一种防治樟疫霉的方法
1.本发明属于植物病害防治研究技术领域,更具体地,涉及一种防治樟疫霉的方法。
背景技术:
2.樟疫霉(phytophthora cinnamomi)是一种全世界广泛分布的重要植物病原菌,寄主范围较广,可侵染林木、果树、农作物和园林花卉等多种植物,引起枝枯、腐烂、溃疡、猝倒等症状(hardham,2005)。由于其寄主范围广泛,目前已在68个国家的950多种寄主植物上分离得到(hardham and blackman,2018)。近年来,樟疫霉在澳大利亚西南地区森林迅速传播开来,已经严重威胁到当地自然生态系统以及生物多样性。在美国,疫霉根腐病(phytophthora root rot)是杜鹃的一种破坏性病害,在受侵染的田间和容器苗生产区造成这种苗圃作物的大量损失(weiland et al.,2020)。在地中海地区,樟疫霉引起的橡树大量死亡,严重威胁了地中海混交橡树林(frisullo et al.,2018;g
ó
mez et al.,2020)。因此,寻求有效的樟疫霉防治策略迫在眉睫。但由于樟疫霉属于卵菌,其与真菌在遗传关系上具有较大的差异,因此樟疫霉引起的病害一旦发生,应用传统的杀真菌剂治理效果一直不理想。目前已经开发并应用了几种针对樟疫霉的杀菌剂,例如亚磷酸盐和乙磷铝等(gonz
á
lez et al.,2019;ros
á
rio et al.,2021)。然而,化学农药的频繁使用造成了日益严重的抗药性和环境污染。相比之下,使用微生物拮抗剂的生物控制作为管理植物病原体的化学防治的替代方式引起了相当大的兴趣(jiang et al.,2015)。微生物制剂的其他优点包括控制时间长、耐药性低、易于开发和成本相对较低。因此,生物防治是植物病害防治的最佳策略之一(zheng et al.,2011)。
3.假单胞菌(pseudomonas)是自然界中分布最广泛的革兰氏阴性细菌之一,作为pgpr,假单胞菌可以通过产生和分泌嗜铁素、磷酸酶、植物激素等物质促进植物的生长;对病原菌的抑制作用则主要是通过产生一些抗生素类物质。据报道,荧光假单胞菌(pseudomonas fluorescens)brz63能够在体外抑制立枯丝核菌(rhizoctonia solani)w70、炭疽菌(colletotrichum dematium)k、核盘菌(sclerotinia sclerotiorum)k2291和燕麦镰刀菌(fusarium avenaceum)的生长,而且在盆栽试验中,还能刺激用真菌病原菌处理的油菜幼苗的萌发和生长(chlebek et al.,2020)。绿针假单胞菌(pseudomonas chlororaphis)g05产生的硝吡咯菌素能抑制禾谷镰刀菌(fusarium graminearum)的菌丝生长(huang et al.,2018)。
4.此外,微生物挥发性有机化合物(volatile organic compounds,voc)也引起了更多的关注。voc是具有特定优势的关键生物防治因素,例如良好的膜渗透性、高扩散效率和广泛的拮抗作用(morath et al.,2012)。低浓度的voc可以被感知,因此它们除了直接抑制病原真菌的生长还能诱导植物的系统抗性(cordero et al.,2014;velivelli et al.,2015)。一个成功的生防菌通常需要激活多种代谢物来协同控制病原体,但之前的研究主要集中在单一种类的抗菌代谢物上,而不是多种类型的组合,因此还不能完全揭示其抗菌特性和机制而满足使用需求。
