发现治疗靶标的方法与流程
交叉引用
本申请要求在2012年9月5日提交的名称为“发现治疗靶标的方法”的美国临时申请号61/696,873和在2013年8月29日提交的名称为“免疫特征分析:通向早期诊断和健康监测的途径”的美国申请号14/014,168的权益,这些专利申请通过引用整体并入本文。本说明书中提及的所有出版物、专利和专利申请均通过引用并入本文,其程度如同特别且单独地指出每个单独的出版物、专利或专利申请通过引用而并入。
背景技术:
诸如dna、rna、蛋白质、抗体和肽微阵列等高通量技术通常用于调查药物治疗、疾病、转基因动物等中的差异。对从此类高通量技术获得的数据的解读并非简单的过程:需要创建、开发和优化特殊的算法用于分析。此外,当前技术的局限性阻碍了其应用于靶标鉴定筛选。
技术实现要素:
在一些实施方案中,本发明包括一种筛选治疗靶标的方法,该方法包括:a)使肽阵列与来自具有感兴趣的已知状况的个体的第一生物样品接触;b)检测所述第一生物样品中的抗体与所述肽阵列的结合以获得第一免疫特征谱(immunosignatureprofile);c)使肽阵列与来源于没有已知状况的个体的对照样品接触;d)检测所述对照样品中的抗体与所述肽阵列的结合以获得第二免疫特征谱;e)将所述第一免疫特征谱与所述第二免疫特征谱进行比较,并鉴定与所述第二免疫特征谱相比在所述第一免疫特征谱中结合更少或更多抗体的差异结合的肽;以及f)将与所鉴定的差异结合的肽相对应的蛋白质鉴定为针对感兴趣的状况的治疗靶标。
在一些实施方案中,本发明包括一种鉴定疫苗靶标的方法,该方法包括:a)使肽阵列与来自具有感兴趣的已知状况的个体的第一生物样品接触;b)检测所述第一生物样品中的抗体与所述肽阵列的结合以获得第一免疫特征谱;c)使肽阵列与来源于没有已知状况的个体的对照样品接触;d)检测所述对照样品中的抗体与所述肽阵列的结合以获得第二免疫特征谱;e)将所述第一免疫特征谱与所述第二免疫特征谱进行比较,并鉴定与所述第二免疫特征谱相比在所述第一免疫特征谱中结合更少或更多抗体的差异结合的肽;以及f)将与所鉴定的差异结合的肽相对应的蛋白质鉴定为针对感兴趣的状况的疫苗靶标。
在一些实施方案中,本发明包括一种鉴定针对癌症的治疗靶标的方法,该方法包括:a)使肽阵列与来自患有感兴趣的已知癌症的个体的第一生物样品接触;b)检测所述第一生物样品中的抗体与所述肽阵列的结合以获得第一免疫特征谱;c)使肽阵列与来源于没有已知癌症的个体的对照样品接触;d)检测所述对照样品中的抗体与所述肽阵列的结合以获得第二免疫特征谱;e)将所述第一免疫特征谱与所述第二免疫特征谱进行比较,并鉴定与所述第二免疫特征谱相比在所述第一免疫特征谱中结合更少或更多抗体的差异结合的肽;以及f)将与所鉴定的差异结合的肽相对应的蛋白质鉴定为针对感兴趣的癌症的靶标。
在一些实施方案中,本发明包括一种鉴定针对自身免疫病症的治疗靶标的方法,该方法包括:a)使肽阵列与来自患有感兴趣的已知自身免疫病症的个体的第一生物样品接触;b)检测所述第一生物样品中的抗体与所述肽阵列的结合以获得第一免疫特征谱;c)使肽阵列与来源于没有已知自身免疫病症的个体的对照样品接触;d)检测所述对照样品中的抗体与所述肽阵列的结合以获得第二免疫特征谱;e)将所述第一免疫特征谱与所述第二免疫特征谱进行比较,并鉴定与所述第二免疫特征谱相比在所述第一免疫特征谱中结合更少或更多抗体的差异结合的肽;以及f)将与所鉴定的差异结合的肽相对应的蛋白质鉴定为针对感兴趣的自身免疫病症的靶标。
附图说明
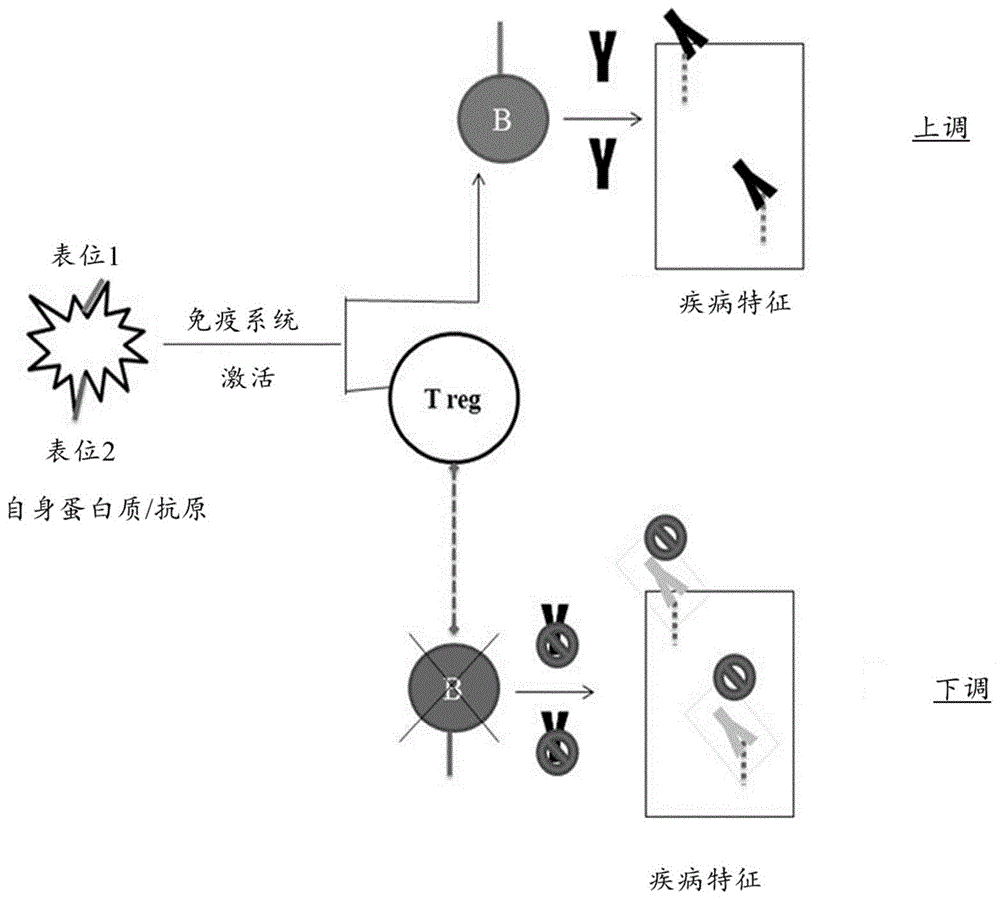
图1为显示自身蛋白质/抗原如何能在随机序列肽微阵列中引起免疫特征的上调和下调的途径。
图2为盒型图,其显示了2组i型糖尿病患者在具有高和低gad-65rip滴度时的13种肽的强度。
图3为免疫特征阵列上的肽强度与抗gad滴度之间的关系的线性回归分析。
图4为鉴定ksfhgrviqdvvgepyggsc(seqidno.1)为感兴趣的肽。a图为示出了野生型小鼠与转基因小鼠的归一化强度的生物信息学平均值之间的肽关系的图。b图为线图,其中将单个小鼠的归一化的强度绘制为线图。b图中的线代表各个单个肽。
图5对应于肽ksfhgrviqdvvgepyggsc(seqidno.1)与排在前两位的小鼠蛋白质组候选物的blast比对。a图对应于最可能的匹配即p190rhogap的blast比对。b图对应于tubby候选基因的blast比对。
图6为示出了可与本发明的示例实施方案结合使用的计算机系统的第一示例架构的框图。
图7为示出了可与本发明的示例实施方案结合使用的计算机网络的示意图。
图8为示出了可与本发明的示例实施方案结合使用的计算机系统的第二示例架构的框图。
图9为本发明的免疫特征分析(immunosignaturing)系统的组件的示意图。
图10示出了具有不同肽密度的本发明的示例性阵列。
图11,a图示出了噬菌体展示文库。b图示出了肽微阵列。
图12为免疫特征肽与pa-x移码序列和合成的平铺肽(tiledpeptide)的guitope比对。a图示出了具有来自免疫特征的两种肽的翻译的pa-x移码序列。相同的氨基酸对于epme肽加下划线,而对于pamk肽加粗。用星号(*)指示相似的氨基酸残基,而用垂直线(|)指示相同的残基。b图示出了具有突出显示的残基的平铺肽。
图13为显示出预测的表位越来越多地被感染流感(h1n1)a/pr/8/34后的小鼠的血清抗体所结合的图。
图14为经免疫的小鼠在采用流感a/pr/8/34进行致死攻击后的结果的图示。a图为相对于起始体重的平均每日百分比的图示。b图为攻击后存活的小鼠的百分比的图示。
图15为对免疫应答的传统测量的图示。a图为针对灭活prb(2006-2007和2007-2008季节性疫苗,根据终点滴度)测量的抗原特异性循环igg的量的图示。b图至e图分别示出了il-2、il-4、il-5、inf-γ和il-12的浓度。
图16示出了活的和灭活的流感a/pr/8/34的免疫特征的比较。a图是散点图,其中活pr8在x-轴上,而灭活pr8在y-轴上。b图为示出了显著不同的肽的重叠的维恩图。c图示出了通过pca分析测定的个体间免疫应答的差异。d图示出了重叠,其中对前两个主分量绘图,并且将个体按疫苗进行颜色区分。
图17说明了免疫特征可区分较弱的灭活疫苗与较强的疫苗。a图示出了主分量间的差异。b图为示出了所列项之间的重叠的维恩图。
图18为在免疫特征验证研究中收集的数据的图示,其表明抗体与na195-219的交叉反应性在保护季节性疫苗接受者免受pr8影响方面是重要的。a图示出了在pr8攻击后存活或死亡的2006-2007疫苗接受者的全病毒elisa终点滴度。b图为接受灭活疫苗的所有个体间的差异的图示。c图为y-轴上的guitope得分和x-轴上的氨基酸的图示。d图为来自合并的血清的抗体反应性的条形图。
图19示出了对随机肽阵列上的免疫特征的解读,以确定被每种疫苗所识别的表位。
图20为本发明在靶标鉴定筛选中的一种应用的总体示意图。
具体实施方式
新出现的病原体、流行性疾病和越来越多的抗生素抗性细菌正在推动对新的药物和疫苗靶标的需求。对新的治疗靶标的探寻已经采用了蛋白质组学或大规模pcr筛选,并且此类技术通常用于调查药物治疗、疾病、转基因动物等中的差异。然而,这些方法在靶标鉴定筛选中的应用常常受到限制。
在针对例如癌症、心脏病、糖尿病、大流行性疾病、多种慢性状况、自身免疫状况、新出现的病原体和越来越多的抗生素抗性细菌的药物开发中的挑战,正在推动对新药和疫苗靶标鉴定的需求。对发现基因组中编码的新型蛋白质,例如发现人类病原体中编码的新型蛋白质的兴趣日渐提高。然而,现有的发现方法并非经济有效的、实用的、可靠的和/或一致的。此外,对于开发如下的测定存在迫切需求:该测定例如通过鉴定与那些新开发的蛋白质结合的表位,可容易地表征新发现的蛋白质。
本文公开的方法和装置还能够个体化或定制针对患有状况(包括癌症、心脏病、糖尿病、自身免疫病症和感染(包括病原性感染和微生物感染))的个体的治疗性处理。可鉴定对于个体可能是唯一的、突出的或可用的针对状况的靶标,并且针对该靶标开发治疗性处理。例如,可采用本文公开的方法和装置来检测由于状况而可能上调或下调的靶标,尤其在存在遗传多态性或疾病起源的变化的情况下。因此,可利用本文公开的方法和装置来实现开发针对特定个体的治疗方案。
免疫系统持续监测自身和非自身蛋白质,并且当遇到时,免疫系统可在1周内将来自被检测为异常的分子靶标的信号扩增1011倍。这提示免疫系统的自然应答可提供对受试者体内分子(例如,新型蛋白质)的存在的容易获取的读出。通过比较健康个体和患病个体的血清免疫特征,可在计算机中使用疾病所共有的肽来预测病变细胞中的潜在移码或其他新型蛋白质。开发具有高得多的多样性的肽阵列可促进该分析。如果广泛可用,可考虑针对新型蛋白质对所有病原体和慢性疾病的普通血清学重新评估。
免疫特征分析微阵列基于抗体的复杂混合物与肽阵列的结合。它依赖于抗体与肽的多对多结合。每种肽可结合多种抗体,并且每种抗体可结合多种肽。
对免疫系统活性的测量可鉴定至少一种抗体的存在、至少一种抗体的缺失、多种抗体的上调和/或多种抗体的下调。所有这些活性可包含可利用例如免疫特征分析方法测量、表征和破译的健康变化的体液免疫代理(proxy)。
免疫特征分析是对受试者中的循环抗体的动力学进行量化的基于阵列的技术。循环抗体的动力学包括来自受试者系统的一种抗体或多种抗体的存在和缺失。该方法基于个体的抗体谱对异常细胞发育的敏感性。甚至少量的引发癌细胞或其他病变细胞可引发可在1周内扩增1011倍的b细胞应答。
在一些实施方案中,一种破译免疫特征的方法包括以下步骤:(a)使肽阵列与来自患有感兴趣的病症的个体的血清接触;(b)检测血清中的抗体与阵列上的肽的结合以产生疾病免疫谱;(c)将疾病免疫谱与正常对照进行比较并基于以下一项或两项来鉴定差异结合的肽:(i)与正常对照相比在疾病免疫谱中结合较多抗体的肽;和(ii)与正常对照相比在疾病免疫谱中结合较少抗体的肽;其中与差异结合的肽相对应的蛋白质为针对感兴趣的疾病的治疗靶标。
通过在肽阵列上展开抗体组库(免疫特征分析),并将疾病与正常对照(包括但不限于在相同实验条件下与相同阵列接触的非疾病血清)进行比较,可鉴定步骤(c)(i)和(c)(ii)中的反应性肽以确定正在与抗体反应的蛋白质。例如,可利用信息学方法来鉴定肽。在信息学无法鉴定推定的匹配的情况下,例如在非连续表位的情况下,可使用信息性的(informative)肽作为亲和试剂来纯化反应性抗体。而后可在标准免疫学技术中使用纯化的抗体来鉴定靶标。
可使用任何合适的肽阵列,在该肽阵列上将肽固定于基底。在一些实施方案中,所述阵列包含500-1,000,000个肽、500-500,000个肽、500-250,000个肽、500-100,000个肽、500-50,000个肽或500-10,000个肽。在一些实施方案中,所述肽为8-35、12-35、15-25、10-30或9-25个氨基酸的长度。在一些实施方案中,随机选择所述肽的氨基酸序列。在一些实施方案中,预先限定微阵列中存在的氨基酸的模式,并且该阵列不是随机肽阵列。
在一些实施方案中,所述肽的氨基酸序列与已知蛋白质具有小于90%的序列同一性。在一些实施方案中,阵列上的肽之间的平均间距为约2-15nm、约7-12nm、约8-11nm、约9nm、约2-4nm或约3nm。
如本文所用的,术语“基底”是指将肽固定于其上的任何类型的固体支持体。基底的实例包括但不限于:微阵列;珠;柱;光纤;擦拭物;硝化纤维;尼龙;玻璃;石英;重氮化的膜(纸或尼龙);硅酮;聚甲醛;纤维素;醋酸纤维素;纸;陶瓷;金属;准金属(metalloids);半导体材料;涂覆的珠;磁性颗粒;塑料,诸如聚乙烯、聚丙烯和聚苯乙烯等;形成凝胶的材料;硅酸盐;琼脂糖;聚丙烯酰胺;甲基丙烯酸甲酯聚合物;溶胶凝胶;多孔聚合物水凝胶;纳米结构化表面;纳米管(如碳纳米管);以及纳米颗粒(如金纳米颗粒或量子点)。当与基底结合时,肽可直接连接至支持体或通过连接体附接至表面。因此,可使用本领域中已知的方法来衍生化固体基底和/或肽以促进肽与固体支持体的结合,只要该衍生化不消除对肽与血清中的抗体之间的结合的检测。
其他分子如参照或对照分子也可任选地固定在基底上。用于将各种类型的分子固定于多种基底上的方法是本领域技术人员所熟知的。很多种材料可用于固体表面。多种不同的材料可用于制备支持体以获得各种性质。例如,蛋白质(例如,牛血清白蛋白)或大分子的混合物(例如,denhardt溶液)可用于最小化非特异性结合、简化共价偶联和/或提高信号检测。
可使用本领域已知的技术从个体中获得血清。个体可以是患有任何感兴趣的病症的个体,所述病症包括但不限于癌症、糖尿病、感染、动脉粥样硬化、心血管心脏病、中风和神经障碍如帕金森病和阿尔茨海默(alzheimer)病。
肽阵列可在任何合适的条件下与血清接触以促进血清中的抗体与阵列上固定的肽的结合。因此,本发明的方法不受所采用的任何特定类型的结合条件的限制。此类条件将根据所使用的阵列、基底的类型、基底上排列的肽的密度、结合相互作用的期望严格性和结合溶液中竞争材料的性质而变化。在一个优选的实施方案中,所述条件包括从可寻址阵列中去除未结合的抗体的步骤。确定对该步骤的需要以及用于该步骤的合适的条件完全在本领域的技术水平内。
相似地,任何合适的检测技术可以在本发明的方法中使用,其检测血清中的抗体与阵列上的肽的结合以产生疾病免疫谱。在一个实施方案中,任何类型的可检测标记物可用于标记阵列上的肽,包括但不限于放射性同位素标记物、荧光标记物、发光标记物和电化学标记物(即,具有不同的电极中点电位的配体标记物,其中检测包括检测标记物的电位)。或者,例如可使用可检测地标记的第二抗体来检测结合的抗体。
对来自可检测标记物的信号的检测完全在本领域的技术水平内。例如,荧光阵列读取仪是本领域熟知的,用于记录基底上的电位的仪器也是本领域熟知的(关于电化学检测,参见,例如,j.wang(2000)analyticalelectrochemistry,vol.,第2版,wiley-vch,newyork)。
对照可以是任何合适的对照。在一个实施方案中,对照包含在相同的实验条件下与相同的阵列接触的非疾病血清。疾病免疫谱与正常对照的比较以及差异结合的肽的鉴定可通过任何合适的技术进行。如在实施例中更详细讨论的,(i)与正常对照相比在疾病免疫谱中结合更多抗体的肽;和(ii)与正常对照相比在疾病免疫谱中结合更少抗体的肽,是针对感兴趣的疾病的治疗靶标。
感兴趣的差异结合的肽/相应的蛋白质治疗靶标可通过任何合适的方法来鉴定,所述方法包括但不限于:序列分析,与蛋白质组的比较,对阵列上特定位置处的肽的预先了解,以及使用差异结合作为亲和试剂来纯化反应性抗体,而后可在标准免疫学技术中使用该反应性抗体来鉴定靶标。例如,通过将i型糖尿病(tid)患者的免疫特征与年龄匹配的对照进行比较,鉴定出在i型糖尿病患者中结合更多抗体的约200种肽(实施例1)。在将这些肽与人蛋白质组或其部分进行比较时,将多种肽与已知在i型糖尿病中将被上调并释放至细胞外间隙从而引起自身免疫应答的蛋白质进行比对。
由对应于人蛋白质序列的肽组成的阵列可直接与相应蛋白质中的表位进行比对。随机肽阵列和非随机或伪随机肽阵列可用于鉴定肽序列并将肽序列与人蛋白质进行比对。通过将疾病样品与非疾病样品进行比较,可鉴定与疾病样品相比在非疾病样品中结合更多抗体的大量肽。
在将疾病样品与非疾病样品进行比较时,下调的信号可能对于状况具有特异性。例如,实施例1说明了对i型糖尿病中约400种下调的肽的鉴定。在将这400种肽的序列与人蛋白质比较时,约70种肽与已知在i型糖尿病中起作用的蛋白质匹配。这暗示相对于正常血清上升和下降的抗体反应性都是关于疾病中的相关蛋白质的信息的来源。这证明自身抗体可因疾病的发生而下调。
本发明方法的特色特征为可以鉴定早期疾病的独特靶标。由于免疫系统可以非常早地与甚至少量的异常细胞反应,因此可以发现早期治疗靶标。
抗体生物标志物的多重检测
被设计用于检测宿主产生的抗体(而非其他较不丰富的生物标志物)的诊断方法更有可能足够灵敏以检测罕见事件。由于在循环血液中已经存在这些标志物的非常多样化的来源,因此无需形成高亲和力、高特异性抗体的充足供应。在设计用于检测抗体的多重阵列中,将多组(panels)蛋白质或肽附接至固体支持体上,随后暴露于血液。
蛋白质阵列作为能够在单一实验中同时检测大量参数的高容量方法而出现。蛋白质靶标提供了构象表位的来源以供抗体结合,尽管并不总是暴露线性表位。invitrogen生产了一种更全面的蛋白质微阵列,其包含在单一载玻片上排列的约9000种不同的杆状病毒产生的人蛋白质。这些蛋白质阵列的大规模化潜能受到每个载玻片的高成本、可扩大性的缺乏以及重组蛋白质生产、纯化和稳定性的不一致性的限制。使用体外合成的蛋白质已改善了蛋白质生产的通量和成功率,但在排列的量、稳定性、翻译后修饰的不一致性上以及对膜(表面)、多聚体和大蛋白质的偏性上仍然存在问题。这两种方法均局限于检测自身抗体,除非特异性地合成已知的突变体或由病原体衍生的候选蛋白质。对病变细胞的生化分级分离使得能够探查针对修饰的和突变的抗原的抗体,但这是一个实质上更复杂的过程(hanash,s.(2003)diseaseproteomics.nature422,226-232)。
与蛋白质相比,肽能够化学合成,从而可大量获得高度可再现且纯的产品,且该产品具有长贮存期。生物相关修饰或检测分子的附接是简单的,并且非天然的设计也是可能的(reddy,m.m.等人,identificationofcandidateiggbiomarkersforalzheimer'sdiseaseviacombinatoriallibraryscreening.cell144,132-142)。
