一种Na
一种na
+
/磷酸根协同转运体基因pvptb在富集砷和/或吸收利用磷元素中的应用
技术领域
1.本发明涉及基因工程技术领域,具体地,涉及一种na
+
/磷酸根协同转运体基因pvptb在富集砷和/或吸收利用磷元素中的应用。
背景技术:
2.砷是一种在自然界中广泛存在的类金属元素,砷及其可溶性化合物都具有很强的毒性。砷污染的水源、土壤、沉积物是许多国家饮用水和食物链污染的主要来源,对植物、动物等多种生命形态都有很大的危害。土壤中的砷积累达到一定程度,会直接影响植物的生长发育、降低农业产量,并可能进入蔬果的可食用组织及水稻、小麦等农作物的籽粒中。经饮用水或饮食暴露,人体摄入一定量的砷易造成各类皮肤癌症、肾脏损伤和心血管疾病等不可逆损害。土壤砷的点位超标率达2.7%,加强土壤砷污染的治理与修复,对于改善土壤环境质量具有重要意义。
3.重金属土壤污染修复方法主要是工程治理措施和物理化学修复方法,但成本相对较高、对环境扰动大,且有可能造成土壤的二次污染。近年来兴起的植物修复技术因具有成本低、操作简单、生态效益高、对环境扰动较小和无二次污染等优点,展现出在土壤修复方面的优势和发展前景。其中,植物提取修复是当前应用最为广泛且最有前景的植物修复方式。
4.蜈蚣草是世界上最强的砷超富集植物、是砷污染土壤植物提取修复的最重要植物材料。其中含有大量与砷超富集相关的关键基因能够用于构建砷提取修复工程植物。但是,目前已报道的一些蜈蚣草基因,特别是在高等(显花)植物中存在高度同源性的基因,基因工程应用中效果并不显著。可能由于序列具有较高同源性,工程植物自身的条件系统会限制其发挥功能。而将一些高等(显花)植物中没有的基因类型进行基因工程应用,往往具有较好效果。目前,缺少对蜈蚣草特有的砷超富集相关关键基因的研究与利用关键基因进行土壤污染修复的应用。
技术实现要素:
5.本发明的目的是克服现有技术的上述不足,提供一种na
+
/磷酸根协同转运体基因pvptb在富集砷和/或吸收利用磷元素中的应用。
6.本发明的第一个目的是提供一种na
+
/磷酸根协同转运体基因pvptb。
7.本发明的第二个目的是提供一种na
+
/磷酸根协同转运体蛋白pvptb。
8.本发明的第三个目的是提供一种重组载体。
9.本发明的第四个目的是提供一种重组工程菌。
10.本发明的第五个目的是提供所述na
+
/磷酸根协同转运体基因pvptb在提高微生物和/或植物富集和/或吸收砷和/或磷元素中的应用。
11.本发明的第六个目的是提供所述na
+
/磷酸根协同转运体蛋白pvptb在提高微生物
和/或植物富集和/或吸收砷和/或磷元素中的应用。
12.本发明的第七个目的是提供所述重组载体在提高微生物和/或植物富集和/或吸收砷和/或磷元素中的应用。
13.本发明的第八个目的是提供所述重组工程菌在提高微生物和/或植物富集和/ 或吸收砷和/或磷元素中的应用。
14.本发明的第九个目的是提供一种富集和/或吸收砷和/或磷元素的方法。
15.本发明的第十个目的是提供一种构建富集砷和/或磷元素微生物和/或植物的方法。
16.为了实现上述目的,本发明是通过以下方案予以实现的:
17.一种na
+
/磷酸根协同转运体基因pvptb,所述na
+
/磷酸根协同转运体基因 pvptb的核苷酸序列如seq id no:7和/或seq id no:8所示。
18.一种na
+
/磷酸根协同转运体蛋白pvptb,所述na
+
/磷酸根协同转运体蛋白 pvptb的氨基酸序列如seq id no:16和/或seq id no:17所示。
19.一种重组载体,所述重组载体中含有所述na
+
/磷酸根协同转运体基因pvptb。
20.一种重组工程菌,所述重组工程菌中含有所述na
+
/磷酸根协同转运体基因 pvptb。
