一种提高马铃薯薯块中维生素C含量的方法
一种提高马铃薯薯块中维生素c含量的方法
技术领域
1.本技术涉及一种提高马铃薯薯块中维生素c含量的方法,属于生物技术领域。
背景技术:
[0002][0003]
近年来,越来越多的人们开始关注马铃薯中对人体有益的成分,特别是抗坏血酸(维生素c)的含量。抗坏血酸是迄今为止在光合生物中最丰富的低分子量的抗氧化剂,也是人体必需营养素。由于缺乏l-葡萄糖酸-1,4-内酯氧化酶(wheeler et al.,1998),人类无法自主合成抗坏血酸,人体所需90%的维生素c来自新鲜蔬菜和水果。因此,作物的维生素c含量是一个重要的营养品质性状。然而,由于马铃薯的维生素c含量相对较低,研究人员试图通过操纵维生素c生物合成途径来培育具有高维生素c水平的新品种。尽管在其他作物中,操纵维生素c生物合成途径可以培育出维生素c含量提高8-10 倍的新品种,但在马铃薯中,据发明人所见的报道,维生素c含量提高并不大,例如在马铃薯中过表达维生素c循环再生基因脱氢抗坏血酸还原酶 (dhar)也可增加其维生素c含量(qin et al.,2011);使用强启动子(35s)或薯块特异性启动子(pat)将拟南芥或马铃薯自身ggp基因转入马铃薯,薯块抗坏血酸含量比野生型提高了2.7倍(35s-atvtc2)和4.7倍(pat-stvtca)(bulleyet al.,2012)。因此,人们希望能获得维生素c含量更高的马铃薯。
[0004]
同时,农作物的产量与品质性状很难兼顾,例如,在高产花青素水稻材料中,该水稻产量明显低于对照材料,说明营养品质的提升降低了水稻的产量(zhu et al.,2017);再如,ems诱变产生的p17c5番茄突变体维生素c含量比野生型高2-5倍,植株表型为花果异常、雄性不育、种子很少或没有种子(deslous et al.,2021),说明维生素c含量高也会影响作物的正常生长。因此,在实际育种中,培育产量高、同时维生素c含量也高的优质马铃薯存在很大挑战。
[0005]
另外,如何降低或去除植物的苦味也是育种工作者长期致力解决的问题。尽管人们已经发现了一些主要的苦味化合物,以及与这些苦味化合物密切关联的基因或蛋白等,但由于植物的苦味的形成涉及植物代谢调控网络中的各成分之间复杂的相互作用,因此,仍然需要更多的该方面的研究。
技术实现要素:
[0006]
为了解决上述背景技术中的问题,本技术首先提供一种多肽,其氨基酸序列如seq id no.1或seq id no.2所示。
[0007]
本技术还提供一种核酸,其为编码上述多肽的核酸;或者其为序列如 seq id no.3或seq id no.4所示的核酸。
[0008]
本技术进一步提供一种提高马铃薯维生素c含量、或降低马铃薯苦味、和/或去除马铃薯苦味的方法,或培育高维生素c含量、或低苦味、和/或无苦味的马铃薯的方法,包括抑制上述多肽的正常翻译,或者修饰上述核酸的序列。
[0009]
在一种实施方式中,将马铃薯ggp酶基因的5’非翻译区修饰为包括 seq id no.5或seq id no.6所示的核酸。
[0010]
本技术还提供一种鉴定高维生素c含量、或低苦味、和/或无苦味的马铃薯的方法,检测马铃薯ggp酶基因的5’非翻译区或上述核酸是否存在干扰uorf正常翻译的修饰,例如,是否存在seq id no.5或seq id no.6所示的核酸。
[0011]
本技术还提供一种判断植物维生素c含量高低的方法,包括检测或比较植物ggp酶基因的5’非翻译区序列或ggp酶基因开放阅读框附近的序列,所述序列中存在修饰的植物的维生素c含量高于未修饰的植物的维生素c含量,存在大片段修饰的植物的维生素c含量高于存在小片段修饰的植物的维生素c含量。
[0012]
在一些实施方式中,修饰为至少一个核苷酸的缺失、添加和/或置换。
[0013]
在一些实施方式中,“至少一个核苷酸”为1-5个、6-9个、10-17个、 18-24个、25-44个、45-81个、82-142个、143-148个、149-152个、或153 以上个。
