新型Zn-尼克酰胺转运蛋白及其应用
新型zn-尼克酰胺转运蛋白及其应用
技术领域
1.本发明属于基因工程技术领域,更具体地,本发明涉及一种新型zn-尼克酰胺转运蛋白及其应用。
背景技术:
2.锌(zn)和铁(fe)是人体不可缺少的营养元素。谷物是发展中国家的主粮,其锌和铁含量过低导致了全世界近20亿人的锌营养不良。zn和fe也是植物必需营养元素,在植物光合作用,呼吸作用等生命活动过程具有不可代替的作用。因此,研究矿质元素zn和fe在谷物种子中的运输和积累过程,不仅利于农作物生产实践,也是使用生物技术手段改造食物中的锌和铁含量进行生物强化的必经之路。
3.锌和铁既能以离子的形式直接被转运蛋白进行转运,也可以和金属螯合物进行结合形成复合物被金属转运蛋白转运。na(nicotianamine)就是这些有机小分子中的一员,它在植物体内可以和铁、锌、铜、锰、镍等多种金属离子螯合,参与这些金属离子的运输和分布,维持植物体内金属离子的平衡和控制金属离子在籽粒中的积累。
4.禾本科植物玉米是目前世界上最重要的粮食作物之一,也是大多数牲畜的主要饲料,其所含营养元素的含量关系着人类和众多生物的健康。目前的报道发现了玉米籽粒中不同营养元素的分布情况,但对这些矿质元素的转运和积累机制研究较少。锌作为重要的微量元素之一,分离调控玉米籽粒锌含量自然变异的基因,研究其在玉米籽粒中的吸收和转运的分子机制,将对培育富锌玉米新品种提供理论指导和基因资源。
5.目前玉米中参与zn或者zn-na复合物转运的基因鲜有报道。zn在玉米中的吸收,长距离运输以及在籽粒中的积累必然有离子转运蛋白的参与,那么有哪些成员参与这些过程?籽粒zn含量自然变异的遗传基础是什么?这些问题都有待系统的解答。
技术实现要素:
6.本发明的目的在于提供一种新型zn-尼克酰胺转运蛋白及其应用。
7.在本发明的第一方面,提供一种zn-尼克酰胺转运蛋白或其上调剂的用途,用于:增加禾本科植物籽粒中zn或fe的含量,促进籽粒的发育;其中,所述的zn-尼克酰胺转运蛋白具有seq id no:2所示的氨基酸序列或其保守性序列变体。
8.在一种或多种优选方式中,所述增加籽粒中zn或fe的含量包括:以所述zn-尼克酰胺转运蛋白将zn、fe或zn-尼克酰胺复合物转运到细胞内;较佳地,所述zn-尼克酰胺转运蛋白表达胚乳基底转移层(细胞);较佳地,所述zn-尼克酰胺转运蛋白表达于细胞膜上。
9.在一种或多种优选方式中,所述增加籽粒中zn或fe的含量包括增加籽粒的胚或胚乳中zn或fe的含量。
10.在一种或多种优选方式中,所述促进籽粒的发育包括促进胚乳的发育;较佳地,包括促进粉质淀粉向硬质淀粉的转化,增加醇溶蛋白的含量。
11.在一种或多种优选方式中,所述保守性序列变体包括:(1)由seq id no:2所示序
列的多肽经过一或多个(如1-30个,1-20个,1-10个,1-5个,1-3个或1-2个)氨基酸残基的取代、缺失或添加而形成的,且具有seq id no:2所示序列的多肽功能的多肽;(2)氨基酸序列与seq id no:2所示序列的多肽有80%以上(较佳地≥85%,≥90%,≥95%,≥98%或≥99%)相同性,且具有seq id no:2所示序列的多肽功能的多肽;或(3)在seq id no:2所示序列的多肽的n或c末端添加标签序列,或在其n末端添加信号肽序列后形成的多肽。
12.在一种或多种优选方式中,所述的上调剂包括:与zn-尼克酰胺转运蛋白相互作用、从而提高其表达或活性的上调剂;或过表达zn-尼克酰胺转运蛋白的表达盒或表达构建物(如表达载体)。
13.在本发明的另一方面,提供一种增加禾本科植物籽粒中zn或fe的含量,促进籽粒的发育的方法,包括:在植物中提高zn-尼克酰胺转运蛋白的表达或活性;其中,所述的zn-尼克酰胺转运蛋白具有seq id no:2所示的氨基酸序列或其保守性序列变体。
14.在一种或多种优选方式中,所述的提高zn-尼克酰胺转运蛋白的表达或活性包括:以与zn-尼克酰胺转运蛋白相互作用的上调剂进行调控,从而提高zn-尼克酰胺转运蛋白的表达或活性;或,在植物中过表达zn-尼克酰胺转运蛋白。
15.在一种或多种优选方式中,所述的禾本科(gramineae)植物包括下组植物或所述zn-尼克酰胺转运蛋白或其同源物来自于包括下组植物:水稻、玉米、高粱、小米、黍、小麦、大麦、燕麦、黑麦、brachypodium stacei、短柄草。
16.在一种或多种优选方式中,所述的促进籽粒的发育包括:维持籽粒或其胚/胚乳的品质或提高籽粒或其胚/胚乳的品质。
17.在一种或多种优选方式中,所述增加籽粒中zn或fe的含量包括:以所述zn-尼克酰胺转运蛋白将zn、fe或zn-尼克酰胺复合物转运到细胞内;较佳地,所述zn-尼克酰胺转运蛋白表达胚乳基底转移层(细胞);较佳地,所述zn-尼克酰胺转运蛋白表达于细胞膜上。
18.在一种或多种优选方式中,所述增加籽粒中zn或fe的含量包括增加籽粒的胚或胚乳中zn或fe的含量。
19.在一种或多种优选方式中,所述促进籽粒的发育包括促进胚乳的发育;较佳地,包括促进粉质淀粉向硬质淀粉的转化,增加醇溶蛋白的含量。
20.在本发明的另一方面,提供一种zn-尼克酰胺转运蛋白或其编码基因的用途,用作鉴定禾本科植物的籽粒性状的分子标记物;所述性状包括:籽粒中zn或fe的含量,籽粒的发育;其中,所述的zn-尼克酰胺转运蛋白具有seq id no:2所示的氨基酸序列或其保守性序列变体。
21.在本发明的另一方面,提供一种定向选择籽粒性状改良的禾本科植物的方法,所述籽粒性状改良包括:籽粒中zn或fe含量增加,籽粒发育促进;所述方法包括:鉴定测试植物体内zn-尼克酰胺转运蛋白的表达或活性,若该测试植物中zn-尼克酰胺转运蛋白的表达或活性高于(显著高于,如高5%以上、10%以上、20%以上、40%以上、60%以上、100%以上或更高)该类植物(对照植物)中zn-尼克酰胺转运蛋白的表达或活性的平均值,则其为籽粒性状改良的植物;其中,所述的zn-尼克酰胺转运蛋白具有seq id no:2所示的氨基酸序列或其保守性序列变体。
22.在一种或多种优选方式中,所述高表达(或表达高)或高活性(或活性高),是指与同类或同种植物的表达或活性的平均值相比,表达或活性具有统计学意义的提高,如提高
10%、20%、40%、60%、80%、90%或更高。