21.优选地,所述微生物为酵母菌。
22.优选地,所述植物为拟南芥植物。
23.所述na
+
/磷酸根协同转运体基因pvptb在提高微生物和/或植物富集和/或吸收砷和/或磷元素中的应用。
24.优选地,所述微生物为酵母菌。
25.优选地,所述植物为拟南芥植物。
26.优选地,所述na
+
/磷酸根协同转运体基因pvptb的核苷酸序列如seq id no: 7所示。
27.优选地,所述植物为拟南芥植物地上部分。
28.所述na
+
/磷酸根协同转运体蛋白pvptb在提高微生物和/或植物富集和/或吸收砷和/或磷元素中的应用。
29.优选地,所述微生物为酵母菌。
30.优选地,所述植物为拟南芥植物。
31.优选地,所述na
+
/磷酸根协同转运体蛋白pvptb的氨基酸序列如seq id no: 16所示。
32.优选地,所述植物为拟南芥植物地上部分。
33.所述重组载体在提高微生物和/或植物富集和/或吸收砷和/或磷元素中的应用。
34.优选地,所述微生物为酵母菌。
35.优选地,所述植物为拟南芥植物。
36.优选地,所述重组载体中含有所述na
+
/磷酸根协同转运体基因pvptb的核苷酸序列如seq id no:7所示。
37.优选地,所述植物为拟南芥植物地上部分。
38.所述重组工程菌在提高微生物和/或植物富集和/或吸收砷和/或磷元素中的应
用。
39.优选地,所述微生物为酵母菌。
40.优选地,所述植物为拟南芥植物。
41.优选地,所述重组工程菌中含有所述na
+
/磷酸根协同转运体基因pvptb的氨基酸序列如seq id no:7所示。
42.优选地,所述植物为拟南芥植物地上部分。
43.一种富集和/或吸收砷和/或磷元素的方法,在微生物和/或植物中过表达核苷酸序列如seq id no:7和/或seq id no:8所示的na
+
/磷酸根协同转运体基因 pvptb。
44.优选地,所述微生物为酵母菌。
45.优选地,所述植物为拟南芥植物。
46.优选地,在微生物和/或植物中过表达核苷酸序列如seq id no:7所示的na
+
/磷酸根协同转运体基因pvptb。
47.一种构建富集砷和/或磷元素微生物和/或植物的方法,在微生物和/或植物中过表达核苷酸序列如seq id no:7和/或seq id no:8所示的na
+
/磷酸根协同转运体基因pvptb。
48.优选地,所述微生物为酵母菌。
49.优选地,所述植物为拟南芥植物。
50.优选地,在微生物和/或植物中过表达核苷酸序列如seq id no:7所示的 na
+
/磷酸根协同转运体基因pvptb。
51.与现有技术相比,本发明具有以下有益效果:
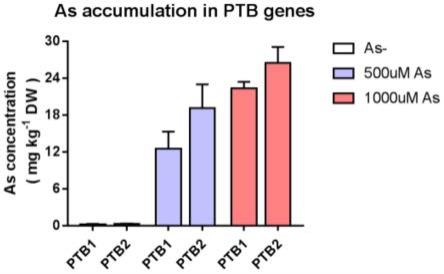
52.本发明公开了一种na
+
/磷酸根协同转运体基因pvptb在富集砷和/或吸收利用磷元素中的应用。在蜈蚣草中提取出2个na
+
/磷酸根协同转运体基因pvptb,其核苷酸序列分别如seq id no:7和seq id no:8所示,为pvptb1和pvptb2 基因。pvptb1和pvptb2基因是常见的高等植物中未存在的一类磷转运蛋白基因,将其转运到ey917酵母突变株,使酵母具有砷和磷转运功能。在拟南芥中表达所述pvptb基因,提高了其地上部的砷富集能力90%~110%,总磷积累量 12%~46%。在工程植物或微生物中过表达pvptb基因,用于修复环境中的砷污染和提高磷利用效率。
附图说明
53.图1为蜈蚣草pvptb的进化分析和组织表达模式;a为蜈蚣草pvptb1和 pvptb2的系统发育树分析图;b为基于定量pcr分析的蜈蚣草pvptb1和 pvptb2基因在根系和地上部的相对表达模式。
