntd,n端域;rbd,受体结合域;ctd,c 端域;fp,融合肽;hr1,七肽重复序列1;hr2,七肽重复序列2;td,跨膜结构域;图1b是本发明实施例1中sars
‑
2 s蛋白胞外段的整体结构(pdb编号:6vxx);图1c是本发明实施例1中ctsl切割s蛋白的sds
‑
page电泳图,纯化的sars
‑
cov
‑
2的s蛋白胞外域与不同浓度的 ctsl (2
‑
8 μg/ml) 一起孵育,切割后条带 1 (60 kda) 和带 2 (100 kda)的n 端测序结果展示在图下;图1d是本发明实施例1中 6
‑
gly突变的sars
‑
cov
‑
2 s蛋白(即为mutant,获得方法为:野生型s 蛋白的cs
‑
1a 和 cs
‑
1b 位点、cs
‑
2 位点的p1残基、p2残基共6个氨基酸残基均突变为gly)不能被 ctsl 切割成 60 kda(带 1)和 100 kda(带 2)片段, 野生型s 蛋白 (1 μg) 和6
‑
gly突变的sars
‑
cov
‑
2 s蛋白 (500 ng) 与 ctsl (s 蛋白与ctsl 的混合物中,ctsl 终浓度为8 μg/ml) 一起孵育,所有样品均进行sds
‑
page后银染检测条带;野生型s 蛋白为实施例1的第“一”中sars
‑
cov
‑
2 的s蛋白胞外段在 k986 和 v987 处用脯氨酸取代,弗林蛋白酶裂解位点(s1/s2 位点,r682 至 r685)处用“gsas”取代以增强的稳定性,所得s蛋白突变体。
24.图1e是本发明实施例1中 cs
‑
1与cs
‑
2切点附近的多肽可被ctsl切割,cs
‑
1多肽(即图中的cs
‑
1 peptide)或cs
‑
2的多肽(即图中的cs
‑
2 peptide)与ctsl共同孵育后进行质谱检测。其中,cs
‑
1多肽可在249l(即seq id no.1第19位和20位之间)与259t处(即seq id no.1第29位和30位之间)被切割,cs
‑
2多肽可在636y处(即seq id no.2第36位和37位之间)被切割。
25.图2a是本发明实施例1 sars
‑
cov
‑
2突变株中cs
‑
1和cs
‑
2周围残基的氨基酸序列比对;符号“*”表示在所有测试序列中保守的氨基酸残基, 符号“:”和“.”表示具有高度相似或相似生化特性的异质氨基酸残基的位置;图2b是本发明实施例1源自蝙蝠和穿山甲的 sars
‑
cov
‑
2 和 sars
‑
cov
‑
1 相关冠状病毒中cs
‑
1和cs
‑
2周围残基的氨基酸序列比对;符号“*”表示在所有测试序列中保守的氨基酸残基, 符号“:”和“.”表示具有高度相似或相似生化特性的异质氨基酸残基的位置;图2c是本发明实施例1能够感染人的所有冠状病毒中cs
‑
1和cs
‑
2周围残基的氨基酸序列比对;符号“*”表示在所有测试序列中保守的氨基酸残基, 符号“:”和“.”表示具有高度相似或相似生化特性的异质氨基酸残基的位置;图3a是本发明实施例1中cs
‑
1、cs
‑
2 和 s1/s2 切割位点的突变体介绍;图3b是本发明实施例1中在 llc
‑
mk2、vero、293t/hace2 和 huh7 细胞中评估了在 cs
‑
1 和 cs
‑
2 中具有不同点突变的假病毒的感染性;假病毒感染性通过萤光素酶测定法测量并显示为始萤光素酶活性 (n=4), 通过单向方差分析和 tukey检验评估统计显着
性;纵坐标为萤光素酶活性;图3 c是本发明实施例1中荧光素酶基因表达由 ere 启动子驱动,esr1(激活子)结合并激活ere启动子以上调荧光素酶表达示意图;图3d是本发明实施例1中细胞融合实验原理示意图;huh7 效应细胞用表达 ere
‑
荧光素酶和不同 s 蛋白的质粒(详见图3e说明)共转染;另一皿 huh7 靶细胞用表达 esr1 的质粒转染。24小时后,将效应细胞分离并加入靶细胞中30
‑