技术实现要素:
5.针对现有技术中存在的不足,本技术所要解决的技术问题是提供一种防治樟疫霉的方法,使用桔黄假单胞菌(p.aurantiaca)st-tj4作为一种有效的生物防治剂来控制樟疫霉。
6.为解决上述技术问题,本技术采用的技术方案为:
7.一种防治樟疫霉的方法,利用桔黄假单胞菌st-tj4的代谢物对樟疫霉进行防治。
8.所述的防治樟疫霉的方法,所述的代谢物为发酵滤液、发酵滤液提取物或氨气。
9.所述的发酵滤液的制备方法为:将菌株st-tj4单菌落接种在20ml kb液体培养基中过夜生长,然后将过夜培养物按5%的比例重新接种到50ml新鲜kb液体培养基中,并在28℃下以200rpm振荡培养3天;发酵液在4℃、8000rpm离心15min,上清液用0.22μm微孔滤膜进行过滤。
10.所述的发酵滤液提取物为采用有机溶剂萃取法分离纯化获得。
11.所述的有机溶剂萃取方法为:将有机溶剂与发酵滤液以1∶1的体积比例加入发酵滤液中,然后剧烈振荡几分钟后收集有机相;将水相加入等体积有机溶剂重新萃取,该过程重复3次;收集所有有机相并使用旋转蒸发器蒸发至干以产生粗提物;粗提物完全溶解在1ml甲醇中并通过0.22μm滤膜过滤后用无菌水稀释备用。
12.所述的有机溶剂选自三氯甲烷。
13.一种防治樟疫霉的方法,利用phenazine-1-carboxylic acid、2-hydroxyphenazine和氨气拮抗樟疫霉。
14.一株防治樟疫霉的桔黄假单胞菌,其分类命名为桔黄假单胞菌(pseudomonas chlororaphis subsp.aurantiaca)st-tj4,保存在中国典型培养物保藏中心,保藏日期为2020年8月19号,保藏编号cctcc no:m2020435,保藏地址为中国武汉武汉大学。
15.所述的桔黄假单胞菌st-tj4在防治樟疫霉中的应用。
16.有益效果:与现有技术相比,本技术采用不同拮抗方式、观察菌体形态、研究代谢产物和确定拮抗物质等方法来明确桔黄假单胞菌st-tj4对樟疫霉的拮抗机制;结果表明,桔黄假单胞菌st-tj4能够明显抑制樟疫霉菌丝的生长,导致菌丝畸形,破坏内部细胞结构。此外,菌株st-tj4还能够破坏菌丝细胞膜通透性,影响细胞内容物的合成。超高效液相色谱-质谱(uplc-ms)和气相色谱-质谱(gc-ms)分析发现,吩嗪类化合物和2-undecanone是主要的抑菌化合物。并且st-tj4菌株产生的挥发性氨也是抑制病原体生长的原因之一。可见,桔黄假单胞菌st-tj4可通过多重机制抑制樟疫霉,具有作为生物防治樟疫霉引起的各种植物病害的潜力。
附图说明
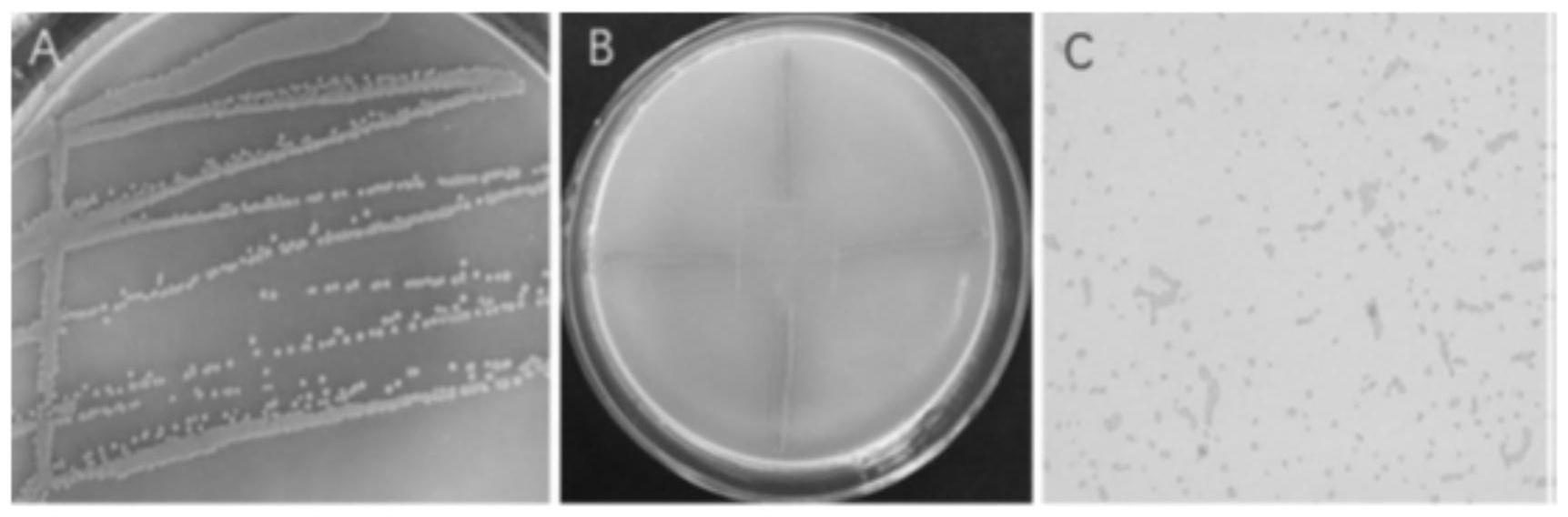
17.图1是菌株st-tj4形态学鉴定结果图:a:桔黄假单胞菌菌落形态;b:需氧量测试结果;c:革兰氏染色;
18.图2是桔黄假单胞菌st-tj4对樟疫霉生长的影响结果图;a:ck;b:平板对峙;c:发酵滤液;d:voc;
19.图3是桔黄假单胞菌st-tj4产生的发酵滤液和vocs对樟疫霉侵染杜鹃叶片的防治效果结果图;
20.图4是菌株st-tj4对樟疫霉菌丝形态和超微结构的影响结果图;a、d:蔡司显微镜下菌丝形态;b、e:扫描电镜下菌丝形态;c、f:透射电镜下菌丝内部结构;
