一种N-乙酰神经氨酸的制备方法与流程
一种n-乙酰神经氨酸的制备方法
技术领域
1.本发明属于化合物制备领域,具体涉及一种n-乙酰神经氨酸的制备方法。
背景技术:
2.n-乙酰神经氨酸是最普遍存在的唾液酸种类,占据细胞膜中糖蛋白、糖脂或寡糖的末端位置,在生物学、病理学和免疫学过程中起着重要作用,如参与细胞识别、信号转导、肿瘤发生和受精过程。因此,n-乙酰神经氨酸己被用作增强免疫力和促进婴儿大脑发育的营养保健品,同时n-乙酰神经氨酸由于其显着的抗癌、抗粘连和抗炎作用在药物方面有重大应用。因此,随着新应用的推出,n-乙酰神经氨酸的需求将不断增加。
3.目前n-乙酰神经氨酸的生产方法包括:天然产物提取、化学合成、生物酶催转化、基因工程菌直接发酵及聚唾液酸发酵水解法。绝大多数天然产物中的neu5ac含量低,而且组分非常复杂,从中提取neu5ac伴随着复杂的过程,回收率低,因此天然产物提取很难满足大规模生产的要求。化学合成法由于繁琐的基团保护、去保护等操作,以及手性异构副产物的存在,该方法亦无法应用于工业生产。生物酶催转化因为转化前体n-乙酰葡萄糖胺和丙酮酸成本高,导致合成成本高。聚唾液酸水解法由于发酵产量低及后处理复杂,成本也居高不下。基因工程菌直接发酵通过对大肠杆菌模式菌株基因改造,使之能够利用普通碳源进行高密度发酵,并达到较高的neu5ac产量,近年来该方法受到广泛重视。
4.中国专利文献
″
一种提高发酵产n-乙酰神经氨酸的方法及发酵液(专利申请号为cn201811124497.2)
″
公开了一种提高发酵产n-乙酰神经氨酸的方法,其步骤为:在液体发酵培养基中接种产n-乙酰神经氨酸菌种的种子培养液进行发酵,液体发酵培养基和种子培养液的体积比为25-35∶1,发酵过程维持ph 6.5-7,并在液体发酵培养基中的碳源耗完后,以5-6g/(l
·
h)的速率补充碳源;每隔36-60h排出85%-95%体积的发酵液,并补入相同体积、不含碳源的新鲜液体培养基。该发明方法工序简单,生产效率高,但是仍然存在有n-乙酰神经氨酸产量有待提高的问题。
技术实现要素:
5.本发明的目的是提供一种n-乙酰神经氨酸的制备方法,以解决在中国专利文献
″
一种提高发酵产n-乙酰神经氨酸的方法及发酵液(专利申请号为cn201811124497.2)
″
公开的基础上,如何优化组分、用量、工艺等,从而提高n-乙酰神经氨酸的产量的问题。
6.为了解决以上技术问题,本发明采用以下技术方案:
7.一种n-乙酰神经氨酸的制备方法,包括以下步骤:
8.s1:以大肠杆菌e.colik-12mg1655为出发菌,敲除其基因组中的nage基因和manxyz、nanatek基因簇,并将cneub基因和bage基因克隆到ptrcc99a载体,分别加上trc启动子实现2个基因在大肠杆菌中的共表达;
9.s2:敲除与n-乙酰神经氨酸转运载体相关基因nant以及与葡萄糖胺转运相关的磷酸转移酶系统相关基因nage;敲除影响葡萄糖胺6-磷酸合成酶的编码基因glms表达水平的
yhbj基因来提高转录水平;
10.s3:克隆s.cerevisiae eby100的葡萄糖胺酰基转移酶的编码基因gnat,通过基因敲除的方法整合到大肠杆菌染色体的naga-nagb基因位点,同时将naga-nagb基因删除,阻断6-磷酸-n-乙酰葡萄糖胺的分解代谢途径,从而催化6-磷酸葡萄糖胺转化生成6-磷酸-n-乙酰葡萄糖胺;将集胞藻age编码基因slr整合到染色体的laci基因位点,同时将laci基因删除;
11.s4:敲除醋酸激酶的编码基因acka与磷酸乙酰转移酶的编码基因pta阻断乙酸合成的其中一条支路;同时敲除乳酸脱氢酶的编码基因idha阻断乳酸生成途径;继续敲除丙酮酸氧化酶的编码基因poxb以阻断乙酸合成的另一条代谢支路;