60分钟。然后,去除上清液并用 pbs 或 ctsl处理 20 分钟。加入培养基终止反应,继续培养 24小时,使细胞融合。当靶细胞和效应细胞融合形成合胞体时,esr1 结合并激活 ere 启动子以上调荧光素酶表达。然后测量荧光素酶活性作为融合率的代表。
26.图3e是本发明实施例1中细胞融合实验结果;横坐标wt、cs
‑
1m、cs
‑
2m或cs
‑
1m+2m为不同 s 蛋白的质粒,huh7效应细胞分别转染上述不同质粒后,按照d所述方法进行细胞融合实验,所得结果;不同 s 蛋白的质粒获得方法:包括将wt、cs
‑
1m、cs
‑
2m或cs
‑
1m+2m蛋白的编码基因插入pcdna3.1所得质粒;wt为实施例1的第“一”中sars
‑
cov
‑
2 的s蛋白胞外段在 k986 和 v987 处用脯氨酸取代,弗林蛋白酶裂解位点(s1/s2 位点,r682 至 r685)处用“gsas”取代以增强的稳定性,所得s蛋白突变体;cs
‑
1m将wt的seq id no.1第18、19、28和29位的氨基酸均替换为甘氨酸,其他氨基酸残基保持不变得到的突变体;cs
‑
2 突变体为将s蛋白假病毒(wt)的seq id no.2第35和36位氨基酸均替换为甘氨酸,其他氨基酸残基保持不变得到的突变体;组合突变体为将s蛋白假病毒(wt)的seq id no.1第18、19、28和29位的氨基酸和seq id no.2第35和36位的氨基酸均替换为甘氨酸,其他氨基酸残基保持不变得到的突变体;纵坐标为融合率。
27.图3f是本发明实施例1中明场显微镜直接观察合胞体的形成结果;huh7 效应细胞用表达 ere
‑
荧光素酶(吉凯基因)和不同 s 蛋白的质粒(wt、cs
‑
1m、cs
‑
2m或cs
‑
1m+2m)共转染,另一皿 huh7 靶细胞用表达esr1的质粒转染。24小时后,将效应细胞分离并加入靶细胞中30
‑
60分钟。然后,去除上清液并用 pbs 或 ctsl(4 μg/ml,ph=5.8)处理 20 分钟。在培养基(比例尺,50 μm)中额外培养 10
‑
16 小时后获取图像,黑色箭头表示合胞体,显示了来自三个独立实验的代表性数据;不同 s 蛋白的质粒包括将wt、cs
‑
1m、cs
‑
2m或cs
‑
1m+2m蛋白的编码基因插入pcdna3.1所得质粒;wt为实施例1的第“一”中sars
‑
cov
‑
2 的s蛋白胞外段在 k986 和 v987 处用脯氨酸取代,弗林蛋白酶裂解位点(s1/s2 位点,r682 至 r685)处用“gsas”取代以增强的稳定性,所得s蛋白突变体;cs
‑
1m将wt的seq id no.1第18、19、28和29位的氨基酸均替换为甘氨酸,其他氨基酸残基保持不变得到的突变体;cs
‑
2 突变体为将s蛋白假病毒(wt)的seq id no.2第35和36位氨基酸均替换为甘氨酸,其他氨基酸残基保持不变得到的突变体;组合突变体为将s蛋白假病毒(wt)的seq id no.1第18、19、28和29位的氨基酸和seq id no.2第35和36位的氨基酸均替换为甘氨酸,其他氨基酸残基保持不变得到的突变体。
28.图3g是本发明实施例1中 huh7 细胞中的ctsl 基因的过表达或敲低分别剂量依赖性地促进或抑制了野生型和三个弗林蛋白酶突变的psv 突变体(fm
‑
delta, fm
‑
araa 和 fm
‑
gsas)的感染。psv 感染性通过荧光素酶测定法测量,并显示为相对荧光素酶活性 (n = 3);通过单向方差分析和 tukey事后检验评估统计显著性;control和scramble都是空白载体(对照);纵坐标为萤光素酶活性;横坐标sictsl表示的是敲减ctsl所用的sirna序列混合
物的浓度,混合物由ctsl
‑1‑ꢀ
ctsl
‑
6组成(ctsl
‑
1、ctsl
‑
2、ctsl
‑
3、ctsl
‑1‑ꢀ
ctsl
‑
4、ctsl