[0014]
本技术还提供一种核酸,其序列为seq id no.5或seq id no.6所示。
[0015]
本技术还提供与该经修饰的核酸相关的生物材料,所述生物材料为含有所述核酸分子的表达盒;或含有所述核酸分子的载体;或含有所述核酸分子的细胞。
[0016]
本技术进一步提供上述的多肽、核酸、或生物材料在马铃薯育种中的用途;优选的,所述育种为培育维生素c含量高、苦味降低、和/或无苦味的马铃薯。
[0017]
本技术的突出贡献在于:
[0018]
①
发现马铃薯ggp酶基因的uorf不能正常翻译可以提高维生素c含量,并获得维生素c含量提升了24.12倍的优质马铃薯。
[0019]
②
根据ggp基因的5’utr区或uorf附近序列的修饰片段大小分为两类:小片段修饰和大片段修饰;被修饰的片段越大,植物中维生素c 含量就越高,即修饰片段的大小和植物中维生素c含量呈正相关。
[0020]
③
发现马铃薯薯块中维生素c含量大幅升高时(如高于野生型8.72倍时)会显著抑制植株生长和薯块发育,而适度提高薯块维生素c含量 (1.46-8.72倍)则没有影响,据此,获得了产量高、且维生素c含量也高的马铃薯。
[0021]
④
发现了提高维生素c含量能降低植物苦味。通过修饰ggp基因的 5’utr区或uorf附近序列,提高了维生素c含量的同时,也使苦味化合物含量降低,并使与苦味化合物紧密关联的基因下调表达。
附图说明
[0022]
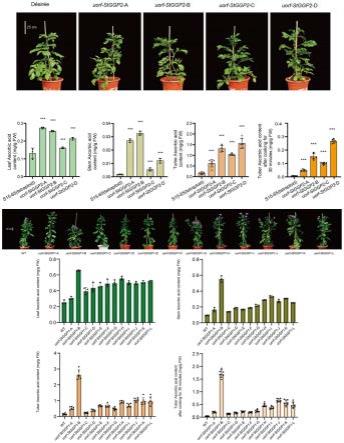
图1、uorf-stggp不同程度突变的马铃薯的叶、茎、薯块及蒸煮后薯块中维生素c含量均比野生型的高。
[0023]
图2、uorf-stggp1-a/-b的修饰方式。其中,uorf序列为自方框内atc 开始(蓝色),sgrna靶点用粗体下划线表示,删除的核苷酸用连字符表示, uorf起始密码子用红色方框展示。
[0024]
图3、uorf-stggp1-a/-b纯合突变材料叶、茎、薯块及蒸煮后薯块中维生素c含量显著提高。
[0025]
图4、uorf-stggp1-a/-b纯合突变材料地上/地下表型测定。
[0026]
图5、uorf-stggp1-a/-b马铃薯叶片和薯块中sga生物合成通路代谢物(a),基因通路图(b)和qp结果(c)。*表明代谢物含量的值通过t检验确定与有显着差异(p《0.05)。标记为红色的代谢物表示其相对含量相对于对照升高,而标记为蓝色的代谢物表示其相对含量相对于对照降低。
[0027]
图6、uorf-stggp系列修饰方式示例。
具体实施方式
[0028]
下述实施例中以具体试验为例,所使用的实验方法如无特殊说明,均为常规方法。下述实施例中所用的材料、试剂等,如无特殊说明,均可从商业途径得到。
[0029]
在高等植物中,l-半乳糖途径被认为是合成维生素c的主要途径。该途径以d-葡萄糖作为前体,通过十步酶促反应最终合成l-抗坏血酸。其中, gdp-l-半乳糖磷酸酶(ggp)在调控维生素c合成中具有非常重要作用 (macknight et al.,2017)。
[0030]
uorfs(upstream open reading frames)是真核生物部分基因在mrna 5’端的一段开放阅读框,是常见的顺式作用元件,可负调控下游mrna序列的转录(calvo et al.,2009,von arnim et al.,2014)。本技术提供一种uorfs多肽,其氨基酸序列如seq id no.1或seq id no.2所示。本技术还提供编码该 uorfs多肽的核酸。
[0031]
本技术从马铃薯中鉴定到两个ggp基因,即stggp1和stggp2,其5’非翻译区的序列分别如seq id no.3、seq id no.4所示。
[0032]
本技术进一步提供一种提高马铃薯维生素c含量、或降低马铃薯苦味、和/或去除马铃薯苦味的方法,或培育高维生素c含量、或低苦味、和/或无苦味的马铃薯的方法,包括抑制上述多肽的正常翻译;优选的,抑制的方式为修饰上述核酸的序列。