肽在溶液中以相似的方式呈现,即使与固体支持体结合时也是如此;因此,不论批次间的生产差异如何,均可针对高度一致的结构来筛选抗体相互作用。肽微阵列已可使用的时间远远长于蛋白质微阵列(panicker,r.c.等人,(2004)recentadvancesinpeptide-basedmicroarraytechnologies.combchemhighthroughputscreen7,547-556),并且已经用于多种应用。酶(fu,j.等人,(2010)exploringpeptidespaceforenzymemodulators.jamchemsoc132,6419-6424;以及fu,j.等人,(2011)peptide-modifiedsurfacesforenzymeimmobilization.plosone6,e18692)、蛋白质(diehnelt,c.w.等人,discoveryofhigh-affinityproteinbindingligands-backwards.plosone5,e10728;greving,m.p.等人,high-throughputscreeningintwodimensions:bindingintensityandoff-rateonapeptidemicroarray.analbiochem402,93-95;greving,m.p.等人,thermodynamicadditivityofsequencevariations:analgorithmforcreatinghighaffinitypeptideswithoutlargelibrariesorstructuralinformation.plosone5,e15432;gupta,n.等人,engineeringasyntheticligandfortumornecrosisfactor-alpha.bioconjugchem22,1473-1478)、dna和小分子(boltz,k.w.等人,(2009)peptidemicroarraysforcarbohydraterecognition.analyst134,650-652;foong,y.m.等人,(2012)currentadvancesinpeptideandsmallmoleculemicroarraytechnologies.curropinchembiol16,234-242;moralesbetanzos,c.等人,(2009)bacterialglycoprofilingbyusingrandomsequencepeptidemicroarrays.chembiochem10,877-888)、全细胞(falsey,j.r.等人,(2001)peptideandsmallmoleculemicroarrayforhighthroughputcelladhesionandfunctionalassays.bioconjugchem12,346-353)和抗体(cerecedo,i.等人,(2008)mappingoftheigeandigg4sequentialepitopesofmilkallergenswithapeptidemicroarray-basedimmunoassay.jallergyclinimmunol122,589-594;cretich,m.等人,(2009)epitopemappingofhumanchromograninabypeptidemicroarrays.methodsmolbiol570,221-232;lin,j.等人,(2009)developmentofanovelpeptidemicroarrayforlarge-scaleepitopemappingoffoodallergens.jallergyclinimmunol124,315-322,322e311-313;lorenz,p.等人,(2009)probingtheepitopesignaturesofiggantibodiesinhumanserumfrompatientswithautoimmunedisease.methodsmolbiol524,247-258;perez-gordo,m.等人,(2012)epitopemappingofatlanticsalmonmajorallergenbypeptidemicroarrayimmunoassay.int.archallergyimmunol157,31-40;以及shreffler,w.g.等人,(2005)igeandigg4epitopemappingbymicroarrayimmunoassayrevealsthediversityofimmuneresponsetothepeanutallergen,arah2.jallergyclinimmunol116,893-899)仅是可对其与肽的结合进行分析的生物分子的子集。一个经典的实例是表位作图:可将跨越抗原的肽平铺以有效地破译单克隆抗体的表位。通常,高特异性抗体将识别并结合其表位序列,而与抗原衍生的其他肽没有或具有很少的可测量的结合。利用这种方法,可以区分并表征针对相同抗原产生的不同的单克隆抗体。
交叉反应性的相关性
免疫系统已经进化为产生并扩增忽略自身蛋白质并以显著的强度结合非自身靶标的抗体。免疫系统已经进化为产生并扩增忽略自身蛋白质并以显著的强度结合非自身靶标的抗体。这些不一致的压力在分子水平上变得明显。典型的抗体识别约15个氨基酸的表位,其中约5个氨基酸主导结合能。这5个残基中任何一个的变化将极大地影响结合强度。
其他表位位置的序列变化将改变结合区的空间构象并适度地影响总强度。因此,如果欲将结合强度最大化,则必须将条件调整为允许高亲和力和低亲和力残基相互作用。这意味着严格性降低,因此允许表位序列的变体结合抗体。抗体具有包含许多重叠互补位的50个氨基酸的可变区(抗体的表位识别部分),从而进一步有助于交叉反应性的潜能(mohan,s.等人,(2009)associationenergeticsofcross-reactiveandspecificantibodies.biochemistry48,1390-1398;thorpe,i.f.和brooks,c.l.,第3版(2007)molecularevolutionofaffinityandflexibilityintheimmunesystem.proceedingsofthenationalacademyofsciencesoftheunitedstatesofamerica104,8821-8826;以及zhou,z.h.等人,(2007)propertiesandfunctionofpolyreactiveantibodiesandpolyreactiveantigen-bindingbcells.jautoimmun.29,219-228.epub2007sep2020)。
这些互补位中的每一个均由约15个氨基酸组成,使得互补位和表位为大小相似的序列段,其限定形状和电荷的互补区。互补位可结合多于一个表位,并且单一表位可与多于一个互补位结合,每一对都表现出独特的结合性质。由于单一抗体携带多个互补位,因此抗体能以不同的强度结合不同的而潜在多样化的一组表位。这种交叉反应性以及特异性与亲和性的复杂相互影响是复杂的免疫系统的标志,该免疫系统协调对抗直接威胁的直接攻击以及对抗将来可能的变体暴露的间接攻击。
在体外,对特定线性表位具有特异性的抗体不仅显示出结合序列相关肽,也结合序列非相关肽(folgori,a.等人,(1994)ageneralstrategytoidentifymimotopesofpathologicalantigensusingonlyrandompeptidelibrariesandhumansera.emboj13,2236-2243)。通常表现出构象相关性的这些序列非相关肽被称为模拟表位,并且最初描述于早期噬菌体展示研究中(folgori,a.等人,(1994)ageneralstrategytoidentifymimotopesofpathologicalantigensusingonlyrandompeptidelibrariesandhumansera.emboj13,2236-2243;christian,r.b.等人,(1992)simplifiedmethodsforconstruction,assessmentandrapidscreeningofpeptidelibrariesinbacteriophage.journalofmolecularbiology227,711-718;liu,r.等人,(2003)combinatorialpeptidelibrarymethodsforimmunobiologyresearch.experimentalhematology31,11-30;wang,y.等人,(1995)detectionofmammarytumorvirusenvgene-likesequencesinhumanbreastcancer.cancerresearch55,5173-5179)。
基于噬菌体的系统提供了构建和筛选比使用大多数其他系统所可能达到的具有大得多的配体多样性的文库的机会。例如,针对特定单克隆抗体淘选展示肽的大随机序列文库。反复数轮选择常常导致鉴定出同源表位,但也会鉴定出几种非相关肽序列。随机肽多样性比生物序列多样性大几个数量级的事实意味着该肽将不对应于任何生物肽。所有的结合反应均依赖于非同源交叉反应性(抗体的固有性质)。这意味着可以鉴定针对任何类别抗体的配体:自身抗原、修饰的抗原、突变的表位或非肽模拟表位。尽管在随机序列空间中进行筛选具有这些优势,但噬菌体展示技术受到使用噬菌体和细菌培养物进行反复数轮淘选、二元选择过程和缺乏可扩大性的限制(derda,r.等人,(2011)diversityofphage-displayedlibrariesofpeptidesduringpanningandamplification.molecules16,1776-1803;szardenings,m.(2003)phagedisplayofrandompeptidelibraries:applications,limits,andpotential.jreceptsignaltransductres23,307-34953,54)。迄今为止,随机噬菌体文库尚未产生抗体生物标志物。
免疫特征分析
免疫特征分析是上述技术的综合。首先,并非在噬菌体上以生物学方式展示肽,而是以可寻址有序阵列将合成的较长的肽连接至载玻片上,这是一种更为系统化的方法。虽然噬菌体文库可以有超过1010个单独的克隆,但微阵列已经从每个载玻片几千个斑点增加到数百万个斑点。成本、可靠性、精确度和分析速度使微阵列具有显著的优势。微阵列由于其低成本和可扩大性已证明其对于基因组学和蛋白质组学是无价的,并且商业化的阵列室和扫描仪已经存在多年。
其次,使用抗体作为疾病的生物标志物利用了稳定且易于获得的分子以及免疫系统的多样性、监视和生物扩增的方便的性质。哺乳动物免疫系统的复杂性令人吃惊(janeway,c.和travers,j.(1997)immunobiology:theimmunesysteminhealthanddisease.currentbiologylimited),因此信息量也是如此。随着免疫学家探索免疫组,日渐达成以下共识:能够有>1010个不同的分子种类的抗体组库(nobrega,a.等人,(1998)functionaldiversityandclonalfrequenciesofreactivityintheavailableantibodyrepertoire.europeanjournalofimmunology28,1204-1215)是个体健康状态的过去、现在乃至前驱扰动(prodromicperturbation)的动态数据库。
第三,随机序列肽的使用能够使抗体组库的多样性与待筛选的无偏、综合配体文库相匹配。随机序列肽可用于噬菌体展示文库,但它们携带偏性,并且不是无序的、控制不良的形式。由于随机肽序列对生物空间没有限制且没有刻意的同源性,因此微阵列包含极少的但覆盖范围非常广的序列空间。可在相同的微阵列上筛选与任何疾病或有机体相对应的正常的、突变的、翻译后修饰的和模拟的表位。本领域中最近的出版物已使用了10,000个独特的随机序列20-mer肽来表征大量的疾病状态1、10(brown,j.r.等人,(2011)statisticalmethodsforanalyzingimmunosignatures.bmcbioinformatics12,349;hughes,a.k.等人,(2012)immunosignaturingcandetectproductsfrommolecularmarkersinbraincancer.plosone7,e40201;kroening,k.等人,(2012)autoreactiveantibodiesraisedbyselfderiveddenovopeptidescanidentifyunrelatedantigensonproteinmicroarrays.areautoantibodiesreallyautoantibodies?expmolpathol92,304-311;kukreja,m.等人,(2012)comparativestudyofclassificationalgorithmsforimmunosignaturingdata.bmcbioinformatics13,139;kukreja,m.等人,(2012)immunosignaturingmicroarraysdistinguishantibodyprofilesofrelatedpancreaticdiseases.journalofproteomicsandbioinformatics;以及legutki,j.b.等人,(2010)ageneralmethodforcharacterizationofhumoralimmunityinducedbyavaccineorinfection.vaccine28,4529-4537)。
由免疫特征分析微阵列获得的结果与由噬菌体展示获得的结果相比存在几个显著的差异。免疫特征分析探查阵列上所有的肽,并产生对于每个肽的结合值。噬菌体展示产生在限制性选择后留下的序列,并且通常仅鉴定共有序列。处理免疫特征分析微阵列需要数小时,而非数周。图10显示了这些技术之间的差别。在技术上,‘免疫特征’是指肽的统计学上显著的模式,每一个具有可稳健地将一种疾病状态与其他疾病状态区分开的特定结合值。
技术的这种整合可代表朝向普遍适用的早期诊断平台的目标的进步。有待解决的关键问题为:i)免疫系统是否对传染病和慢性疾病均引起一致的疾病特异性体液应答,ii)抗体是否足够早地响应于疾病的病因学从而是临床上有用的,以及iii)该测定是否足够灵敏、提供足够多的信息、足够廉价以及可扩大,从而筛选大量的患者样品以用于可信度确定。如果这几点能够得到满足,则可以发现任何免疫相关疾病的免疫特征。而后可使用这些限定的反应性模式来早期及全面地诊断疾病。如果可以使这些检测广泛适用于群体,则免疫特征分析可以构成在个体以及流行病学水平上具有重要意义的长期健康监测系统的基础。我们提出在这方面很有前景的平台的几个特点。
免疫特征分析是上述技术的综合。首先,并非在噬菌体上以生物学方式展示肽,而是以可寻址有序阵列将合成的较长的肽连接至载玻片上,这是一种更为系统化的方法。虽然噬菌体文库可以有超过1010个单独的克隆,但微阵列已经从每个载玻片几千个斑点增加到数百万个斑点。成本、可靠性、精确度和分析速度使微阵列具有显著的优势。微阵列由于其低成本和可扩大性已证明其对于基因组学和蛋白质组学是无价的,并且商业化的阵列室和扫描仪已经存在多年。其次,使用抗体作为疾病的生物标志物利用了稳定且易于获得的分子以及免疫系统的多样性、监视和生物扩增的方便的性质。
哺乳动物免疫系统的复杂性令人吃惊,因此信息量也是如此。随着免疫学家探索免疫组,日渐达成以下共识:能够具有>1010个不同的分子种类的抗体组库是个体健康状态的过去、现在乃至前驱扰动的动态数据库。第三,随机序列肽的使用能够使抗体组库的多样性与待筛选的无偏、综合配体文库相匹配。随机序列肽可用于噬菌体展示文库,但它们携带偏性,并且不是无序的、控制不良的形式。由于随机肽序列对生物空间没有限制且没有刻意的同源性,因此微阵列包含极少的但覆盖范围非常广的序列空间。
可在相同的微阵列上筛选与任何疾病或有机体相对应的正常的、突变的、翻译后修饰的和模拟的表位。本领域的出版物已使用了10,000个独特的随机序列20-mer肽来表征大量的疾病状态。由免疫特征分析微阵列获得的结果与由噬菌体展示获得的结果相比存在几个显著的差异。免疫特征分析探查阵列上所有的肽,并产生对于每个肽的结合值。噬菌体展示产生在限制性选择后留下的序列,并且通常仅鉴定共有序列。
处理免疫特征分析微阵列可能需要数小时而非数周。‘免疫特征’是指肽的统计学上显著的模式,每一个具有可稳健地将一种疾病状态与其他疾病状态区分开的特定结合值。因此,本文公开的实施方案的一个方面是用复杂生物样品探查免疫特征阵列的相对较短的处理时间,其中探查和处理时间可能花费最多10分钟、最多20分钟、最多30分钟、最多45分钟、最多60分钟、最多90分钟、最多2小时、最多3小时、最多4小时或最多5小时。或者,探查和处理时间可能花费不超过10分钟、不超过20分钟、不超过30分钟、不超过45分钟、不超过60分钟、不超过90分钟、不超过2小时、不超过3小时、不超过4小时或不超过5小时。