‑
5和 ctsl
‑
6的摩尔比为1:1:1:1:1:1);ctsl
‑1‑ꢀ
ctsl
‑
6的序列见表1,过表达为ctsl 基因序列插入pcdna3.1质粒所得。
29.图3h是本发明实施例1中 ctsl 促进由 fm
‑
araa 的突变s 蛋白诱导的合胞体形成;用 ere
‑
荧光素酶质粒和 fm
‑
araa s 或空白载体(对照)共转染效应细胞,用esr1表达质粒转染靶细胞,效应细胞和靶细胞混合后,去除上清液并用pbs或ctsl(8μg/ml和16μg/ml)处理。然后测量荧光素酶活性并将其标准化为对照组(n=4)。通过单向方差分析和 tukey 事后检验评估统计显着性;纵坐标为融合率。
30.图3i是本发明实施例1中ctsl在fm
‑
araa突变s蛋白引起的合胞体形成中的作用;空白载体或fm
‑
araa突变s蛋白表达质粒转染huh7细胞,在不存在(pbs)或存在 ctsl(2 或 4 μg/ml)的情况下处理细胞,在培养基(比例尺,50 μm)中额外培养 10
‑
16 小时后获取图像。黑色箭头表示合胞体。显示了来自四个独立实验的代表性数据;fm
‑
araa 突变s蛋白表达质粒为将fm
‑
araa 突变s蛋白的编码基因插入pcdna3.1后,所得;fm
‑
araa 突变s蛋白为将wt中的qtnsprrarsva中的
682
rrar
685
残基替换为
682
araa
685
后,其他氨基酸残基保持不变得到的突变体;数据表示为平均值
ꢀ±ꢀ
sem 值, * p < 0.05,** p < 0.01,*** p < 0.001,*** p < 0.0001。
31.图4a是本发明实施例1中六个化合物在无细胞毒性剂量下都能有效地阻断sars
‑
cov
‑
2感染;vero e6 细胞用递增浓度的每种化合物预处理 16 小时,然后以 0.01 的 moi 感染 sars
‑
cov
‑
2,在感染后 24 小时,通过 rt
‑
qpcr 定量上清液中的病毒 rna 拷贝。用 cck 试剂盒 (transgen biotech) (n=3) 评估细胞活力。纵坐标为相对感染百分比。
32.图4b是本发明实施例1中六个化合物的抗病毒效果;vero e6 细胞用递增浓度的每种化合物预处理 16 小时,然后用不同的 sars
‑
cov
‑
2 变体 psv 感染;在感染后 24 小时,通过荧光素酶测定法测量感染性;通过带有 dunnett 事后检验的双向 anova 评估指定变体和 wt psv 之间的统计显著性。* p < 0.05,** p < 0.01;纵坐标为抑制率(n = 3);图5a是本发明实施例1中e64d 和 z
‑
fy
‑
cho预防性和治疗性给药方案时间表; e64d 和 z
‑
fy
‑
cho 在
ꢀ‑
2~3 dpi 腹膜内给药作为预防性治疗,并在 0 dpi 时用 10
6 pfu的sars
‑
cov
‑
2真病毒感染小鼠;两种药物均在0~3 dpi进行治疗,在 4 dpi 收集组织样本;图5b是本发明实施例1中小鼠肺和鼻甲组织中的病毒 rna 拷贝(n = 5 只小鼠/组),虚线表示检测下限 (lod),通过单向方差分析和 tukey 事后检验评估指定组和对照组之间的统计显著性;图5c是本发明实施例1中来自感染 sars
‑
cov
‑
2 的 hace2转基因小鼠肺组织学分析的代表性图像,每个图像中框区域的放大视图显示在相应图像下方,表示炎症细胞浸润,表示细支气管上皮细胞变性, 表示肺泡间隔增厚;图5d是本发明实施例1中小鼠组织评分结果;通过对细支气管上皮细胞损伤 (0
‑
10分)、肺泡损伤 (0
‑
10分) 和血管和细支气管中炎症细胞浸润 (0
‑
10分)的严重程度进行分级并求和,对每个肺组织进行半定量组织学评分这些分数来计算总分,正常 = 0分,不确定 = 1
‑
2分,轻度 = 3
‑
4分,中度 = 5
‑