[0033]
本技术还提供一种鉴定高维生素c含量、或低苦味、和/或无苦味的马铃薯的方法,检测马铃薯ggp酶基因(stggp)的5’非翻译区或上述核酸是否存在修饰。
[0034]
在一些实施方式中,修饰为至少一个核苷酸的缺失、添加和/或置换;优选的,所述修饰抑制ggp酶基因uorf多肽的正常翻译;优选的,“至少一个核苷酸”为1-5个、6-9个、10-17个、18-24个、25-44个、45-81个、82
‑ꢀ
142个、143-148个、149-152个、或153以上个。
[0035]
在一些实施方式中,修饰为在seq id no.3中:删除第234-257位的核苷酸(删除24bp);或者删除第207-358位的核苷酸(删除152bp)。
[0036]
本技术提供一种经修饰的核酸,其为修饰的stggp1基因的5’非翻译区,序列为seq id no.5或seq id no.6。
[0037]
本技术提供一种核酸,其为马铃薯ggp酶基因的经修饰的5’utr区 (即5’非翻译区),其特征在于,所述修饰抑制了uorf编码的多肽的正常翻译,所述多肽的序列如seq id no.1或seq id no.2所示。
[0038]
在一种实施方式中,马铃薯ggp酶基因为stggp1和/或stggp2。
[0039]
在一种实施方式中,马铃薯ggp酶基因5’utr区的序列为seq idno.3,seq id no.4,seq id no.5和/或seq id no.6。
[0040]
本技术提供一种生产维生素c含量高、苦味降低、和/或无苦味的马铃薯的方法,其特征在于,包括使马铃薯ggp酶基因的5’utr区为前述经修饰的核酸。
[0041]
本技术提供一种获得产量高同时维生素c含量也高的、苦味降低、和/或无苦味的
马铃薯的方法,包括使马铃薯ggp酶基因的5’utr区为前述经修饰的核酸。
[0042]
本技术还提供与上述所有核酸相关的生物材料,其为含有所述核酸分子的表达盒;或含有所述核酸分子的载体;或含有所述核酸分子的细胞;或含有所述核酸分子的植物(例如马铃薯)组织或器官(根、茎、叶、花或果实);或含有所述核酸分子的整个植物或种质,所述植物或种质(例如为马铃薯) 可以为杂合突变或纯合突变;二倍体或四倍体;杂合或纯合突变的二倍体、或杂合或纯合突变的四倍体。
[0043]
本技术提供一种提高马铃薯维生素c含量、或降低马铃薯苦味、和/或去除马铃薯苦味的方法,或培育高维生素c含量、或低苦味、和/或无苦味的马铃薯的方法,将马铃薯ggp酶基因的5’非翻译区修饰为seq id no.5或 seq id no.6表征的核酸。
[0044]
本技术提供一种鉴定高维生素c含量、或低苦味、和/或无苦味的马铃薯的方法,检测马铃薯ggp酶基因的5’非翻译区是否存在序列为seq id no.5 或seq id no.6的核酸。“存在”是指包含或者本身即是。
[0045]
本技术进一步提供经修饰的核酸或与该经修饰的核酸相关的生物材料在马铃薯育种中的用途;优选的,所述育种为培育维生素c含量高、苦味降低、和/或无苦味的马铃薯。
[0046]
本技术提供一种培育产量高同时维生素c含量也高的、苦味降低、和/或无苦味的马铃薯的方法,将马铃薯ggp酶基因的5’非翻译区修饰为seq idno.5表征的核酸。
[0047]
本技术提供一种降低马铃薯苦味化合物含量的方法,包括提高维生素c 的含量的步骤。
[0048]
本技术提供一种降低或去除马铃薯苦味的方法,包括增加维生素c的含量的步骤。
[0049]
本技术提供一种判断植物维生素c含量高低的方法,比较植物ggp酶基因的5’非翻译区序列或ggp酶基因开放阅读框附近的序列,存在大片段修饰的植物比存在小片段修饰或未修饰的植物的维生素c含量高。
[0050]
本技术提供一种降低植物苦味化合物含量,或者降低植物苦味,和/或去除植物苦味的方法,包括提高植物中维生素c的含量的步骤。
[0051]
在一种实施方式中,苦味化合物为α-茄碱,α-查茄碱,和/或茄尼定。
[0052]