21.图5是桔黄假单胞菌st-tj4发酵滤液对樟疫霉细胞膜通透性和细胞内含物的影响结果图;a:伊文思蓝染色,左图为ck,右图为st-tj4发酵滤液处理;b:电导率;c:还原性糖含量;d:可溶性蛋白含量;
22.图6是st-tj4菌株培养滤液的提取物通过不同方法对樟疫霉的拮抗作用结果图;a:酸沉淀产生的环脂肽(clps)及不同有机溶剂萃取;b:tlc分析;
23.图7是st-tj4菌株产生的phenazine-1-carboxylic acid(a)和2-hydroxyphenazine(b)质谱图;
24.图8是桔黄假单胞菌st-tj4产氨能力检测及对培养基ph的改变结果图;a:st-tj4产氨能力;b:溴甲酚紫指示剂显示ph的改变;
25.图9是不同ph下樟疫霉生长情况和2-undecanone的抑制效果图;a:未碱化培养基;b:碱化培养基;c:未碱化培养基+2-undecanone;d:碱化培养基+2-undecanone处理。
具体实施方式
26.下面结合具体实施例对本技术做进一步的说明。
27.以下实施例所使用的材料如下:
28.1)樟疫霉,是在南京林业大学校园内从受感染的杜鹃根部中分离出来,经鉴定为樟疫霉。
29.2)从杨树根际分离得到菌株st-tj4,lb平板上的st-tj4菌落呈薄、扁平、橙色、不透明、圆形,边缘光滑,直径约1.2-3mm。需氧量测试表明st-tj4是一种需氧细菌。它在盖玻片外生长良好,但在盖玻片下几乎不生长。革兰氏染色显示st-tj4是一种革兰氏阴性细菌(图1)。通过16s rdna(seq id no.1)构建系统发育树,确定菌株st-tj4的种属,命名为桔黄假单胞菌(pseudomonas chlororaphis subsp.aurantiaca)st-tj4。并保存在中国典型培养物保藏中心,保藏日期为2020年8月19号,保藏编号cctcc no:m2020435,保藏地址为中国武汉武汉大学。
30.实施例1桔黄假单胞菌st-tj4对樟疫霉的拮抗活性
31.1、桔黄假单胞菌st-tj4抑制菌丝体生长的测定
32.从lb琼脂培养基中挑取一个桔黄假单胞菌st-tj4的单菌落,接种到lb肉汤培养基中,并在28℃下200rpm振荡孵育12小时。取对数生长期(od
600
=1.6)的桔黄假单胞菌st-tj4的10μl细菌悬液和生长4天的樟疫霉菌落边缘的单个5nm直径菌饼在pda培养基共培养5天,试验重复三次。
33.为了评估发酵滤液的抑菌活性,将菌株st-tj4单菌落接种在20ml kb液体培养基中并过夜生长,然后将过夜培养物按5%的比例重新接种到50ml新鲜kb液体培养基中,并在28℃下以200rpm振荡培养3天。发酵液在4℃、8000rpm离心15min,上清液用0.22μm微孔滤膜进行过滤。取活跃生长的樟疫霉菌落边缘的5mm直径的菌饼转移到v8平板的中心。将100μl上清液样品倒入距樟疫霉2.5cm的两个孔中,并设置培养基空白对照。在25℃培养5天。试验重复三次。
34.为评估细菌voc的抑菌活性,将st-tj4菌株的悬浮液(100μl)均匀涂布在lb平板
上,使用取自活跃生长的菌落边缘的5mm菌饼接种到另一个培养皿中。将带有樟疫霉的培养皿倒置在带有细菌的培养皿的顶部,接着用m封口膜密封,以避免voc从顶部空间逸出。密封后在25℃下培养5天。试验重复三次。