7分,严重 = 8
‑
10分, (n=3) 通过单向方差分析和 tukey 事后检验评估指定组和对照组之间
的统计显著性。
具体实施方式
33.表1:针对人 ctsl 的 sirna 序列混合物实施例1一、ctsl在三个新位点切割 sars
‑
cov
‑
2 的s蛋白在sars
‑
cov
‑
2病毒表面的三聚体 s 蛋白包含 s1 和 s2 亚基(图1a 和图1b)。s1 亚基与宿主细胞受体结合,而 s2 亚基参与病毒
‑
细胞膜融合过程,然后将病毒遗传物质释放到宿主细胞中。为了研究经 ctsl 处理的s蛋白的结构和功能变化,本实施例利用哺乳动物细胞表达和纯化了sars
‑
cov
‑
2 的s蛋白(氨基酸序列见genbank id: mn908947.3)胞外段(残基 m1
–
q1208)。将sars
‑
cov
‑
2 的s蛋白胞外段在 k986 和 v987 处用脯氨酸取代,弗林蛋白酶裂解位点(s1 /s2 位点,r682 至 r685)处用“gsas”取代以增强的稳定性,该s蛋白突变体广泛用于结构和功能分析。s蛋白突变体在与不同浓度的ctsl(0~8 μg/ml)共孵育后,被裂解为两个主要片段,在sds
‑
page中分子量分别为~60 kda(带 1)和~100 kda(带 2)。切割效率与ctsl浓度呈剂量依赖性(图 1c)。针对这两个片段进行n端氨基酸测序,本实施例确定了sars
‑
cov
‑
2 s蛋白上ctsl的两个新切割位点,259t(命名为 ctsl 切割位点 1,或 cs
‑
1,即seq id no.1 :ginitrfqtlclalhrsyltpgdsssgwtagaaayyvgylqprtfllk的第29位)和636y(命名为 ctsl 切割位点 2,或 cs
ꢀ‑
2,即seq id no.2:gtntsnqvavlyqdvnctevpvaihadqltptwrvystgsnvfqtragcl的第36位)(图1b)。 cs
‑
1位于s1亚基的n端结构域(ntd),而cs
‑
2位于s1亚基的c端结构域(ctd)(图1a)。与弗林蛋白酶的 s1/s2 位点相同,两个 ctsl 切割位点都位于s蛋白暴露位置的氨基酸环中,蛋白酶可接近这些位点(图1b)。
34.为了验证 ctsl 可以在 cs
‑
1 和 cs
‑
2 位点切割s蛋白,本实施例根据sars
‑
cov
‑
2的s 蛋白的三级结构,根据259t和636y的氨基酸环范围设计了两个多肽(图1e)。与 ctsl 酶孵育后,通过质谱鉴定序列,cs
‑
1多肽(氨基酸245
‑
262)可在 259t 位点裂解,cs
‑
2多肽(氨基酸618
‑
641)可在 636y 处裂解。意外的是,本实施例发现了另一个新的 249l 切割位点,它位于cs
‑
1切点259t的上游侧。考虑到249l和259t在s蛋白上的位置比较接近,经过ctsl处理后,在249l位点被切割的250
‑
636片段可以在259t位点进一步被切割为260
‑
636片段,因此249l位点很难用n端测序法。 因此,在cs
‑
1区域应该有两个ctsl切割位点,249l(seq id no.1的第19位)和259t,分别命名为 cs
‑
1a 和 cs
‑
1b 位点。
35.由于ctsl与其底物的识别主要取决于切割位点残基(命名为p1残基)及其上游残
基(命名为p2残基),本实施例将多肽cs
‑
1a、cs
‑
1b及cs
‑
2的p1和p2残基突变为甘氨酸,再加入ctsl切割,质谱结果显示,这些突变的多肽都不能被 ctsl 在相应位点切割。此外,本实施例制备了在这些切割位点具有双甘氨酸突变的 s 蛋白,并用 ctsl 切割它们。结果表明,当切割位点被甘氨酸取代时,ctsl不能切割s蛋白(图1d)。这些结果不仅证明s的249l(cs
‑
1a)、259t(cs
‑
1b)和636y(cs
‑