在一种实施方式中,降低植物苦味,和/或去除植物苦味为下调参与茄尼定代谢的关键基因表达。
[0053]
在一种实施方式中,关键基因是game4,6,7,8a,8b,11,12,sgt1 和/或sgt3。
[0054]
在一种实施方式中,所述提高维生素c的含量的步骤包括修饰植物ggp 酶基因的5’非翻译区或ggp酶基因上游开放阅读框附近的区域,从而抑制 uorf正常翻译。
[0055]
本技术的植物为单子叶植物或双子叶植物(例如马铃薯),可指植物叶、植物茎、植物根、植物花或植物果实。
[0056]
本技术的马铃薯可指马铃薯叶、马铃薯茎段、马铃薯薯块、或经烹饪的马铃薯薯块。
[0057]
在一种实施方式中,提高维生素c的含量的步骤包括使马铃薯ggp1基因的5’非翻译区或上游开放阅读框附近的区域、和/或ggp2基因的5’非翻译区或上游开放阅读框附近的区域发生突变的步骤。
[0058]
本技术构建了敲除靶基因的crispr-cas9载体,利用含靶基因敲除载体的农杆菌eha105侵染经过2天预培养的4周苗龄、不带腋芽的马铃薯茎段。侵染后的外植体在含有抗
生素的诱导再生培养基上培养8周,期间可两周更换一次培养基。再生苗经过生根抗性筛选获得阳性植株。对阳性植株外源 dna片段进行pcr检测,对靶位点序列进行扩增,并通过sanger测序确定靶位点突变情况。将成功突变的植株进行定植并通过qpcr检测靶基因的表达量最终确定目标基因已被敲除。为了进一步研究马铃薯产量与维生素c含量动态协调的生物学机理,本技术还做了转录组和代谢组的研究。具体实施方式如下。
[0059]
实施例1、克隆stggp1和stggp2的uorf及5’utr
[0060]
从二倍体马铃薯克隆s.tuberosum组phureja s15-65和四倍体系 d
é
sir
é
e中克隆基因stggp1和stggp2。其中,s15-65由国际马铃薯中心友情提供,d
é
sir
é
e由云南师范大学栽培教研室友情提供。发现并确认stggp1 和stggp2基因的uorf及5’utr,使用引物对其进行序列克隆和测序。测序结果:stggp1和stggp2的uorf编码的多肽序列分别如seq idno.1或seq id no.2所示;stggp1和stggp2的5’utr序列分别如seqid no.3或seq id no.4所示。
[0061]
实施例2、突变stggp1和stggp2的uorf附近区域
[0062]
通过基因编辑技术,设计靶向uorf-stggp1或uorf-stggp2起始密码子的单向导rna(sgrna)。由于基因编辑技术的自我修复带有不确定性,可能引入随机的插入和缺失;而且马铃薯ggp-uorf哪些位置容易发生基因编辑、进行基因编辑后是否可以大幅提高翻译水平等,都是不确定的。因此,下面将采用多种方式来确证最终的结果。
[0063]
stggp1uorf的单向导rna序列和stggp2uorf的19-nt单向导 rna序列均使用pkse402-atu626载体进行构建。其中,使用bsai酶消化 pkse402用于插入sgrna,最终的二元载体用于通过根癌农杆菌菌株 eha105转化马铃薯节间,gfp荧光用于鉴定突变植物。
[0064]
所有野生(wt)和突变株系在受控温室的秋季中正常生长,光照时间为 16h光照/8h暗光周期,白天温度为25℃/夜间温度为18℃。
[0065]
实施例3、植物的基因分型和倍性
[0066]
从gfp荧光阳性马铃薯植株中收集叶样本,并用十六烷基三甲基溴化铵(ctab)方法提取基因组dna。使用特定引物从所有再生植株中扩增目标序列,并测序以鉴定目标区域中的突变。通过对克隆的pcr产物进行测序,对单个品系进行基因分型,其中二倍体植株每个样品测8个单克隆测序,四倍体植株每个样品则至少需要20个单克隆测序。马铃薯愈伤组织再生期间染色体加倍发生的频率非常高,因此可以通过流式细胞术测定(bd, aria sorp)确定突变植物的倍性。
[0067]
实施例4、双荧光素酶报告基因检测
[0068]