35.在平板对峙实验中,菌株st-tj4能够显著抑制樟疫霉的生长(图2a,b)。微生物对病原体的控制涉及多种代谢物,因此接下来测试st-tj4菌株两个部分的抗真菌活性,即发酵滤液和挥发性化合物。结果表明,与st-tj4菌株发酵滤液共培养5d后的樟疫霉可见明显的抑菌圈(图2c)。采用皿皿对扣方式,用菌株st-tj4释放的vocs处理后,在培养基表面生长的气生菌丝被明显抑制,仅在培养基内还有稀疏的菌丝生长(图2d)。两者的抑制作用都不如平板对峙实验时效果明显。
36.2、桔黄假单胞菌st-tj4对樟疫霉侵染杜鹃叶片的防治效果
37.除了在体外试验中测试桔黄假单胞菌st-tj4对樟疫霉的拮抗作用外,本实施例还测试了st-tj4菌株对樟疫霉引起的杜鹃叶片病害生物防治效果。取健康杜鹃叶片,无菌水清洗,用脱脂棉蘸取75%乙醇清洗表面,无菌水清洗3次后自然晾干。按照上述获取桔黄假单胞菌st-tj4的发酵滤液,将健康杜鹃叶片浸入st-tj4菌株发酵滤液15秒,以浸泡无菌水15秒的处理为对照。为了测定st-tj4菌株释放的voc的活性,采用皿皿对扣方式进行。将st-tj4菌株的悬浮液(100μl)均匀涂布在lb平板上,将一片健康杜鹃叶片置于另一培养皿中。
38.每一健康杜鹃叶片中心处用灭菌的梅花针刺伤叶片。上述处理组在叶片伤口处接种直径5mm的病原菌饼,对照组在伤口处接种同样大小的无菌琼脂块。处理组及对照组各接种5片叶,然后置于恒温光照培养箱(mlr-352h-pc,panasonic,japan)内保湿培养,相对湿度为75%,在25℃下,12小时光照和12小时黑暗生长5天后,测量叶片的病斑面积。试验重复三次。
39.结果如图3所示,在25℃下处理5天后,与对照相比,对于用st-tj4菌株发酵滤液或挥发性气体处理的叶片,病斑面积分别限制在0.31
±
0.13em2和0.76
±
0.21cm2,而接种樟疫霉叶片的病斑面积扩大至2.48
±
1.05cm2。而在发酵滤液与挥发性气体共同作用下,病斑面积比单独采用一种方式处理要更小,为0.24
±
0.51cm2。以上结果表明,樟疫霉引起的病斑面积的扩展被桔黄假单胞菌st-tj4发酵滤液和挥发性气体有效抑制,且st-tj4发酵滤液的抑制作用更为明显。
40.3、菌丝体的蔡司、扫描和透射显微镜观察
41.方法如上述,共培养5天后,在菌株st-tj4对峙的樟疫霉菌落边缘切取0.5
×
0.5cm的样品,以正常生长的樟疫霉菌丝为对照,在蔡司显微镜下观察。将每个处理的菌丝体用3%戊二醛和0.1m磷酸盐缓冲液(ph 7.2)在4℃下固定24小时(guevara-avendano et al.,2019)。将样品用无菌水洗涤3次,每次10分钟,并使用乙醇梯度(30、50、80、90和100%)脱水,然后,在临界点干燥器(k850,emitech,uk)中用液态co2喷涂金箔15分钟。在20kv下通过扫描电子显微镜观察(quanta 200,fei,usa)。
42.将前面经乙醇脱水的样品进行包埋、固化后,用3%醋酸铀-枸橼酸铅双染色,然后嵌入taab spurr树脂中。由超薄切片机切片,并将样品在透射电子显微镜(jem-1400,jeol,japan)下观察。每个样品至少检查20个切片。
43.大多数生物防治剂能够抑制菌丝的生长,来控制植物病原体。从光学显微镜可以看出,对照组樟疫霉菌丝细长,生长密集,分支较少;而用p.aurantiaca st-tj4处理的菌丝
