靶向5/B/6金属β-内酰胺酶的2’-全修饰XNA适配体及其获取方法
本发明涉及生物,具体为利用定向进化后的sfm4-6聚合酶在selex每一轮筛选当中转录dna文库得到2’-全修饰的非天然核酸适配体文库,并用该文库筛选靶向5/b/6金属β-内酰胺酶的2’-全修饰非天然核酸适配体,最终得到三条70nt的靶向5/b/6金属β-内酰胺酶的2’-全修饰非天然核酸适配体。
背景技术:
1、β-内酰胺环类抗生素药物如青霉素,头孢等是生活中最常见的抗菌类药物之一,随着这类抗生素的广泛使用,细菌体内进化出了相应的耐药机制。其中,细菌产生耐药性最常见的机制是细菌体内产生的β-内酰胺酶对β-内酰胺环类抗生素进行水解,β-内酰胺环发生开环,从而使β-内酰胺环类抗生素失活(如图1所示为β-内酰胺酶使抗生素开环失活机制)。β-内酰胺酶大致可以分为a、b、c、d四类,其中a、c、d类均以丝氨酸残基作为活性部位,而b类为金属β-内酰胺酶mbls(metallo-β-lactamases,mbls),该类β-内酰胺酶需要以一个或两个金属锌离子作为活性中心。据文献报道,产生mbls的细菌数量正在增加,并在耐药菌中占了很大的比例。
2、适配体是基于selex技术(即指数富集的配体系统进化技术)从随机寡核苷酸文库中筛选获得的对靶物质具有很高特异性与亲和力的单链核苷酸序列,相比化学小分子药物,适配体药物具极高的亲和性和特异性识别作用。与抗体相比,核酸适配体具有无免疫原性、不需要使用动物产生、可以直接化学合成、筛选周期短、可反复变性、易于标记等优点。在过去的三十年,核酸适配体越来越受欢迎,被视为可用作亲和试剂,分子成像,诊断,甚至是治疗药物的潜在分子。但是,由于ssdna/rna的不稳定性,在适配体进入细菌体内后很快被核酸酶降解而失去活性,因此增强适配体的稳定性成为热点研究方向。大量的研究证明:rna核糖2’位的羟基被甲氧基(2’-ome)或氟(2’-f)修饰取代的核酸链可以显著提高核酸分子的稳定性。因此,如果可以建立一个2’-f,2’-ome修饰的单链核酸文库,再从中筛选出可以靶向靶标蛋白的全修饰核酸适配体,就可以得到既对核酸酶稳定又具有结合活性的核酸适配体。
3、xna(这里主要指以2’-f或2’-ome修饰的核苷酸为底物,通过酶法或者化学合成的核酸链)适配体是由非天然核苷酸单体通过3’-5’磷酸二酯键连接,在一定的缓冲液下,通过自身卷曲和折叠,形成特定的空间二级结构,并对靶标分子具有高亲和力和高特异性的核酸单链。与天然的dna或rna适配体相比,xna适配体具有更强的稳定性以及耐核酸酶降解能力,这使得xna核酸适配体替代天然的dna或rna适配体成为可能。