12.s5:将上述菌种接种于培养基中,待细胞浓度上升后添加诱导剂,诱导胞内酶的表达;
13.s6:将菌体收集浓缩后悬浮于添加了底物n-乙酰葡萄糖胺的转化培养基;
14.s7:用高效液相色谱测定n-乙酰神经氨酸的含量。
15.优选地,所述步骤s5的培养基中含有胰蛋白胨8-15g/l、酵母2-8g/l、氯化钠8-15g/l和氨苄青霉素0.08-0.12g/l,ph为7.0。
16.优选地,所述步骤s5中将培养基接种于摇瓶中,并置于36-38℃摇床中培养2-4h,当细胞浓度达到1.2时,添加诱导剂至浓度为0.2mmol/l。
17.优选地,所述步骤s5中将培养基继续振荡培养4h,将菌液收集于冰浴中静置10min,然后于2-5℃的条件下离心15min,除去上清液,用预冷的生理盐水洗涤菌体2次。
18.优选地,所述培养基置于37℃摇床中,旋转速度为200-250r/min,培养3h。
19.优选地,所述诱导剂为异丙基硫代半乳糖苷。
20.优选地,所述步骤s6中向转化培养基中再次添加诱导剂至浓度为0.2mmol/l,于37℃摇床中,旋转速度为200-250r/min,培养3h。
21.优选地,所述步骤s6中转化培养基的成分为甘油10-14g/l、n-乙酰葡萄糖胺75-90g/l、硫酸镁1-3g/l、k2hpo4·
3h2o 28-34g/l,ph为7.0。
22.本发明具有以下有益效果:
23.(1)本发明在n-乙酰神经氨酸的制备过程中添加甘油、n-乙酰葡萄糖胺和异丙基硫代半乳糖苷起到了协同作用,协同提高了n-乙酰神经氨酸的产量,这是因为:在转化培养基中添加的n-乙酰葡萄糖胺在cneub基因的催化下与胞内的磷酸烯醇式丙酮酸合成n-乙酰神经氨酸,从而可以提高n-乙酰神经氨酸的产量。并且,在转化培养基中添加诱导剂异丙基硫代半乳糖苷时,重组大肠杆菌能够重新合成cneub基因,使得胞内cneub基因活性维持在较高的水平,从而促进n-乙酰神经氨酸的生成。在转化培养基中添加甘油对细胞膜的流动性和通透性具有促进作用,有利于n-乙酰葡萄糖胺进入胞内和n-乙酰神经氨酸向胞外分泌;并且,以甘油为碳源时,甘油的跨膜运输不依赖磷酸转运系统,因此不消化胞内的磷酸烯醇式丙酮酸,从而使得更多的磷酸烯醇式丙酮酸流向n-乙酰神经氨酸合成的方向,在生物量相同的条件下,提高了n-乙酰神经氨酸的速率和产量。
24.(2)步骤s4中阻断胞内的磷酸烯醇式丙酮酸的降解途径,增加胞内的磷酸烯醇式丙酮酸的积累,敲除醋酸激酶的编码基因acka与磷酸乙酰转移酶的编码基因pta阻断乙酸合成的其中一条支路;同时敲除乳酸脱氢酶的编码基因idha阻断乳酸生成途径;继续敲除
丙酮酸氧化酶的编码基因poxb以阻断乙酸合成的另一条代谢支路,进一步提高胞内的磷酸烯醇式丙酮酸的产量,从而提高n-乙酰神经氨酸的产量。
25.(3)背景技术所引用的专利文献
″
一种提高发酵产n-乙酰神经氨酸的方法及发酵液(专利申请号为cn201811124497.2)
″
,虽然该发明方法工序简单,生产效率高,但是仍然存在有n-乙酰神经氨酸产量有待提高的问题。基于为了解决以上技术问题,本发明才对该发明的配方和工艺进行进一步的优化和改良,经过多次试验研究发现,当n-乙酰神经氨酸制备方法中在转化培养基中添加甘油、n-乙酰葡萄糖胺和异丙基硫代半乳糖苷时,n-乙酰葡萄糖胺在cneub基因的催化下与胞内的磷酸烯醇式丙酮酸合成n-乙酰神经氨酸;异丙基硫代半乳糖苷使得胞内cneub基因活性维持在较高的水平;提高n-乙酰神经氨酸的产量;甘油对细胞膜的流动性和通透性具有促进作用,使得更多的磷酸烯醇式丙酮酸流向n-乙酰神经氨酸合成的方向,能够解决背景技术文件中出现的技术问题,产生了意想不到的效果。
具体实施方式
26.为了更好地理解本发明,现采用以下实施例加以说明,以下实施例属于本发明的保护范围,但不限制本发明的保护范围。