54.图2为酵母系统研究蜈蚣草pvptb的磷转运功能结果,具体为不同磷酸盐梯度(1mm、2mm、5mm、10mm和20mm)的酵母打点实验结果。
55.图3为酵母系统研究蜈蚣草pvptb的砷转运功能结果,具体为在1mm的 na2haso4·
7h2o条件下,转入pvptb的酵母对砷的抗性实验结果。
56.图4为酵母系统研究蜈蚣草pvptb的砷转运功能结果,具体为转入pvptb 的酵母对砷的吸收积累结果。
57.图5为pvptb1转基因拟南芥的组培实验中对砷和磷的吸收转运情况,其中 a为
pvptb1转基因拟南芥地上部对砷积累量;b为pvptb1转基因拟南芥根对砷积累量;c为pvptb1转基因拟南芥地上部对总磷的积累量;d为pvptb1转基因拟南芥根对总磷的积累量。不同字母的标注代表显著差异p《0.05。
58.图6为pvptb2转基因拟南芥的组培实验中对砷和磷的吸收转运情况,其中 a为pvptb2转基因拟南芥地上部对砷积累量;b为pvptb2转基因拟南芥根对砷积累量;c为pvptb2转基因拟南芥地上部对总磷的积累量;d为pvptb2转基因拟南芥根对总磷的积累量。不同字母的标注代表显著差异p《0.05。
59.图7为组培pvptb1和pvptb2转基因拟南芥在加砷条件下的生长状况,其中a为缺磷加砷条件下的野生型(wt)、pvptb1转基因拟南芥(ex1-2和ex1-10)、 pvptb2转基因拟南芥(ex2-2和ex2-3),b为加磷加砷条件下的野生型(wt)、 pvptb1转基因拟南芥(ex1-2和ex1-10)、pvptb2转基因拟南芥(ex2-2和 ex2-3)。
具体实施方式
60.下面结合说明书附图及具体实施例对本发明作出进一步地详细阐述,所述实施例只用于解释本发明,并非用于限定本发明的范围。下述实施例中所使用的试验方法如无特殊说明,均为常规方法;所使用的材料、试剂等,如无特殊说明,为可从商业途径得到的试剂和材料。
61.实施例1蜈蚣草pvptb1和pvptb2的克隆
62.一、实验方法
63.提取蜈蚣草根系rna(ncbi登录号为taxonomy id:13821),并反转录为cdna,分别使用克隆引物pvptb1-pcr-f(seq id no:1)、pvptb1-pcr-r (seq id no:2)和pvptb2-pcr-f(seq id no:3)、pvptb2-pcr-r(seqid no:4)进行pcr扩增获得克隆产物。
64.将克隆产物使用擎科公司的凝胶回收试剂盒(tsingke tsp601-200,china) 进行纯化回收,得到纯化产物,并将纯化产物用pclone007 blunt vector kit (tsingke,china)连接克隆载体,最终转入大肠杆菌感受态细胞dh5α chemically competent cell(tsingke,china)中,利用测序引物m13f(seq idno:5)及m13r(seq id no:6)进行测序,引物具体核苷酸序列见表1。
65.表1引物核苷酸序列表
[0066][0067][0068]
二、实验结果
[0069]
克隆载体的测序结果显示,所获得的克隆产物与预测的pvptb1和pvptb2 序列完全一致,表明成功克隆获得了核苷酸序列分别如seq id no:7和seq idno:8所示的pvptb1和pvptb2基因序列(编码区cds序列区域)。
[0070]
实施例2蜈蚣草pvptb1和pvptb2的生物信息学分析和表达模式鉴定
[0071]
一、实验方法
[0072]
(1)蜈蚣草的培养
[0073]