23.在一种或多种优选方式中,所述低表达(或表达低)或低活性(或活性低),是指与同类或同种植物的表达或活性的平均值相比,表达或活性具有统计学意义的降低,如降低5%、10%、20%、40%、60%、80%、90%或更低。
24.在一种或多种优选方式中,所述“zn或fe含量增加”是指与同类或同种植物的籽粒相比,zn或fe含量有统计学意义地提高,如高3%、5%、10%、20%、40%、60%、80%、90%或更高。
25.在一种或多种优选方式中,所述“籽粒发育促进”是指与同类或同种植物的籽粒相比,籽粒饱满性、粒宽、粒长、粒厚或粒重在统计学上被促进,如促进/提高/增加1%、2%、3%、5%、10%、20%、40%、60%、80%、90%或更高。
26.在本发明的另一方面,提供一种筛选增加禾本科植物籽粒中zn或fe的含量,促进籽粒的发育的物质(潜在物质)的方法,包括:(1)将候选物质加入到表达zn-尼克酰胺转运蛋白的体系中;(2)检测所述体系,观测其中zn-尼克酰胺转运蛋白的表达或活性,若其表达或活性提高(显著提高,如提高5%、10%、20%、40%、60%、80%、90%或更高),则表明该候选物质为可用于增加禾本科植物籽粒中zn或fe的含量、促进籽粒的发育的物质;其中,所述的zn-尼克酰胺转运蛋白具有seq id no:2所示的氨基酸序列或其保守性序列变体。
27.在一种或多种优选方式中,所述体系包括:细胞(培养物)体系、亚细胞(培养物)体系、组织(培养物)体系;所述组织体系包括植物籽粒。
28.在一种或多种优选方式中,所述体系为细胞,(2)中还包括:观测细胞上所述zn-尼克酰胺转运蛋白对于zn、fe或zn-尼克酰胺的转运情况,若所述候选物质能够促进其向胞内的转运作用,则表明该候选物质为可用于增加禾本科植物籽粒中zn或fe的含量、促进籽粒的发育的物质;较佳地,所述的细胞包括胚乳基底转移层细胞或酵母细胞。
29.在一种或多种优选方式中,所述体系为籽粒、胚或胚乳,(2)中还包括:观测籽粒粉质淀粉或醇溶蛋白含量;若籽粒粉质淀粉量降低,醇溶蛋白量增加,则表明该候选物质为可用于增加禾本科植物籽粒中zn或fe的含量、促进籽粒的发育的物质。
30.在一种或多种优选方式中,所述方法还包括:设置不添加所述候选物质的对照组,从而明确分辨测试组中所述zn-尼克酰胺转运蛋白表达或活性、所述zn、fe或zn-尼克酰胺的转运情况或所述籽粒粉质淀粉或醇溶蛋白含量与对照组的差异。
31.在一种或多种优选方式中,所述的候选物质包括(但不限于):针对所述zn-尼克酰胺转运蛋白或其编码基因或其上游或下游蛋白或基因设计的调控分子(如上调剂、小分子化合物基因编辑构建物等。
32.在本发明的另一方面,提供一种植物细胞、组织或器官,其中含有外源的zn-尼克酰胺转运蛋白或其上调剂;所述上调剂包括:与zn-尼克酰胺转运蛋白相互作用、从而提高其表达或活性的上调剂;或过表达zn-尼克酰胺转运蛋白的表达盒或表达构建物(如表达载体)。
33.在一个或多个实施方式中,所述的植物细胞、组织或器官不具有繁殖能力,不是植物繁殖材料,不能直接繁育为植物。
34.本发明的其它方面由于本文的公开内容,对本领域的技术人员而言是显而易见的。
附图说明
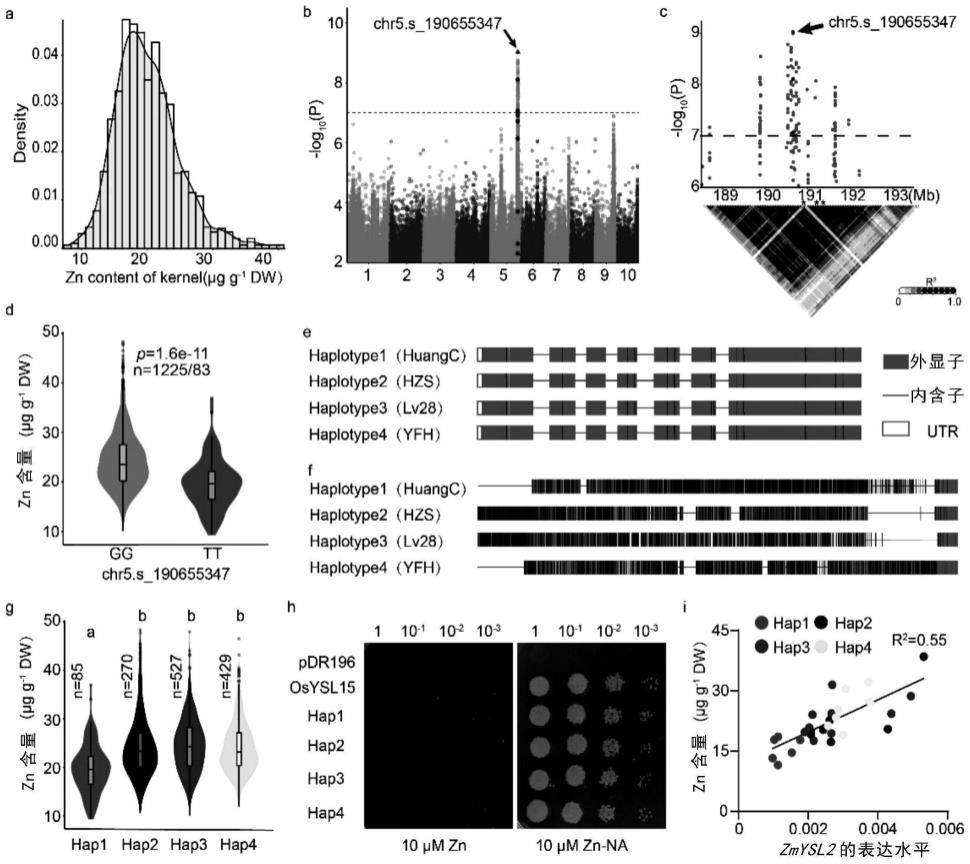
35.图1、玉米籽粒zn含量的全基因组关联分析定位到zmysl2基因。图a是1311份cubic群体子代籽粒zn含量频率分布直方图。图b是gwas分析的曼哈顿图。候选基因内的snp被标成红色,红色箭头指示最显著的snp chr5.s_190655347。图c是候选区间内曼哈顿图和连锁不平衡分析。显示区间为最高snp上下游2mb,候选基因内的snp被标成红色,红色箭头指示最显著的snp。图d是cubic群体被最高snp基因型分型的zn含量差异。图e和图f分别是zmysl2基因区和atg上游5kb启动子区单倍型分析。不同单倍型内的snp分别用不同的颜色表示,其中红色代表t,黑色代表g,蓝色代表c,绿色代表a,灰色代表各个单倍型之间的序列一致。图g是不同单倍型之间zn含量比较。n代表不同单倍型里的子代个数。图h是zmysl2的底物转运活性分析。zn转运能力缺失酵母突变体zrt1zrt2被转进不同的质粒,阳性克隆被点在添加了znso4(10μm)或者zn-na(10μm)的sd/ura培养基上,30℃生长3天。图i是籽粒中zn含量和zmysl2基因表达量之间相关性分析。每个单倍型分别用不同的颜色表示。
36.图2、zmysl2四个突变体criysl2-1,criysl2-2,zmysl2-1和zmysl2-2的表型。图a为四个zmysl2突变体的基本信息。图b-g是杂合的criysl2-1,criysl2-2,zmysl2-1突变体和纯合的zmysl2-2突变体的雌穗表型。白色箭头代表鉴定纯合的criysl2-1,criysl2-2,zmysl2-1和zmysl2-2突变体籽粒,鉴定的对应的野生型籽粒用红色箭头表示。图h-j是用μxrf展示四个纯合突变体籽粒中zn含量以及分布与野生型的差异。不同颜色的虚线代表不同的组织,其中红色虚线是整个籽粒区域,黄色虚线为胚和胚乳所在的区域,绿色虚线为胚所在的区域。图k-s为用icp-ms检测不同突变体与野生型在整个籽粒,胚以及胚乳中zn含量差异。
37.图3、zmysl2的组织定位和亚细胞定位。图a为用qrt-pcr分析zmysl2在各个组织中的表达强度。图b为用qrt-pcr分析zmysl2在籽粒发育过程中在胚,胚乳以及去除betl细胞的胚乳组织中的表达。图c为原位杂交显示zmysl2在授粉后16天b73籽粒中的组织定位。阳性信号显示特异定位在betl细胞。图d为在拟南芥叶片原生质体中瞬时表达zmysl2-gfp,观察其亚细胞定位。图e为用zmysl2抗体免疫荧光实验显示zmysl2特异在betl细胞表达。
38.图4、不同发育时期的纯合criysl2-1,criysl2-2,zmysl2-1和zmysl2-2突变体中zn含量以及分布变化。图a-c为icp-ms检测不同发育时期的纯合criysl2-1,criysl2-2,zmysl2-1和zmysl2-2突变体中zn含量变化。图d-i为用μxrf展示zmysl2-1与其野生型w22籽粒不同发育时期中zn的分布差异。图j-l为授粉后20天的各个突变体与其野生型籽粒不同部位zn含量。基因型鉴定后的籽粒被解剖为不同部位,分别为整个籽粒,胚,胚乳和母体组织。