2)是ctsl的特异性切割位点,而且表明本实施例已经无遗漏地鉴定了所有的切割位点。
36.二、ctsl 切割位点在 sars
‑
2 突变株中高度保守由于之前没有报道过s蛋白上的这三个 ctsl切割位点,本实施例研究了它们在不同sars
‑
cov
‑
2突变株之间的序列保守性。到目前为止,已经在不同国家发现了许多 sars
‑
cov
‑
2突变株(图2a)。 这些突变株可能会拥有增强传播、致病性、或免疫逃逸这三个特点,并且是大多数国家的主要流行突变株。许多中和抗体的靶点位于s蛋白rbd 区域,这些靶点在这些突变株中发生了突变。然而,cs
‑
1a 处的 249l(p2位点)和 250t(p1位点)、cs
‑
1b 处的 258w(p2位点)和 259t(p1位点)以及 cs
‑
2的635v(p2位点)和 636y(p1位点),在所有关注的 sars
‑
cov
‑
2的突变株中具有高度保守性(图2a)。它表明ctsl的切割对病毒突变逃逸具有高度抵抗力,表明这些 ctsl 切割位点对 sars
‑
cov
‑
2生命周期至关重要。
[0037] 据报道,sars
‑
cov
‑
2可能源自蝙蝠冠状病毒,例如ratg13,因此本实施例对蝙蝠和穿山甲中发现的冠状病毒中ctsl的切点周围氨基酸序列进行了序列比对。如图2b 所示,cs
‑
2位点在大多数sars
‑
cov和 sars
‑
cov
‑
2 相关的冠状病毒中高度保守,而cs
‑
1b位点仅在 sars
‑
cov
‑
2 相关的冠状病毒中保守,而在 sars
‑
cv相关的冠状病毒中不保守。更有趣的是,cs
‑
1a 位点在所有这些冠状病毒中都是可变的,仅存在于ratg13 和pangolin cov guangxi中。这表明这三个ctsl切点在不同动物宿主的进化过程中可能具有不同的作用。cs
‑
2 位点似乎对病毒的生命周期最为重要,而 cs
‑
1b 可能是从sars
‑
cov逐渐进化到 sars
‑
cov
‑
2才出现的,以增强病毒感染,而 cs
‑
1a可能进化得最晚。此外,本实施例还比较了已知可以感染人类的7种冠状病毒中,三个 ctsl 裂解位点的保守性。cs
‑
2存在于 sars
‑
cov 和 sars
‑
cov
‑
2 中,而 cs
‑
1a 和 cs
‑
1b 仅存在于 sars
‑
cov
‑
2(图2c),表明 ctsl 可能在 sars
‑
cov
‑
2 感染过程中发挥某种独特的重要作用。
[0038]
三、ctsl 切割位点对于 sars
‑
cov
‑
2 感染和细胞融合至关重要由于 ctsl 在体外有效切割 sars
‑
cov
‑
2的s蛋白,本实施例研究了 ctsl 在病毒感染过程中的功能。本实施例根据三个切割位点制备了几种sars
‑
cov
‑
2 s蛋白假病毒(psv)突变体,包括 cs
‑
1 突变体(cs
‑
1m)、cs
‑
2 突变体(cs
‑
2m)及其组合突变体(cs
‑
1m+2m)(图3a),cs
‑
1 突变体为将s蛋白假病毒(wt)的seq id no.1第18、19、28和29位的氨基酸均替换为甘氨酸,其他氨基酸残基保持不变得到的突变体;cs
‑
2 突变体为将s蛋白假病毒(wt)的seq id no.2第35和36位氨基酸均替换为甘氨酸,其他氨基酸残基保持不变得到的突变体;组合突变体为将s蛋白假病毒(wt)的seq id no.1第18、19、28和29位的氨基酸和seq id no.2第35和36位的氨基酸均替换为甘氨酸,其他氨基酸残基保持不变得到的突变体。作为适当的对照,本实施例制备了三个弗林蛋白酶突变的psv 突变体,可以逃避弗林蛋白酶切割,分别命名为弗林蛋白酶突变体(fm)
‑
delta(残基 682
‑
685 缺失,即s蛋白假病毒的qtnsprrarsva中的残基 682
rrar
685 缺失,其他氨基酸残基保持不变得到的突变体)、fm
‑