蜈蚣草孢子采集自广东省广州市番禺区,于实验室内进行繁殖。将蜈蚣草孢子(0.01~0.02g)均匀地撒在盛营养土的塑料盆中,盖上薄膜,定期浇水,摆放在温室中育苗。待蜈蚣草幼苗长至5~6叶,移至1/5霍格兰营养液(hns) 中曝气培养,每周换一次营养液,预培养两周。所有植株均生长在光照14h、昼 /夜温度26/20℃、相对湿度60%、光强3000lx的温室中。
[0074]
(2)rna提取和转录分析
[0075]
为了鉴定蜈蚣草中核苷酸序列分别如seq id no:7和seq id no:8所示的pvptb1和pvptb2基因的组织表达模式,用fastpure plant total rna isolationkit试剂盒(vazyme biotech,nanjing,china),以水培的蜈蚣草为材料,并进行加磷、缺磷、加磷加砷三种处理,提取不同处理组的蜈蚣草rna。将得到的rna 使用hiscript ii一步rt-pcr试剂盒(vazyme biotech,nanjing,china)反转录成 cdna。
[0076]
以pvactin基因(seq id no:9)作为蜈蚣草pvptb1和pvptb2基因的内参,并以所得cdna作为模板,使用sybr green pcr master mix试剂盒(vazymebiotech,nanjing,china)进行实时荧光定量pcr(qrt-pcr),结果采用2-δδct
法进行统计。定量引物信息见表1。
[0077]
表2 qrt-pcr引物列表
[0078]
序号引物名称序列seq id no:10qrt-pvactin-fgggcagtatttccaagcatagtgggseq id no:11qrt-pvactin-rtgcctcgctttgattgagcctcatcseq id no:12pvptb1_q_faccgcagccttattttcccaseq id no:13pvptb1_q_raacgtcattctcgtaccgcaseq id no:14pvptb2_q_fgacatcccgcttcacgatctseq id no:15pvptb2_q_rcagggcatcgacagagtgtt
[0079]
(3)系统进化树的构建
[0080]
利用dnaman软件翻译核苷酸序列分别如seq id no:7和seq id no: 8所示的pvptb1和pvptb2基因序列,获得氨基酸序列分别如seq id no:16 和seq id no:17所示的pvptb1和pvptb2蛋白序列,作为植物基因研究的模式植物,我们在ncbi网站上获得了水稻中的13个pht1基因和拟南芥中的9 个pht1基因(见表3),并得到了它们的氨基酸序列。在水稻和拟南芥中利用 ncbi网站检索对比pvptb的同源基因,但并未得到结果。
[0081]
通过对蜈蚣草中的pvptb1和pvptb2蛋白、模式植物水稻中的pht1家族蛋白和模式植物拟南芥中的pht1家族蛋白的氨基酸序列进行分析和对比,利用 mega x的neighbor-joining(邻接法)构建系统发育树。
[0082]
表3水稻和拟南芥中的pht1家族蛋白的基因的ncbi编号
[0083][0084][0085]
二、实验结果
[0086]
图1a反映了在本研究中发现的2个na
+
/磷酸根协同转运体基因pvptb与常见的模式植物中的磷酸盐转运蛋白并不属于同一分支,且ncbi的同源基因的比对结果显示,本发明的pvptb1和pvptb2基因在显花植物中丢失,是常见的高等植物中未存在的一类磷转运蛋白。
[0087]
通过qrt-pcr对蜈蚣草中的pvptb1和pvptb2基因进行分析,发现pvptb1 和pvptb2基因在蜈蚣草的根系种特异性表达,且缺磷与加砷处理并不会下调该基因的表达,说明该基因可在砷暴露条件下持续发挥砷吸收功能(图1b)。
[0088]