39.图5、zmysl2过表达材料籽粒zn和fe含量。图a-d用icp-ms分别检测三个独立的pbetl9::zmysl2(a,b)和ubi:zmysl2
b73
(c,d)转基因植株籽粒zn(a,c)和fe(b,d)的含量。图e和f为用μxrf分别检测三个独立的pbetl9::zmysl2(e)和ubi:zmysl2
b73
(f)转基因植株籽粒zn和fe的含量。
具体实施方式
40.本发明人经过广泛的研究,揭示了一种新型的与禾本科植物籽粒性状相关的基因,本发明人将之命名为zn-尼克酰胺转运蛋白基因(ysl2),其为禾本科植物籽粒(包括胚
以及胚乳)zn或fe含量的正调控基因,其功能缺失则会导致产量和品质下降,但是对于植物其他性状没有影响。本发明为植物的性状改良提供了新途径。
41.术语
42.如本文所用,所述的“植物”是具有籽粒、且所述籽粒具有胚、胚乳结构的植物。本领域人员熟知,植物胚或胚乳的组成成分是相似的,具有胚或胚乳结构的植物具有共同的特性,其基因组中存在许多保守的调控基因转录、表达的基因或调节元件,例如一系列调控植物形成胚或胚乳的元件。本发明中,所述的“植物”表达zn-尼克酰胺转运蛋白基因(包括其保守性序列变体或同源物)或包含zn-尼克酰胺转运蛋白基因及其所参与的信号通路。根据本领域的知识,表达zn-尼克酰胺转运蛋白的植物,其内在存在如本发明所主张的作用机制,可以实现如本发明所主张的技术效果。在一些优选方式中,所述的植物为作物,较佳地为禾谷类作物,所述禾谷类作物为具有籽粒(穗粒)的作物。所述的“禾谷类作物”可以是禾本科植物。较佳地,所述的禾本科植物包括:水稻、玉米、高粱、小米、黍、小麦、大麦、燕麦、黑麦、brachypodium stacei、短柄草等。
43.如本文所用,术语“提高”、“改良”或“增强”是相互可以交换的并且在应用含义上应当意指与本文中定义的对照植物相比较,至少2%、3%、4%、5%、6%、7%、8%、9%或10%、优选的至少15%或20%、更优选25%、30%更高的调节。
44.关于“对照植物”,选择合适的对照植物是实验设计的例行部分,可以包括对应的野生型植物或无目的基因的相应转基因植物。对照植物一般是相同的植物物种或甚至是与待评估植物相同或属于同一类的品种。对照植物也可以是因分离而丢失转基因植物的个体。如本文所用的对照植物不仅指完整植物,也指植物部分,包括种子和种子部分。
45.如本文所用,所述的“籽粒”是指植物的果实或种子,在玉米、水稻、小麦、大麦等作物中也称为穗粒。
46.如本文所用,所述的“可操作(性)地连接”或“操作性连接”是指两个或多个核酸区域或核酸序列的功能性的空间排列。例如:启动子区被置于相对于目的基因核酸序列的特定位置,使得核酸序列的转录受到该启动子区域的引导,从而,启动子区域被“可操作地连接”到该核酸序列上。
47.如本文所用,“目的基因”是指本发明所指出的可应用于调控植物性状的新型靶蛋白/靶基因zn-尼克酰胺转运蛋白/基因。
48.如本文所用,术语“特异(性)表达”是指目的基因在特定的时间和/或特定的组织表达。“组织特异性”又称“器官特异性”,在一些调控元件的调控下,基因往往只在某些特定的器官或组织部位表达,并表现出其相关的发育调节的特性。本发明中,所述的“特异表达”是指在植物胚乳基底转移层(betl,basal endosperm transfer layer)特异表达。通常,如果在某组织或器官中mrna以比在其它组织或器官中高至少1.5倍、2倍、3倍或4倍,优选至少高5,更优选至少高10倍,最优选至少高100倍水平被表达,则认为相关基因的表达是组织或器官特异性的。
49.如本文所用,“外源的”或“异源的”是指来自不同来源的两条或多条核酸或蛋白质序列之间的关系。例如,如果启动子与目的基因序列的组合通常不是天然存在的,则启动子对于该目的基因来说是外源的。特定序列对于其所插入的细胞或生物体来说是“外源的”。
50.如本文所用,所述的“非繁殖材料”是指一种生物材料,其不具有借助光合作用,以
水、二氧化碳和无机盐等无机物合成碳水化合物、蛋白质来维系生存的特性。
51.正调控分子
52.本发明人根据通过对玉米cubic(complete diallel design plus unbalanced breeding-like intercross)群体成熟籽粒中锌含量的全基因组关联分析鉴定到一个位于5号染色体上的主效qtl并提名候选基因zmysl2,该基因的编码蛋白定位于细胞质膜且在授粉十天后玉米籽粒的胚乳基底转移层(betl,basal endosperm transfer layer)存在特异表达。单倍型分析表明zmysl2启动子序列变异引起的表达量差异是导致自然群体籽粒中锌含量的变异的原因。xrf(micro-x-ray fluorescence spectrometer)和icp-ms(inductively coupled plasma-mass spectrometry)检测结果表明,与野生型相比,zmysl2突变体成熟籽粒中的锌含量显著降低;在发育过程中突变体的锌被局限于胚乳基底转移层。zmysl2突变体籽粒也呈现出粉质淀粉增多,醇溶蛋白含量降低的表型,说明了锌含量对玉米淀粉品质具有重要的影响。
53.在本发明中,除非特别说明,所述ysl2包括了其同源物(同源蛋白/同源基因)。在一些实施方式中,所述ysl2蛋白为具有seq id no:2所示氨基酸序列的蛋白(多肽);本发明还包括具有与ysl2蛋白相同功能的序列变异形式。
54.蛋白的序列变异形式包括(但并不限于):若干个(通常为1-50个,较佳地1-30个,更佳地1-20个,最佳地1-10个,还更佳如1-8个、1-5个)氨基酸的缺失、插入和/或取代,以及在c末端和/或n末端添加或缺失一个或数个(通常为20个以内,较佳地为10个以内,更佳地为5个以内)氨基酸。任何与所述的ysl2蛋白同源性高(比如与seq id no:2所示的多肽序列的同源性为70%或更高;优选地同源性为80%或更高;更优选地同源性为90%或更高,如同源性95%,98%或99%)的、且具有所述ysl2蛋白相同功能的蛋白也包括在本发明内。
