一种mTOR抑制剂及其应用
一种mtor抑制剂及其应用
技术领域
1.本发明涉及生物技术领域,尤其是涉及一种mtor抑制剂及其应用。
背景技术:
2.mtor(mechanistic mammalian target of rapamycin),是哺乳类等动物的参与细胞内信号传递的蛋白激酶的一种,其在蛋白质转译、细胞自噬、免疫调节、细胞骨格调节、基因转录等发挥重要作用,与细胞成长、分裂及死亡等基本生命现象密切相关。mtor,已知是作为表现出强效免疫抑制作用及抗癌作用的雷帕霉素的细胞内标靶而被发现的蛋白质,且是分子量29万的巨大丝氨酸-苏氨酸激酶。
3.目前mtor抑制剂已有广泛研究,但作为化疗药物,其适用均存在不同的缺陷。mtor抑制剂的发展大致可分为三种阶段,首先是最早发现的雷帕霉素,但因为它们仅部分阻断4e-bp依赖性翻译途径,并且不能抑制mtorc2
–
akt调节的促生存途径,同时仅对于肾癌细胞有疗效,因此临床上治疗效果并不理想。第二代药物(torin1,pp242,ku-0063794)主要依靠与atp竞争占据激酶活性位点抑制mtorc1和mtorc2底物发挥作用。但早期临床数据表明催化mtor抑制剂会出现耐药性。第三代药物主要是依靠联合用药,使用pi3k和mtor双重抑制发挥作用。而利用蛋白抑制mtor,进而治疗肿瘤的用药策略至今尚未被开发。
技术实现要素:
4.本发明的目的在于克服上述现有技术的不足之处而提供一种mtor抑制剂及其应用,本发明通过新型邻近标记技术筛选方法,找到了atp6ap1蛋白的c端片段(c tail)作为rheb的鸟苷酸交换因子(gef)可激活rheb,在此基础上,将c末端30个氨基酸中第14、17、18位的氨基酸进行突变,其突变体可结合并阻断内源性atp6ap1对rheb的抑制,进而抑制mtorc1信号通路。
5.为实现上述目的,本发明采取的技术方案为:
6.第一目的,本发明提供了一种mtor抑制剂,所述mtor抑制剂为atp6ap1蛋白的c末端30个氨基酸中的第14、17、18位的天冬氨酸均突变为丙氨酸所获得,30个氨基酸的序列如seq id no:1所示。
7.本发明通过新型邻近标记技术筛选方法,找到了atp6ap1蛋白,将c末端30个氨基酸中的第14、17、18位的天冬氨酸均突变为丙氨酸,获得抑制rheb-mtorc1的短肽(mtor抑制剂),抑制mtorc1信号通路。
8.由于mtorc1、mtorc2复合物均含有mtor蛋白,而rheb为mtorc1特有。传统直接靶向mtor的抑制剂,能同时抑制mtorc1、mtorc2下游信号,而本发明获得的mtor抑制剂并不影响mtorc2下游。
9.更优选地,mtor抑制剂为mtorc1抑制剂。
10.第二目的,本发明提供了atp6ap1蛋白突变体在抑制mtor信号通路活性中的应用,其特征在于,atp6ap1蛋白突变体为atp6ap1蛋白的c末端30个氨基酸中的第14、17、18位的
天冬氨酸均突变为丙氨酸所获得,30个氨基酸的序列如seq id no:1所示。
11.作为本发明所述atp6ap1蛋白突变体在抑制mtor信号通路活性中的应用的优选实施方式,mtor信号通路包括mtorc1信号通路和mtorc2信号通路。
12.第三目的,本发明提供了一种筛选mtor抑制剂的方法,包括以下步骤:
13.1)构建生物素连接酶与rheb融合表达载体,将载体转入宿主细胞内,药物筛选稳定表达rheb蛋白的细胞株,同时构建对照细胞株;
14.2)向稳定表达rheb蛋白的细胞株中加入胰岛素和生物素进行刺激,向对照细胞株中加入生物素刺激,然后进行质谱检测,分析蛋白;
15.3)在有/无进行胰岛素刺激条件下,分别将稳定表达rheb蛋白的细胞株中的富集蛋白扣除(扣除表示为将两组数据中重叠的蛋白去掉,以达到去除对照组干扰的目的)对照细胞株中的富集蛋白,获得有/无进行胰岛素刺激条件下的rheb邻近蛋白,分析rheb邻近蛋白,经鸟嘌呤核苷酸交换因子筛选条件,得atp6ap1蛋白;将atp6ap1蛋白c末端30个氨基酸中的第14、17、18位的天冬氨酸均突变为丙氨酸,获得mtor抑制剂。