27.以下实施例中所述的一种n-乙酰神经氨酸的制备方法,包括以下步骤:
28.s1:以大肠杆菌e.colik-12mg1655为出发菌,敲除其基因组中的nage基因和manxyz、nanatek基因簇,并将cneub基因和bage基因克隆到ptrcc99a载体,分别加上trc启动子实现2个基因在大肠杆菌中的共表达;
29.s2:敲除与n-乙酰神经氨酸转运载体相关基因nant以及与葡萄糖胺转运相关的磷酸转移酶系统相关基因nage;敲除影响葡萄糖胺6-磷酸合成酶的编码基因glms表达水平的yhbj基因来提高转录水平;
30.s3:克隆s.cerevisiae eby100的葡萄糖胺酰基转移酶的编码基因gnat,通过基因敲除的方法整合到大肠杆菌染色体的naga-nagb基因位点,同时将naga-nagb基因删除,阻断6-磷酸-n-乙酰葡萄糖胺的分解代谢途径,从而催化6-磷酸葡萄糖胺转化生成6-磷酸-n-乙酰葡萄糖胺;将集胞藻age编码基因slr整合到染色体的laci基因位点,同时将laci基因删除;
31.s4:敲除醋酸激酶的编码基因acka与磷酸乙酰转移酶的编码基因pta阻断乙酸合成的其中一条支路;同时敲除乳酸脱氢酶的编码基因idha阻断乳酸生成途径;继续敲除丙酮酸氧化酶的编码基因poxb以阻断乙酸合成的另一条代谢支路;
32.s5:将上述菌种接种于培养基中,培养基中含有胰蛋白胨8-15g/l、酵母2-8g/l、氯化钠8-15g/l和氨苄青霉素0.08-0.12g/l,ph为7.0;将培养基接种于摇瓶中,并置于36-38℃摇床中,旋转速度为200-250r/min,培养2-4h,当细胞浓度达到1.2时,添加异丙基硫代半乳糖苷至浓度为0.2mmol/l,诱导胞内酶的表达;将培养基继续振荡培养4h,将菌液收集于冰浴中静置10min,然后于2-5℃的条件下离心15min,除去上清液,用预冷的生理盐水洗涤菌体2次;
33.s6:将菌体收集浓缩后悬浮于添加了底物n-乙酰葡萄糖胺的转化培养基,转化培养基的成分为甘油10-14g/l、n-乙酰葡萄糖胺75-90g/l、硫酸镁1-3g/l、k2hpo4·
3h2o28-34g/l,ph为7.0;向转化培养基中再次添加异丙基硫代半乳糖苷至浓度为0.2mmol/l,于37
乙酰葡萄糖胺;将集胞藻age编码基因slr整合到染色体的laci基因位点,同时将laci基因删除;
49.s4:敲除醋酸激酶的编码基因acka与磷酸乙酰转移酶的编码基因pta阻断乙酸合成的其中一条支路;同时敲除乳酸脱氢酶的编码基因idha阻断乳酸生成途径;继续敲除丙酮酸氧化酶的编码基因poxb以阻断乙酸合成的另一条代谢支路;
50.s5:将上述菌种接种于培养基中,培养基中含有胰蛋白胨15g/l、酵母6g/l、氯化钠8g/l和氨苄青霉素0.12g/l,ph为7.0;将培养基接种于摇瓶中,并置于38℃摇床中,旋转速度为220r/min,培养2h,当细胞浓度达到1.2时,添加异丙基硫代半乳糖苷至浓度为0.2mmol/l,诱导胞内酶的表达;将培养基继续振荡培养4h,将菌液收集于冰浴中静置10min,然后于3℃的条件下离心15min,除去上清液,用预冷的生理盐水洗涤菌体2次;
51.s6:将菌体收集浓缩后悬浮于添加了底物n-乙酰葡萄糖胺的转化培养基,转化培养基的成分为甘油13g/l、n-乙酰葡萄糖胺90g/l、硫酸镁1g/l、k2hpo4·
3h2o 28g/l,ph为7.0;向转化培养基中再次添加异丙基硫代半乳糖苷至浓度为0.2mmol/l,于37℃摇床中,旋转速度为220r/min,培养3h;
52.s7:用高效液相色谱测定n-乙酰神经氨酸的含量。
53.实施例3
54.一种n-乙酰神经氨酸的制备方法,包括以下步骤:
55.s1:以大肠杆菌e.colik-12mg1655为出发菌,敲除其基因组中的nage基因和manxyz、nanatek基因簇,并将cneub基因和bage基因克隆到ptrcc99a载体,分别加上trc启动子实现2个基因在大肠杆菌中的共表达;
56.s2:敲除与n-乙酰神经氨酸转运载体相关基因nant以及与葡萄糖胺转运相关的磷酸转移酶系统相关基因nage;敲除影响葡萄糖胺6-磷酸合成酶的编码基因glms表达水平的yhbj基因来提高转录水平;
57.s3:克隆s.cerevisiae eby100的葡萄糖胺酰基转移酶的编码基因gnat,通过基因敲除的方法整合到大肠杆菌染色体的naga-nagb基因位点,同时将naga-nagb基因删除,阻断6-磷酸-n-乙酰葡萄糖胺的分解代谢途径,从而催化6-磷酸葡萄糖胺转化生成6-磷酸-n-乙酰葡萄糖胺;将集胞藻age编码基因slr整合到染色体的laci基因位点,同时将laci基因删除;
58.s4:敲除醋酸激酶的编码基因acka与磷酸乙酰转移酶的编码基因pta阻断乙酸合成的其中一条支路;同时敲除乳酸脱氢酶的编码基因idha阻断乳酸生成途径;继续敲除丙酮酸氧化酶的编码基因poxb以阻断乙酸合成的另一条代谢支路;
59.s5:将上述菌种接种于培养基中,培养基中含有胰蛋白胨8g/l、酵母8g/l、氯化钠12g/l和氨苄青霉素0.08g/l,ph为7.0;将培养基接种于摇瓶中,并置于36℃摇床中,旋转速度为250r/min,培养3h,当细胞浓度达到1.2时,添加异丙基硫代半乳糖苷至浓度为0.2mmol/l,诱导胞内酶的表达;将培养基继续振荡培养4h,将菌液收集于冰浴中静置10min,然后于5℃的条件下离心15min,除去上清液,用预冷的生理盐水洗涤菌体2次;
60.s6:将菌体收集浓缩后悬浮于添加了底物n-乙酰葡萄糖胺的转化培养基,转化培养基的成分为甘油14g/l、n-乙酰葡萄糖胺75g/l、硫酸镁2g/l、k2hpo4·
3h2o 30g/l,ph为7.0;向转化培养基中再次添加异丙基硫代半乳糖苷至浓度为0.2mmol/l,于37℃摇床中,旋
转速度为250r/min,培养3h;
61.s7:用高效液相色谱测定n-乙酰神经氨酸的含量。
62.实施例4
63.一种n-乙酰神经氨酸的制备方法,包括以下步骤:
64.s1:以大肠杆菌e.colik-12mg1655为出发菌,敲除其基因组中的nage基因和manxyz、nanatek基因簇,并将cneub基因和bage基因克隆到ptrcc99a载体,分别加上trc启动子实现2个基因在大肠杆菌中的共表达;
65.s2:敲除与n-乙酰神经氨酸转运载体相关基因nant以及与葡萄糖胺转运相关的磷酸转移酶系统相关基因nage;敲除影响葡萄糖胺6-磷酸合成酶的编码基因glms表达水平的yhbj基因来提高转录水平;
66.s3:克隆s.cerevisiae eby100的葡萄糖胺酰基转移酶的编码基因gnat,通过基因敲除的方法整合到大肠杆菌染色体的naga-nagb基因位点,同时将naga-nagb基因删除,阻断6-磷酸-n-乙酰葡萄糖胺的分解代谢途径,从而催化6-磷酸葡萄糖胺转化生成6-磷酸-n-乙酰葡萄糖胺;将集胞藻age编码基因slr整合到染色体的laci基因位点,同时将laci基因删除;
67.s4:敲除醋酸激酶的编码基因acka与磷酸乙酰转移酶的编码基因pta阻断乙酸合成的其中一条支路;同时敲除乳酸脱氢酶的编码基因idha阻断乳酸生成途径;继续敲除丙酮酸氧化酶的编码基因poxb以阻断乙酸合成的另一条代谢支路;
68.s5:将上述菌种接种于培养基中,培养基中含有胰蛋白胨14g/l、酵母5g/l、氯化钠12g/l和氨苄青霉素0.1g/l,ph为7.0;将培养基接种于摇瓶中,并置于36℃摇床中,旋转速度为220r/min,培养3h,当细胞浓度达到1.2时,添加异丙基硫代半乳糖苷至浓度为0.2mmol/l,诱导胞内酶的表达;将培养基继续振荡培养4h,将菌液收集于冰浴中静置10min,然后于3℃的条件下离心15min,除去上清液,用预冷的生理盐水洗涤菌体2次;