实施例3利用酵母系统研究pvptb1和pvptb2的砷和磷转运功能
[0089]
一、实验方案
[0090]
(1)酵母表达载体的构建
[0091]
为构建酵母表达载体,分别以pvptb1和pvptb2克隆载体为模板,利用诺维赞公司ce design在线设计网站设计的引物,将同源臂分别克隆到核苷酸序列分别如seq id no:7和seq id no:8所示的pvptb1和pvptb2基因的cds 序列两端。克隆获得的pcr产物经凝胶纯化,通过一步法快速克隆试剂盒(hieff plus one step cloning kit,yesen)同源重组到pag426gpd-ccdb表达载体的spei和hindiii限制性酶切位点之间,构建的重组载体被命名为 pag426::pvptb1和pag426::pvptb2。
[0092]
将重组载体转化大肠杆菌dh5α感受态细胞,挑取阳性单克隆,经pcr验证及测序通过后,保存菌液,分别提取pvptb1和pvptb2重组载体质粒,备用。
[0093]
(2)酵母突变株的转化及阳性转化子的鉴定
[0094]
利用酵母快速转化试剂盒(yeast transformation kit,coolaber)将空载体 pag426-v与重组载体pag426::pvptb1和pag426::pvptb2分别转入酵母突变株ey917中,于
sd-u选择培养基平板筛选阳性单克隆,获得pvptb1酵母转化株和pvptb2酵母转化株。ey917酵母突变株缺失了酵母内源全部5个磷转运体 (δpho84(ncbi登录号:854916)、δpho89(ncbi登录号:852599)、δpho87 (ncbi登录号:850403)、δpho90(ncbi登录号:853241)、δpho91(ncbi 登录号:4838906)),完全失去了对磷(pi)的吸收和转运功能。挑取于30℃恒温培养箱中生长4~7天后大小适宜的酵母阳性单克隆,200mm、30℃摇床中培养至od600约为1.0,富集菌体,使用酵母质粒提取试剂盒(yeast dnaextraction kit,thermo)提取表达酵母质粒并回转到大肠杆菌中进行二次测序验证。测序通过后,-80℃保存酵母表达菌株,备用。
[0095]
(3)酵母平板生长实验
[0096]
设置不同的磷酸盐浓度(1mm、2mm、5mm、10mm和20mm)处理,进行酵母打点实验,30℃培养3天,拍摄。
[0097]
(4)酵母砷抗性实验
[0098]
将od600约1.0的pvptb1和pvptb2酵母转化株,以3000rpm转速离心富集5min,清洗两次后用少量无菌水重悬,得到pvptb1酵母转化株重悬液和 pvptb2酵母转化株重悬液。
[0099]
将pvptb1酵母转化株重悬液分装至含有0μm na2haso4·
7h2o(as-)和 1mm na2haso4·
7h2o(as+)的30ml sd-u酵母液体培养基中,酵母初始 od600约为0.2。酵母细胞在200rpm、30℃的摇床中摇动培养,每隔一段时间测定波长为600nm处酵母细胞液的吸光度,绘制酵母生长曲线。pvptb2酵母转化株培养操作与pvptb1酵母转化株相同。
[0100]
(5)酵母砷积累实验
[0101]
将pvptb1酵母转化株重悬液分装至含有0μm na2haso4
·
7h2o、500μm na2haso4·
7h2o和1mm na2haso4·
7h2o的sd-u酵母液体培养基中,使培养液初始od600介于0.8~1.0之间。
[0102]
酵母细胞在200rpm、30℃的摇床中摇动培养,3h后,以3000rpm转速离心富集5min,用4℃超纯水清洗菌体4次并离心,保存菌体及最后一次洗脱液,待检测其中的总砷含量。
