香港巨牡蛎新型Lectin基因全长序列及其抗菌应用
香港巨牡蛎新型lectin基因全长序列及其抗菌应用
技术领域:
1.本发明属于生物化学与分子生物学,具体涉及香港巨牡蛎新型lectin基因全长序列及其抗菌应用。
背景技术:
2.海洋生物资源是人类尚待开发使用的一个重要资源领域。随着国家海洋战略的调整和发展,沿海海水养殖业特别是海洋贝类的养殖已经成为海洋经济发展的一个重要的支柱产业,据2012年商务部统计,当年海洋贝类的养殖就达到了1179.6万吨,年产量占世界贝类总产量的60%以上。其中牡蛎375.63万吨,经济价值达到200多亿元人民币,占海水贝类养殖产量的30%以上,因此,牡蛎的养殖在我国沿海经济社会发展中占据举足轻重的地位,并具有巨大的发展潜力。但是,我国海洋贝类的发展是以传统养殖育种为主,采取的是集约化、工业化的养殖模式,水产养殖动物对病害的抵抗能力下降,极易发生传染性疾病,给养殖行业带来巨大的经济损失。
技术实现要素:
3.本发明的目的是提供香港巨牡蛎新型lectin基因全长序列及其抗菌应用。
4.本发明在牡蛎转录组测序的基础上,首次在牡蛎中发现了一个新的抗菌基因,属于c型凝集素,含有1个或者多个糖基识别域(carbohydrate-recognition domain,crd),其生物学活性对于ca
2+
具有依赖性,该型凝集素作为先天性免疫的模式识别受体,在宿主的免疫防御系统中,参与到识别蛋白对病原微生物的识别中,c型凝集素在识别微生物细胞膜表面的糖分子后,通过激活生物体内的免疫系统来清除入侵的微生物。通过race技术,我们获得了该基因的全长,该基因的多肽序列具有高度的物种特异性,其蛋白序列约有20-30%同源于其他物种的同源蛋白。因此,该基因是一个全新的基因,其功能有待于进一步证实。定量pcr实验表明,该基因在细菌感染后显著上调,因此,该基因参与了宿主的免疫应答反应。我们通过构建酵母表达载体,利用毕赤酵母对该基因进行蛋白表达,通过纯化获得纯化蛋白,发现该蛋白的具有抑菌功能,将有利于提升养殖贝类的种质创新能力,支撑产业的高效可持续性发展。
5.本发明提供了一种新的贝类免疫相关的基因-香港巨牡蛎新型lectin基因全长序列,其核苷酸序列如seq id no.1的22bp至504bp所示。
6.该基因cdna全长577p,自22bp至504bp区段为其开放阅读框,编码160个氨基酸,5'非编码区长21bp,3'非编码区长73bp,有多聚腺苷酸加尾信号(tataaa)和多聚腺苷酸尾巴。
7.本发明的第二个目的是提供上述香港巨牡蛎新型lectin基因编码的蛋白,其氨基酸序列如seq id no.2所示。
8.本发明的第三个目的是提供上述蛋白在制备抑菌药物中的应用。
9.优选,所述的抑菌药物是抑制溶藻弧菌生长的药物。
10.本发明还提供了一种抑菌药物,其含有上述蛋白作为活性成分。
11.本发明的第四个目的是提供一种检测香港巨牡蛎中的新型lectin基因表达水平的制剂在制备预测香港牡蛎感染菌株制剂中的应用。
12.优选,所述的检测香港巨牡蛎的lectin基因表达水平是检测香港巨牡蛎的血细胞中lectin基因表达水平。
13.优选,所述的预测香港牡蛎感染菌株制剂是预测香港牡蛎感染溶藻弧菌、葡萄球菌或酵母菌的制剂。
14.本发明通过构建cdna文库的方法,从香港牡蛎中发现了lectin的基因序列,利用rt-pcr的方法以及race技术,克隆到了该基因序列全长,结果表明该基因cdna全长为577bp,自22bp至504bp区段为开放阅读框,编码160个氨基酸,5'非编码区长22bp,3'非编码区长73bp,末尾有多聚腺苷酸加尾信号(tataaa)和多聚腺苷酸尾巴。定量pcr实验表明,给基因在成年的香港牡蛎中各个组织中均有表达,该基因分别在微生物溶藻弧菌,葡萄球菌和酵母感染下表达上调;通过构建酵母表达载体,通过酵母的诱导表达,获得了一个18kd左右的蛋白;通过大量表达、纯化,获得目标蛋白,在抑菌实验中,发现该蛋白对于溶藻弧菌的生长具有抑制作用。
附图说明
15.图1是lectin基因及蛋白编码序列分析;a,lectin核酸及蛋白编码,方框为信号肽序列,阴影部分为lectin结构域,下划线为加尾信号;b,lectin蛋白的结构域预测结果。
16.图2是实时定量pcr检测lectin mrna的表达模式;a.lectin在不同组织中的分布,实验结果表明lectin在性腺中有显著性表达,在消化腺、外套膜、鳃、心脏、肌肉和外套膜中有中度表达,在血细胞表达量相对较少(图2a);b,在溶藻弧菌(v.alginolyticus)、葡萄球菌(s.haemolyticus)以及酵母菌(s.cerevisiae)感染时lectin在血淋巴细胞中的表达情况;星号(*,**)指示和对照组差异显著或者极其显著(p《0.5或者p《0.05)。