55.来源于玉米以外其它物种的与seq id no:2所示序列的多肽序列的同源性较高、或在同样或相近的信号通路中发挥同样或相近作用的多肽也包括在本发明中。
56.应理解,虽然本发明中优选研究了获自特定物种的ysl2,但是获自其它物种、特别是禾本科植物的与所述ysl2高度同源(如具有70%以上,更特别80%,85%、90%、95%、甚至98%以上序列相同性)的其它多肽或基因也在本发明考虑的范围之内。
57.本发明还涉及编码本发明的ysl2或其序列变异形式的多核苷酸序列。所述的多核苷酸可以是dna形式或rna形式。dna形式包括cdna、基因组dna或人工合成的dna。dna可以是单链的或是双链的。dna可以是编码链或非编码链。编码成熟多肽的编码区序列可以与seq id no:1或seq id no:3所示的编码区序列相同或者是简并的变异体。如本文所用,“简并的变异体”在本发明中是指编码具有seq id no:2序列的多肽,但与seq idno:1或seq id no:3所示的编码区序列有差别的核酸序列。本发明还涉及上述多核苷酸的变异体(变体),其编码与本发明有相同的氨基酸序列的多肽或多肽的片段、类似物和衍生物。
58.本发明也涉及包含所述的多核苷酸的载体,以及用所述的载体或多肽编码核酸经基因工程产生的宿主细胞。
59.本发明中,编码本发明的多肽的多核苷酸序列可插入到重组表达载体中。术语“重组表达载体”指本领域熟知的细菌质粒、噬菌体、酵母质粒、植物细胞病毒、哺乳动物细胞病毒或其他载体。总之,只要能在宿主体内复制和稳定,任何质粒和载体都可以用。表达载体的一个重要特征是通常含有复制起点、启动子、标记基因和翻译控制元件。较佳地,所述表
达载体还可选择性地加入抗性元件、筛选(选择)元件或报告基因元件,如bar、gus。
60.所述的多核苷酸在高等真核细胞中表达时,如果在载体中插入增强子序列时将会使转录得到增强。增强子是dna的顺式作用因子,通常大约有10到300个碱基对,作用于启动子以增强基因的转录。
61.用重组dna转化宿主细胞可用本领域技术人员熟知的常规技术进行。转化植物可使用农杆菌转化或基因枪转化等方法,例如喷洒法、叶盘法、幼胚转化法等。
62.植物改良的应用
63.本发明人通过大量的系统性分析研究,克隆获得ysl2的全长序列并鉴定了其生物学功能,该基因在籽粒(包括胚/胚乳)发育过程中发挥了重要作用。
64.本发明中,首次通过正向遗传学全基因组关联分析的方法鉴定到玉米来源的zn-尼克酰胺转运蛋白基因(ysl2)启动子区的变异引起的表达量的差异是造成玉米自然群体籽粒zn含量变异的原因。通过酵母异源表达体系,发现ysl2能够将zn-na复合物转运到酵母体内。通过时空表达模式的分析,ysl2特异表达在籽粒的胚乳基底转移层细胞,在授粉十天后表达逐渐上调。通过对四个独立的ysl2的突变体的离子表型分析发现,突变体籽粒中zn含量显著降低。通过分别用betl9和ubiquitin的启动子驱动ysl2实现对ysl2的超表达,发现这两种转基因材料中,籽粒zn含量显著提高。
65.这些结果表明,zmysl2可能通过调控锌离子的积累进而影响玉米籽粒发育。通过分别用zmbetl9和zmubiquitin的启动子驱动zmysl2实现对zmysl2的超表达,发现这两种转基因材料中,籽粒zn含量显著提高,其中pubi::zmysl2-1材料籽粒zn含量提高了31.2%,超过玉米籽粒zn强化目标20%,具有极大的应用潜力。本研究发现并证实了zmysl2特异调控锌在籽粒中的转运和积累,进而调控籽粒胚乳淀粉合成和玉米籽粒发育,阐明了其潜在的利用价值和利用途径。
66.基于本发明人的上述新发现,提供了一种ysl2或其上调剂的用途,用于:增加禾本科植物籽粒中zn或fe的含量,促进籽粒的发育。所述增加籽粒中zn或fe的含量包括:以所述zn-尼克酰胺转运蛋白将zn、fe或zn-尼克酰胺复合物转运到细胞内。所述增加籽粒中zn或fe的含量包括增加籽粒的胚或胚乳中zn或fe的含量。所述促进籽粒的发育包括促进胚乳的发育。
67.同时,本发明也提供了一种增加禾本科植物籽粒中zn或fe的含量,促进籽粒的发育的方法,包括:在植物中提高zn-尼克酰胺转运蛋白的表达或活性。
68.应理解,在得知了所述ysl2在禾本科植物籽粒性状调控中的作用后,可以根据实际所需,采用本领域人员熟知的多种方法来调节所述的ysl2的表达或活性,这些方法均被包含在本发明中。
69.可以利用ysl2的表达或活性的上调剂来上调ysl2的活性。所述的ysl2的表达或活性的上调剂包括了促进剂、激动剂、激活剂。所述的“上调”、“促进”包括了蛋白活性的“上调”、“促进”或蛋白表达的“上调”、“促进”。任何可提高ysl2的活性、提高ysl2基因或蛋白的稳定性、上调ysl2基因表达、增加ysl2蛋白的有效作用时间的物质,这些物质均可用于本发明,作为对于上调gras11蛋白或其编码的蛋白有用的物质。它们可以是化合物、化学小分子、生物分子。所述的生物分子可以是核酸水平(包括dna、rna)的,也可以是蛋白水平的。
70.作为一种优选的实施方式,提供一种上调植物中ysl2的表达的方法,所述的方法
包括:将表达ysl2的表达构建物或载体引入植物中。
71.优选地,提供了一种制备转基因植物的方法,包括:(1)将外源的ysl2的编码核酸引物植物器官或组织,获得转化入所述多肽的编码核酸的植物组织或器官。
72.作为一种优选的实例,所述的方法包括步骤:(s1)提供携带表达载体的农杆菌,所述的表达载体含有ysl2的编码核酸;(s2)将植物组织或器官与步骤(s1)中的农杆菌接触,从而使所述多肽的编码核酸转入并且整合到植物细胞的染色体上;(s3)选择出引入所述ysl2的编码核酸的植物组织或器官。
73.鉴于本发明的ysl2的调控作用可发生于籽粒的胚或胚乳中,因此,作为本发明的优选方式,可以以胚或胚乳特异性表达启动子来驱动ysl2的表达。
74.作为分子标记的应用
75.基于本发明人的新发现,本发明提供了适用于鉴定籽粒性状改良的禾本科植物的分子标记,即ysl2。本发明还涉及针对所述ysl2设计的特异性分子标记,以及鉴定策略。
76.因此,本发明提供了一种定向选择或鉴定农艺性状被调节的植物的方法,包括:鉴定测试植物体内ysl2的表达或活性,若该测试植物中ysl2的表达或活性高于该类植物(对照植物)中ysl2的表达或活性的平均值,则其为籽粒品质好(籽粒中zn或fe含量高,籽粒发育好)的植物;或,若该测试植物中ysl2的表达或活性低于该类植物(对照植物)中ysl2的表达或活性的平均值,则其为籽粒性状不理想的植物。