[0103]
总砷含量的测定采用使用usepa 3050b的浓硝酸-过氧化氢体系(10ml 1:1 (w/v)hno3和2ml 30%h2o2),于石墨炉消解仪中105℃消解酵母菌体。利用铟(in)内标法,使用电感耦合等离子体质谱仪(icp-ms)测定样品中的总砷含量。pvptb2酵母转化株培养、总砷含量测定操作与pvptb1酵母转化株相同。
[0104]
二、实验结果
[0105]
图2的酵母平板生长实验的结果显示,在低磷(pi)浓度条件下,酵母并不能正常生长,而在磷(pi)浓度达到5mm时,酵母可正常生长,且pvptb1酵母转化株生长优于pvptb2酵母转化株,所以pvptb1及pvptb2可以在高磷(pi) 环境回补ey917酵母突变株的磷吸收能力,具有低亲和磷(pi)转运功能。
[0106]
图3的酵母砷抗性实验结果显示,pvptb1和pvptb2酵母转化株在砷的胁迫下,生长受到了抑制,这表明在酵母中表达pvptb1和pvptb2蛋白能够增强酵母菌株对砷的吸收转运能力,进而证明pvptb1和pvptb2蛋白具有砷转运能力。
[0107]
图4的酵母砷积累实验结果表明,在添加500μm as和1mm as的酵母培养基中培养固定时间与相同生长状态的pvptb1和pvptb2酵母转化株,pvptb1 和pvptb2酵母转化株能够吸收培养基中的as,且随着培养基中砷浓度的提高, as吸收量也会有所上身,所以
pvptb1和pvptb2蛋白具备砷转运能力。
[0108]
因此,pvptb蛋白同时具备磷酸盐和砷酸盐转运功能。
[0109]
实施例4构建pvptb1和pvptb2转基因拟南芥植株
[0110]
一、实验方法
[0111]
1.拟南芥转基因载体的构建和阳性转基因拟南芥株系的鉴定
[0112]
将核苷酸序列分别如seq id no:7和seq id no:8所示的pvptb1和 pvptb2基因序列转入植物表达载体psn1305中,构建psn1305-pvptb1和psn1305-pvptb2重组载体。将重组载体psn1305-pvptb1和psn1305-pvptb2 导入农杆菌菌株gv3101中。
[0113]
通过农杆菌转化法构建转基因拟南芥株系,具体的实验方案为:
[0114]
将构建好的含有重组载体psn1305-pvptb1和psn1305-pvptb2的农杆菌菌株gv3101划线培养,用含kana和rif抗性的lb培养基扩培含有重组载体 psn1305-pvptb1和psn1305-pvptb2的农杆菌,在农杆菌摇至活性最佳od (0.8~1.0)后富集菌体,转移进入侵染液(质量比5%蔗糖,体积比0.02% sliwet77)。
[0115]
在基质土中种植培养野生型拟南芥,在拟南芥出现绿色花苞后,将拟南芥的未开花絮(稍微露点白色比较好)浸泡在上述含有农杆菌的侵染液中45s,继续种植培养待拟南芥成熟后收种。
[0116]
通过种植培养收获转基因株系t1代拟南芥种子,在添加30mg/l潮霉素的 1/2ms培养基上萌发t1代拟南芥幼苗,筛选出阳性转化植株。筛选出阳性苗后在含有30mg/l潮霉素的1/2ms培养基上继续培养,等到其生物量达到一定量之后,收集植物组织,提取植物rna,进行qrt-pcr鉴定pvptb1和pvptb2 转基因拟南芥株系。
[0117]
二、实验结果
[0118]
通过潮霉素筛选和qrt-pcr鉴定,pvptb1转基因拟南芥筛选出两个生长状态良好的拟南芥株系(ex1-2和ex1-10),pvptb2转基因拟南芥筛选出两个生长状态良好的拟南芥株系(ex2-2和ex2-3)。
[0119]
实施例5组培环境下pvptb1和pvptb2转基因拟南芥对磷和砷吸收利用的情况