技术的这种整合可代表朝向普遍适用的早期诊断平台的目标的进步。有待解决的关键问题为:i)免疫系统是否对传染病和慢性疾病均引起一致的疾病特异性体液应答,ii)抗体是否足够早地响应于疾病病因学从而是临床上有用的,以及iii)该测定是否足够灵敏、提供足够多的信息、足够廉价以及可扩大,从而筛选大量的患者样品以用于可信度确定。如果这几点能够得到满足,则可以发现任何免疫相关疾病的免疫特征。
而后可以使用这些限定的反应性模式来早期及全面地诊断疾病。如果可以使这些检测广泛适用于群体,则免疫特征分析可以构成在个体以及流行病学水平上具有重要意义的长期健康监测系统的基础。我们提出在这方面很有前景的平台的几个特点。
免疫特征分析的独特特点
除了抗体的互补位对配体的亲和力外,结合强度还能受血清中的抗体种类的浓度影响。与噬菌体展示不同,免疫特征分析可以定量地测量这些参数的结果,并且可在非常大的动态范围内进行这种测量(legutki,j.b.等人,(2010)ageneralmethodforcharacterizationofhumoralimmunityinducedbyavaccineorinfection.vaccine28,4529-4537;stafford,p.和johnston,s.(2011)microarraytechnologydisplaysthecomplexitiesofthehumoralimmuneresponse.expertrevmoldiagn11,5-8;halperin,r.f.等人,(2011)exploringantibodyrecognitionofsequencespacethroughrandom-sequencepeptidemicroarrays.molecular&cellularproteomics10)。
科学家利用这种能力来检查高亲和力单克隆抗体与免疫特征分析微阵列的结合。他们发现单一单克隆抗体识别数百个随机序列,并且可以测量和比较这些独特结合反应的不同强度(halperin,r.f.等人,(2011)exploringantibodyrecognitionofsequencespacethroughrandom-sequencepeptidemicroarrays.molecular&cellularproteomics10)。令人好奇的是,这些脱靶模拟表位相互作用中有许多比同源表位具有更强的结合。虽然这些相互作用的相应溶液相结合较低,但构建免疫特征分析微阵列的方式增强了这些相互作用。抗体与免疫特征分析微阵列脱靶结合的这种免疫学现象对该技术极为重要。
另一重要的观察结果是对于低亲和力相互作用的检测而言,免疫特征分析比噬菌体展示或基于elisa的测定具有更高的灵敏度(stafford,p.和johnston,s.(2011)microarraytechnologydisplaysthecomplexitiesofthehumoralimmuneresponse.expertrevmoldiagn11)。高灵敏度是载玻片表面上肽的高密度的结果,并且曾被称为“免疫特征分析效应”。这已通过将肽打印在官能化玻璃表面上并检测肽在其上的不同空间布置而建立。如果打印阵列使得肽间隔约9至约12nm,则同源表位比脱靶随机肽(除非常强的模拟表位以外)更有利地竞争抗体。
我们通常使肽平均间隔1-2nm,但在肽间隔3-4nm时观察到脱靶结合。如果肽间隔约1至约1.5nm,则观察到脱靶结合的增加。紧密填充的肽似乎通过亲合力和快速重结合来捕获抗体。这一概念已经显示出极大的可再现性,并示于图11中(stafford,p.等人,(2012)physicalcharacterizationofthe"immunosignaturingeffect".molcellproteomics11,m111011593;chase,b.a.等人,(2012)evaluationofbiologicalsamplepreparationforimmunosignature-baseddiagnostics.clinvaccineimmunol19,352-358;hughes,a.k.等人,(2012)immunosignaturingcandetectproductsfrommolecularmarkersinbraincancer.plosone7,e40201;restrepo,l.等人,(2011)applicationofimmunosignaturestotheassessmentofalzheimer'sdisease.annalsofneurology70,286-295)。虽然肽的序列是完全随机的,但它们对抗体的脱靶捕获明显不是这样;相反,血清与阵列结合的模式非常一致。早期对这种技术的担心是任何血清样品中抗体种类的高度多样性可能导致重叠的结合竞争,从而导致扁平的(flat)、无法提供信息的强度场。数据不支持这一观点。事实上,即使是稀释至血清中的纯化的单克隆抗体仍会保持其独特的反应性模式,没有或具有极少的结合损失(uhlen,m.和hober,s.(2009)generationandvalidationofaffinityreagentsonaproteome-widelevel.jmolrecognit22,57-64)。
用于解释常规核酸微阵列的经典统计模型(draghici,s.(2012)statisticsanddataanalysisformicroarraysusingrandbioconductor.chapman&hall/crc)对于解决由免疫特征阵列所呈现的新复杂性不具有灵活性。并非是描述rna或dna与微阵列上的互补探针结合的一对一结合模型,免疫特征分析肽可与多于一种抗体结合,并且许多不同的抗体可与相同的肽结合。三个不同的报告特别针对免疫特征分析比较了图像分析方法(yang,y.等人,(2011)segmentationandintensityestimationformicroarrayimageswithsaturatedpixels.bmcbioinformatics12,462)、因素分析和混合模型(brown,j.r.等人,(2011)statisticalmethodsforanalyzingimmunosignatures.bmcbioinformatics12,349)以及分类(kukreja,m.等人,(2012)comparativestudyofclassificationalgorithmsforimmunosignaturingdata.bmcbioinformatics13,139)。免疫特征分析微阵列存在许多能够区分疾病的基本性质。
首先,来自健康志愿者的对照血清表现出相当宽的基线结合反应性分布。这要求使用该技术的大规模研究必须从大量未患病个体中采集血清以适应群体变异性。其次,来自患有给定疾病的人的血清的特征(signatures)非常一致,不同于非疾病血清的特征。这一观察结果暗示免疫系统不断探查局部环境并与局部环境进行反应,从而导致特征的巨大差异。然而,一旦针对于抗原,抗体趋于形成具有很小的个体变异性的窄且明确定义的特征。
即使如此,该技术能够识别疾病的亚型(hughes,a.k.等人,(2012)immunosignaturingcandetectproductsfrommolecularmarkersinbraincancer.plosone7,e40201),同时仍提供对照与患病者之间的区别。对多种肽之间的共同关系和协方差的分析提供了极大的辨别能力,该能力在单一表位水平上是不可能的。在免疫学上,抗体:肽结合模式并非由非特异性危险信号或天然抗体的活性而创建:它们由可识别的刺激而创建。抗体吸附实验证明,来自流感感染的肽主要结合病毒特异性抗体,而阿尔茨海默病的特征结合许多抗aβ抗体(legutki,j.b.等人,(2010)ageneralmethodforcharacterizationofhumoralimmunityinducedbyavaccineorinfection.vaccine28,4529-4537;restrepo,l.等人,(2011)applicationofimmunosignaturestotheassessmentofalzheimer'sdisease.annalsofneurology70,286-295)。
通过免疫特征分析进行的疾病确定已经与使用目前的诊断测定获得的结果良好地关联(hughes,a.k.等人,(2012)immunosignaturingcandetectproductsfrommolecularmarkersinbraincancer.plosone7,e40201;kukreja,m.等人,(2012)immunosignaturingmicroarraysdistinguishantibodyprofilesofrelatedpancreaticdiseases.journalofproteomicsandbioinformatics;legutki,j.b.等人,(2010)ageneralmethodforcharacterizationofhumoralimmunityinducedbyavaccineorinfection.vaccine28,4529-4537)。免疫特征携带采用传统诊断法无法获得的历史健康信息;即,可以检测速发型应答和记忆应答(legutki,j.b.等人,(2010)ageneralmethodforcharacterizationofhumoralimmunityinducedbyavaccineorinfection.vaccine28,4529-4537)。迄今为止,该方法已经应用于超过33种不同的疾病和后遗症,包括小鼠、大鼠、狗、猪和人宿主中的病毒、细菌、真菌和寄生虫感染,癌症,糖尿病,自身免疫疾病,移植患者和许多慢性疾病。可建立与病理学相关的肽结合模式的高度可再现模式。
这些结合谱将从患者和健康志愿者获得的盲血清样品正确地分类,并且在灵敏度和准确度上优于经典的免疫学检验。在大规模研究中,免疫特征能够以非常高的准确度诊断溪谷热(valleyfever)患者,包括正确诊断最初标准elisa检测为阴性的患者。对患者的免疫特征的分析能够在癌症之间和癌症内进行区分(brown,j.r.等人,(2011)statisticalmethodsforanalyzingimmunosignatures.bmcbioinformatics12,349;hughes,a.k.等人,(2012)immunosignaturingcandetectproductsfrommolecularmarkersinbraincancer.plosone7,e40201;kukreja,m.等人,(2012)immunosignaturingmicroarraysdistinguishantibodyprofilesofrelatedpancreaticdiseases.journalofproteomicsandbioinformatics;以及yang,y.等人,(2011)segmentationandintensityestimationformicroarrayimageswithsaturatedpixels.bmcbioinformatics12,462),甚至达到准确地诊断将会响应于以及将不会响应于药物治疗的癌症类型的程度(hughes,a.k.等人,immunosignaturingcandetectproductsfrommolecularmarkersinbraincancer.plosone7,e40201)。
免疫特征分析技术的最独特的特点之一可能被证明是对特定肽:抗体反应性(先前不可测量的一类相互作用)的降低的测量。即,尽管来自患病个体的血清产生比正常血清更高的信号,但也存在始终表现出相对于健康人降低的结合的肽。(kukreja,m.等人,(2012)immunosignaturingmicroarraysdistinguishantibodyprofilesofrelatedpancreaticdiseases.journalofproteomicsandbioinformatics;以及legutki,j.b.等人,(2010),ageneralmethodforcharacterizationofhumoralimmunityinducedbyavaccineorinfection.vaccine28,4529-4537)。这些“下降”肽在免疫应答中的作用是令人好奇的。尽管在其最简单的层面上,这些“下降”肽提高了疾病分类,但可能存在一些以其他方式无法观察到的潜在免疫学现象。
分子与阵列的结合
根据美国国家癌症研究所(nationalcancerinstitute),存在约150类癌症以及(取决于人们如何定义它们)数百种不同的亚型。抗体通常针对于肿瘤细胞表达的抗原而产生,并且随后在b细胞成熟期间扩增。抗体也在对疫苗或感染应答期间产生。在受试者日常暴露于各种病原性以及非病原性刺激期间,也可产生抗体。
受试者体内抗体扩增的过程可产生与状况相关的受试者特异性标志物的充足供应。抗体扩增可提供与受试者的特定健康状态和/或状况相关的充足数目的抗体。样品中充足数目的抗体的存在可降低健康监测方法中对人工生物标志物扩增的需求。样品中充足数目的抗体的存在可允许将少量样品成功应用于例如健康监测方法中。
本发明的方法和阵列允许,例如,采用少量来自受试者的生物样品鉴定治疗靶标、疫苗,或筛选针对癌症和/或免疫病症的治疗靶标的方法。在一些实施方案中,生物样品可以无需进一步处理而以少量应用于本发明的方法中。在一些实施方案中,生物样品包括血液、血清、唾液、汗液、细胞、组织或任何体液。在一些实施方案中,由本发明的阵列和方法进行的分析需要约0.5nl、约1nl、约2nl、约3nl、约4nl、约5nl、约6nl、约7nl、约8nl、约9nl、约10nl、约11nl、约12nl、约13nl、约14nl、约15nl、约16nl、约17nl、约18nl、约19nl、约20nl、约21nl、约22nl、约23nl、约24nl、约25nl、约26nl、约27nl、约28nl、约29nl、约30nl、约31nl、约32nl、约33nl、约34nl、约35nl、约36nl、约37nl、约38nl、约39nl、约40nl、约41nl、约42nl、约43nl、约44nl、约45nl、约46nl、约47nl、约48nl、约49nl或约50nl、约51nl、约52nl、约53nl、约54nl、约55nl、约56nl、约57nl、约58nl、约59nl、约60nl、约61nl、约62nl、约63nl、约64nl、约65nl、约66nl、约67nl、约68nl、约69nl、约70nl、约71nl、约72nl、约73nl、约74nl、约75nl、约76nl、约77nl、约78nl、约79nl、约80nl、约81nl、约82nl、约83nl、约84nl、约85nl、约86nl、约87nl、约88nl、约89nl、约90nl、约91nl、约92nl、约93nl、约94nl、约95nl、约96nl、约97nl、约98nl、约99nl、约0.1、约0.2μl、约0.3μl、约0.4μl、约0.5μl、约0.6μl、约0.7μl、约0.8μl、约0.9μl、约1μl、约2μl、约3μl、约4μl、约5μl、约6μl、约7μl、约8μl、约9μl、约10μl、约11μl、约12μl、约13μl、约14μl、约15μl、约16μl、约17μl、约18μl、约19μl、约20μl、约21μl、约22μl、约23μl、约24μl、约25μl、约26μl、约27μl、约28μl、约29μl、约30μl、约31μl、约32μl、约33μl、约34μl、约35μl、约36μl、约37μl、约38μl、约39μl、约40μl、约41μl、约42μl、约43μl、约44μl、约45μl、约46μl、约47μl、约48μl、约49μl或约50μl的生物样品。
来自受试者的生物样品可以例如从受试者采集并直接与本发明的阵列接触。在一些实施方案中,生物样品在与本发明的阵列接触之前,不需要制备或处理步骤。在一些实施方案中,来自受试者的干血样在与本发明的阵列接触之前,在稀释步骤中重建。稀释可为免疫特征分析提供来自受试者的生物样品的抗体的最佳浓度。
本发明的方法和阵列允许采用少量来自受试者的生物样品鉴定治疗靶标、疫苗,或筛选针对癌症和/或免疫病症的治疗靶标。在一些实施方案中,本发明的方法需要不多于约0.5nl至约50nl、不多于约1nl至约100nl、不多于约1nl至约150nl、不多于约1nl至约200nl、不多于约1nl至约250nl、不多于约1nl至约300nl、不多于约1nl至约350nl、不多于约1nl至约400nl、不多于约1至约450nl、不多于约5nl至约500nl、不多于约5nl至约550nl、不多于约5nl至约600nl、不多于约5nl至约650nl、不多于约5nl至约700nl、不多于约5nl至约750nl、不多于约5nl至约800nl、不多于约5nl至约850nl、不多于约5nl至约900nl、不多于约5nl至约950nl、不多于约5nl至约1μl、不多于约0.5μl至约1μl、不多于约0.5μl至约5μl、不多于约1μl至约10μl、不多于约1μl至约20μl、不多于约1μl至约30μl、不多于约1μl至约40μl或不多于约1μl至约50μl。