araa(即s蛋白假病毒的qtnsprrarsva中的
682
rrar
685
残基替换为
682
araa
685
后,其他氨基酸残
基保持不变得到的突变体)和 fm
‑
gsas(即s蛋白假病毒的qtnsprrarsva中的
682
rrar
685
残基替换为
682
gsas
685
后,其他氨基酸残基保持不变得到的突变体)。然后,本实施例使用四种人类和猴细胞系(分别为lic
‑
mk2、vero、hek
‑
293t/hace2(简称293t/hace2)、huh7)比较了野生型 (wt) 和突变型 psvs 的病毒感染性,现有技术已经记载了这些细胞系对 sars
‑
cov
‑
2 感染的易感性最高。与 wt 组相比,fm
‑
delta、fm
‑
araa 和 fm
‑
gsas 的感染性没有降低太多。然而,一旦 ctsl 切割位点突变为甘氨酸,这些 psv突变体在所有四种细胞系中都失去了大部分感染能力。当三个 ctsl 切割位点都发生突变时,与 wt psv 相比,病毒感染性降低了 100~700 倍(图3b)。结果表明,cs
‑
1 和 cs
‑
2 的三个新 ctsl 切割位点对于 sars
‑
cov
‑
2 感染至关重要。此外,与 cs
‑
1 位点相比,cs
‑
2 位点似乎在 sars
‑
cov
‑
2 病毒进入方面发挥了更重要的作用。在不同的 psv 浓度下观察到类似的结果。
[0039]
sars
‑
cov
‑
2的s蛋白可导致细胞融合,这是 sars
‑
cov
‑
2 感染的一个特征。在covid
‑
19 患者肺组织中发现了大量的合胞体,这表明该作用在疾病致病过程中起作用。本实施例通过 esr1
‑
ere转录激活系统(图 3c、图 3d和图3e,图3c
‑
图3e是由 cs 突变体 sars
‑
cov
‑
2 s 蛋白诱导的合胞体形成的定量分析)对合胞体的形成进行量化,并通过明场显微镜直接观察合胞体的形成(图3f)。与对照组相比,cs
‑
1m组显著减少合胞体形成,cs
‑
2m组对合胞体形成的抑制更为严重,而cs
‑
1m+2m组几乎没有观察到合胞体形成(图3e和图3f)。 更重要的是,这些突变体的抑制作用不能通过将 ctsl 酶添加到系统中来挽救(图3e和图3f),这证实了ctsl是通过切割本实施例发现的cs
‑
1和 cs
‑
2位点来增强合胞体形成的。总之,本实施例证实了,ctsl通过在 cs
‑
1 和 cs
‑
2 处切割病毒s 蛋白对 sars
‑
cov
‑
2 感染起至关重要的作用。
[0040]
四、ctsl促进sars
‑
cov
‑
2感染的作用不依赖于弗林蛋白酶由于cs
‑
2区域非常接近弗林蛋白酶切割位点,本实施例接下来用不同的弗林蛋白酶突变(fm)的psvs来比较这两种蛋白酶的功能。在过表达或敲减ctsl的情况下,在人huh7细胞系中进行psv感染实验。结果显示,ctsl基因的过表达明显增加了sars
‑
cov
‑
2 psv的感染,而ctsl基因的敲减则导致其感染效率明显降低(图3g),证实了ctsl在sars
‑
cov
‑
2感染过程中起着关键作用。然而,与wt psv相似,ctsl基因的过表达或敲减也会以剂量依赖的方式明显影响三个fm突变体psvs的感染水平(图3g)。这表明ctsl增强sars
‑
cov
‑
2病毒的进入是独立于弗林蛋白酶发挥作用的。
[0041]
据报道,弗林蛋白酶切位点对sars
‑
cov
‑
2 s蛋白导致的细胞融合是重要的。这里本实施例用同样的细胞融合系统(图3d)来评估ctsl在fm突变s蛋白引起的合胞体形成中的作用。fm
‑
araa的突变s蛋白诱导的合胞体形成很少,其水平与对照组相似(图3h),fm
‑
araa的突变s蛋白为wt中的qtnsprrarsva序列上的
682
rrar
685
残基替换为
682
araa
685
后,其他氨基酸残基保持不变得到的突变体,wt为实施例1的第“一”中sars