16.更优选地,鸟嘌呤核苷酸交换因子筛选条件包括以下条件:
17.(1)gef(鸟嘌呤核苷酸交换因子)与rheb存在体内外相互作用;
18.(2)过表达gef后可以激活mtorc1下游信号通路;
19.(3)敲低或敲除gef后,mtorc1下游信号通路活性受到大幅度抑制;
20.(4)体外实验证明gef可以直接促进rheb完成gdp-gtp转换过程。
21.本发明通过新型邻近标记技术筛选得到rheb蛋白激活剂-atp6ap1蛋白,atp6ap1蛋白的c末端30个氨基酸可以直接靶向激活rheb。
22.作为本发明所述筛选mtor抑制剂的方法的优选实施方式,胰岛素的浓度为0.9μm,生物素的浓度为50μm。
23.胰岛素、生物素选用上述浓度时,使得生长的细胞株可以较好的发挥功能。
24.作为本发明所述筛选mtor抑制剂的方法的优选实施方式,刺激的时间为15min。采用上述刺激时间时,可以较为准备的捕捉到gef蛋白。
25.作为本发明所述筛选mtor抑制剂的方法的优选实施方式,生物素连接酶包括生物素连接酶xxid,生物素连接酶与rheb融合表达载体为haflag-xxid-rheb。本发明中提到的生物素连接酶不仅限于本发明,还可以为本领域中常规使用的生物素连接酶。
26.优选地,稳定表达rheb蛋白的细胞株为hela-xxid-rheb稳定表达细胞株,对照细胞株为hela-xxid稳定表达细胞株。
27.更优选地,宿主细胞包括hela细胞,还可以为其他细胞。
28.第四目的,本发明提供了一种抑癌药物,包含上述mtor抑制剂,以及医学上可接受的载体。
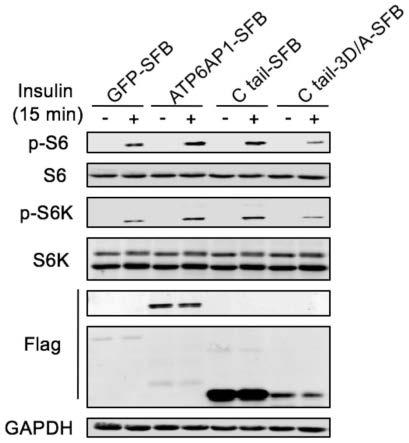
29.第五目的,本发明提供了一种抗衰老药物,包含上述mtor抑制剂,以及医学上可接受的载体。
30.第六目的,本发明提供了上述mtor抑制剂在制备提高由于atp6ap1蛋白敲低导致的磷酸化药物中的应用。
31.第七目的,本发明提供了上述mtor抑制剂在制备抑制mtorc1信号通路的活性而预防和/或治疗癌症、以及抗衰老药物中的应用,所述癌症包括乳腺癌或胰腺癌。
32.与现有技术相比,本发明具有以下有益效果:
33.本发明提供了一种mtor抑制剂及其应用,利用新型邻近标记技术筛选方法找到了atp6ap1蛋白的c端片段(c tail)作为rheb的鸟苷酸交换因子(gef)可激活rheb,在此基础上,将c末端30个氨基酸中第14、17、18位的天冬氨酸均突变为丙氨酸,获得新的mtor抑制剂,该mtor抑制剂可结合并阻断内源性atp6ap1对rheb的抑制,进而抑制mtorc1信号通路,相比于现存的mtor抑制剂,本发明获得的mtor抑制剂特异性较高,更能有针对性的发挥抑制剂作用。本发明获得的mtor抑制剂在治疗糖尿病、肿瘤中具有广泛的药用前景。
附图说明
34.图1为hela-xxid-rheb稳定表达细胞株和hela-xxid稳定表达细胞株构建过程图及质谱检测图;
35.图2为实施例1中come/go/stay组的差异倍数分析图;
36.图3为过表达mtor抑制剂抑制mtor信号通路活性的示意图;
37.图4为回补过表达mtor抑制剂抑制mtor信号通路活性的示意图。
具体实施方式
38.为更好的说明本发明的目的、技术方案和优点,下面将结合附图和具体实施例对本发明作进一步说明。
39.在以下实施例中,所使用的实验方法如无特殊说明,均为常规方法,所用的材料、试剂等,如无特殊说明,均可从商业途径得到。
40.实施例1、一种筛选mtor抑制剂的方法
41.一种筛选mtor抑制剂的方法,包括以下步骤:
42.1)构建生物素连接酶(xxid)与rheb融合表达载体(haflag-xxid-rheb),将载体转入hela细胞内,药物筛选hela-xxid-rheb稳定表达细胞株,同时构建对照细胞系hela-xxid稳定表达细胞株,利用含10%透析血清培养基培养;实验前16小时,使用无血清培养基培养,规避胰岛素的干扰。