4、由于聚合酶对底物的特异性,天然的聚合酶难以识别修饰的核苷酸。但是随着聚合酶定向进化技术的发展,使得聚合酶识别非天然核苷酸底物成为可能,实验室酶法合成非天然核酸序列变得更加容易。本专利通过使用定向进化的taq dna聚合酶stoffel片段stoffel fragment(sf)突变株——sfm4-6转录出糖环2’-c不同修饰的xna文库,利用selex技术对得到的文库与固定好的靶标蛋白蜡状芽孢杆菌5/b/6中的金属β-内酰胺酶进行孵育结合。通过不同的洗脱条件,将未结合的xna序列除掉,最后用变性条件洗脱得到结合靶标蛋白的序列。然后再通过反转录,扩增,转录构建后续筛选的文库。经过多轮筛选,最终得到靶向蜡状芽孢杆菌5/b/6中的金属β-内酰胺酶的xna适配体。最后通过emsa测试xna适配体对金属β-内酰胺酶5/b/6的结合活性,证明筛选到的适配体均具有较高的结合5/b/6中的金属β-内酰胺酶的活性,未来有开发为检测分子的潜在应用。
技术实现思路
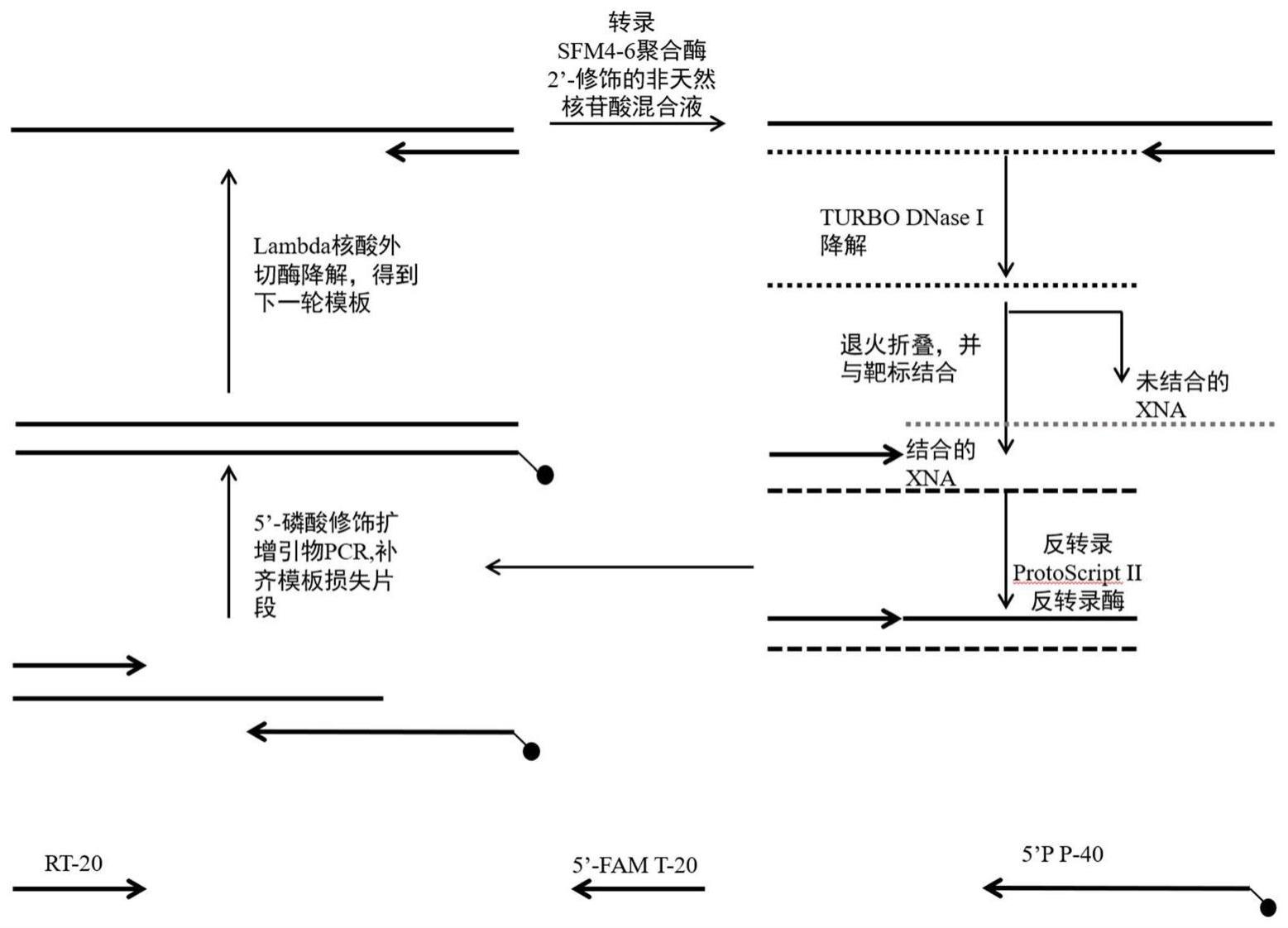
1、本发明包括2’-全修饰非天然核酸适配体文库的制备方法,其具体操作是采用四种不同组合(包括2’-f修饰的尿嘧啶核苷酸,2’-f修饰的胞嘧啶核苷酸,2’-ome修饰的腺嘌呤核苷酸以及2’-ome修饰的鸟嘌呤核苷酸;2’-f修饰的胞嘧啶核苷酸,2’-ome修饰的尿嘧啶核苷酸,2’-ome修饰的腺嘌呤核苷酸以及2’-ome修饰的鸟嘌呤核苷酸;2’-f修饰的尿嘧啶核苷酸,2’-f修饰的鸟嘌核苷酸,2’-ome修饰的腺嘌呤核苷酸以及2’-ome修饰的胞嘧啶核苷酸;2’-f修饰的腺嘌呤核苷酸,2’-f修饰的鸟嘌核苷酸,2’-ome修饰的尿嘧啶核苷酸以及2’-ome修饰的胞嘧啶核苷酸)的非天然核苷酸为元件,在sfm4-6聚合酶的作用下,以l90-n30文库为模板,直接转录合成dna/xna杂合体产物,经turbo dnase降解后,获取到2’-全修饰的xna文库。
2、本发明包括筛选靶向蜡状芽孢杆菌5/b/6中的金属β-内酰胺酶全修饰非天然核酸适配体的技术。筛选靶向5/b/6金属β-内酰胺酶非天然核酸适配体的过程中,第二轮以后的转录文库获取方法主要采用5’-磷酸化扩增引物对筛选到的xna序列反转录产物做pcr扩增,该引物与模板3’-端互补40nt,包含与转录产物5’-端互补的20nt以及转录引物部分。扩增产物用lambda核酸外切酶降解,通过控制降解时间,能最大效率降解掉磷酸化的序列以获取单链dna文库,然后再使用sfm4-6聚合酶转录获取下一轮的xna文库(筛选流程如图2所示)。在筛选靶向5/b/6金属β-内酰胺酶非天然核酸适配体的过程中,每轮筛选所用的盐离子浓度、洗脱时间、洗脱次数会有所不同,随着轮数的增加,逐渐增加选择压力。
3、本发明采用高通量测序的方法,通过对不同修饰的文库添加不同的识别碱基进行合并测序,共测得有效序列为1,946,602条,其中2’-f-c,2’-f-u,2’-ome-a,2’-ome-g的文库共测序193,889条,2’-f-c,2’-ome-u,2’-ome-a,2’-ome-g的文库共测序360,992条,2’-f-a,2’-f-g,2’-ome-c,2’-ome-u,的文库共测序62,886条,2’-f-g,2’-f-u,2’-ome-a,2’-ome-c共测序1,328,440条。
4、本发明共使用了四种不同修饰的xna文库进行selex,且四种不同修饰的文库在筛选之后均富集到以下三条相同的序列,利用rna-fold预测其可能的二级结构如图2所示;