77.根据本发明的新发现,本领域技术人员可以采用任何本领域公知的或正在发展的多种技术来进行核酸序列的分析,这些技术均可被包含在本发明中。所述的方法例如包括但不限于:测序法,pcr扩增法,探针法,杂交法,限制性酶切分析法,等位基因多态性分析法(如溶解曲线法)进行核酸序列的鉴定,等等。
78.本发明的鉴定方法,只需进行pcr反应和/或琼脂糖凝胶电泳,并通过判断相应的pcr产物的长度,就可以准确、快速地判断待测样品的表型或产量,成本低廉,适合于大规模鉴定,而且所需的样品量很少。如果需要,本领域技术人员能够设计出鉴定所述分子标记的引物。
79.获取待测样品的dna的方法是本领域技术人员所熟知的技术,例如可采取传统的酚/氯仿/异戊醇法,或者可采用一些商购的dna提取试剂盒,这类试剂盒是本领域技术人员熟知的。聚合酶链反应(pcr)技术是本领域技术人员熟知的技术,其基本原理是体外酶促合成特异dna片段的方法。本发明的方法可采用常规的pcr技术进行。
80.本发明在分子设计育种及利用基因工程技术进行农作物品种改良等方面具有良好的应用前景。
81.在得知了ysl2的功能和启动子变异以后,可以以其或其启动子序列为分子标记物,来进行植物的定向筛选。也可基于该新发现来筛选通过调节这一机制,从而定向调控产量、胚乳品质的物质或潜在物质。
82.本发明提供了一种筛选增加禾本科植物籽粒中zn或fe的含量,促进籽粒的发育的物质(潜在物质)的方法;该方法包括:(1)将候选物质加入到表达zn-尼克酰胺转运蛋白的体系中;(2)检测所述体系,观测其中zn-尼克酰胺转运蛋白的表达或活性,若其表达或活性提高,则表明该候选物质为可用于增加禾本科植物籽粒中zn或fe的含量、促进籽粒的发育的物质。
83.以蛋白或基因或其上特定的区域作为靶点,来筛选作用于该靶点的物质的方法是本领域人员所熟知的,这些方法均可用于本发明。所述的候选物质可以选自:肽、聚合肽、拟肽、非肽化合物、碳水化合物、脂、抗体或抗体片段、配体、有机小分子、无机小分子和核酸序列等。根据待筛选的物质的种类,本领域人员清楚如何选择适用的筛选方法。
84.检测蛋白与蛋白之间相互作用以及相互作用的强弱可采用多种本领域技术人员熟知的技术,比如gst沉降技术(gst-pull down)、双分子荧光互补实验、酵母双杂交系统或免疫共沉淀技术等。
85.经过大规模的筛选,可以获得一类特异性作用于ysl2,对植物性状改良有调控作用的物质。
86.下面结合具体实施例,进一步阐述本发明。应理解,这些实施例仅用于说明本发明而不用于限制本发明的范围。下列实施例中未注明具体条件的实验方法,通常按照常规条件如j.萨姆布鲁克等编著,分子克隆实验指南,第三版,科学出版社中所述的条件,或按照制造厂商所建议的条件。
87.材料和方法
88.1、全基因组关联分析
89.在cubic群体中,本发明人将该群体在2016年海南收获的籽粒用icp-ms经过log2转化的离子组的含量作为表型,以前期严建兵老师课题组分析得到的高质量的共1180万个snp(最小等位基因频率maf≥0.02)(获自华中农业大学)作为基因型,用tassel 3.0软件里的混合线性模型进行单标记关联分析(即sgwas);为了控制稀有的等位基因的效应并且同时保留其潜在的作用,利用maf≥2%的共1180万个snp作为基因型数据,将snp基因型与群体结构作为固定效应,同时将亲缘关系矩阵作为随机效应,阈值设为1/n,其中n为每个性状实际所用到的snp标记的总数,最终p值小于设定阈值的snp为显著的snp。p值最低的snp为最显著peak snp,每个snp位点所覆盖的物理区间就被定义为该sqtl的区间。
90.2、zmysl2的组织定位和亚细胞定位
91.(1)zmysl2在原生质体中瞬时表达
92.根据zmysl2的cdna序列,以cdna为模板利用高保真酶kodplus(toyobo公司)进行pcr扩增,同时设计相应的引物扩增gfp片段,用overlappcr的方式将zmysl2在n端和gfp片段融合。连接成功pcr产物经电泳回收后克隆到瞬转载体pa7上,阳性克隆进行测序验证正确后保存。
93.(2)瞬转及表达
94.将瞬时表达载体利用peg-ca
2+
的方法转化到原生质体中,利用leica sp8激光共聚焦显微镜进行观察。
95.3、酵母互补试验验证zmysl2将zn-na复合物转运到酵母体内
96.(1)zmysl2酵母表达表达载体的构建
97.根据zmysl2的cdna序列,分别设计去掉终止密码子的特异引物,以cdna为模板利用高保真酶kodplus(toyobo公司)对不同基因型的cdna进行pcr扩增,pcr产物经电泳回收后克隆到酵母表达载体pdr196上。阳性克隆进行测序验证正确后保存。
98.(2)酵母转化
99.将不同单倍型的pdr196-zmysl2质粒,pdr196-zmys1质粒和空载体用peg-lich的
方法转化到zn转运缺失酵母突变体zrt1zrt2中,用缺ura缺陷型培养基筛选。
100.(3)酵母点板实验
101.将转化阳性的酵母用sd-ura的酵母培养基摇到od=1(5ml),1500g,离心5min,用18.2mω去离子水重复清洗三次,梯度稀释5个梯度后取5μl点在添加10μm zn和10μm zn-na的板子上,30℃生长3天之后观察酵母生长情况。
102.4、icp-ms检测离子含量
103.(1)样品的选择与处理
104.对于群体籽粒的样品,选取每个子代三个生长一致的成熟雌穗,每个棒子取3粒种子混在一起磨成均匀的干粉,称取5mg左右作为待测样品。对于突变体和野生型成熟籽粒的不同部位,用塑料刀解剖之后分别称干重,每一粒种子的整体,胚或者胚乳作为一个样品。对于突变体和野生型发育过程中籽粒的不同部位,用塑料刀解剖之后分别称湿重,每一粒种子的整体,胚或者胚乳作为一个样品。
105.(2)将玉米籽粒样品装入玻璃试管中,置于65℃烘箱中过夜烘干。次日,拿出玻璃试管,冷却至室温。
106.(3)打开通风橱,向每个玻璃试管中加2ml的含in(铟)元素的浓硝酸,放入石墨炉,调至115℃处理4h。用超纯水补至10ml(此步骤需考虑加热挥发的量),轻轻混合均匀。吸取1ml混合液至一套新的玻璃试管中。