在一些实施方案中,本发明的方法需要至少0.5nl至约50nl、至少约1nl至约100nl、至少约1nl至约150nl、至少约1nl至约200nl、至少约1nl至约250nl、至少约1nl至约300nl、至少约1nl至约350nl、至少约1nl至约400nl、至少约1至约450nl、至少约5nl至约500nl、至少约5nl至约550nl、至少约5nl至约600nl、至少约5nl至约650nl、至少约5nl至约700nl、至少约5nl至约750nl、至少约5nl至约800nl、至少约5nl至约850nl、至少约5nl至约900nl、至少约5nl至约950nl、至少约5nl至约1μl、至少约0.5μl至约1μl、至少约0.5μl至约5μl、至少约1μl至约10μl、至少约1μl至约20μl、至少约1μl至约30μl、至少约1μl至约40μl、至少约1μl至约50μl或至少50μl。
受试者可以提供多种生物样品,包括固体生物样品,例如,来自活检或组织。在一些实施方案中,本发明的阵列和方法需要约1mg、约5mg、约10mg、约15mg、约20mg、约25mg、约30mg、约35mg、约40mg、约45mg、约50mg、约55mg、约60mg、约65mg、约7mg、约75mg、约80mg、约85mg、约90mg、约95mg或约100mg的生物样品。
在一些实施方案中,本发明的方法和阵列需要不多于约1mg至约5mg、不多于约1mg至约10mg、不多于约1mg至约20mg、不多于约1mg至约30mg、不多于约1mg至约40mg、不多于约1mg至约50mg、不多于约50mg至约60mg、不多于约50mg至约70mg、不多于约50mg至约80mg、不多于约50mg至约90mg、不多于约50mg至约100mg的生物样品。
在一些实施方案中,本发明的方法和阵列需要至少约1mg至约5mg、至少约1mg至约10mg、至少约1mg至约20mg、至少约1mg至约30mg、至少约1mg至约40mg、至少约1mg至约50mg、至少约50mg至约60mg、至少约50mg至约70mg、至少约50mg至约80mg、至少约50mg至约90mg、至少约50mg至约100mg的生物样品。
本发明的方法和阵列提供了用于鉴定治疗靶标和/或疫苗的灵敏方法。本发明的方法和阵列提供了采用少量来自受试者的生物样品筛选针对癌症和/或免疫病症状况的治疗靶标的灵敏方法。在一些实施方案中,来自受试者的生物样品的浓度过高,因此在与本发明的阵列接触之前需要稀释。在生物样品与本发明的阵列接触之前,可对该样品进行多次稀释。稀释可以是系列稀释,其可以以对数方式产生几何级数的浓度。例如,10倍系列稀释可以为1m、0.01m、0.001m及其几何级数。稀释可以为,例如,一倍稀释、两倍稀释、三倍稀释、四倍稀释、五倍稀释、六倍稀释、七倍稀释、八倍稀释、九倍稀释、十倍稀释、十六倍稀释、二十五倍稀释、三十二倍稀释、六十四倍稀释和/或一百二十五倍稀释。
生物样品可来源于受试者体内的多种来源,并且生物样品可在多种不同情况下从受试者采集。例如,可以在日常医疗咨询期间采集生物样品,如在年度体检期间抽血。生物样品可以在非日常咨询过程中采集,例如,生物样品可在活检过程中采集。受试者还可从自身采集生物样品,并且受试者可以以直接面向消费者的方式提供将要通过本发明的方法和系统来分析的生物样品。在一些实施方案中,可将生物样品邮寄至本发明的方法和阵列的提供者。在一些实施方案中,将干生物样品,如在滤纸上的来自受试者的干血样邮寄至本发明的方法和阵列的提供者。
分子与本发明阵列的结合创造出可与状况相关的结合模式。分子与阵列中的肽的结合亲和力可在数学上与状况相关。抗体与本发明的多个不同肽的脱靶结合模式可在数学上与状况相关。分子与本发明的多个不同肽的结合亲合力可在数学上与状况相关。脱靶结合和亲合力可包含生物样品中的分子与肽阵列中的多个不同肽的相互作用。分子与肽阵列中的多个不同肽的结合亲合力可决定该分子与肽阵列的缔合常数。在一些实施方案中,样品中的抗体的浓度有助于与肽阵列的结合亲合力,例如,通过捕获阵列中临界数目的抗体,并允许抗体与阵列的快速重结合。
生物分子与阵列的结合亲合力可由多种键相互作用的组合来决定。抗体与肽阵列中的多个肽的交叉反应性可有助于结合亲合力。在一些实施方案中,抗体可识别约3个氨基酸、约4个氨基酸、约5个氨基酸、约6个氨基酸、约7个氨基酸、约8个氨基酸、约9个氨基酸、约10个氨基酸、约11个氨基酸、约12个氨基酸、约13个氨基酸、约14个氨基酸、约15个氨基酸、约16个氨基酸或约17个氨基酸的表位。在一些实施方案中,约5个氨基酸的序列主导抗体与肽的结合能。
分子与本发明阵列的脱靶结合和/或亲合力可以,例如,有效地将跨越飞摩尔(fm)至微摩尔(μm)解离常数的结合亲和力压缩至仅使用3log的动态范围即可定量测量的范围。分子可以以103m-1或更高的缔合常数与阵列中的多个肽结合。分子可以以范围为103至106m-1、2×103m-1至106m-1的缔合常数和/或范围为104m-1至106m-1的缔合常数与阵列中的多种肽结合。分子可以以约1fm、约2fm、约3fm、约4fm、约5fm、约6fm、约7fm、约8fm、约9fm、约10fm、约20fm、约30fm、约40fm、约50fm、约60fm、约70fm、约80fm、约90fm、约100fm、约200fm、约300fm、约400fm、约500fm、约600fm、约700fm、约800fm、约900fm、约1皮摩尔(pm)、约2pm、约3pm、约4pm、约5pm、约6pm、约7pm、约8pm、约9pm、约10pm、约20pm、约30pm、约40pm、约50pm、约60pm、约7pm、约80pm、约90pm、约100pm、约200pm、约300pm、约400pm、约500pm、约600pm、约700pm、约800pm、约900pm、约1纳摩尔(nm)、约2nm、约3nm、约4nm、约5nm、约6nm、约7nm、约8nm、约9nm、约10nm、约20nm、约30nm、约40nm、约50nm、约60nm、约70nm、约80nm、约90nm、约100nm、约200nm、约300nm、约400nm、约500nm、约600nm、约700nm、约800nm、约900nm、约1μm、约2μm、约3μm、约4μm、约5μm、约6μm、约7μm、约8μm、约9μm、约10μm、约20μm、约30μm、约40μm、约50μm、约60μm、约70μm、约80μm、约90μm或约100μm的解离常数与阵列中的多种肽结合。
分子可以以至少1fm、至少2fm、至少3fm、至少4fm、至少5fm、至少6fm、至少7fm、至少8fm、至少9fm、至少10fm、至少20fm、至少30fm、至少40fm、至少50fm、至少60fm、至少70fm、至少80fm、至少90fm、至少100fm、至少200fm、至少300fm、至少400fm、至少500fm、至少600fm、至少700fm、至少800fm、至少900fm、至少1皮摩尔(pm)、至少2pm、至少3pm、至少4pm、至少5pm、至少6pm、至少7pm、至少8pm、至少9pm、至少10pm、至少20pm、至少30pm、至少40pm、至少50pm、至少60pm、至少7pm、至少80pm、至少90pm、至少100pm、至少200pm、至少300pm、至少400pm、至少500pm、至少600pm、至少700pm、至少800pm、至少900pm、至少1纳摩尔(nm)、至少2nm、至少3nm、至少4nm、至少5nm、至少6nm、至少7nm、至少8nm、至少9nm、至少10nm、至少20nm、至少30nm、至少40nm、至少50nm、至少60nm、至少70nm、至少80nm、至少90nm、至少100nm、至少200nm、至少300nm、至少400nm、至少500nm、至少600nm、至少700nm、至少800nm、至少900nm、至少1μm、至少2μm、至少3μm、至少4μm、至少5μm、至少6μm、至少7μm、至少8μm、至少9μm、至少10μm、至少20μm、至少30μm、至少40μm、至少50μm、至少60μm、至少70μm、至少80μm、至少90μm或约100μm的解离常数与阵列中的多个肽结合。
来自生物样品的抗体与肽微阵列结合的动态范围可被描述为检测到的结合信号的最大值与最小值之间的比值。结合信号可以是,例如,利用第二抗体检测到的荧光信号。传统测定受限于结合的预定的且狭窄的动态范围。本发明的方法和阵列可检测宽动态范围的抗体与本发明阵列中的肽的结合。在一些实施方案中,可在对数尺度上检测宽动态范围的抗体结合。在一些实施方案中,本发明的方法和阵列允许使用最高2log的动态范围、最高3log的动态范围、最高4log的动态范围或最高5log的动态范围来检测多种抗体与阵列的结合模式。
阵列中的分子组成可决定分子与阵列的结合亲合力。在健康状况的预防、治疗、诊断或监测中使用的阵列中可存在多种不同的分子。生物分子的非限制性实例包括氨基酸、肽、肽模拟物、蛋白质、重组蛋白质抗体(单克隆或多克隆的)、抗体片段、抗原、表位、碳水化合物、脂质、脂肪酸、酶、天然产物、核酸(包括dna、rna、核苷、核苷酸、其结构类似物或组合)、营养物、受体和维生素。在一些实施方案中,阵列中的分子为模拟表位,即模拟表位的结构并能够结合由表位引发的抗体的分子。在一些实施方案中,阵列中的分子为互补位或互补位模拟物,包含与抗原表位结合的抗体(或t细胞受体)的可变区中的位点。在一些实施方案中,本发明的阵列是包含随机肽序列的肽阵列。
肽阵列中的氨基酸内距离为肽微阵列中的每个肽之间的距离。氨基酸内距离可有助于脱靶结合和/或分子与阵列的结合亲合力。氨基酸内差异可以是约0.5nm、约1nm、约1nm、1.1nm、约1.2nm、约1.3nm、约1.4nm、约1.5nm、约1.6nm、约1.7nm、约1.8nm、约1.9nm、约2nm、约2.1nm、约2.2nm、约2.3nm、约2.4nm、约2.5nm、约2.6nm、约2.7nm、约2.8nm、约2.9nm、约3nm、约3.1nm、约3.2nm、约3.3nm、约3.4nm、约3.5nm、约3.6nm、约3.7nm、约3.8nm、约3.9nm、约4nm、约4.1nm、约4.2nm、约4.3nm、约4.4nm、约4.5nm、约4.6nm、约4.7nm、约4.8nm、约4.9nm、约5nm、约5.1nm、约5.2nm、约5.3nm、约5.4nm、约5.5nm、约5.6nm、约5.7nm、约5.8nm、约5.9nm和/或约6nm。在一些实施方案中,氨基酸内距离小于6纳米(nm)。
氨基酸内差异可以是至少0.5nm、至少1nm、至少1nm、至少1.1nm、至少1.2nm、至少1.3nm、至少1.4nm、至少1.5nm、至少1.6nm、至少1.7nm、至少1.8nm、至少1.9nm、至少2nm、至少2.1nm、至少2.2nm、至少2.3nm、至少2.4nm、至少2.5nm、至少2.6nm、至少2.7nm、至少2.8nm、至少2.9nm、至少3nm、至少3.1nm、至少3.2nm、至少3.3nm、至少3.4nm、至少3.5nm、至少3.6nm、至少3.7nm、至少3.8nm、至少3.9nm、至少4nm、至少4.1nm、至少4.2nm、至少4.3nm、至少4.4nm、至少4.5nm、至少4.6nm、至少4.7nm、至少4.8nm、至少4.9nm、至少5nm、至少5.1nm、至少5.2nm、至少5.3nm、至少5.4nm、至少5.5nm、至少5.6nm、至少5.7nm、至少5.8nm或至少5.9nm。
氨基酸内差异可以是不大于0.5nm、不大于1nm、不大于1nm、不大于1.1nm、不大于1.2nm、不大于1.3nm、不大于1.4nm、不大于1.5nm、不大于1.6nm、不大于1.7nm、不大于1.8nm、不大于1.9nm、不大于2nm、不大于2.1nm、不大于2.2nm、不大于2.3nm、不大于2.4nm、不超过2.5nm、不大于2.6nm、不大于2.7nm、不大于2.8nm、不大于2.9nm、不大于3nm、不大于3.1nm、不大于3.2nm、不大于3.3nm、不大于3.4nm、不大于3.5nm、不大于3.6nm、不大于3.7nm、不大于3.8nm、不大于3.9nm、不大于4nm、不大于4.1nm、不大于4.2nm、不大于4.3nm、不大于4.4nm、不大于4.5nm、不大于4.6nm、不大于4.7nm、不大于4.8nm、不大于4.9nm、不大于5nm、不大于5.1nm、不大于5.2nm、不大于5.3nm、不大于5.4nm、不大于5.5nm、不大于5.6nm、不大于5.7nm、不大于5.8nm、不大于5.9nm和/或不大于6nm。在一些实施方案中,氨基酸内距离不大于6纳米(nm)。
氨基酸内差异的范围可以是0.5nm至1nm、0.5nm至2nm、0.5nm至3nm、0.5nm至3nm、0.5nm至4nm、0.5nm至5nm、0.5nm至6nm、1nm至2nm、1nm至3nm、1nm至4nm、1nm至5nm、1nm至6nm、2nm至3nm、2nm至4nm、2nm至5nm、2nm至6nm、3nm至4nm、3nm至5nm、3nm至6nm、4nm至5nm、4nm至6nm和/或5nm至6nm。
肽阵列可以在表面上包含多种不同的肽模式。肽阵列可包含,例如,不同的多个肽和/或分子的单份、一式两份、一式三份、一式四份、一式五份、一式六份、一式七份、一式八份、一式九份和/或一式十份重复。在一些实施方案中,将多个不同的肽重复地点印在肽阵列表面上。肽阵列可以,例如,包含均匀地分布在阵列上的多个肽。肽阵列可以,例如,包含非均匀地分布在阵列上的多个肽。
可将肽“点印”于肽阵列中。肽斑点(peptidespot)可具有各种几何形状,例如,肽斑点可以是圆形、正方形、矩形和/或三角形。肽斑点可具有多种直径。肽斑点直径的非限制性实例为约3μm至约8μm、约3至约10mm、约5至约10mm、约10μm至约20μm、约30μm、约40μm、约50μm、约60μm、约70μm、约80μm、约90μm、约100μm、约110μm、约120μm、约130μm、约140μm、约150μm、约160μm、约170μm、约180μm、约190μm、约200μm、约210μm、约220μm、约230μm、约240μm和/或约250μm。
肽阵列可包含许多不同的肽。在一些实施方案中,肽阵列包含约10个肽;约50个肽;约100个肽;约200个肽;约300个肽;约400个肽;约500个肽;约750个肽;约1000个肽;约1250个肽;约1500个肽;约1750个肽;约2,000个肽;约2,250个肽;约2,500个肽;约2,750个肽;约3,000个肽;约3,250个肽;约3,500个肽;约3,750个肽;约4,000个肽;约4,250个肽;约4,500个肽;约4,750个肽;约5,000个肽;约5,250个肽;约5,500个肽;约5,750个肽;约6,000个肽;约6,250个肽;约6,500个肽;约7,500个肽;约7,725个肽;8,000个肽;约8,250个肽;约8,500个肽;约8,750个肽;约9,000个肽;约9,250个肽;约10,000个肽;约10,250个肽;约10,500个肽;约10,750个肽;约11,000个肽;约11,250个肽;约11,500个肽;约11,750个肽;约12,000个肽;约12,250个肽;约12,500个肽;约12,750个肽;约13,000个肽;约13,250个肽;约13,500个肽;约13,750个肽;约14,000个肽;约14,250个肽;约14,500个肽;约14,750个肽;约15,000个肽;约15,250个肽;约15,500个肽;约15,750个肽;约16,000个肽;约16,250个肽;约16,500个肽;约16,750个肽;约17,000个肽;约17,250个肽;约17,500个肽;约17,750个肽;约18,000个肽;约18,250个肽;约18,500个肽;约18,750个肽;约19,000个肽;约19,250个肽;约19,500个肽;约19,750个肽;约20,000个肽;约20,250个肽;约20,500个肽;约20,750个肽;约21,000个肽;约21,250个肽;约21,500个肽;约21,750个肽;约22,000个肽;约22,250个肽;约22,500个肽;约22,750个肽;约23,000个肽;约23,250个肽;约23,500个肽;约23,750个肽;约24,000个肽;约24,250个肽;约24,500个肽;约24,750个肽;约25,000个肽;约25,250个肽;约25,500个肽;约25,750个肽;和/或约30,000个肽。