为了产生用于双荧光素酶报告基因检测的构建体,将35s启动子和野生型和突变形式的stggp1和stggp2的5'前导序列(或非翻译区)克隆到具有bamhi和ncoi的pgreenii0800-luc载体中。野生(wt)和突变的构建体通过农杆菌介导的转化在烟叶中瞬时表达。将含有双荧光素酶报告载体的根癌农杆菌菌株gv3101重新悬浮在含有1m mgcl 2和100mm 乙酰丁香酮的500mm mes缓冲液中,至最终od600为0.6。使用无针注射器将缓冲液渗透到4周龄烟草的完全展开的叶子中。渗透后,植物在黑暗下生长12小时,然后在16小时光照/8小时黑暗下生长2天。luc/ren 活性用双荧光素酶报告基因检测系统测量。
[0069]
实施例5、抗坏血酸含量的测定
[0070]
成熟的马铃薯叶子、茎、块茎组织和蒸煮30分钟后的马铃薯薯块在液氮中彻底冷
冻。使用提取缓冲液溶解粉末,该缓冲液含有74.45mg edta、 286.65mg tcep和5ml 98%正磷酸,终体积为100ml milli-q水。将悬浮液涡旋30秒。然后将样品置于4℃30分钟。随后,将它们在4℃下以 12,000g离心30分钟,保留上清液并使用4mm亲水性ptfe注射器过滤器过滤。使用液相色谱三重四极杆质谱法(lc-ms/ms agilent 6460)测量抗坏血酸浓度。使用aquility csh c18(waters)分析过滤后的样品。色谱分离以0.2ml/min的流速进行梯度洗脱,使用0.1%甲酸水溶液作为流动相 a,使用甲醇和0.1%甲酸作为流动相b。梯度如下:5%b至45%b作为分析运行,然后50秒上升至95%b,然后在95%b下保持2分钟,然后在5%b下重新平衡2分钟,进样体积为15微升。在分离过程中,分析柱在恒温柱温箱中保持在35℃的温度。基于质量丰度比对所有分析物进行了母离子和产物离子的鉴定。lc洗脱液以负esi模式引入agilent 6490系统。详细实验条件为:离子喷雾电压3500v,气帘35psi,碰撞气体介质,温度350℃。在多反应监测(mrm)模式下检测分析物离子。
[0071]
实施例6、修饰结果
[0072]
对于uorf-stggp2,我们设计了两个sgrna,靶向远离uorf-stggp2 起始密码子的序列,最终在d
é
sir
é
e背景中获得了四个嵌合系(uorf-stggp2
‑ꢀ
a/-b/-c/-d)。每个系在uorf-stggp2的序列中具有2至44bp缺失的组合,除了uorf-stggp2-b除了缺失突变外还具有1bp插入。luc/ren分析证实,从这些突变体中鉴定出的所有uorf-stggp2破坏均能有效地操纵下游报告基因的转录,提高马铃薯维生素c含量(图1)。
[0073]
对于uorf-stggp1,获得了二倍体,且stggp1-uorf突变材料薯块中维生素c含量均值为1.03mg/g fw,约为stggp2-uorf突变株系的2倍。因此,以下主要对uorf-stggp1进行研究。
[0074]
对发明人创制的一系列马铃薯stggp-uorf突变及薯块维生素c含量升高的种质材料(图1,图6)的表型进行观察发现,这些材料叶片、茎、薯块中维生素c含量均有不同程度提高,而且stggp-uorf片段缺失大小与植物中维生素c含量之间存在正相关,即被突变的片段越大,维生素c的含量越高。其中,两个分别含有24bp和152bp缺失的纯合突变种质uorf-stggp1
‑ꢀ
a/-b(或者写作:ggp1-uorf-a、ggp1-uorf-b)是最终筛选出来的最具代表性的马铃薯(图2)。
[0075]
就突变体整体而言,与对照相比,突变材料叶片中的维生素c含量的提高为1.20-2.71倍;茎中增加倍数为1.50-6.14倍;薯块中的维生素c含量提升显著,增幅为1.51-24.12倍。尤其令人惊讶的是,与对照s15-65和其他二倍体突变体相比,具有最大片段(152bp)缺失的uorf-stggp1-b突变体的叶、茎和块茎中的维生素c水平分别显着增加了2.71、5.78、24.12倍。