[0120]
一、实验方法
[0121]
(1)拟南芥的组培和处理方法
[0122]
为了明确实施例4得到的pvptb1和pvptb2转基因拟南芥对磷和砷吸收的能力,对组培pvptb1和pvptb2转基因拟南芥植株和野生型拟南芥植株中总磷和砷的含量进行测定。
[0123]
将实施例4获得的阳性pvptb1转基因拟南芥株系(ex1-2和ex1-10)和 pvptb2转基因拟南芥株系(ex2-2和ex2-3)的种子在含有30mg/l潮霉素的无菌1/2ms培养基上萌发,在同时间和半天后分别在不含抗生素的无菌1/2ms培养基上萌发野生型拟南芥株系(wt),5天后,将长势一致的拟南芥幼苗分别移入ck(未添加as
ⅴ
)和含有50μm as
ⅴ
两种处理的1/2ms培养基上处理。 2周后,分离拟南芥的根系和地上部组织于液氮中速冻,随后保存于-80℃环境中备用。
[0124]
(2)总磷含量的测定
[0125]
用低温冷冻干燥机冻干拟南芥的根系和地上部组织,称取约0.05g经干燥粉碎后的植物组织于消解管中,用usepa 3050b的浓硝酸-过氧化氢体系(10ml 1: 1(w/v)hno3和2ml 30%h2o2),于石墨炉消解仪中105℃消解植物组织。用钼酸盐比色法测定样品植物组织
总磷含量。
[0126]
(3)砷含量的测定
[0127]
用步骤(2)的方法得到消解植物组织,用铟(in)内标法,电感耦合等离子体质谱仪(icp-ms),测定样品植物组织中的总砷含量。
[0128]
二、实验结果
[0129]
与野生型相比,pvptb1转基因拟南芥地上部的砷含量显著增加了90%~ 110%,其中ex1-2转基因株系的效果最好,地上部的砷含量增加了110%;ex1-10 转基因株系地上部的砷含量增加90%,根系中的砷含量没有显著差异,说明转入 pvptb1基因促进拟南芥对砷的吸收和向地上部的转运(图5a、图5b)。
[0130]
与野生型相比,pvptb1转基因拟南芥地上部的总磷含量显著增加了12%~ 46%,其中ex1-2转基因株系的地上部的总磷含量增加了46%;ex1-10转基因株系地上部的总磷含量增加12%,根系的总磷含量不变,说明转入pvptb1基因促进拟南芥对磷的吸收(图5c、图5d)。
[0131]
与野生型相比,pvptb2转基因拟南芥地上部的砷含量显著增加了27~51%,其中ex2-3转基因株系的效果最好,地上部的砷含量增加了51%;ex2-2转基因株系地上部的砷含量增加27%,根系中的砷含量没有显著差异,说明转入pvptb2 基因促进拟南芥对砷的吸收和向地上部的转运(图6a、图6b)。与野生型相比, pvptb2转基因拟南芥地上部的总磷含量并未发生显著变化,根系的总磷含量不变(图6c、图6d)。
[0132]
与野生型相比,pvptb1和pvptb2转基因拟南芥能够提高地上部的砷积累量,但从图7中拟南芥的表型可以看出,更高的砷积累量并不会影响拟南芥的生长。
[0133]
以上结果说明,转入pvptb1和pvptb2基因促进了拟南芥地上部对砷和磷的吸收和积累,其中转基因拟南芥株系ex1-2对拟南芥地上部对砷和磷的吸收和积累的促进效果最好,而且这种对砷的富集作用并不会影响拟南芥本身的生长。
[0134]
最后所应当说明的是,以上实施例仅用以说明本发明的技术方案而非对本发明保护范围的限制,对于本领域的普通技术人员来说,在上述说明及思路的基础上还可以做出其它不同形式的变化或变动,这里无需也无法对所有的实施方式予以穷举。凡在本发明的精神和原则之内所作的任何修改、等同替换和改进等,均应包含在本发明权利要求的保护范围之内。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1