在一些实施方案中,在健康监测方法、治疗方法、诊断方法和预防状况的方法中使用的肽阵列包含多于30,000个肽。在一些实施方案中,在健康监测方法中使用的肽阵列包含约330,000个肽。在一些实施方案中,该阵列包含约30,000个肽;约35,000个肽;约40,000个肽;约45,000个肽;约50,000个肽;约55,000个肽;约60,000个肽;约65,000个肽;约70,000个肽;约75,000个肽;约80,000个肽;约85,000个肽;约90,000个肽;约95,000个肽;约100,000个肽;约105,000个肽;约110,000个肽;约115,000个肽;约120,000个肽;约125,000个肽;约130,000个肽;约135,000个肽;约140,000个肽;约145,000个肽;约150,000个肽;约155,000个肽;约160,000个肽;约165,000个肽;约170,000个肽;约175,000个肽;约180,000个肽;约185,000个肽;约190,000个肽;约195,000个肽;约200,000个肽;约210,000个肽;约215,000个肽;约220,000个肽;约225,000个肽;约230,000个肽;约240,000个肽;约245,000个肽;约250,000个肽;约255,000个肽;约260,000个肽;约265,000个肽;约270,000个肽;约275,000个肽;约280,000个肽;约285,000个肽;约290,000个肽;约295,000个肽;约300,000个肽;约305,000个肽;约310,000个肽;约315,000个肽;约320,000个肽;约325,000个肽;约330,000个肽;约335,000个肽;约340,000个肽;约345,000个肽;和/或约350,000个肽。在一些实施方案中,在健康监测方法中使用的肽阵列包含多于330,000个肽。
肽阵列可包含许多不同的肽。在一些实施方案中,肽阵列包含至少2,000个肽;至少3,000个肽;至少4,000个肽;至少5,000个肽;至少6,000个肽;至少7,000个肽;至少8,000个肽;至少9,000个肽;至少10,000个肽;至少11,000个肽;至少12,000个肽;至少13,000个肽;至少14,000个肽;至少15,000个肽;至少16,000个肽;至少17,000个肽;至少18,000个肽;至少19,000个肽;至少20,000个肽;至少21,000个肽;至少22,000个肽;至少23,000个肽;至少24,000个肽;至少25,000个肽;至少30,000个肽;至少40,000个肽;至少50,000个肽;至少60,000个肽;至少70,000个肽;至少80,000个肽;至少90,000个肽;至少100,000个肽;至少110,000个肽;至少120,000个肽;至少130,000个肽;至少140,000个肽;至少150,000个肽;至少160,000个肽;至少约170,000个肽;至少180,000个肽;至少190,000个肽;至少200,000个肽;至少210,000个肽;至少220,000个肽;至少230,000个肽;至少240,000个肽;至少250,000个肽;至少260,000个肽;至少270,000个肽;至少280,000个肽;至少290,000个肽;至少300,000个肽;至少310,000个肽;至少320,000个肽;至少330,000个肽;至少340,000个肽;至少350,000个肽。在一些实施方案中,在健康监测方法中使用的肽阵列包含至少330,000个肽。
肽可通过连接体分子物理链接至肽阵列上。肽的n末端或c末端可附接至连接体分子。连接体分子可以为,例如,存在于阵列表面上的多种官能分子,如酰亚胺官能团、胺官能团、羟基官能团、羧基官能团、醛官能团和/或巯基官能团。连接体分子可以为,例如,聚合物。在一些实施方案中,连接体为马来酰亚胺。在一些实施方案中,连接体为甘氨酸-丝氨酸-半胱氨酸(gsc)或甘氨酸-甘氨酸-半胱氨酸(ggc)连接体。在一些实施方案中,连接体由不同长度或组成的多肽组成。在一些情况下,连接体为不同长度的聚乙二醇。在另外一些情况下,连接体为羟甲基苯甲酸、4-羟基-2-甲氧基苯甲醛、4-氨磺酰苯甲酸或适用于将肽附接至固体基底的其他连接体。
肽阵列的表面可包含多种不同的材料。肽阵列的表面可以是,例如,玻璃。可构成肽阵列表面的材料的非限制性实例包括玻璃、官能化玻璃、硅、锗、砷化镓、磷化镓、二氧化硅、氧化钠、氮化硅(siliconnitrade)、硝化纤维、尼龙、聚四氟乙烯、聚偏二氟乙烯(polyvinylidendiflouride)、聚苯乙烯、聚碳酸酯、甲基丙烯酸酯或其组合。
肽阵列的表面可以是平的、凹的或凸的。肽阵列的表面可以是均质的,并且阵列的表面可以是异质的。在一些实施方案中,肽阵列的表面是平的。
肽阵列的表面可涂覆有涂层。涂层可以例如改善本发明的阵列的粘附能力。涂层可以例如降低生物样品与本发明的阵列的背景粘附。在一些实施方案中,本发明的肽阵列包含具有氨基硅烷涂层的载玻片。
肽阵列可具有多种尺寸。肽阵列可以是肽微阵列。
检测
可以以多种形式检测样品成分与阵列之间的结合相互作用。在一些形式中,对样品的成分进行标记。标记物可以是放射性同位素或染料等。可通过在获得样品前对患者施用标记物或通过将标记物连接至样品或其选择性成分上来提供标记物。
结合相互作用还可使用第二检测试剂如抗体来检测。例如,样品中的抗体与阵列的结合可使用对抗体的同种型(例如,igg(包括任何亚型,如igg1、igg2、igg3和igg4)、iga、igm)具有特异性的第二抗体来检测。第二抗体通常被标记,并且可与被分析样品中特定同种型的所有抗体结合。可使用具有不同的同种型特异性的不同的第二抗体。虽然在被相同样品中不同同种型的抗体所结合的化合物中通常存在大量重叠,但在谱中也存在差异。
结合相互作用还可使用无标记方法(如表面等离子体共振(spr)和质谱分析法)来检测。spr可提供对解离常数和解离率的测量。例如,a-100biocore/ge仪器适合于这种类型的分析。flex芯片(flexchip)可用于在同一支持体上分析多达400个结合反应。
任选地,样品成分与阵列之间的结合相互作用可以以竞争形式来检测。在存在与不存在结合的竞争性抑制剂的情况下阵列与样品的结合谱的差异在表征样品时可能是有用的。该竞争性抑制剂可以是,例如,与疾病状况相关的已知蛋白质,如病原体或病原体的抗体。在这样的竞争剂的存在下阵列成员与样品的结合的降低提供了存在该病原体的指示。可通过改变盐、离子强度、有机溶剂含量和文库成员与靶标接触时的温度来调节严格性。
基于抗体的检测方法,如酶联免疫吸附测定(elisa)方法,可用于检测与本发明阵列的结合模式。例如,检测免疫球蛋白的特定同种型(例如,igm同种型)的第二抗体可用于检测来自受试者的复杂生物样品的多种igm抗体与阵列的结合模式。例如,第二抗体可以缀合至可检测标记物,如荧光部分或放射性标记物。
本发明提供了用于检测多种不同抗体与本发明阵列的脱靶结合的阵列和方法。复杂生物样品中的多种抗体能够与肽微阵列中的多个肽脱靶结合。在一些实施方案中,检测至少一种抗体与肽阵列中的多个肽的脱靶结合可形成免疫特征。在其他实施方案中,检测至少一种抗体与肽阵列中的至少10个不同肽的脱靶结合可形成免疫特征。在其他实施方案中,检测至少一种抗体与肽阵列中的至少20个不同肽的脱靶结合可形成免疫特征。在其他实施方案中,检测至少一种抗体与肽阵列中的至少50个不同肽的脱靶结合可形成免疫特征。在其他实施方案中,检测至少一种抗体与肽阵列中的至少100个不同肽的脱靶结合可形成免疫特征。抗体的众多类别或同种型可提供与阵列的脱靶结合模式。抗体或免疫球蛋白可以为iga、igd、ige、igg和/或igm抗体。
来自复杂生物样品的至少一种igm抗体与肽阵列的结合模式可形成免疫特征。igm抗体可形成聚合物,在该聚合物中多个免疫球蛋白通过二硫键共价连接在一起。igm聚合物可为五聚体。igm同种型的抗体的聚合性质可增强样品与阵列的脱靶结合。抗体的聚合性质可提高样品与阵列的结合亲合力。例如,聚合igm同种型抗体的抗体与肽微阵列的结合模式可形成独特的五聚体驱动的免疫特征。
iga抗体可以是iga1或iga2抗体。iga同种型的抗体可形成二聚体。igg抗体可以是igg1、igg2、igg3或igg4抗体。igg同种型的抗体可作为单体存在。igd和/或ige抗体可形成单体。在一些实施方案中,本发明可检测来自受试者的复杂生物样品的至少一种igm抗体与肽阵列的脱靶结合。
治疗和状况
本发明的方法和阵列提供了用于鉴定治疗靶标和/或疫苗的灵敏方法。本发明的方法和阵列可用于筛选针对癌症和/或免疫病症或状况的治疗靶标。本发明的阵列和方法可用于,例如,鉴定受试者的多种不同状况的治疗靶标。受试者可以是人、豚鼠、狗、猫、马、小鼠、兔和各种其他动物。受试者可为任何年龄,例如,受试者可为婴儿、学步孩童、儿童、青春期前儿童、青少年、成年或老年个体。
受试者的状况可对应于疾病或健康状况。在一些实施方案中,受试者的状况为健康状况,并且本发明的方法监测该健康状况。在一些实施方案中,受试者的状况为疾病状况,并且本发明的方法用于诊断/监测该状况的状态和/或进展。本发明的方法还可用于预防状况。在一些实施方案中,本发明的方法与预防性处理结合使用。
在一些实施方案中,本发明的方法是一种鉴定针对癌症的治疗靶标的方法,该方法包括:a.使肽阵列与来自患有感兴趣的已知癌症的个体的第一生物样品接触;b.检测第一生物样品中的抗体与所述肽阵列的结合以获得第一免疫特征谱;c.使肽阵列与来源于没有已知癌症的个体的对照样品接触;d.检测所述对照样品中的抗体与所述肽阵列的结合以获得第二免疫特征谱;e.将第一免疫特征谱与第二免疫特征谱进行比较,并鉴定与第二免疫特征谱相比在第一免疫特征谱中结合更少或更多抗体的差异结合的肽;以及f.将与所鉴定的差异结合的肽相对应的蛋白质鉴定为针对感兴趣的癌症的靶标。
本发明的阵列和方法可用于,例如,鉴定治疗靶标、疫苗,或筛选针对癌症的治疗靶标。可用本发明的阵列和方法诊断、监测、预防和/或治疗的癌症的非限制性实例可包括:急性淋巴母细胞性白血病、急性髓样白血病、肾上腺皮质癌、aids相关癌症、aids相关淋巴瘤、肛门癌、阑尾癌、星形细胞瘤、基底细胞癌、胆管癌、膀胱癌、骨癌、脑肿瘤(如小脑星形细胞瘤、大脑星形细胞瘤/恶性胶质瘤)、室管膜瘤、髓母细胞瘤、幕上原始神经外胚层肿瘤、视觉通路和下丘脑神经胶质瘤、乳腺癌、支气管腺瘤、伯基特淋巴瘤(burkittlymphoma)、未知原发来源的癌症、中枢神经系统淋巴瘤、小脑星形细胞瘤、宫颈癌、儿童期癌症、慢性淋巴细胞性白血病、慢性髓细胞性白血病、慢性骨髓增生性病症、结肠癌、皮肤t细胞淋巴瘤、促结缔组织增生性小圆细胞肿瘤、子宫内膜癌、室管膜瘤、食管癌、尤因氏肉瘤(ewing'ssarcoma)、生殖细胞肿瘤、胆囊癌、胃癌、胃肠道类癌瘤、胃肠道间质瘤、神经胶质瘤、毛细胞白血病、头颈癌、心脏癌、肝细胞(肝)癌、霍奇金淋巴瘤、下咽癌、眼内黑素瘤、胰岛细胞癌、卡波西肉瘤、肾癌、喉癌、唇及口腔癌、脂肪肉瘤、肝癌、肺癌(例如非小细胞和小细胞肺癌)、淋巴瘤、白血病、巨球蛋白血症、骨恶性纤维组织细胞瘤/骨肉瘤、髓母细胞瘤、黑素瘤、间皮瘤、隐性原发性转移性鳞状颈癌、口癌、多发性内分泌肿瘤综合征、骨髓增生异常综合征、髓样白血病、鼻腔和副鼻窦癌、鼻咽癌、神经母细胞瘤、非霍奇金淋巴瘤、非小细胞肺癌、口腔癌、口咽癌、骨肉瘤/骨恶性纤维组织细胞瘤、卵巢癌、卵巢上皮瘤、卵巢生殖细胞肿瘤、胰腺癌、胰岛细胞胰腺癌、副鼻窦和鼻腔癌、甲状旁腺癌、阴茎癌、咽癌、嗜铬细胞瘤、松果体星形细胞瘤(pinealastrocytoma)、松果体生殖细胞瘤(pinealgerminoma)、垂体腺瘤、胸膜肺母细胞瘤、浆细胞瘤、原发性中枢神经系统淋巴瘤、前列腺癌、直肠癌、肾细胞癌、肾盂和输尿管移行细胞癌、视网膜母细胞瘤、横纹肌肉瘤、唾液腺癌、肉瘤、皮肤癌、梅克尔细胞皮肤癌(skincarcinomamerkelcell)、小肠癌、软组织肉瘤、鳞状细胞癌、胃癌、t细胞淋巴瘤、喉癌、胸腺瘤、胸腺癌、甲状腺癌、滋养细胞肿瘤(妊娠期)、未知原发部位的癌症、尿道癌、子宫肉瘤、阴道癌、外阴癌、瓦尔登斯特伦巨球蛋白血症(
在一些实施方案中,本发明的阵列和方法提供了鉴定针对癌症的治疗靶标的方法,其中所述癌症选自肺癌、白血病、胰腺癌、前列腺癌、乳腺癌、膀胱癌、子宫内膜癌和结肠直肠癌。
在一些实施方案中,本发明的方法是一种用于鉴定针对自身免疫病症的治疗靶标的方法,该方法包括:a)使肽阵列与来自患有感兴趣的已知自身免疫病症的个体的第一生物样品接触;b)检测第一生物样品中的抗体与所述肽阵列的结合以获得第一免疫特征谱;c)使肽阵列与来源于没有已知自身免疫病症的个体的对照样品接触;d)检测所述对照样品中的抗体与所述肽阵列的结合以获得第二免疫特征谱;e)将第一免疫特征谱与第二免疫特征谱进行比较,并鉴定与第二免疫特征谱相比在第一免疫特征谱中结合更少或更多抗体的差异结合的肽;以及f)将与所鉴定的差异结合的肽相对应的蛋白质鉴定为针对感兴趣的自身免疫病症的靶标。
在一些实施方案中,本发明的方法可用于,例如,鉴定治疗靶标、疫苗,或筛选针对免疫病症的治疗靶标。与免疫系统相关的病症的非限制性实例可包括:自身免疫病症、炎性疾病、hiv、类风湿性关节炎、i型糖尿病、系统性红斑狼疮、硬皮病、多发性硬化、重症联合免疫缺陷(scid)、迪乔治综合征(digeorgesyndrome)、共济失调性毛细血管扩张症(ataxia-telangiectasia)、季节性变态反应、常年性变态反应、食物变态反应、过敏反应(anaphylaxis)、肥大细胞增多症、变应性鼻炎、特应性皮炎、帕金森病、阿尔茨海默病、脾功能亢进、白细胞粘附缺陷症、x连锁淋巴增生性疾病、x连锁无丙种球蛋白血症、选择性免疫球蛋白a缺乏症、高igm综合征、自身免疫性淋巴增生性综合征、wiskott-aldrich综合征、慢性肉芽肿病、常见变异型免疫缺陷病(cvid)、高免疫球蛋白e综合征和桥本甲状腺炎(hashimoto'sthyroiditis)。
在一些实施方案中,所述免疫病症为自身免疫病症。在一些实施方案中,所述自身免疫病症选自i型糖尿病、类风湿性关节炎、多发性硬化、炎性肠病、系统性红斑狼疮、银屑病和硬皮病。
本发明可提供一种预防状况的方法,该方法包括:a)提供来自受试者的复杂生物样品;b)使所述复杂生物样品与肽阵列接触,其中所述肽阵列包含能够脱靶结合复杂生物样品中的至少一种抗体的不同肽;c)测量所述复杂生物样品与多个不同肽的脱靶结合以形成免疫特征;d)将所述免疫特征与状况相关联;以及e)接受针对所述状况的处理。
在一些实施方案中,本发明的方法可与预防性处理结合使用。例如,疫苗可以是预防性处理。起到预防性处理作用的疫苗的非限制性实例包括脊髓灰质炎疫苗、天花疫苗、麻疹疫苗、腮腺炎疫苗、人乳头瘤病毒(hpv)疫苗和流感疫苗。在一些实施方案中,本发明的方法用于监测,例如,受试者对预防性疫苗的应答。
在一些实施方案中,本发明提供了一种鉴定疫苗靶标的方法,该方法包括:a.使肽阵列与来自具有感兴趣的已知状况的个体的第一生物样品接触;b.检测第一生物样品中的抗体与所述肽阵列的结合以获得第一免疫特征谱;c.使肽阵列与来源于没有已知状况的个体的对照样品接触;d.检测所述对照样品中的抗体与所述肽阵列的结合以获得第二免疫特征谱;e.将第一免疫特征谱与第二免疫特征谱进行比较,并鉴定与第二免疫特征谱相比在第一免疫特征谱中结合更少或更多抗体的差异结合的肽;以及f.将与所鉴定的差异结合的肽相对应的蛋白质鉴定为针对感兴趣的状况的疫苗靶标。
在一些实施方案中,所述疫苗对抗病原体、微生物有机体、病毒、癌症或自身免疫病症。病原体可为病原性病毒或病原性细菌。病原性病毒和/或病原性细菌的感染可引起状况,例如,炎症。病原性细菌的非限制性实例可见于:a)博德特氏菌属(bordetellagenus),如百日咳博德特氏菌(bordetellapertussis)种;b)疏螺旋体属(borreliagenus),如布氏疏螺旋体(borreliaburgdorferi)种;c)布鲁氏菌属(bruceliagenus),如流产布鲁氏菌(brucellaabortus)、狗布鲁氏菌(brucellacanis)、羊布鲁氏菌(brucelameliterisis)和/或猪布鲁氏菌(brucellasuis)种;d)弯曲杆菌属(campylobactergenus),如空肠弯曲杆菌(campylobacterjejuni)种;e)衣原体和嗜衣原体属(chlamydiaandchlamydophilagenus),如肺炎衣原体(chlamydiapneumonia)、沙眼衣原体(chlamydiatrachomatis)和/或鹦鹉热嗜衣原体(chlamydophilapsittaci)种;f)梭菌属(clostridiumgenus),如肉毒梭菌(clostridiumbotulinum)、艰难梭菌(clostridiumdifficile)、产气荚膜梭菌(clostridiumperfringens)、破伤风梭菌(clostridiumtetani)种;g)棒状杆菌属(corynebacteriumgenus),如白喉棒状杆菌(corynebacteriumdiphtheria)种;h)肠球菌属(enterococcusgenus),如粪肠球菌(enterococcusfaecalis)和/或屎肠球菌(enterococcusfaecium)种;i)埃希氏菌属(escherichiagenus),如大肠杆菌(escherichiacoli)种;j)弗朗西丝氏菌属(francisellagenus),如土拉热弗朗西丝氏菌(francisellatularensis)种;k)嗜血杆菌属(haemophilusgenus),如流感嗜血杆菌(haemophilusinfluenza)种;l)螺杆菌属(helicobactergenus),如幽门螺杆菌(helicobacterpylori)种;m)军团菌属(legionellagenus),如嗜肺军团菌(legionellapneumophila)种;n)钩端螺旋体属(leptospiragenus),如问号钩端螺旋体(leptospirainterrogans)种;o)利斯特氏菌属(listeriagenus),单核细胞增生利斯特氏菌(listeriamonocytogenes)种;p)分枝杆菌属(mycobacteriumgenus),如麻风分枝杆菌(mycobacteriumleprae)、结核分支杆菌(mycobacteriumtuberculosis)和/或溃疡分枝杆菌(mycobacteriumulcerans)种;q)支原体属(mycoplasmagenus),如肺炎支原体(mycoplasmapneumonia)种;r)奈瑟氏球菌属(neisseriagenus),如淋病奈瑟氏球菌(neisseriagonorrhoeae)和/或脑膜炎奈瑟氏球菌(neisseriameningitidia)种;s)假单胞菌属(pseudomonasgenus),如绿脓假单胞菌(pseudomonasaeruginosa)种;t)立克次氏体属(rickettsiagenus),如立氏立克次氏体(rickettsiarickettsii)种;u)沙门氏菌属(salmonellagenus),如伤寒沙门氏菌(salmonellatyphi)和/或鼠伤寒沙门氏菌(salmonellatyphimurium)种;v)志贺氏菌属(shigellagenus),如索氏志贺氏菌(shigellasonnei)种;w)葡萄球菌属(staphylococcusgenus),如金黄色葡萄球菌(staphylococcusaureus)、表皮葡萄球菌(staphylococcusepidermidis)和/或腐生葡萄球菌(staphylococcussaprophyticus)种;x)链球菌属(streptpcoccusgenus),如无乳链球菌(streptococcusagalactiae)、肺炎链球菌(streptococcuspneumonia)和/或酿脓链球菌(streptococcuspyogenes)种;y)密螺旋体属(treponemagenus),如苍白密螺旋体(treponemapallidum)种;z)弧菌属(vibriogenus),如霍乱弧菌(vibriocholera);和/或aa)耶尔森氏菌属(yersiniagenus),如鼠疫耶尔森氏菌(yersiniapestis)种。