17.图3是通过毕赤酵母诱导的lectin蛋白表达;m:蛋白marker;1泳道:洗脱纯化蛋白;2泳道:穿透液;3泳道:原液。
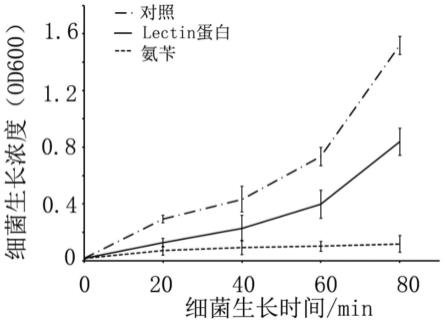
18.图4是lectin蛋白具有抑制溶藻弧菌生长的功能。
具体实施方式
19.以下实施例是对本发明的进一步说明,而不是对本发明的限制。
20.实施例1:
21.1.rna提取
22.a).香港巨牡蛎细胞或组织加trizol后,采用组织研磨器研磨,室温放置5min,使其充分裂解。
23.b).12,000rpm离心5min,弃沉淀。
24.c).按200ul氯仿/ml trizol加入氯仿,振荡混匀后室温放置15min。
25.d).4℃12,000g离心15min。
26.e).吸取上层水相,至另一离心管中。
27.f).按0.5ml异丙醇/ml trizol加入异丙醇混匀,室温放置5-10min。
28.g).4℃12,000g离心10min,弃上清,rna沉于管底。
29.h).按1ml 75%乙醇/ml trizol加入75%乙醇,温和振荡离心管,悬浮沉淀。
30.i).4℃8,000g离心5min,尽量弃上清。
31.j).室温晾干或真空干燥5-10min。
32.k).可用50ul h2o,te buffer或0.5%sds溶解rna样品,55-60℃,5-10min。
33.l).在室温孵育15min之后,加入250μl dnase stop solution(dsa),在13,000
×
g离心1min。
34.m).重复步骤a-j,加100μl无核酸酶水,然后,在13,000
×
g离心1min,弃离心柱,使用分光光度计测量所收集的溶液中的rna的浓度,分装后,贮存于-80℃。
35.2.第一条链cdna的合成
36.1)dnase i消化
37.按照如下组分分别将各组的rna加入到pcr离心管中
[0038][0039]
2)用枪头轻轻吹打混匀并稍作离心;37℃,15min,消化rna中的dna
[0040]
3)加入1μl edta混匀,65℃,10min使dnase i失活
[0041]
4)按照如下组分依次加到离心管:
[0042][0043]
用枪头轻轻吹打混匀并稍作离心,放置于37℃,30min;
[0044]
5)85℃,5s使反转录酶失活,然后4℃短期保存,-20℃长期保存。
[0045]
3、差减杂交文库构建
[0046]
差减杂交文库使用pcr-select cdna subtraction kit(clontech,usa)试剂盒进行构建。香港巨牡蛎受弧菌刺激8h后,收集血细胞,按照上述的方法提取血细胞总rna,并使用同样的方法提取对照组(未受弧菌处理)血细胞总rna。以刺激后香港巨牡蛎作为tester,对照组作为driver,进行扣除杂交。将差减杂交后的cdnas亚克隆到pgem-teasy vector(promega,usa)并转化到escherichia coli jm109(promega,usa)感受态细胞。随机挑选2000个克隆并进行测序。
[0047]
4、lectin基因全长的克隆
[0048]
依据步骤2所得到的克隆测序,通过blasten等生物软件分析并确定lentin序列,再以此序列为模版,通过smarter race cdna试剂盒(invitrogen,usa)进行该基因的5'/3'
race,具体操作参照说明书。得到race序列后,通过拼接,以获得该lectin基因的全长cdna,其序列入seq id no.1所示,该基因cdna全长577bp,自22bp至504bp区段为其开放阅读框,编码160个氨基酸,5'非编码区长21bp,3'非编码区长73bp,有多聚腺苷酸加尾信号(tataaa)和多聚腺苷酸尾巴(图1),其编码的氨基酸序列如seq id no.2所示:
[0049]
seq id no.1
[0050][0051]
seq id no.2
[0052]
mgatrklsslilvfsyitvahgsgcrsgwtqyknkcymfsrmtepwgmassycqayhsklaepvtseesaflsshahslhgtfwigitdltaenewvyssnlqrvqttnwashepngkgvencavlyyprhsqwvdlhcdthekficemsedesgpviig。