[0076]
另外,鉴于维生素c在烹饪过程中极易降解,我们还进一步检测了在高温下这些材料薯块中维生素c含量的变化。结果表明蒸煮30分钟后,对照和突变材料薯块中维生素c均存在一定程度的损耗。但与对照相比(损耗了 80%-90%),基因编辑材料的薯块维生素c损耗约20%-50%,仍然保留了足够营养成分(图1)。
[0077]
就最具代表性的马铃薯uorf-stggp1-a和uorf-stggp1-b而言,与对照相比,小片段缺失(24bp)的uorf-stggp1-a材料(5’utr区的核酸序列如 seq id no.5所示)叶、茎和薯块中维生素c含量各增加了2.02、2.60和8.72 倍;大片段(152bp)缺失的uorf-stggp1-b材料(5’utr区的核酸序列如seqid no.6所示)叶、茎和薯块中的维生素c水平分别增加2.71、
5.78和24.12 倍。其中,uorf-stggp1-b的薯块维生素c含量增加倍数是已知报道中维生素c含量提升最显著和浓度变化最大的案例(图1,图3)。但是,大片段敲除的uorf-stggp1-b株系与对照材料和uorf-stggp1-a相比,地上部分植株出现矮化、花期延迟、生物量减少和薯块减产表型。其中,株高、地上鲜重和单株块茎重分别降低了29.24%、27.53%和69.29%。然而,uorf-stggp1-a 薯块中维生素c含量显著提高(8.72倍)的同时,并没有出现生长发育抑制现象(图1,图4)。
[0078]
可见,uorf-stggp1-a是兼顾了产量和营养品质的种质材料,即是产量高、且维生素c含量也高的马铃薯;而uorf-stggp1-b则是能量流过度转化成代谢流的种质材料,是维生素c含量超高的马铃薯。
[0079]
实施例7、组学分析
[0080]
(1)转录组数据测定与分析
[0081]
样品测序后,进行reads过滤,去除3'端带接头的序列和平均质量分数低于q20的reads。利用hisat2(http://ccb.jhu.edu/software/hisat2/index.shtml) 软件将过滤后的reads比对参考基因组 (http://spuddb.uga.edu/dm_v6_1_download.shtml),利用stringtie统计比对到每一个基因上read count值作为基因的原始表达量,并采用fpkm值对表达量进行标准化。利用deseq2软件对各样品的基因表达量进行pca分析,并对标准化后基因数据进行差异表达分析,筛选差异基因的条件为显著性p
‑ꢀ
value《0.05且|log2 fold change|》1。利用top go进行富集分析,利用超几何分布确定差异基因显著富集的go term。利用r语言clusterprofiler进行keggpathway富集分析,确定差异表达基因相关的信号通路。
[0082]
(2)建立马铃薯代谢数据库
[0083]
使用高质量的二级碎片信息和高分辨的质谱数据可建立马铃薯代谢数据库。主要通过标准品比对、代谢数据库匹配、谱图解析和参考文献等四个方面来解析物质的结构。首先,将具有高分辨数据的代谢物进行分子式计算,然后与公开的代谢数据库进行分子式、碎片裂解模式的匹配,实现代谢物结构的初步解析。对于有标准品可以参考的代谢物,通过比对标准品和未知物质的m/z、rt和碎片裂解模式,如果以上三个条件都是相同的,则确定为同一物质。对于没有标准品可以参考的代谢物,主要通过谱图解析来鉴定代谢物,综合运用已发表的文献和公开的代谢数据库等信息,系统的总结代谢物的洗脱规律、裂解规律等,运用到未知物质的解析。
[0084]
(3)代谢组数据测定与分析:
[0085]