病毒的非限制性实例可见于以下病毒科,并用示例性种来说明:a)腺病毒科(adenoviridae),如腺病毒种;b)疱疹病毒科(herpesviridae),如1型单纯疱疹、2型单纯疱疹、水痘-带状疱疹病毒、eb病毒(epstein-barrvirus)、人巨细胞病毒、8型人疱疹病毒种;c)乳头瘤病毒科(papillomaviridae),如人乳头瘤病毒种;d)多瘤病毒科(polyomaviridae),如bk病毒、jc病毒种;e)痘病毒科(poxviridae),如天花种;f)肝脱氧核糖核酸病毒科(hepadnaviridae),如乙型肝炎病毒种;g)细小病毒科(parvoviridae),如人博卡病毒、细小病毒b19种;h)星形病毒科(astroviridae),如人星形病毒种;i)嵌杯样病毒科(caliciviridae),如诺沃克病毒(norwalkvirus)种;j)黄病毒科(flaviviridae),如丙型肝炎病毒、黄热病病毒、登革热病毒、西尼罗病毒种;k)披膜病毒科(togaviridae),如风疹病毒种;l)戊型肝炎病毒科(hepeviridae),如戊型肝炎病毒种;m)逆转录病毒科(retroviridae),如人免疫缺陷病毒(hiv)种;n)正粘病毒科(orthomyxoviridaw),如流感病毒种;o)砂粒样病毒科(arenaviridae),如瓜纳里托病毒(guanaritovirus)、呼宁病毒(juninvirus)、拉沙病毒(lassavirus)、马秋博病毒(machupovirus)和/或萨比亚病毒(sabiávirus)种;p)本扬病毒科(bunyaviridae),如克里米亚-刚果出血热病毒(crimean-congohemorrhagicfevervirus)种;q)纤丝病毒科(filoviridae),如埃博拉病毒和/或马尔堡病毒(marburgvirus)种;副粘病毒科(paramyxoviridae),如麻疹病毒、腮腺炎病毒、副流感病毒、呼吸道合胞体病毒、人偏肺病毒、亨德拉病毒(hendravirus)和/或尼帕病毒(nipahvirus)种;r)弹状病毒科(rhabdoviridae)属,如狂犬病病毒种;s)呼肠孤病毒科(reoviridae),如轮状病毒、环状病毒、科罗拉多壁虱热病毒(coltivirus)和/或版纳病毒(bannavirus)种。在一些实施方案中,未指定病毒的病毒科,如丁型肝炎。
在一些实施方案中,所述疫苗对抗微生物有机体。微生物有机体可以是单细胞或多细胞有机体。微生物有机体可以是细菌、病毒、真菌或原生动物。
在一些实施方案中,本发明提供了一种提供治疗的方法,该方法包括:a)接收来自受试者的复杂生物样品;b)使复杂生物样品与肽阵列接触,其中所述肽阵列包含能够脱靶结合生物样品中的至少一种抗体的不同肽;c)测量抗体与多个不同肽的脱靶结合以形成免疫特征;d)将所述免疫特征与状况相关联;以及e)提供针对所述状况的治疗。
在一些实施方案中,本发明可提供一种诊断方法,该方法包括:a)接收来自受试者的复杂生物样品;b)使所述复杂生物样品与肽阵列接触,其中所述肽阵列包含能够脱靶结合生物样品中的至少一种抗体的不同肽;c)测量抗体与肽阵列中的一组不同肽的脱靶结合以形成免疫特征;以及d)基于所述免疫特征诊断状况。
在一些实施方案中,本发明的方法可用作诊断、监测和治疗状况的方法。治疗状况的方法可能需要开出以治疗受试者的状况或疾病为目标的治疗剂的处方。在一些实施方案中,可以以下述剂量范围开出治疗剂:约1mg至约2000mg;约5mg至约1000mg、约10mg至约500mg、约50mg至约250mg、约100mg至约200mg、约1mg至约50mg、约50mg至约100mg、约100mg至约150mg、约150mg至约200mg、约200mg至约250mg、约250mg至约300mg、约300mg至约350mg、约350mg至约400mg、约400mg至约450mg、约450mg至约500mg、约500mg至约550mg、约550mg至约600mg、约600mg至约650mg、约650mg至约700mg、约700mg至约750mg、约750mg至约800mg、约800mg至约850mg、约850mg至约900mg、约900mg至约950mg或约950mg至约1000mg。
在一些实施方案中,开出至少1mg、至少5mg、至少15mg、至少15mg、至少20mg、至少25mg、至少30mg、至少35mg、至少40mg、至少45mg、至少50mg、至少55mg、至少60mg、至少65mg、至少70mg、至少80mg、至少85mg、至少90mg、至少100mg、至少150mg、至少200mg、至少250mg、至少300mg、至少350mg、至少400mg、至少450mg、至少500mg、至少550mg、至少600mg、至少650mg、至少700mg、至少750mg、至少800mg、至少850mg、至少900mg、至少950mg或至少1000mg的治疗剂。
本发明的阵列和方法可由使用者使用。众多使用者可使用本发明的方法来鉴定和/或提供状况的治疗。使用者可以是,例如,希望监测其自身健康的人。使用者可以是,例如,卫生保健提供者。卫生保健提供者可以是,例如,医师。在一些实施方案中,使用者是照顾受试者的卫生保健提供者。可以是本发明的使用者的医师和卫生保健提供者的非限制性实例可包括麻醉师、减肥手术专业人员、血库输血医学专业人员、心脏电生理学医生、心脏外科医生、心脏病科医生、注册护理助理、临床心脏电生理学专业人员、临床神经生理学专业人员、临床护理专业人员、结直肠外科医生、重症护理医学专业人员、重症护理外科手术专业人员、牙科保健员、牙科医生、皮肤科医师、急诊医疗技师、急诊医学医师、胃肠外科医生、血液病医生、临终关怀和姑息医学专业人员、顺势疗法专业人员、传染病专业人员、内科医生、颌面外科医生、医务助理、体检医生(medicalexaminer)、医学遗传学家、医疗肿瘤学家、助产师、新生儿-围产期专业人员、肾病医生、神经科医生、神经外科医生、核医学专业人员、护士、开业护士、产科医师、肿瘤科医生、口腔外科医生、口腔正畸医师、整形外科专业人员、疼痛控制专业人员、病理学医师、儿科医师、灌注师、牙周病医师、整形外科医生、足病医生、直肠科医生、假肢专业人员、精神科医生、肺脏科医生、放射科医生(radiologist)、外科医生、胸科专业人员、移植专业人员、血管专业人员、血管外科医生和兽医。利用本发明的阵列和方法确定的诊断可并入受试者的病历中。获得的免疫特征随后可用于鉴定治疗靶标并针对根据本文公开的方法和装置鉴定的病症开发对于个体的治疗。
本文公开的阵列装置和方法提供了同时从多种疾病和/或状况鉴定治疗靶标和/或疫苗的灵敏方法。本发明的方法和阵列还提供了同时从多种疾病和/或状况筛选针对癌症和/或免疫病症或状况的治疗靶标的灵敏方法。例如,本文公开的阵列装置和方法能够在同一阵列上同时检测炎性状况、癌症疾病和病原性感染。因此,检测广谱的疾病和状况仅需要一个阵列,即一个免疫特征测定。因此,在受试者的整个生命期间对受试者的监测将利用所进行的每次免疫特征测定来提供受试者的健康状态随时间的快照。这提供了检测受试者健康状态的总体和具体改变的有力手段,并且与高灵敏度的免疫特征测定一起提供了一种能够非常早期地检测个体健康状态的任何变化的系统。
因此,本文公开的方法、系统和阵列装置能够筛选、鉴定治疗靶标,鉴定疫苗靶标,和/或在疾病和/或状况的早期阶段治疗该疾病和/或状况。例如,本文公开的方法、系统和阵列装置能够在传统的基于生物标志物的测定之前数天或数周检测、诊断和监测疾病和/或状况。此外,仅需要一个阵列,即一个免疫特征测定,即可检测、诊断和监测疾病和状况(包括炎性状况、癌症和病原性感染)的副谱(sidespectra)。
分类算法
多种算法和分类器可用于对在免疫特征分析阵列中获得的数据进行分类和/或分析。朴素贝叶斯(
实施例
实施例1:
以下实施例描述了用于鉴定肽阵列中的肽的示例性系统和方法。
破译在i型糖尿病免疫特征中的肽的身份。
实施例1阐述了用于鉴定i型糖尿病免疫特征的方法和系统。将来自40名i型糖尿病受试者和40名年龄匹配的对照受试者的血清施加到包含约10,000个肽的随机肽微阵列上。检测并量化血清中的抗体的结合模式。使用t-检验来选择满足至少p<0.0001假发现率的阈值的肽。在该假发现率下,在40名i型糖尿病受试者和40名年龄匹配的对照受试者中鉴定出689种差异肽。表1描述了不同的肽在i型糖尿病的免疫特征分析中的性能测量值,以及这些差异肽的分类准确度、特异性和灵敏度。表1的说明:a表示从10,000种中(双尾(2-tail))选出的肽的数目,它们根据welsht-检验具有小于0.0001的总体i型误差率且在2种条件下有差异。b表示通过朴素贝叶斯分类算法对40名i型糖尿病和40名对照受试者进行弃一法交叉验证(leave-one-outcrossvalidation)所获得的性能测量值。
我们通过免疫特征分析使用朴素贝叶斯分类算法弃一法交叉验证能够获得>90%的特异性。弃一法交叉验证是一种消除分类准确度的方法,在该方法中将一个个体排除在样品组之外。将该算法用该组的其余成员进行训练,并用于预测弃除的样品。依次测试所有样品并计算百分比准确度。679种差异肽的分布有偏斜,与对照相比,只有210种肽上调,而479种肽下调。我们然后检查有多少肽可以经生物信息学方式被定位(map)到i型糖尿病中的已知抗原。
i型糖尿病免疫特征的破译
我们首先考虑i型糖尿病中的210种上调的肽,并使用表位匹配工具guitope将其以生物信息学方式定位至8种已知的i型糖尿病抗原。这8种已知抗原的蛋白质序列与210种肽同源性匹配。选择出以p<0.0001,与来自我们10,000种肽的文库的相等数目的肽在蛋白质的区域(表位)上显示出显著匹配的肽。
平均而言,我们能够成功地将来自210种肽的肽定位到已知抗原,每个抗原具有1-2个表位。抗原的一个表位平均定位3-4种肽。从上调的这组肽中针对抗原平均定位了7种肽。
表2对应于来自免疫特征的210种上调肽向i型糖尿病中的8种已知抗原的定位。表2显示了对于210种上调肽在i型糖尿病中的8种已知抗原上,表位的数目、每个表位的肽的数目以及每种抗原所定位的肽总数。表2的说明:a表示i型糖尿病中已报道的自身蛋白质/抗原。
b表示对蛋白质使用guitope以生物信息学方式观察到的表位的总数。
c表示针对蛋白质的特定表位e1/e2所定位的肽的数目。
d表示i型糖尿病的210种上调肽中定位至抗原的肽的总数。e表示错误发现率,即当从10,000个随机肽的文库中选择同等数目的肽时观察到针对抗原的等同匹配的概率。
接下来,我们考虑其余479种下调肽并将它们定位至i型糖尿病中已知的8种抗原。我们对此的假设示于图1中。图1为显示自身蛋白质/抗原如何能在随机序列肽阵列中引起免疫特征分析的上调和下调的途径的示意图。图1通过示出具有两个表位的自身蛋白质阐明了疾病相关蛋白质包含多个表位的概念。在图1中,包含两个表位的自身蛋白质遇到抗原呈递细胞并激活b细胞。表位1导致b细胞的激活,b细胞开始产生对表位1具有反应性的抗体,然后通过抗体反应性肽上信号的增加在肽阵列上检测到这些抗体。相反地,当表位2激活b细胞时存在表位特异性调节性t细胞(treg),该treg抑制b细胞的抗体分泌。因此当抗体在循环中被消耗时它们没有被补充,从而导致这些抗体与本发明阵列的结合的下调或缺乏。
表3示出了定位至i型糖尿病中的8种已知抗原的来自免疫特征的479种下调肽。在下调肽中,肽仅定位到gad-65、znt8和胰岛素,这些是i型糖尿病中的强抗原。与上调组的肽相比,下调中被定位的肽的数目更多,但对于上调和下调来说,每种抗原的表位的数目都是相似的。两种情况下定位到抗原的肽不仅因为它们来自两个不同的组而不同,而且它们在蛋白质序列上定位的表位也是不同的。
因此,抗原的特定表位可导致肽的上调,而在相同蛋白质上的其他表位可导致某一其他组的肽的下调。表3.来自免疫特征的479种下调肽向i型糖尿病中的8种已知抗原的定位。表3的说明:a表示i型糖尿病中已报道的自身蛋白质/抗原。b表示对蛋白质使用guitope以生物信息学方式观察到的表位的总数。c表示针对蛋白质的特定表位e1/e2所定位的肽的数目。d表示i型糖尿病的479种下调肽中定位到抗原的肽的总数。
e表示错误发现率,即当从10,000个随机肽的文库中选择同等数目的肽时观察到针对抗原的等同匹配的概率。
为了建立空对照(nullcontrol)和方法验证,我们针对8种随机蛋白质定位有差异的(上调和下调的)肽,其中一些蛋白质与酶有关,其余的是随机的,并不参与抗体产生或i型糖尿病途径。
为了提供验证并证实针对已知抗原所定位的肽,进行正交测量。图2为盒型图,其显示了2组i型糖尿病患者在具有高和低gad-65放射免疫沉淀(rip)滴度时的13种肽的强度。对个体t1d患者进行在cim1ok阵列上的免疫特征分析测定。生物信息学选择导致鉴定出定位回gad-65的13种肽。将个体患者血清对13种肽的反应性取平均并将组值示于盒型图中。基于高或低抗gad-65滴度定义各组。两组的平均值之间存在显著性差异(p<0.001)。图2表明没有肽被定位到8种随机蛋白质的组。
表4.来自i型糖尿病的免疫特征的679种差异肽向8种随机蛋白质/抗原的定位。表4的说明:a表示作为阴性对照的一部分的随机蛋白质/自身抗原。b表示对蛋白质使用guitope以生物信息学方式观察到的表位的总数。c表示在i型糖尿病的所有689种差异肽(上调和下调的)中的肽的总数。d表示当从10,000个随机肽的文库中选择同等数目的肽时观察到针对相同抗原的等同匹配的错误发现率。
正交测量和结果验证
我们使用放射免疫沉淀(rip)测定来测量gad-65滴度,并将40个i型样品分成高/低gad-65滴度的两个类别,并且探寻在13种定位的肽的强度与gad-65和rip滴度之间是否存在任何对应关系。图2示出了13种肽在高和低滴度gad-65上的强度的平均值的盒型图,表明在两组的平均值之间存在显著性差异(p<0.001)。我们然后探寻在gad-65r.i.p滴度与13种肽的强度的相应平均值之间是否存在任何线性对应关系。
图3示出了免疫特征阵列上的肽强度与抗gad滴度之间的关系的线性回归分析。图3中具有线性回归线的散点图显示了gad-65rip滴度和log肽强度的95%置信带。回归线的斜率指示线性关系。gad-65的定位的肽显示了与rip滴度的线性对应关系。
鼠乳腺癌模型免疫特征的破译
对乳腺癌新型疗法和疫苗候选物的鉴定存在迫切需求。本实施例描述了本发明的方法和系统在her2/neut转基因鼠乳腺癌模型表征中的应用。将免疫特征分析应用于野生型(健康小鼠)与易患乳腺癌的小鼠(her2/neut小鼠)的比较分析中。
图4示出了对作为感兴趣的肽的ksfhgrviqdvvgepyggsc(seqidno.1)的破译/鉴定。在cim10k示例阵列上比较年龄匹配的野生型balb/c小鼠10(n=8)与转基因balb/c-neut小鼠(n=11)的免疫特征。小鼠为10、15或16周龄。转基因balb/c-neut小鼠在采血时没有可感知的肿瘤。使用小于0.005的t-检验p值选出23种肽。这23种肽能够使用朴素贝叶斯分类算法对个体小鼠进行分类,得到89%正确率的弃一法交叉验证。23种肽被鉴定为在组间具有显著性,p<0.005(图4,a图),并且能够采用朴素贝叶斯模型中的弃一法交叉验证,基于疾病状态区别个体小鼠。这些肽中的一种肽在除了一个之外的所有野生型中相对于转基因小鼠强烈上调(图4,b图)。图4的a图为散点图,其示出了野生型与转基因小鼠的归一化强度的生物信息学平均值之间的肽关系。图4的a图中的箭头指向与转基因小鼠相比在野生型小鼠中显著升高的肽(p=2.33×10-5)。