[0053]
3.实时定量pcr
[0054]
进行实时定量pcr所使用的试剂盒为lightcycler 480sybr green i(roch),所使用的仪器为lightcycler 480system(roche),所使用的模板为上述所获得的反转录产物,所使用的内参gapdh,所使用的引物有:
[0055]
lectin-f:5'-tccgaggagtccgcctttcta-3'
[0056]
lectin-r:5'-tcgtggcttgcccagtttgt-3'
[0057]
gapdh-f:5'-ggattggcgtggtggtagag-3'
[0058]
gapdh-r:5'-gtatgatgcccctttgttgagtc-3'
[0059]
qrt-pcr的反应体系如下:2
×
mix:10μl,chlectin-f:0.2μm;chlectin-r:0.2μm;反转录产物(cdna):50ng。
[0060]
热循环条件如下:95℃处理10min;接下来运行40个循环:95℃变性5秒,60℃退火延伸20秒。qrt-pcr运行完成后,进行溶解曲线分析以确定pcr扩增是否特异。使用2
-δδct
方法对转录本进行相对定量。对于每一份样品,实验组和对照组均设置3个平行反应。
[0061]
4、组织分布及样品处理
[0062]
分别提取5只牡蛎的各个组织的总rna(见步骤1)并反转录为cds,采用real-time pcr技术检测了lectin基因在香港巨牡蛎的淋巴细胞,腮,性腺,消化腺,外套膜,围心腔,闭
壳肌中的分布情况。结果显示它在各个组织中广泛表达,其中在血淋巴中表达量最低,性腺最高(图2a)。
[0063]
取正常牡蛎,分别注射1.0
×
109个活的病原菌,以pbs为参照,体积为100微升,在不同的时间点以5个一组的方式取样,提取血细胞的总rna并反转录为cdna,见步骤1。依此为模版,进行样品的real-time pcr检测,结果发现溶藻弧菌、葡萄球菌、以及酵母菌(图2b)都引起lectin基因的显著表达。
[0064]
5、表达载体的克隆,表达与纯化
[0065]
为了得到真核表达载体,以包含lectin的核酸序列为模版,进行pcr克隆扩增,随后采用takara pcr产物凝胶回收试剂盒纯化,酶切,进行picza载体的亚克隆,所有引物如下:
[0066][0067]
5.1.piczaa载体构建
[0068]
1)采用ecor i/kpn i对picza a载体(invitrogen,k1740-01)进行双酶切,回收载体,采用lectin pi f/r引物,利用高保真酶对目标基因进行克隆,纯化该pcr产物,利用上述2种纯化产物(载体/lectin),采用clonexpress
tm ii试剂盒(南京诺唯赞公司),进行连接反应,将piczaa-lectin连接产物转化至top10菌株中,涂布于含25μg/ml zeocin的lb平板上,37℃过夜培养。
[0069]
2)挑取lb平板上形态大小良好的单菌落接种到3ml lb液体培养基中,37℃过夜培养,抽提质粒(步骤见质粒抽提试剂盒,maga),进行酶切和pcr检测,结果正确的克隆载体,进一步进行送样测序(华大基因公司);对于正确的克隆载体,无菌条件下吸取菌液700ul于400ul 50%甘油(已灭菌)中保种,随后转接剩下的菌液于300ml lb(25μg/ml的zeocin)液体培养基中,过夜培养。
[0070]
3)大提质粒,质粒纯度100%,浓度在10ug/ul左右。
[0071]
4)用sac1酶对piczaa-lectin质粒进行线性化,37℃水浴过夜后回收。
[0072]
5)回收后用eppendorf电转化仪分别电转化piczaa-lectin于gs115感受态中。电转参数为:电压1500v;时间5ms。
[0073]
6)涂布150ul转化后的piczaa-lectin菌液于rdb+g418平板上,其中g418的浓度为1mg/ml,30℃培养4-5天。涂布150ul转化后的piczaa-lectin菌液于ypg+zeocin平板上,30℃培养4-5天。
[0074]
5.2piczaa载体小量表达筛选
[0075]
1)挑取rdb+g418,ypg+zeocin与平板上的单菌落20株进行pcr鉴定。
[0076]
2)将pcr鉴定为阳性的菌株点在ypg平板上保种,同时挑在装有1.5mlypg液体培养基的试管中,30℃,200rpm培养48h。保种的平板放在30℃恒温培养箱中培养72h。
[0077]