69.s6:将菌体收集浓缩后悬浮于添加了底物n-乙酰葡萄糖胺的转化培养基,转化培养基的成分为甘油14g/l、n-乙酰葡萄糖胺85g/l、硫酸镁2g/l、k2hpo4·
3h2o 32g/l,ph为7.0;向转化培养基中再次添加异丙基硫代半乳糖苷至浓度为0.2mmol/l,于37℃摇床中,旋转速度为220r/min,培养3h;
70.s7:用高效液相色谱测定n-乙酰神经氨酸的含量。
71.对比例1
72.与实施例4的制备方法基本相同,不同之处在于制备n-乙酰神经氨酸的制备方法中不添加甘油、n-乙酰葡萄糖胺和异丙基硫代半乳糖苷。
73.对比例2
74.与实施例4的制备方法基本相同,不同之处在于制备n-乙酰神经氨酸的制备方法中不添加甘油。
75.对比例3
76.与实施例4的制备方法基本相同,不同之处在于制备n-乙酰神经氨酸的制备方法中不添加n-乙酰葡萄糖胺。
77.对比例4
78.与实施例4的制备方法基本相同,不同之处在于制备n-乙酰神经氨酸的制备方法
中不添加异丙基硫代半乳糖苷。
79.对比例5
80.采用中国专利文献
″
一种提高发酵产n-乙酰神经氨酸的方法及发酵液(专利申请号为cn201811124497.2)
″
中实施例1中所述方法制备n-乙酰神经氨酸。
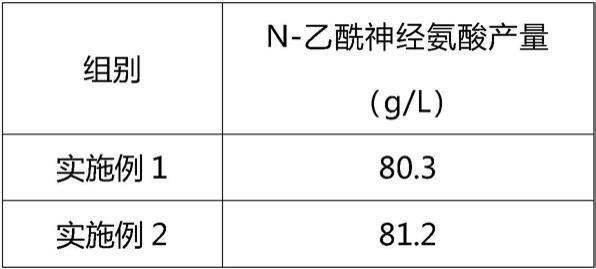
81.按照实施例1-4和对比例1-5的制备n-乙酰神经氨酸,用高效液相色谱测定n-乙酰神经氨酸的含量,其结果见下表。
[0082][0083][0084]
由上表可知:(1)由实施例1-4和对比例5的数据可见,实施例1-4制得的n-乙酰神经氨酸的产量显著高于对比例5制得的n-乙酰神经氨酸的产量,且实施例4为最优实施例。
[0085]
(2)由实施例4和对比例1-4的数据可见,在n-乙酰神经氨酸的制备过程中添加甘油、n-乙酰葡萄糖胺和异丙基硫代半乳糖苷起到了协同作用,协同提高了n-乙酰神经氨酸的产量,这是因为:在转化培养基中添加的n-乙酰葡萄糖胺在cneub基因的催化下与胞内的磷酸烯醇式丙酮酸合成n-乙酰神经氨酸,从而可以提高n-乙酰神经氨酸的产量。并且,在转化培养基中添加诱导剂异丙基硫代半乳糖苷时,重组大肠杆菌能够重新合成cneub基因,使得胞内cneub基因活性维持在较高的水平,从而促进n-乙酰神经氨酸的生成。在转化培养基中添加甘油对细胞膜的流动性和通透性具有促进作用,有利于n-乙酰葡萄糖胺进入胞内和n-乙酰神经氨酸向胞外分泌;并且,以甘油为碳源时,甘油的跨膜运输不依赖磷酸转运系统,因此不消化胞内的磷酸烯醇式丙酮酸,从而使得更多的磷酸烯醇式丙酮酸流向n-乙酰神经氨酸合成的方向,在生物量相同的条件下,提高了n-乙酰神经氨酸的速率和产量。
[0086]
(3)步骤s4中阻断胞内的磷酸烯醇式丙酮酸的降解途径,增加胞内的磷酸烯醇式丙酮酸的积累,敲除醋酸激酶的编码基因acka与磷酸乙酰转移酶的编码基因pta阻断乙酸合成的其中一条支路;同时敲除乳酸脱氢酶的编码基因idha阻断乳酸生成途径;继续敲除丙酮酸氧化酶的编码基因poxb以阻断乙酸合成的另一条代谢支路,进一步提高胞内的磷酸烯醇式丙酮酸的产量,从而提高n-乙酰神经氨酸的产量。
[0087]
以上内容不能认定本发明具体实施只局限于这些说明,对于本发明所属技术领域的普通技术人员来说,在不脱离本发明构思前提下,还可以做出若干简单推演或替换,都应当视为属于本发明由所提交的权利要求书确定的专利保护范围。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1