图4的b图示出了绘制为线型图的个体小鼠的归一化强度,图中线条代表单个肽,而灰色的垂直图线代表单个小鼠。ksfhgrviqdvvgepyggsc(seqidno.1)肽被绘制为黑线并且在除了一只之外的所有小鼠中恒定地升高。
将这种肽定位回两个推定的靶标:p190rhogap和tubby候选基因。图5示出了肽ksfhgrviqdvvgepyggsc(seqidno.1)与前两种小鼠蛋白质组候选物的blast比对。图5的a图为最可能的匹配即p190rhogap的blast比对。图5的b图为第二可能的匹配即tubby候选基因的blast比对。使用已鉴定的肽序列来搜索经翻译的小家鼠(musmusculus)基因组,build37.2,以使用2011年10月28日在线访问的ncbi基本局部比对搜索工具(ncbibasiclocalalignmentsearchtool)进行匹配。
插入转基因小鼠中的her2基因通过p190rhogap进行信号传导,而tubby先前已与同视网膜病相关的癌症相关联。
破译小鼠的流感a/pr/8/34感染和klh免疫的免疫特征
为了证明破译方法在免疫期间鉴定免疫原性靶标的能力,进行了表征感染流感(h1n1)a/pr/8/34或用klh免疫的balb/c小鼠的免疫特征的实验。在第0天、第14天和第28天向用klh免疫的小鼠给予在明矾中的100μgklh。对感染的小鼠给予l×104个病毒颗粒的亚致死鼻内剂量。两组小鼠都在第35天取血,且在本发明的示例性阵列(在这种情况下为cim10k阵列)上操作血清。使用guitope程序进行的分析揭示了klh免疫的小鼠中一种感兴趣的表位(表5)。表5以表格形式显示了guitope分析的结果,其在p<0.005下定位了与pbs(模拟(mock))免疫的小鼠相比来自klh免疫的小鼠的免疫特征分析的305种差异肽。表5的说明:a表示3只小鼠中的klh免疫。b表示与pbs免疫的小鼠(模拟)相比上调和下调的肽的数目。c表示对蛋白质使用guitope以生物信息学方式观察到的表位的总数。d表示定位到第一表位的肽的数目。e表示在klh免疫的小鼠中上调和下调的定位的肽的总数。f表示当从10,000个随机肽的文库中选择同等数目的肽时观察到针对相同抗原的等同匹配的错误发现率。
使用guitope程序进行的分析揭示了感染流感的小鼠中的一种越来越多地被识别的表位以及一种越来越少地被识别的表位(表6)。表6以表格形式显示了guitope分析的结果,其在p<0.01下定位了与pbs(模拟)免疫的小鼠相比来自pr8免疫的小鼠的免疫特征分析的325种差异肽。表6的说明:a表示小鼠中的a/pr/8/34免疫。b表示与pbs免疫的小鼠(模拟)相比上调和下调的肽的数目。c表示对蛋白质使用guitope以生物信息学方式观察到的表位的总数。d表示定位到第一表位的肽的数目。e表示在pr8免疫的小鼠中上调和下调的定位的肽的总数。f表示当从10,000个随机肽的文库中选择同等数目的肽时观察到针对相同抗原的等同匹配的错误发现率。
实施例2:与免疫特征系统一起使用的计算机架构
由本发明的阵列检测到的数据可通过具有不同计算机架构的多台计算机进行分析。图6为示出了可与本发明的示例实施方案结合使用的计算机系统600的第一示例架构的框图。如图6所示,示例计算机系统可包括用于处理指令的处理器602。处理器的非限制性实例包括:intelcorei7tm处理器、intelcorei5tm处理器、intelcorei3tm处理器、intelxeontm处理器、amdopterontm处理器、samsung32-bitriscarm1176jz(f)-sv1.0tm处理器、armcortex-a8samsungs5pc100tm处理器、armcortex-a8applea4tm处理器、marvellpxa930tm处理器或功能等同的处理器。多线程执行可用于并行处理。在一些实施方案中,可使用多个处理器或多核处理器,无论是在单一计算机系统中,在集群中,还是分布在经由网络包含多台计算机、蜂窝电话和/或个人数据助理设备的系统中。
数据采集、处理和存储
如图6所示,高速缓冲存储器601可连接到或并入处理器602,以便为处理器602最近使用或频繁使用的指令或数据提供高速存储。处理器602通过处理器总线605连接到北桥606。北桥606通过存储器总线604连接到随机存取存储器(ram)603,并且管理处理器602对ram603的访问。北桥606还通过芯片组总线607连接到南桥608。南桥608又连接至外围总线609。例如,外围总线可以是pci、pci-x、pciexpress或其他外围总线。北桥和南桥通常被称作处理器芯片组并管理处理器、ram和外围总线609上的外围组件之间的数据传送。在一些架构中,北桥的功能可并入处理器,而不是使用单独的北桥芯片。
在一些实施方案中,系统600可包括连接至外围总线609的加速器卡612。加速器可包括现场可编程门阵列(fpga)或用于加速某些处理的其他硬件。
软件接口
软件和数据被存储在外部存储613中,并且可加载到ram603和/或高速缓冲存储器601中以供处理器使用。系统600包括用于管理系统资源的操作系统;操作系统的非限制性实例包括:linux、windowstm、macostm、blackberryostm、iostm和其他功能等同的操作系统,以及在操作系统之上运行的应用软件。
在本实施例中,系统600还包括连接到外围总线的网络接口卡(nic)610和611,以用于提供通向外部存储(如网络附加存储(nas)和其他可用于分布式并行处理的计算机系统)的网络接口。
计算机系统
图7为示出了具有多个计算机系统702a和702b、多个蜂窝电话和个人数据助理702c以及网络附加存储(nas)701a和701b的网络700的示意图。在一些实施方案中,系统702a、702b和702c可管理数据存储并优化对存储在网络附加存储(nas)701a和702b中的数据的数据存取。数学模型可用于该数据,并使用在计算机系统702a和702b以及蜂窝电话和个人数据助理系统702c中的分布式并行处理进行评估。计算机系统702a和702b以及蜂窝电话和个人数据助理系统702c还可提供并行处理,以用于在网络附加存储(nas)701a和702b中存储的数据的适应性数据重构。图7仅示出了一个实例,而很多种其他计算机架构和系统可与本发明的各个实施方案一起使用。例如,刀片式服务器可用于提供并行处理。处理器刀片可通过底板连接以提供并行处理。存储还可连接至底板或通过单独的网络接口作为网络附加存储(nas)连接。
在一些实施方案中,处理器可保持单独的存储空间并通过网络接口、底板或其他连接器传输数据以便由其他处理器进行并行处理。在一些实施方案中,一些或所有处理器可使用共享虚拟地址存储空间。
虚拟系统
图8为使用共享虚拟地址存储空间的多处理器计算机系统的框图。该系统包括多个处理器801a-f,所述处理器可访问共享存储器子系统802。该系统将多个可编程的硬件存储器算法处理器(map)803a-f并入存储器子系统802中。各个map803a-f可包括存储器804a-f以及一个或多个现场可编程门阵列(fpga)805a-f。map提供可配置的功能单元,可向fpga805a-f提供特定算法或算法的一部分,以与相应的处理器密切配合地进行处理。在本实施例中,为了这些目的,每个map可被所有处理器全局访问。在一种配置中,每个map可使用直接存储器存取(dma)来访问相关存储器804a-f,从而允许其独立于相应的微处理器801a-f并与之异步地执行任务。在这种配置中,map可将结果直接提供给另一个map,以用于算法的流水操作(pipelining)及并行执行。
上述计算机架构和系统仅为示例,且很多种其他计算机、蜂窝电话以及个人数据助理架构和系统可与示例实施方案结合使用,包括使用通用处理器、协处理器、fpga和其他可编程逻辑设备、片上系统(systemonchip)(soc)、专用集成电路(asic)以及其他处理和逻辑元件的任意组合的系统。任何种类的数据存储介质均可与示例实施方案结合使用,包括随机存取存储器、硬盘驱动器、闪速存储器、磁带驱动器、磁盘阵列、网络附加存储(nas)及其他本地或分布式数据存储设备和系统。
在示例实施方案中,计算机系统可使用在任一上述或其他计算机架构和系统上执行的软件模块来实现。在其他实施方案中,该系统的功能可部分地或完全地在固件、可编程逻辑设备(例如图8中示出的现场可编程门阵列(fpga))、片上系统(soc)、专用集成电路(asic)或其他处理和逻辑元件中实现。例如,集处理器和优化器(setprocessorandoptimizer)可通过使用硬件加速器卡,例如图6中示出的加速器卡612,采用硬件加速来实现。
实施例3:免疫特征分析系统
下述实施例描述了用于免疫特征分析的自动化系统。
该自动化系统包括几个组件:1)用于接收、记录和稀释来自受试者的生物样品(如血液或唾液样品)的自动化系统。该自动化系统使生物样品与本发明的肽微阵列接触。
可在对受试者的免疫特征测定中获得受试者的免疫特征分析,该测定由以下自动化步骤组成:a)将稀释的样品施加至肽阵列;b)孵育特定时间;c)除去样品并洗涤该阵列;d)施加第二抗体溶液特定时间;e)用洗涤步骤除去未结合的和/或过量的第二抗体;以及f)干燥并扫描该阵列以测定斑点的荧光。图9为本发明的免疫特征分析系统的组件的示意图。
数据采集和分析
将阵列进行比对,并相对于标准特征确定特征。标准特征可以是健康受试者的特征或未结合的肽的参考信号。
基于利用本发明系统获得的免疫特征,可提供诊断。本发明的发现治疗靶标和破译免疫特征的方法可在任何位置进行。如图9所示,可将利用发现治疗靶标和破译的方法鉴定的数据传送到任何位置。在一些实施方案中,数据在不同社区、县、城市、州和/或国家间传送,例如,通过图9所示的云网络系统。
实施例4:发现和破译来自病原体的未注释的蛋白质
下述实施例描述了用于发现先前未鉴定的和/或未注释的蛋白质的本发明的方法的验证。将容易理解,本文所述的方法也可应用于未充分地或未能令人满意地表征的蛋白质的表征。
近来,jagger等人报道了pa-x蛋白质中的第二开放阅读框的鉴定,其在通过核糖体移码获取时产生具有生物活性的新型pa-x蛋白质(b.w.jagger等人,science337,1999,2012年7月13日;本文称为“jagger”)。我们在本实施例中通过证明免疫特征分析可应用于pa-x蛋白质的新型抗原的鉴定而显示了本发明的方法和系统作为抗原发现平台的应用。
使用鼠流感的长期免疫特征对pa-x移码的guitope搜索
使来自感染流感(h1n1)a/pr/8/34的小鼠的血清抗体与肽阵列接触,如legutki(j.b.legutki,d.m.magee,p.stafford,s.a.johnston,vaccine28,4529(2010年5月5日))所述,并且对cim10k阵列上的总计10,000个肽中的283个肽的强度进行关联。
检索来自a/pr/8/34的pa的genbank序列(登录号cy084187),并除去核苷酸598以再现jagger报道的移码。使用www.expasy.org上的在线翻译器来翻译所得到的核苷酸序列。将guitope软件(r.f.halperin,p.stafford,j.s.emery,k.a.navalkar,s.a.johnston,bmcbioinformatics13,1,2012)配置为使用在(j.b.legutki,d.m.magee,p.stafford,s.a.johnston,vaccine28,4529,2010年5月5日)中报道的156种未吸附的肽的列表作为输入序列,使用cim10k文库作为背景,并且使用翻译的pa-x蛋白质作为查询序列。在8个氨基酸的移动平均窗口上使用默认的替代矩阵、为1的反演权重(inversionweight)、扣除文库的得分和为10的截止比对得分来计算得分。
平铺pa-x移码阵列的构建和探查
将pa-x的orf-x部分分成4个平铺的肽,每一个均具有c末端gsc连接体,以促进与马来酰亚胺活化的氨基硅烷载玻片的附接。将载玻片活化并使用具有smp2针的nanoprint60打印。平铺的pa-x和对照肽以21向上格式(21upformat)打印,该格式的间隔适配array-it24向上微阵列盒。在加载到24向上盒中之前,将载玻片洗涤以除去未偶联的肽并封闭,如先前所述(j.b.legutki,d.m.magee,p.stafford,s.a.johnston,vaccine28,4529,2010年5月5日;以及b.a.chase,s.a.johnston,j.b.legutki,clinvaccineimmunol19,352,2012年3月)。所有的孵育均在室温下在摇动平台上进行。使用优化的洗涤方案在96针微孔板洗板机(biotekts405)上进行洗涤。血清在含有0.05%吐温的磷酸盐缓冲盐水(pbst)中的3%bsa中以1:500施加,每孔200ul。用5.0nm抗小鼠igg(h+l)-alexafluor647检测结合的血清抗体。将洗涤并干燥的载玻片在agilent‘c’型扫描仪中扫描,并使用genepix提取数据。从genepix结果文件中提取扣除背景的强度,并在excel中进行分析。将pearsonr2大于0.85的样品包括在最后的分析中。使用student双尾t检验以0.005的截止值来确定显著性。
对结合pa-x蛋白质的肽序列的鉴定
开发guitope软件以通过将随机序列肽与蛋白质序列进行比对使用免疫特征来预测表位(r.f.halperin,p.stafford,j.s.emery,k.a.navalkar,s.a.johnston,bmcbioinformatics13,1(2012))。我们使用流感(h1n1)a/pr/8/34基因组来翻译预测的pa-x转录物,并使用guitope软件搜索具有156种未吸附的肽的所得序列。将两种肽进行比对,其中扣除文库的得分为10.931(pamkhrephwvipgiiwgsc;seqidno.:2)和10.06(epmemhddrtmrpngafgsc,seqidno.:3),均大于在8个氨基酸的移动平均中使用的最小截止值10。
存在于每种肽上的gsc连接体并不包括在guitope搜索中以避免假象(artifact)。图12的a图中示出了这种比对,肽1的身份用加粗突出显示而肽2的身份用下划线突出显示。为了验证这种预测,合成代表来自a/pr/8/34的orf-x的四种平铺肽(图12,b图),并将其打印在小型微阵列上。预测的表位pa-x3越来越多地被感染a/pr/8/34后的小鼠的血清抗体结合(n=12,p=4.16×10-4)(图13)。阳性对照和阴性对照肽的表现如同所预测的一样。
实施例5:预测疫苗功效及发现表位
以下实施例描述了本发明的方法用于预测疫苗功效和靶标鉴定筛选的应用。
为了测试免疫特征分析预测疫苗功效的能力,使用具有已知个体结果的模型攻击系统。对五组各12只雌性balb/c小鼠进行免疫,单独取血,而后用流感h1n1a/pr/8/34(pr8)病毒攻击小鼠。三个组先前已经接种过灭活病毒。这些灭活病毒为福尔马林灭活的pr8(杀死的pr8),以及可购买到的2006/2007和2007/2008季节性三价流感疫苗。这两种季节性流感疫苗具有相同的a/wisconsin/67/2005(h3n2)和b/malaysia/2506/2004,但分别含有a/新喀里多尼亚(newcaledonia)/20/99和a/所罗门群岛(solomonislands)/3/2006而在h1n1部分上不同。除了模拟接种组外,还对第5组单次接种亚致死剂量的活pr8病毒。
用2-5mld的活pr8攻击小鼠,结果示于图14中。图14示出了经免疫的小鼠在采用流感a/pr/8/34进行致死攻击后的结果。用pbs(模拟)、亚致死剂量的活a/pr/8/34、灭活的a/pr/8/34、2006/2007季节性流感疫苗或2008/2007季节性流感疫苗对小鼠进行免疫。以每只小鼠5×105pfu经鼻内对小鼠进行攻击。相对于起始体重的平均每日百分比在a图中图示出,其中基于存活小鼠计算平均值,并且误差条代表标准差。存活曲线在b图中示出,并且代表攻击后存活的小鼠的百分比。
用亚致死攻击或杀死的pr8进行免疫的小鼠均没有死于该攻击,并且这些小鼠中没有小鼠具有有症状的感染,如由缺乏毛皮皱褶以及体重减轻所表明的。这两种季节性疫苗是部分保护性的,导致60%和80%的存活率。所有存活小鼠均具有显著的体重减轻。血清在攻击前两天采集并用于探查cim10k阵列以及建立免疫特征。
系统疫苗基线比较
为了创建用于与免疫特征分析进行比较的基线,对疫苗组的抗体和t细胞应答进行评估。在攻击前两天通过elisa评估抗病毒的血清抗体(图15)。图15示出了对免疫应答的传统测量。在攻击之前从所有小鼠中采集血清。通过终点滴度对灭活的pr8、2006-2007和2007-2008季节性疫苗测量抗原特异性循环igg的量,并将其示于a图中。误差条为合并血清的一式三份测量的标准差。对每组的两只小鼠施以安乐死,并且使用bioplexth1/th2细胞因子测定来测量用灭活病毒刺激的单独脾细胞和48小时分泌的细胞因子,并将测量结果示于(b图-f图)中。绘制的值代表四次测量的平均值+/-标准差。显著性表示如下:*表示相对于介质具有显著性,**表示相对于模拟免疫具有显著性。从a图中明显看出,仅有接受活疫苗或杀死的pr8疫苗的小鼠具有可检测的抗pr8抗体。对pr8攻击仅具有部分保护性的两种季节性疫苗没有可检测的针对pr8的应答。