3)将试管取出,放在试管架上静置5~6小时,待菌体全部沉淀于试管底部后,在超净工作台中倒掉试管中的上清,向试管中加入含1%甲醇的yp 1ml。继续30℃,200rpm培养24h。
[0078]
4)在超净工作台内,向试管中加入200ul含5%甲醇的yp进行诱导,30℃,200rpm培养24h。
[0079]
5)将试管中的菌液倒至1.5mlep管中,11000rpm,10min离心。
[0080]
6)取离心后的上清900ul于另一个1.5mlep管中,并向其中加入150ultca,震荡均匀,放在插入冰中静置2h。
[0081]
7)取出冰中的ep管,11000rpm,10min离心,弃上清。
[0082]
8)向ep管中加入200ul丙酮,11000rpm,10min离心,弃上清。将沉淀放置于60℃烘箱中15-20min,去除丙酮。
[0083]
9)向ep管中加入20ulbuffer,超声5min溶解沉淀,送样跑page胶。
[0084]
各步骤的电泳图如图3所示。
[0085]
5.3.摇瓶发酵
[0086]
1)根据上述小量表达的结果,从之前保种的ypg平板上,将有表达的菌株接种至15mlypg液体培养中,30℃,200rpm培养24h。
[0087]
2)无菌条件下吸取菌液700ul于400ul50%甘油(已灭菌)中保种,将剩下的菌液接种至1l的ypg液体培养基中,30℃,200rpm培养24h。
[0088]
3)将培养24h的菌液在超净工作台中倒入500ml灭菌的backman离心瓶中,8000rpm,3min离心,用yp(含1%甲醇)重悬菌体,倒入原来的5l三角瓶中,并加入1l yp(含1%甲醇)和200ul消泡剂,30℃,200rpm培养24h。
[0089]
4)在超净工作台内,向摇瓶中加入10ml甲醇诱导,30℃,200rpm培养24h。
[0090]
5)继续加入10ml甲醇诱导,30℃,200rpm培养24h后,将菌液倒入500ml的backman离心瓶中,8000rpm,5min离心,保留上清。
[0091]
5.4 ni柱纯化
[0092]
1)样品处理:用1mtris ph9.0将浓缩后的上清调ph至8.0。
[0093]
2)取层析柱,用3倍柱床体积的去离子水冲洗柱床。
[0094]
3)用3倍柱床体积的nta-0冲洗柱床。
[0095]
4)用5倍柱床体积的0.1m niso4冲洗柱床。
[0096]
5)用2倍柱床体积的醋酸钠&氯化钠(ph4.0)溶液冲洗柱床。
[0097]
6)用3倍柱床体积的nta-0冲洗柱床。
[0098]
7)以1ml/min的流速将处理好的样品过柱。
[0099]
8)上样结束后,用溶液nta-0冲洗柱床到g250检测基本不显色。
[0100]
9)用10mm咪唑冲洗柱床。g250检测有蓝色开始收集洗脱峰,g250检测基本无色停止收集洗脱峰。
[0101]
10)用30mm咪唑冲洗柱床。g250检测有蓝色开始收集洗脱峰,g250检测基本无色停止收集洗脱峰。
[0102]
11)用60mm咪唑冲洗柱床。g250检测有蓝色开始收集洗脱峰,g250检测基本无色停止收集洗脱峰。
[0103]
12)用250mm咪唑冲洗柱床。g250检测有蓝色开始收集洗脱峰,g250检测基本无色停止收集洗脱峰。
[0104]
13)用3倍柱床体积的0.1m edta冲洗柱床。
[0105]
14)用水冲洗柱床,至ph试纸检测流出液为中性。
[0106]
15)用3倍柱床体积的20%乙醇冲洗。封柱置于4℃。
[0107]
16)用10kd超滤管超滤洗脱液至1~2ml,得到lectin蛋白。
[0108]
5.5抗菌功能研究
[0109]
37℃过夜生长的溶藻弧菌作为菌种,按照1:100的接种量进行接种,9个100ml的三角瓶,按照阴性对照组(不加任何抗生素,bsa 50ug/ml),实验组(lectin蛋白的工作浓度50ug/ml),氨苄阳性对照组(amp工作浓度50ug/ml)分为3组,每组3个重复,摇床37度,220rpm,每隔20分钟测一次od600,每次测3次,取平均值。
[0110]
结果如图4所示,从图4可以看出,lectin蛋白具有抑制溶藻弧菌生长的功能。
[0111]
以上所述的实施例仅是对本发明的优选方式进行描述,并非对本发明的范围进行限定,在不脱离本发明设计精神的前提下,本领域普通技术人员对本发明的技术方案做出的各种变形和改进,均应落入本发明群里要求书确定的保护范围。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1