表现出畸形且生长稀疏,出现肿胀的囊状结构,分支增多(图4a,d)。从扫描电镜可以看出,用p.aurantiaca st-tj4处理的菌丝出现明显的皱缩和塌陷,而对照组菌丝表面整齐、丰满、光滑(图4b,e)。进一步采用透射电子显微镜下观察菌丝的内部变化可以发现,对照组菌丝内结构完整,线粒体和液泡形态正常。而经p.aurantiaca st-tj4处理后细胞内部结构被破坏,线粒体消失,液泡增多破裂(图4c,f)。因此,p.aurantiaca st-tj4会严重改变樟疫霉的菌丝形态,破坏其细胞内部结构。
44.实施例2樟疫霉细胞膜通透性和细胞内含物的测定
45.在20mlv8培养液中接入1块直径为5mm樟疫霉菌饼,于25℃下静置培养4天。将得到的新鲜菌丝用超纯水洗涤3次后放入10ml离心管中,加入4ml培养72h的st-tj4无菌发酵滤液;以kb培养基作为对照。使用电导仪(dds-307,中国)测量样品的电导率。之后再将菌丝煮沸5min,测量最终电导率,并计算相对电导率(luo et al.,2015),实验重复三次。计算公式如下:
46.相对电导率(%)=各时间电导率/最终电导率
×
100
47.再次获得不同处理的菌丝,并用无菌超纯水反复洗涤,用无菌滤纸擦干,称重并记录。之后,以菌丝体与冷tris-hcl缓冲液(0.05mol/l,ph 7.5,4℃)的比例为1∶5(v∶w)研磨菌丝体。匀浆在4℃下以10000r/min离心15分钟。上清液用于测定可溶性蛋白(dai et al.,2021)和还原性糖(dai et al.,2017)。试验重复三次。
48.伊文思蓝染色法(shehata et al.,2016)检测樟疫霉细胞膜的完整性:在生长4天的樟疫霉中加入4ml培养72h的st-tj4菌株的发酵滤液,25℃下静置处理12h,以磷酸缓冲液浸泡处理的菌丝体为对照;12h后倒掉发酵滤液,加入0.05%的伊文思蓝过夜染色,取出菌丝丛,用无菌水充分洗涤脱色,待脱色完全,将菌丝丛置于蔡司荧光显微镜下(zeiss microscope system standard 16;carl zeiss ltd;germany)观察。
49.为了进一步探究st-tj4发酵滤液对菌丝造成的破坏,本实施例测定了其发酵滤液对樟疫霉细胞膜通透性和细胞内含物的影响。如图5所示,st-tj4发酵滤液处理的菌丝被伊文思蓝染成蓝色(图5a),且菌丝的电解质渗漏率始终高于对照组(图5b),表明菌株st-tj4发酵滤液处理影响了樟疫霉细胞膜的通透性。
50.此外,随着处理时间的增加,菌株st-tj4发酵滤液处理的菌丝体中还原性糖和可溶性蛋白的含量均始终低于对照组(图5c-d)。以上结果表明,经菌株st-tj4发酵滤液处理的樟疫霉菌丝细胞膜受损,细胞内含物外泄。
51.实施例3桔黄假单胞菌st-tj4发酵滤液中脂肽和有机提取物的抑菌活性
52.1、脂肽类代谢物
53.将收集的发酵滤液用6mol/l的hcl将ph调至2,4℃保存过夜。通过离心(4℃下以8,000r/min离心30分钟)从培养液中获得粗提取物,并将沉淀物溶解在甲醇中。再次离心后,收集上清液并用冷冻干燥机(alpha 1-2 ld,christ,germany)进行真空干燥,收集残留物。然后将收集的残余物溶解在甲醇中,真空干燥甲醇提取物,固体残渣重悬于10%甲醇水溶液中,经cleanert s c18管过滤,获得含脂肽混合物的滤液。
54.2、有机溶剂萃取