107.(4)再加入1ml的含in(铟)元素的浓硝酸,放入石墨炉,调至115℃处理4h。
108.(5)再次用超纯水补至10ml(此步骤需考虑加热挥发的量),轻轻混合均匀,向特定测样板中按顺序加入1.7ml混合液,使用本组icp-ms(nexion300d;perkinelmer)仪器,即可对样品中的离子含量测定。
109.5、玉米籽粒离子分布与含量分析
110.μxrf观察玉米籽粒离子分布与含量,具体实施过程如下:
111.(1)用陶瓷刀将玉米籽粒切开,切面朝上粘在xrf检测专用的无杂质胶带上,打开m4 tornado仪器和软件。
112.(2)将粘有待测样品的胶带小心放置于m4 tornado载物台的中心位置,关闭盖子,适当地调整载物台高度,左右移动载物台位置,使样品呈现在镜头的正中央。
113.(3)框选需要扫描的区域,在10倍镜下调整焦距后,再在100倍镜下调整焦距。
114.(4)工作条件设置为:工作电流600μa(对轻元素扫描时为330μa),电压50kv选择所需要的filter,选择步长25μm,单点扫描停顿时长为20ms,设定关注的zn和fe元素以便一边扫描一边观察,开始扫描。
115.(5)扫描完成后,保存扫描结果至个人文件夹,及时关闭机器和软件。
116.6.ysl2序列
117.zmysl2核苷酸序列(seq id no:1):
118.atgccgaggaccacaactccacaaggcttgctcatttataatggccttgtggagtccgaggcactgattgatagtagttcttgctctagccgcaccgccatggagatcaaccccgcggacgagatcgacaagtgcgaggccggcggcggcgcggagccgcaacctgcagatccggagcccgatgccgccgcggcggtcgagcagcagcagccgtggcgcgagcagctgacggcgcggggcttggtggccgcggcgctgatcgggtccatgtacaccgtcgtcgtcatgaagctgaacctgacgacggggttcgtgcccacgatgaacgtgtccgccgcgctgctggcgttcctggcgctccgcggctggacg
ggcgcgctgtcccgcctgggcgtcgccggcgccaggcccttcacgcggcaggagaacaccgtcgtccagacctgcgccgtcgcgtgctactcgctagcgctctgcgggttcgggtcgttcatgctgggcctgaacaggaggacgtacgaggcggccggagtgaacacgccgggaaacgtgccgggtagcgtcaaggagccaggattcgggtggatagccggcttcctcgtcgccagtagcttcggcggcctgctcaccttgatccccctcagaaaggctttggtcatcgactacaagctgacgtacccaagcgggactgcaactgctgttctaataaacgggttccacaccccccaaggagacaagaatgcagagaagcaagtacgtgcatttctgaagtactttgggatcagcttcctatggagcttcttccagtggttctacaccggtggcaacgcctgcggcttcgctcaattccctacgtttggcctcaaggcctggaaacactcgttcttcttcgatttcagcatgacgtacgtcggcgccgggatgatctgcccgcaccttgtcaacctctccgccctcctcggggcggtcctctcctggggaatcctgtggccactcatcagcagacgcaagggctactggtaccctgcagacgtaccggagagcagcatgaccagcttgtatggttacaagtccttcgtgtgcatagctctgatcatgggtgatggcatctaccacctcgtcaaggtcctcggcgtcaccgccaagagcctgcatgagcgctcaaagctcagacgcagcaacaaccgagtggccgacgaggacaacgcggccgcgatcgacagcctgcgccgcgacgaggtcttcaaccgagactccatcccggcgtggctggcgtacgctgggtacgccgtcacgagcgccgtcgcggcggtcgccatcccggcgatgttccggcaggtgcggtggtactacgtggtcgccgcgtacgcgctggcgcccgtgctgggcttctccaactcctacggcgccgggctcacggacatcaacatgggctacagctacggccagctcgcgctcttcgtcctcgcggcgtgggccggccgcgacgacggcgtcgtggccggcctcgtcggctgcggcctggtgaagcagctggtgctcatctccgccgacctcatgcacgacttcaagacggcgcacctcaccctgacgtcgccgcgctccatgctcgccgcgcaggccgtcggcgccgccatgggctgcgtcgtcacgccgctcacgtttctcctcttctacagggcgttcgacgtcggcaaccccgacgggtactggaaagcgccgttcgcgctcatctaccgcaacatggccctcctgggagcccagggcttctcggcgctgcccacgcactgcctgccgctgtccgcggggttcttcgcgctcgccgtgctcgccaacgtgatgaaggacgtcctgccgccgcgctacgcgaggtacgtgcccctgccgacggcgatggccgtgccgttcctcgtcggcgccagcttcgccatcgacatggtggtggggaccgtggtggtcttcgcctggcactggattgacggcaacgagaccacgttgctcgtgccggcggtggcgtcaggcctcatctgcggtgacggcgtatggactttcccttcgtcgttgctttctctggccaagatcaagccacccatctgcctgaaatttacacctggaagctag