5、5/b/6-xap-1:
6、5’-aucucuaugcuaucaaugcgcucucguuucaucgguaucauuacccccaucgguuucacacagucccagu-3’
7、5/b/6-xap-2:
8、5’-aucucuaugcuaucaaugcguggcuacauggacucguuuauccaaucggacgguuucacacagucccagu-3’
9、5/b/6-xap-3:
10、5’-aucucuaugcuaucaaugcgcggcuaaaaaucguaccaauguuuccgagcgguuucacacagtcccagu-3’
11、其中aucg均为糖环2’-c修饰的非天然核苷酸。
12、序列5/b/6-xap-1在2’-f-c,2’-f-u,2’-ome-a,2’-ome-g的文库中富集到11.2910%,在2’-f-c,2’-ome-u,2’-ome-a,2’-ome-g的文库中富集到25.4191%,在2’-f-a,2’-f-g,2’-ome-c,2’-ome-u的文库中富集到36.8270%,在2’-f-g,2’-f-u,2’-ome-a,2’-ome-c的文库中富集到12.6144%。
13、序列5/b/6-xap-2在2’-f-c,2’-f-u,2’-ome-a,2’-ome-g的文库中富集到3.8315%,在2’-f-c,2’-ome-u,2’-ome-a,2’-ome-g的文库中富集到2.8463%,在2’-f-a,2’-f-g,2’-ome-c,2’-ome-u的文库中富集到0.4198%,在2’-f-g,2’-f-u,2’-ome-a,2’-ome-c的文库中富集到1.5169%。
14、序列5/b/6-xap-3在2’-f-c,2’-f-u,2’-ome-a,2’-ome-g的文库中富集到2.2647%,在2’-f-c,2’-ome-u,2’-ome-a,2’-ome-g的文库中富集到2.3895%,在2’-f-a,2’-f-g,2’-ome-c,2’-ome-u的文库中富集到0.29577%,在2’-f-g,2’-f-u,2’-ome-a,2’-ome-c的文库中富集到0.9630%。
15、与现有技术相比,本发明的优势在于:
16、相较于固相合成来说,固相合成全修饰xna难度非常大,通常超过30nt的xna序列的合成成本就非常高,且成功率低。并且固相合成无法完成selex整个筛选过程的循环,即使能合成第一轮的全修饰文库,也无法进行下一轮的扩增-转录-反转录的循环。本发明利用定向进化后的sfm4-6聚合酶转录合成xna适配体文库,不仅合成效率高,合成的文库序列更长,并且在整个selex过程获取每一轮的全修饰xna文库中起到了决定性的作用,是整个筛选流程得以循环的关键。与现今常用的利用其他聚合酶突变株转录得到xna序列相比,定向进化后的sfm4-6聚合酶能够耐高温,通过高温转录得到的文库多样性更大,并且转录效率更高,在适宜的条件下转录得到的全长产物更多,操作流程更加简便。
- 还没有人留言评论。精彩留言会获得点赞!