‑
cov
‑
2 的s蛋白胞外段在 k986 和 v987 处用脯氨酸取代,弗林蛋白酶裂解位点(s1/s2 位点,r682 至 r685)处用“gsas”取代以增强的稳定性,所得s蛋白突变体。然而,一旦ctsl被添加到突变s蛋白组中,大量合胞体开始形成,而且这种拯救效果与ctsl的浓度呈剂量依赖关系(图3h和图3i)。这表明在病毒感染期间,ctsl可以替代弗林蛋白酶来激活病毒s蛋白。
[0042]
五、ctsl抑制剂可预防sars
‑
cov
‑
2的感染由于ctsl在介导sars
‑
cov
‑
2感染中起着关键作用,本实施例选择了6个在50mm浓
度下能抑制ctsl活性超过90%的化合物(z
‑
fy
‑
cho、oxocarbazate、mdl
‑
28170、k777、cathepsin inhibitor1和e64d)来评估它们在预防sars
‑
cov
‑
2感染中的效果,其中z
‑
fy
‑
cho和oxocarbazate是选择性的ctsl抑制剂,其他4个化合物是广泛的组织蛋白酶抑制剂。
[0043]
首先,在体外使用sars
‑
cov
‑
2活病毒,发现所有六个化合物在无细胞毒性剂量下都能有效地阻断sars
‑
cov
‑
2感染,ec
50
从35 nm到10.89mm不等(图4a)。在最大的无毒剂量下,所有化合物都使veroe6细胞中的病毒rna负荷下降了约103至104倍。这一发现证实了ctsl在sars
‑
cov
‑
2感染中起关键作用。
[0044]
其次,本实施例使用最近出现的sars
‑
cov
‑
2突变株的s蛋白psvs进一步研究了这六个化合物的抗病毒效果(图4b)。正如预期的那样,这六个化合物也有效地防止了所有的突变株s蛋白psvs的感染,其中e64d对p.1表现出更高的效率,而mdl
‑
28170和oxocarbazate对b.1.429更有效。这表明ctsl抑制剂可以抵抗包括b.1.351在内的突变株的免疫逃逸作用,而b.1.351降低了目前大多数疫苗的效果。
[0045]
最后,本实施例研究了ctsl抑制剂在人类ace2(hace2)转基因小鼠体内的抗病毒作用,这种小鼠可感染sars
‑
cov
‑
2真病毒(哈尔滨兽医研究所)。本实施例选择了两个已经被验证为对体内研究安全的化合物,e64d和z
‑
fy
‑
cho。e64d和z
‑
fy
‑
cho被给予预防性和治疗性给药方案,并以适当的剂量治疗小鼠。每只小鼠在0dpi时通过鼻内灌注10
6 pfu的sars
‑
cov
‑
2真病毒感染。在4dpi(感染后一天)收集组织样本(图5a),此时病毒量达到高峰,并有明显的组织病理学变化。两种化合物都能使肺组织中的病毒rna拷贝数明显减少,在2只用z
‑
fy
‑
cho治疗的小鼠和4只用e64d治疗的小鼠的肺组织中都检测不到病毒负荷(低于检测下限,lod)(图5b)。其中,预防方案对病毒感染有更好的保护,而治疗方案也表现出明显的疗效。同时,这两种药物也明显减少了鼻甲中的病毒载量(图5b)。肺部病理学结果显示,对照组的小鼠表现出中度病理变化,表现出大面积的肺部隔膜增厚、炎症细胞渗透和支气管上皮细胞变性。相比之下,e64d
‑
和z
‑
fy
‑
cho治疗小鼠的肺部表现出更加正常的形态和较少的渗透(图5c和5d)。综合起来,这些结果表明ctsl抑制剂能有效阻断体外和体内的sars
‑
cov
‑
2感染,对病毒突变逃逸具有抵抗力。
[0046]
显然,上述实施例仅仅是为清楚地说明所作的举例,而并非对实施方式的限定。对于所属领域的普通技术人员来说,在上述说明的基础上还可以做出其它不同形式的变化或变动。这里无需也无法对所有的实施方式予以穷举。而由此所引伸出的显而易见的变化或变动仍处于本发明创造的保护范围之中。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1