119.zmysl2蛋白序列(seq id no:2)
120.mprtttpqglliynglvesealidssscssrtameinpadeidkceagggaepqpadpepdaaaaveqqqpwreqltarglvaaaligsmytvvvmklnlttgfvptmnvsaallaflalrgwtgalsrlgvagarpftrqentvvqtcavacyslalcgfgsfmlglnrrtyeaagvntpgnvpgsvkepgfgwiagflvassfgglltliplrkalvidykltypsgtatavlingfhtpqgdknaekqvraflkyfgisflwsffqwfytggnacgfaqfptfglkawkhsfffdfsmtyvgagmicphlvnlsallgavlswgilwplisrrkgywypadvpessmtslygyksfvcialimgdgiyhlvkvlgvtakslhersklrrsnnrvadednaaaidslrrdevfnrdsipawlayagyavtsavaavaipamfrqvrwyyvvaayalapvlgfsnsygagltdinmgysygqlalfvlaawagrddgvvaglvgcglvkqlvlisadlmhdfktahltltsprsmlaaqavgaamgcvvtpltfllfyrafdvgnpdgywkapfaliyrnmallgaqgfsalpthclplsagffalavlanvmkdvlppryaryvplptamavpflvgasfaidmvvgtvvvfawhwidgnettllvpavasglicgdgvwtfpssllslakikppiclkftpgs
121.实施例1、全基因组关联分析及zmysl2基因的确定
122.本实施例中,利用全基因组关联分析进行基因筛选。本发明人的分析结果如图1。
123.1311份cubic群体子代籽粒zn含量频率分布直方图如图1a。
124.gwas分析的曼哈顿图如图1b。候选基因内的snp被标成红色,红色箭头指示最显著的snp chr5.s_190655347。
125.候选区间内曼哈顿图和连锁不平衡分析如图1c。显示区间为最高snp上下游2mb,候选基因内的snp被标成红色,红色箭头指示最显著的snp。
126.cubic群体被最高snp基因型分型的zn含量差异如图1d。
127.zmysl2基因区和atg上游5kb启动子区单倍型分析分别如图1e和图1f。不同单倍型(haplotype)内的snp分别用不同的颜色表示,其中红色代表t,黑色代表g,蓝色代表c,绿色代表a,灰色代表各个单倍型之间的序列一致。其中huangc为黄玉米c,hzs、lv28及yfh为已知玉米品种。
128.不同单倍型之间zn含量比较结果如图1g。n代表不同单倍型里的子代个数。
129.zmysl2的底物转运活性分析如图1h。zn转运能力缺失酵母突变体zrt1zrt2被转进不同的质粒(pdr196质粒,分别插入osysl15、黄玉米c,hzs、lv28及yfh来源的zmysl2),阳性克隆被点在添加了znso4(10μm)或者zn-na(10μm)的sd/ura培养基上,30℃生长3天。结果显示,zn-na复合物能被zmysl2转运到酵母体内。
130.籽粒中zn含量和zmysl2基因表达量之间相关性分析如图1i。每个单倍型分别以不同的颜色表示。
131.根据图1,应用玉米籽粒zn含量的全基因组关联分析,本发明人定位到zn-尼克酰胺转运蛋白基因(zmysl2基因),其核苷酸序列如seq id no:1,其氨基酸序列如seq id no:2。其是位于5号染色体上的主效qtl。
132.单倍型分析表明,zmysl2启动子序列变异引起的表达量差异是导致自然群体籽粒中zn含量的变异的原因。
133.实施例2、zmysl2突变体及其表型
134.本发明人分析了zmysl2的突变体及其表型。
135.本发明人建立了zmysl2的四个突变体criysl2-1,criysl2-2,zmysl2-1和zmysl2-2,建立方法如下(图2a):
136.criysl2-1及criysl2-2:以玉米kn5585为背景材料,通过crispr技术实施突变,第五外显子删除。
137.zmysl2-1:以玉米w22为背景材料,从美国uniformmu插入突变体库中获得。
138.zmysl2-2:以玉米w64a为背景材料,通过ems诱变技术实施突变,获得发生a113v的位点的突变。
139.四个突变体criysl2-1,criysl2-2,zmysl2-1和zmysl2-2的表型如图2。
140.观测杂合的criysl2-1,criysl2-2,zmysl2-1突变体和纯合的zmysl2-2突变体的雌穗表型。如图2b-g,白色箭头代表鉴定纯合的criysl2-1,criysl2-2,zmysl2-1和zmysl2-2突变体籽粒,鉴定的对应的野生型籽粒用红色箭头表示。与野生型相比,突变体的籽粒表型发生干瘪皱缩,蛋白含量降低的变化);因此突变体籽粒的品质降低。
141.用μxrf展示四个纯合突变体籽粒中zn含量以及分布与野生型的差异。如图2h-j,不同颜色的虚线代表不同的组织,其中红色虚线是整个籽粒区域,黄色虚线为胚和胚乳所在的区域,绿色虚线为胚所在的区域。与野生型相比,突变体的胚未发生明显形态改变,突变体的胚乳干瘪皱缩。图中也显示,胚乳白色区域增多表明粉质淀粉增多、透明的浅红色区域(硬质淀粉部位)呈现减少。因此,突变体籽粒也呈现出粉质淀粉增多,醇溶蛋白含量降低的表型,说明了锌含量对玉米淀粉品质具有重要的影响。
142.用icp-ms检测不同突变体与野生型在整个籽粒,胚以及胚乳中zn含量差异,如图2k-s,可见突变体中,整个籽粒、胚以及胚乳的zn含量均发生显著的降低。
143.因此,zmysl2为籽粒、胚以及胚乳的zn含量的正调控基因;也发挥着调节籽粒的粉质淀粉降低及醇溶蛋白提高作用。
144.实施例3、zmysl2的组织定位和亚细胞定位
145.本实施例中,进一步分析zmysl2的组织定位和亚细胞定位。结果如图3。