55.将溶剂(石油醚(pe)、三氯甲烷(tcm)、正丁醇(nba)、乙酸乙酯(eac)和二甲亚砜(dmso))等比例加入发酵滤液中,然后剧烈振荡几分钟后收集有机相。将水相加入等体积有
机溶剂重新萃取,该过程重复3次。收集所有有机相并使用旋转蒸发器(n-1300,eyela,japan)蒸发至干以产生粗提物。粗提物完全溶解在1ml甲醇中并通过0.22μm滤膜过滤后用无菌水稀释10倍备用。
56.使用牛津杯法评估提取物对樟疫霉生长的抑制作用。将新鲜的樟疫霉菌饼接种在琼脂培养基中,在距菌丝体2cm处放置三个牛津杯,并在其中加入100μl含有脂肽类代谢物和有机溶剂萃取粗提物的甲醇水溶液,对照中加入1ml甲醇和9ml无菌水的混合物100μl。试验重复三次。
57.3、通过uplc-hrms对有机提取物进行化学分析
58.从三氯甲烷提取中获得的粗提取物使用0.22μm尼龙过滤器(syringe filters;millipore)进行过滤。用hplc(alliance e2695,waters,usa)进行分离纯化。对分离物进行uplc-ms分析以表征次级代谢物,所用仪器为uhplc(lc-30,shimadzu,japan)连接质谱仪(tripletof5600+,sciex
tm
,usa)。在35℃下,将滤液注入shimadzu inersustain c18色谱柱中(2μm;100x2.1mm),流速为1ml/min。流动相为乙腈:0.1%ch3cooh-h2o=5%:95%。质谱采用电喷雾电离(esi)正离子和负离子模式进行检测。esi源条件如下:ion source gas1(gas 1):50,ion source gas2(gas 2):50,curtain gas(cur):25,source tempreture:500℃(正离子)和450℃(负离子),ion sapary voltage floating(isvf)5500v(正离子)和4400v(负离子),tof ms scan range:100-1200da,product ion scan range:50-1000da,tof ms scan accumulation time 0.2s,product ion scan accumulation time 0.01s,二级质谱采用information dependent acquisition(ida)获得,并采用high sensitivity模式,declustering potential(dp):
±
60v,collision energy:35
±
15ev。
59.将lc-ms的原始数据导入ms-dial 4.20(ms-dial:data independent ms/ms deavolution for comprehensive metabolome analysis.(nature methods,12,523-526,2015)软件进行预处理,包括峰提取、去噪音、反卷积,峰对齐,导出csv格式的三维数据矩阵(原始数据矩阵)。将提取的峰信息与数据库进行比对,对massbank,respect,gnps(共14951个records)三个库进行全库检索。这个三维矩阵包括的信息有:样品信息、保留时间、质核比和质谱响应强度(峰面积)。