43.2)然后向hela-xxid-rheb稳定表达细胞株中加入胰岛素(0.9μm)和生物素(50μm)进行刺激,刺激15min,向hela-xxid稳定表达细胞株中加入生物素(50μm)刺激,不加入胰岛素,刺激15min,对样品进行裂解,提取蛋白后,利用链霉亲和素下拉,富集生物素化的蛋白,对链霉亲和磁珠进行还原烷基化,胰酶酶解,脱盐,最后将样品真空干燥后干粉保存,进行质谱检测(如图1所示),分析蛋白。
44.3)在有/无进行胰岛素刺激条件下,分别将hela-xxid-rheb稳定表达细胞株中的富集蛋白扣除hela-xxid稳定表达细胞株中的富集蛋白,获得有/无进行胰岛素刺激条件下的rheb邻近蛋白,将这部分富集蛋白(rheb邻近蛋白)进一步分析,可分为come/go/stay组,即刺激后出现蛋白组/刺激前后稳定存在组/刺激后离开蛋白组(如图2所示),come组包括atp6ap1蛋白、reeps蛋白、sc22b蛋白,go组包括vamp3蛋白、scd蛋白,stay组包括rab7a蛋白、tsc1蛋白,由于gef蛋白应该在胰岛素刺激后出现,因此将come组蛋白作为gef候选蛋白进行验证。
45.4)对质谱表单的候选蛋白进行验证,rheb的gef候选蛋白需满足以下条件:
46.(1)gef(鸟嘌呤核苷酸交换因子)与rheb存在体内外相互作用;
47.(2)过表达gef后可以激活mtor下游信号通路;
48.(3)敲低或敲除gef后,mtor下游信号通路活性受到大幅度抑制;
49.(4)体外实验证明gef可以直接促进rheb完成gdp-gtp转换过程。
50.通过筛选,发现atp6ap1蛋白(c末端30个氨基酸,tyglhmilslktmdrfddhkgptisltqiv)可以很好的满足上述条件,证明atp6ap1蛋白是rheb的潜在gef。
51.5)将atp6ap1蛋白c末端30个氨基酸中的第14、17、18位的天冬氨酸均突变为丙氨酸,获得mtor抑制剂。
52.通过ncbi对c末端进行序列保守性分析,发现c末端(30aa)十分保守,在hela细胞系中敲低atp6ap1并过表达atp6ap1全长蛋白(4μg),在胰岛素刺激15min后,发现可以挽救由于atp6ap1敲低而导致的mtor信号通路被抑制的现象(mtor底物s6k及其下游s6的磷酸化水平明显降低);而过表达c末端3d/a(4μg)突变形成的mtor抑制剂(c末端30个氨基酸中的第14、17、18位的天冬氨酸均突变为丙氨酸)则会抑制mtor信号通路的活性(参考图3)。同时,在hela细胞系中过表达atp6ap1(2μg);c末端(2μg)可以明显提升mtor信号通路活性;回补c末端3d/a(8μg)突变形成的mtor抑制剂(c末端30个氨基酸中的第14、17、18位的天冬氨酸均突变为丙氨酸),并不能挽救atp6ap1敲低导致的磷酸化降低的现象,使c末端行使gef功能的能力大幅度降低(参考图4)。
53.本发明利用新型邻近标记技术筛选方法找到了atp6ap1的c端片段(c tail)作为rheb的鸟苷酸交换因子(gef)可激活rheb。在此基础上,将c末端30个氨基酸中第14、17、18位的天冬氨酸均突变为丙氨酸,获得新的mtor抑制剂,该mtor抑制剂可结合并阻断内源性atp6ap1对rheb的抑制,进而抑制mtorc1信号通路,这是首个发现可抑制rheb-mtorc1的短肽。由于mtorc1、mtorc2复合物均含有mtor蛋白,而rheb为mtorc1特有,传统直接靶向mtor的抑制剂,能同时抑制mtorc1、mtorc2下游信号,而抑制rheb-mtorc1的短肽并不影响mtorc2下游。相比于现存的mtor抑制剂,本发明获得的mtor抑制剂特异性较高,更能有针对性的发挥抑制剂作用。此外,mtor信号通路抑制剂在治疗糖尿病、肿瘤中具有广泛的药用前景。
54.最后所应当说明的是,以上实施例仅用以说明本发明的技术方案而非对本发明保护范围的限制,尽管参照较佳实施例对本发明作了详细说明,本领域的普通技术人员应当理解,可以对本发明的技术方案进行修改或者等同替换,而不脱离本发明技术方案的实质和范围。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1