146.用qrt-pcr分析zmysl2在各个组织中的表达强度。如图3a,该基因在授粉10天后玉米籽粒的胚乳基底转移层(betl)存在特异表达。在授粉后天数(dap)为16天的种子中,zmysl2表达强度最高;授粉12天后的种子中表达强度为其次;在授粉后的其它时期中以相对低的水平表达;在根、叶等组织(图中以授粉后1天的组织进行测定)中以相对低的水平表达。
147.用qrt-pcr分析zmysl2在籽粒发育过程中在胚,胚乳以及去除胚乳基底转移层(betl)细胞的胚乳组织中的表达。如图3b,授粉10天后,胚乳中zmysl2含量呈极为显著地增加。
148.通过原位杂交显示zmysl2在授粉后16天b73籽粒中的组织定位。如图3c,阳性信号显示zmysl2特异定位在betl细胞。
149.在拟南芥叶片原生质体中瞬时表达zmysl2-gfp,观察其亚细胞定位。如图3d,可见该基因的编码蛋白定位于细胞质膜。
150.用zmysl2抗体免疫荧光实验显示,zmysl2蛋白特异在玉米籽粒胚乳基底转移层(betl)细胞表达,如图3e。
151.实施例4、zmysl2突变体中zn含量以及分布变化
152.本实施例中,分析不同发育时期的纯合criysl2-1,criysl2-2,zmysl2-1和zmysl2-2突变体中zn含量以及分布变化。
153.icp-ms检测不同发育时期的纯合criysl2-1,criysl2-2,zmysl2-1和zmysl2-2突变体中zn含量变化。结果如图4a-c,与野生型相比,criysl2-1,criysl2-2,zmysl2-1和zmysl2-2突变体中zn含量均显著降低。
154.用μxrf展示zmysl2-1与其野生型w22籽粒不同发育时期中zn的分布差异。结果如图4d-i,与野生型相比,criysl2-1,criysl2-2,zmysl2-1和zmysl2-2突变体中呈现zn含量均显著降低导致的表型。
155.检测授粉后20天的各个突变体与其野生型籽粒不同部位zn含量。结果如图4j-l,基因型鉴定后的籽粒被解剖为不同部位,分别为整个籽粒、胚、胚乳和母体组织。可见,与野生型相比,criysl2-1,criysl2-2,zmysl2-1和zmysl2-2突变体中籽粒、胚、胚乳中zn含量均发生显著降低;而母体组织中zn含量则显著高。
156.上述结果说明,zmysl2发生突变的突变体中,zn滞留于母体组织,未被转运到籽粒、胚、胚乳中。因此,ysl2与zn的定向转运(从母体组织向籽粒、胚、胚乳)密切相关。
157.xrf和icp-ms检测结果表明,与野生型相比,zmysl2突变体成熟籽粒中的锌含量显著降低;在发育过程中突变体的锌被局限于胚乳基底转移层。
158.实施例5、zmysl2过表达材料籽粒zn和fe含量
159.本实施例中,分析zmysl2过表达材料籽粒zn和fe含量。建立了如下过表达转基因
植株:
160.pbetl9::zmysl2:将seq id no:1所示核苷酸序列的zmysl2插入到具有玉米基底胚乳转移层9(betl9)启动子的ptf102质粒(获自华中农业大学)的多克隆位点中。所获得的重组质粒引入到kn5585玉米材料中。
161.ubi:zmysl2
b73
:将seq id no:1所示核苷酸序列的zmysl2插入到具有ubi启动子的prozmubi::eyfp质粒(获自华中农业大学)的多克隆位点中,由ubi启动子驱动表达。所获得的重组质粒引入到kn5585玉米材料中。
162.用icp-ms分别检测三组独立的pbetl9::zmysl2(图5a,b)和ubi:zmysl2
b73
(图5c,d)转基因植株籽粒zn(图5a,c)和fe(图5b,d)的含量。
163.可见,与野生型相比,pbetl9启动子驱动的zmysl2过表达材料的籽粒中zn含量呈现上升。betl特异高表达的启动子驱动能造成zn含量的提高证明了zmysl2能将zn-na转运到籽粒中。
164.ubi启动子驱动的zmysl2过表达材料的籽粒中zn和fe含量均呈现上升,其中pubi::zmysl2-1材料籽粒zn含量提高了31.2%。ubi启动子驱动zmysl2的异位表达,使得籽粒中zn和fe都升高。
165.用μxrf分别检测三组独立的pbetl9::zmysl2(图5e)和ubi:zmysl2
b73
(图5f)转基因植株籽粒zn和fe的含量。可见,该结果与测离子结果对应,与野生型相比,pbetl9启动子驱动的zmysl2过表达材料的籽粒中zn含量呈现上升。ubi启动子驱动的zmysl2过表达材料的籽粒中zn和fe含量均呈现上升。
166.总结
167.1、本发明通过系统研究,首次提供了zn-na复合物转运蛋白zmysl2的生物学功能。
168.2、通过正向遗传学全基因组关联分析鉴定到zmysl2的表达量的变异是引起玉米群体籽粒zn含量变异的原因(图1)。
169.3、通过xrf和icp-ms方法检测zmysl2四个独立突变体籽粒zn和fe的含量和分布变化,确定zmysl2参与控制籽粒zn含量(图2)。
170.4、利用瞬转体系研究了zmysl2的亚细胞定位,利用荧光定量pcr,原位杂交和蛋白免疫荧光的方法研究了zmysl2的表达模式(图3)。
171.5、通过xrf和icp-ms方法检测zmysl2突变体和野生型材料籽粒发育过程中zn的分布和含量变化,确定了突变体籽粒的zn因为zmysl2的缺失被阻断在betl细胞之外(图4)。
172.6、通过xrf和icp-ms方法检测zmbetl9和zmubiquitin的启动子驱动zmysl2的超表达材料籽粒zn和fe的含量,发现这两种转基因材料中,籽粒zn含量均显著提高,其中pubi::zmysl2-1材料籽粒zn含量提高了31.2%,超过玉米籽粒zn强化目标20%。进一步验证了zmysl2的功能,也说明了zmysl2的应用价值(图5)。
173.以上所述实施例仅表达了本发明的几种实施方式,其描述较为具体和详细,但并不能因此而理解为对本发明专利范围的限制。应当指出的是,对于本领域的普通技术人员来说,在不脱离本发明构思的前提下,还可以做出若干变形和改进,这些都属于本发明的保护范围。因此,本发明专利的保护范围应以所附权利要求为准。同时,在本发明提及的所有文献都在本技术中引用作为参考,就如同每一篇文献被单独引用作为参考那样。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1