60.图6结果显示,菌株st-tj4上清液酸沉淀的脂肽类物质没有抑菌作用,而以三氯甲烷、石油醚为萃取剂时,樟疫霉菌丝的生长受到明显抑制。表明st-tj4菌株发酵滤液中的抑菌物质易溶于低极性有机溶剂。为了分离和表征抑菌化合物,通过柱层析从三氯甲烷提取物中获得了在tlc分析中呈现单个点的两个分层,并通过hplc纯化。
61.通过uplc-hrms分析鉴定出两种抑菌物质(图7)。离子分离峰位于m/z 247.0482[m+h]
+
,得到分子式为c
13
h8n2o2;另一个离子分离峰位于197.0714,得到分子式为c
12hs
n2o。基于质谱,这两种物质分别鉴定为2-hydroxyphenazine和phenazine-1-carboxylic acid。
[0062]
实施例4氨和ph值对樟疫霉生长的影响
[0063]
微生物在20ml lb液体培养基中生长,在28℃和200rpm下孵育48小时后,将250μl nessler试剂(cas编号7783-33-7,carlo erba reagents)添加到另一个试管中的4ml培养物等分试样中。培养物从黄色到红褐色的发展表明产生了氨。利用溴甲酚紫指示剂检测氨对培养基ph的改变。将1g琼脂和6mg溴甲酚紫加入100ml蒸馏水中,加热溶解,混匀,冷却至
约50℃,调节ph至溶液颜色呈黄色。将配制好的含指示剂的琼脂均匀快速地倒在培养皿上。琼脂凝固后,将培养皿置于st-tj4菌悬液(100μl)均匀涂布的lb平板上。平板用m密封,以避免vocs从顶部空间逸出。密封后在28℃下避光培养。在3天后,观察含有溴甲酚紫指示剂的琼脂平板的颜色变化。
[0064]
为了评估氨对樟疫霉生长抑制的作用,将st-tj4菌株和樟疫霉与磷酸(h3po4)共培养。磷酸能够与氨反应形成磷酸铵盐(ryden,1984),从而消除氨对樟疫霉的影响。试验以一个90mm培养皿与一个90mm二分皿进行。在二分皿中分别倒入lb培养基和无菌水,在lb培养基中涂布50μl st-tj4菌株悬浮液,将接种5mm樟疫霉菌饼的培养皿置于二分皿上,用m密封,而未接种st-tj4菌株的平板作为对照。实验组将二分皿中的无菌水换成1m磷酸。该试验设置一式三份进行,并在25℃下孵育4天。后将一张石蕊试纸放在琼脂表面以检查其ph值。
[0065]
为了检测氨是直接抑制樟疫霉的生长还是通过改变ph间接抑制,及ph的改变与挥发性气体纯品共同作用对樟疫霉生长的抑制作用,使用0.1m磷酸钾缓冲液(16.37g/l k2hpo4+0.82g/l kh2po4)制备ph为8.5的琼脂培养基。采用皿皿对扣overlapping plate(op)方式进行评估挥发性气体纯品的作用,在另一个平板上添加100μl 2-undecanone。该试验设置一式三份进行,并在25℃下孵育4天。
[0066]
使用nessler试剂在初步测试菌株st-tj4在培养基中产生的氨。在培养48h的菌液中加入nessler试剂,可以明显观察到黄褐色沉淀,说明菌株st-tj4能够产生氨(图8a)。溴甲酚紫指示剂也显示菌株st-tj4产生的挥发性nh3能够使培养基碱化(图8b)。
[0067]
试验通过0.1m磷酸钾缓冲液制备ph为8.5的琼脂培养基。与对照相比,琼脂培养基ph的升高对樟疫霉生长的影响较小,因此菌株st-tj4菌株产生的氨能够直接抑制樟疫霉,而并不是通过改变ph来间接起作用(图9)。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1