利用agilent masshunter qualitative analysis软件处理正、负离子模式下采集的全息质谱数据。主要包括以下步骤:数据导入、色谱峰对齐、实验设计、峰选择、归一化、去卷积、化合物鉴定和统计分析。根据自动选择的质控样品对导入的样品进行分析。选择加合离子模式有:[m-h2o-h]-、[m-h]-、 [m+fa-h]-、[m+na-2h]-、[m+k-2h]-、[2m+fa-h]-和[2m-h]-。质谱数据先去卷积化,然后进行pca主成分分析和opls-da分析,利用s-plot图鉴定各组的差异代谢物,再利用自制马铃薯代谢数据库对保留时间-精确质量对 (rt-em)进行定性分析。最后通过方差分析(p-value≤0.05和max foldchange值≥2)进行数据过滤,确定样品中差异代谢物。
[0086]
(4)基因与代谢物相关分析
[0087]
利用权重基因共表达网络分析(wgcna)软件对多个测序样品中的共有基因表达数据进行共表达分析,使用代谢物作为性状,经过聚类分析,聚类的不同分支代表不同的基因模块(module),同一模块内基因共表达程度高。根据不同组织不同时期的样本中分析获得的差异代谢物和差异基因进行 kmeans聚类分析,再按分类进行绘图,可找到变化趋势一致代谢物和基因。为了进一步确定哪些代谢物和基因高相关性表达,需要使用皮尔逊相关性(pearson)方法计算变化趋势一致基因和代谢物之间的相关性,按照相关性大于0.6,p-value≤0.05筛选出相关性较高的基因和代谢物。利用cytoscape 软件对较高相关性差异基因和差异代谢物绘制网络图,通过网络图可以直观显示代谢物和基因之间的关系。
[0088]
(5)组学分析结果
[0089]
马铃薯维生素c含量升高引发α-茄碱和α-查茄碱极显著下调表达。发明人观察到ggp1-uorf-a和ggp1-uorf-b这两个突变体中维生素c上调时代谢物变化的相似性。例如,与wt相比,茄科中的主要苦味化合物α
ꢀ‑
茄碱和α-查茄碱在uorf-stggp1-a/-b(即uorf-stggp1-a和/或uorf
‑ꢀ
stggp1-b)中均显着降低,根据峰面积估算,约降低了200倍(图5)。此外,在uorf-stggp1-a/-b中,参与茄尼定代谢的目前已知的关键基因,如 game 4,6,7,8a,8b,11,12,sgt1和sgt3全部都下调表达,这表明马铃薯块茎中维生素c的增加将降低其苦味,受到消费者的青睐。因此,维生素c含量升高导致苦味化合物含量降低,从而降低或去除植物苦味是发明人意外发现的新的重要机制。
[0090]
综上,本技术以一系列马铃薯stggp-uorf突变及薯块维生素c含量升高的种质材料为例(作为试材),来深入研究植物ggp-uorf突变与维生素 c含量的关系。发明人发现,ggp-uorf突变后,这些材料叶片、茎、薯块中维生素c含量均有不同程度提高,且维生素c含量增加与植物ggp-uorf 片段缺失大小呈正相关,即小片段缺失导致维生素c含量增长幅度远低于大片段缺失的植株,因此,植物ggp-uorf突变片段越大,其维生素c含量越高。
[0091]
尤其是,本技术获得的维生素c含量最高的株系(uorf-stggp1-b)的薯块维生素c含量提高了24.12倍,是目前已知报道中维生素c含量提升最显著的案例。但是,发明人发现uorf-stggp1-b植株生长发育、薯块产量均受到显著抑制。而在另外一份小片段缺失的株系uorf-stggp1-a中,薯块维生素 c含量提升了8.72倍,但该植株生长发育没有影响。因此,可以得出,uorf
‑ꢀ
stggp1-b是一份能量流向代谢流转化过度的极端突变材料,而uorf-stggp1
‑ꢀ
a则是能量流与代谢流平衡保持较好的材料,兼顾了品质和产量。
[0092]
利用这两份特殊材料以及代谢组、转录组等组学大数据技术进行试验,结果揭示,马铃薯维生素c含量升高导致苦味化合物及其代谢网络中的关键基因极显著地下调表达,提示了植物维生素c的合成可能影响了苦味形成的过程,因此,可以通过提高维生素c含量(如突变ggp-uorf)来去除或降低植物苦味。
[0093]
本技术已经尽力阐述了全部关键的技术细节,实施例仅仅为了示例本技术的构思,不应当反而成为限制本技术保护范围的理由。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1