为了评估t细胞应答,在攻击当天,处死每组的两只小鼠并收获脾脏。对于脾细胞刺激测定,在灭活全病毒的存在下每孔接种2.5×105个细胞。在48小时后,收获上清液,并使用bioradluminexth1/th2测定试剂盒测定细胞因子的分泌。分泌的细胞因子的浓度示于图15的a图-f图中。接受活疫苗的小鼠刺激强t细胞应答,从而分泌所测定的所有细胞因子,包括与th1和th2辅助细胞均相关的那些细胞因子。在灭活疫苗中,仅有杀死的pr8的接受者在受到灭活病毒刺激时分泌显著水平的所有细胞因子。季节性疫苗分泌il-4和il-5,表明其主要为cd4-th2应答。这些细胞因子谱证明,由杀死的pr8株产生的免疫应答为th2偏性的。这提示免疫环境有利于体液部分(humoralarm)的发展。在技术上不可能构建本研究中所有小鼠的t细胞谱。
活的和灭活的流感免疫产生不同的免疫特征。
活的和杀死的pr8疫苗同样地针对攻击提供保护。图15的a图中针对全病毒的elisa表明活的和灭活的流感疫苗产生不同的抗体应答强度。由图16的a图中的散点图可以看出每个疫苗组相对于未接种组(naive)所识别的肽的差异。通过使用fdr校正的p<0.05和倍数变化>1.3倍的选择标准,由活流感产生的血清所识别的肽的数目为灭活疫苗血清的10.75倍。
两种疫苗有7种肽被共同地识别,人们将期望在相似大小的列表之间偶然识别少于一种肽(图16,b图)。主成分分析(pca)图使用方差作为x和y标量值显示了组间相对差异。任一组小鼠所识别的全部593种肽明显地将活疫苗免疫的动物与灭活疫苗免疫的动物区分开(图16,c图)。当被要求预测类别时,支持向量机(svm)显示出0%的弃一法交叉验证(loocv)误差。对重叠肽的分析显示接种活疫苗和灭活疫苗的小鼠聚类在一起,并且与模拟免疫的小鼠区分开(图16,d图)。活疫苗免疫特征中更高数目的肽可能是由于病毒复制的剂量放大效应或不存在于灭活并可能分解的病毒体中的其他表位。这些数据综合起来表明这两种保护性疫苗具有非常不同的免疫特征。
免疫特征分析可区分紧密相关的疫苗
为了评估免疫特征分析进行精细标度概况分析(profiling)的能力,用来自采用灭活的季节性流感疫苗免疫的小鼠的血清探查随机肽阵列。使用两个版本的10k阵列:在cim10k版本1(cim10kv1)上检测合并的样品,而在cim10k版本3(cim10kv3)上检测单个样品。cim10kv3相对于cim10k包含大量的技术改进,并且选择cim10kv3来评估单个小鼠。
图16表明免疫特征可区分较弱的灭活疫苗与较强的灭活疫苗。首先以anova在两种季节性疫苗与杀死的pr8疫苗之间比较其在cim10kv3.0上的免疫特征,其中p<0.0005的55种肽(5个假阳性)能够区分三种疫苗。(图16,a图)中的第一和第二主分量的图中示出了个体间的差异。第一比较使用单因素anova在p<0.0005下寻找与三种疫苗中的总平均值不同的肽。该比较产生55种能够在svm中以0%的loocv误差区分这三种疫苗的肽(图16,a图)。
第二比较使用studentt检验将每种疫苗单独地与模拟免疫的小鼠进行比较。与模拟相比显著改变的肽的数目在疫苗之间不同。与模拟相比的肽之间的重叠示于图16的b图的维恩图中。使用合并血清在cim10kv1.0上进行接种疫苗的小鼠与未接种小鼠之间的比较,其中相对于模拟免疫的小鼠,来自免疫的小鼠的血清中的归一化荧光单位具有最少1.3倍的增加,且使用benjamini和hochberg多重检验校正(multipletestcorrection)时p值小于0.05。这些所列项之间的重叠示于(图16,b图)的维恩图中。
与杀死的pr8相比,在两种季节性疫苗之间观察到更多的重叠。这两种季节性疫苗制剂在所包含的h1n1株上不同。该模式在cim10kv1和cim10kv3两种微阵列上是一致的。这表明免疫特征对于检测疫苗组合物的细微差异是足够敏感的。
已知的保护性应答的免疫特征可预测攻击后的结果
在此,我们表明已知的保护性应答的免疫特征可预测疫苗的功效。sam算法使用置换t检验(per-mutatedt-test),并用于以一种肽的假阳性率(1/25,或fp=4%)选择25种能够区分活疫苗免疫和模拟免疫的肽作为训练集。这25种肽包括在活的与杀死的pr8免疫特征之间的重叠肽。为了克服肽的不同亲和力的影响,我们使用二元分类器,其基于是否已经达到一定的截止得分来对阵列特征进行分箱(bin)。使用这些二元得分来计算成对汉明距离(pairwisehammingdistance)的组平均值,作为表7中示出的免疫特征之间的二元差异数。表7的说明:a使用sam针对每种能够区分活疫苗免疫的小鼠与模拟免疫的小鼠的肽计算个体间的比值的log2;b在活疫苗免疫的小鼠中基于10%肽的截止值将比值分箱为1或0;c使用二元得分计算汉明距离;d平均汉明距离±标准差。e在统计学上不同于季节性疫苗和活疫苗,双尾t检验p=6.5×10-5;f在统计学上不同于季节性疫苗和模拟疫苗,双尾t检验p=1.39×10-6。
使用季节性疫苗作为对相同的25种肽的测试集。发现用杀死的pr8免疫的小鼠更接近于用活疫苗免疫的小鼠,并且与接受季节性疫苗的小鼠相比与模拟免疫的小鼠的差别更大。这与以下事实相符:灭活的pr8疫苗是赋予完全无症状保护的疫苗,而季节性疫苗仅提供部分保护。将杀死的pr8作为具有最大保护性的疫苗的基于免疫特征的预测反映了相对elisa滴度和细胞因子表达模式。假如免疫特征为唯一使用的测定,则其已挑选出正确的疫苗。该数据表明了免疫特征通过选择具有最大保护功效的疫苗而有助于疫苗开发的能力。
季节性疫苗接受者具有与pr8攻击之后的结果相关联的不同的免疫特征
用季节性疫苗免疫的小鼠部分地针对pr8株的攻击提供保护。来自季节性疫苗接受者的脾细胞均不分泌il-12或ifn-γ,提示在缺乏强cd8应答的情况下传送部分保护。
图18表明免疫特征可以预测:抗体与na195-219的交叉反应性在保护季节性疫苗接受者免受pr8影响方面是重要的。在pr8攻击后存活或死亡的2006-2007疫苗接受者的全病毒elisa终点滴度在(图18,a图)中示出,其中水平线代表组平均值,而每个点代表单个小鼠。(图18,a图)未指明组内的解释性趋势。
对于两年,在攻击后存活或死于攻击的季节性疫苗接受者之间比较免疫特征。通过使用截止值为p<0.005的双尾t检验,94种肽被鉴定为显著不同,并且使用svm能够达到100%loocv准确度。在pca图中,存活的小鼠与用杀死的pr8免疫的小鼠聚类在一起(图18,b图)。接受灭活疫苗的所有个体之间的差异示于(图18,b图)中,作为第一和第二主分量的图。
使用38种肽(比被死亡者识别的肽少至少1.3倍)来预测神经氨酸酶中的表位(图18,c图),其中guitope得分在y-轴上,而氨基酸在x-轴上。来自合并血清的抗体与最强预测表位的反应性作为重复阵列的平均值+/-标准差绘制于(图18,d图)中。在这94种肽的选择中不包括pr8免疫的小鼠。这提示免疫特征可用于预测疫苗接种者在感染后的个体结果。
免疫特征可经生物信息学方式被追溯至a/pr/8/34蛋白质上的表位
以上所述的实验表明免疫特征可以产生保护的替代物。如果这些替代物也可指示保护的关联物,例如病毒中的实际保护性表位,则将是有用的。
为了评估被灭活疫苗识别的表位的宽度,我们将我们对肽的选择缩减至在免疫后具有增强的信号的那些肽。针对pr8蛋白质序列搜索与通过用杀死的pr8免疫而产生的抗体相结合的肽,并且ha和na的结果绘图于图19中。图19说明了可解读随机肽阵列上的免疫特征以确定被每种疫苗识别的表位。使用确定为相对于未接种的小鼠在接种疫苗的小鼠中越来越多地被识别的肽来对pr8血凝素(图19,a图)和神经氨酸酶(图19,b图)的蛋白质序列进行guitope搜索。通过在cim10kv1.0上使用合并的血清比较接种疫苗的小鼠与未接种的小鼠来选择肽,其中相对于模拟免疫的小鼠,免疫的小鼠中最少增加1.3倍,并且fdr调节的p值小于0.05。对于两种蛋白质,线型图代表每种氨基酸的guitope预测得分。对实际表位序列的识别在每个线型图上方的方框中示出。方框代表由含有表位序列的平铺肽所覆盖的氨基酸(x-轴)。方框颜色代表来自每种疫苗的合并血清的抗体与这些肽的结合强度。所使用的色标在每幅图的右上角示出。
由于流感株之间有接近的同源性,预测有共同的表位,但用杀死的pr8接种的小鼠识别独特的序列。为了检验这些预测,使用免疫的小鼠的合并血清来探查含有大多数pr8ha和na序列的平铺肽阵列。根据在平铺肽阵列上存在的序列,所有预测均得到支持,但并非所有表位均被预测到。这些表位从预测列表中的排除可能是由于用来选择肽的严格的错误发现校正。如果取消错误发现校正,则在guitope预测与表位阵列之间具有一致性的表位的百分比增加。有趣的是,预测只有用杀死的pr8免疫的小鼠结合na上的表位,包括203-swrkkilrtqes-209(swrkkilrtqes:seqidno.4),其在2009ah1n1病毒中的同源物为已知的中和表位。并非所有包括在免疫特征中的肽均通过guitope预测为与实际序列一致。这些肽可被针对构象表位的抗体所识别。
这些结果表明,免疫特征分析可准确地检测针对生物免疫原的线性表位而内源性产生的抗体。
实施例6:发现及破译能够引发抗原应答的肽
图20为本发明在靶标鉴定筛选中的一种应用的总示意图。
使来自患有感兴趣的病症的个体的复杂生物样品例如血清与肽阵列接触;检测血清中的抗体与阵列上的肽的结合以产生疾病免疫谱;将个体的疾病免疫谱与正常对照进行比较,并基于以下一项或两项来鉴定差异结合的肽:(i)与正常对照相比在疾病免疫谱中结合更多抗体的肽;和(ii)与正常对照相比在疾病免疫谱中结合更少抗体的肽;其中与差异结合的肽相对应的蛋白质为针对感兴趣的疾病的治疗靶标。
本领域技术人员将会理解,本文所述的多种阵列可与这样的方法一起使用。本领域技术人员还将理解,多种检测方法可与此类方法结合使用。本领域技术人员还将理解,采用本文所述的方法可以发现并破译能够引发针对多种不同状况的抗原应答的多种肽。在一些实施方案中,与差异结合的肽相对应的蛋白质为感兴趣的疾病的治疗靶标。
实施方案
以下非限制性实施方案提供了本发明的示例性实例,但并不限制本发明的范围。
实施方案1.在一些实施方案中,本发明提供了一种筛选治疗靶标的方法,该方法包括:a)使肽阵列与来自具有感兴趣的已知状况的个体的第一生物样品接触;b)检测所述第一生物样品中的抗体与所述肽阵列的结合以获得第一免疫特征谱;c)使肽阵列与来源于没有已知状况的个体的对照样品接触;d)检测所述对照样品中的抗体与所述肽阵列的结合以获得第二免疫特征谱;e)将所述第一免疫特征谱与所述第二免疫特征谱进行比较,并鉴定与第二免疫特征谱相比在第一免疫特征谱中结合更少或更多抗体的差异结合的肽;以及f)将与所鉴定的差异结合的肽相对应的蛋白质鉴定为针对感兴趣的状况的治疗靶标。
实施方案2.实施方案1的方法,其中所述蛋白质未经注释。
实施方案3.实施方案1的方法,其中所述蛋白质具有移码。
实施方案4.实施方案1-3中任一个的方法,其中所述治疗靶标为所述蛋白质的表位。
实施方案5.实施方案1-4中任一个的方法,其中所述肽阵列包含至少10,000个不同的肽。
实施方案6.实施方案1-5中任一个的方法,其中所述肽阵列包含至少100,000个不同的肽。
实施方案7.实施方案1-6中任一个的方法,其中所述肽阵列包含至少330,000个不同的肽。
实施方案8.实施方案1-7中任一个的方法,其中所述肽阵列包含至少500,000个不同的肽。
实施方案9.实施方案5-8中任一个的方法,其中所述肽阵列上的不同的肽为8至35个残基的长度。
实施方案10.实施方案5-9中任一个的方法,其中所述肽阵列上的不同的肽具有范围为2-4nm的平均间距。
实施方案11.实施方案5-10中任一个的方法,其中所述肽阵列上的不同的肽具有范围为3-6nm的平均间距。
实施方案12.实施方案5-11中任一个的方法,其中所述不同的肽包含肽模拟物。
实施方案13.实施方案5-12中任一个的方法,其中所述不同的肽具有随机氨基酸序列。
实施方案14.实施方案5-13中任一个的方法,其中所述不同的肽包含非天然氨基酸。
实施方案15.实施方案1-14中任一个的方法,其进一步包括开发针对所述治疗靶标的疫苗。
实施方案16.一种鉴定疫苗靶标的方法,该方法包括:a)使肽阵列与来自具有感兴趣的已知状况的个体的第一生物样品接触;b)检测所述第一生物样品中的抗体与所述肽阵列的结合以获得第一免疫特征谱;c)使肽阵列与来源于没有已知状况的个体的对照样品接触;d)检测所述对照样品中的抗体与所述肽阵列的结合以获得第二免疫特征谱;e)将所述第一免疫特征谱与所述第二免疫特征谱进行比较,并鉴定与第二免疫特征谱相比在第一免疫特征谱中结合更少或更多抗体的差异结合的肽;以及f)将与所鉴定的差异结合的肽相对应的蛋白质鉴定为针对感兴趣的状况的疫苗靶标。
实施方案17.实施方案16的方法,其中所述蛋白质未经注释。
实施方案18.实施方案16的方法,其中所述蛋白质具有移码。
实施方案19.实施方案16-18中任一个的方法,其中所述治疗靶标为所述蛋白质的表位。
实施方案20.实施方案16-19中任一个的方法,其中所述肽阵列包含至少10,000个不同的肽。
实施方案21.实施方案16-20中任一个的方法,其中所述肽阵列包含至少100,000个不同的肽。
实施方案22.实施方案16-21中任一个的方法,其中所述肽阵列包含至少330,000个不同的肽。
实施方案23.实施方案16-22中任一个的方法,其中所述肽阵列包含至少500,000个不同的肽。
实施方案24.实施方案16-23中任一个的方法,其中所述肽阵列上的不同的肽为8至35个残基的长度。
实施方案25.实施方案16-24中任一个的方法,其中所述肽阵列上的不同的肽具有范围为2-4nm的平均间距。
实施方案26.实施方案16-25中任一个的方法,其中所述肽阵列上的不同的肽具有范围为3-6nm的平均间距。
实施方案27.实施方案16-26中任一个的方法,其中所述不同的肽包含肽模拟物。
实施方案28.实施方案16-27中任一个的方法,其中所述不同的肽具有随机氨基酸序列。
实施方案29.实施方案16-28中任一个的方法,其中所述不同的肽包含非天然氨基酸。
实施方案30.实施方案16-29中任一个的方法,其中所述疫苗针对病原体、微生物有机体、癌症或自身免疫病症。
实施方案31.一种鉴定针对癌症的治疗靶标的方法,该方法包括:a)使肽阵列与来自患有感兴趣的已知癌症的个体的第一生物样品接触;b)检测所述第一生物样品中的抗体与所述肽阵列的结合以获得第一免疫特征谱;c)使肽阵列与来源于没有已知癌症的个体的对照样品接触;d)检测所述对照样品中的抗体与所述肽阵列的结合以获得第二免疫特征谱;e)将所述第一免疫特征谱与所述第二免疫特征谱进行比较,并鉴定与第二免疫特征谱相比在第一免疫特征谱中结合更少或更多抗体的差异结合的肽;以及f)将与所鉴定的差异结合的肽相对应的蛋白质鉴定为针对感兴趣的癌症的靶标。
实施方案32.实施方案31的方法,其中所述蛋白质未经注释。
实施方案33.实施方案31的方法,其中所述蛋白质具有移码。
实施方案34.实施方案31-33中任一个的方法,其中所述治疗靶标为所述蛋白质的表位。
实施方案35.实施方案31-34中任一个的方法,其中所述肽阵列包含至少10,000个不同的肽。
实施方案36.实施方案31-35中任一个的方法,其中所述肽阵列包含至少100,000个不同的肽。
实施方案37.实施方案31-36中任一个的方法,其中所述肽阵列包含至少330,000个不同的肽。
实施方案38.实施方案31-37中任一个的方法,其中所述肽阵列包含至少500,000个不同的肽。
实施方案39.实施方案35-38中任一个的方法,其中所述肽阵列上的不同的肽为8至35个残基的长度。
实施方案40.实施方案35-38中任一个的方法,其中所述肽阵列上的不同的肽具有范围为2-4nm的平均间距。
实施方案41.实施方案35-40中任一个的方法,其中所述肽阵列上的不同的肽具有范围为3-6nm的平均间距。
实施方案42.实施方案31-41中任一个的方法,其中所述不同的肽包含肽模拟物。
实施方案43.实施方案31-42中任一个的方法,其中所述不同的肽具有随机氨基酸序列。
实施方案44.实施方案31-43中任一个的方法,其中所述不同的肽包含非天然氨基酸。
实施方案45.实施方案31-44中任一个的方法,其中所述癌症选自肺癌、白血病、胰腺癌、前列腺癌、乳腺癌、膀胱癌、子宫内膜癌和结肠直肠癌。
实施方案46.实施方案31-45中任一个的方法,其中所述癌症为乳腺癌。
实施方案47.一种鉴定针对自身免疫病症的治疗靶标的方法,该方法包括:a)使肽阵列与来自患有感兴趣的已知自身免疫病症的个体的第一生物样品接触;b)检测所述第一生物样品中的抗体与所述肽阵列的结合以获得第一免疫特征谱;c)使肽阵列与来源于没有已知自身免疫病症的个体的对照样品接触;d)检测所述对照样品中的抗体与所述肽阵列的结合以获得第二免疫特征谱;e)将所述第一免疫特征谱与所述第二免疫特征谱进行比较,并鉴定与第二免疫特征谱相比在第一免疫特征谱中结合更少或更多抗体的差异结合的肽;以及f)将与所鉴定的差异结合的肽相对应的蛋白质鉴定为针对感兴趣的自身免疫病症的靶标。
实施方案48.实施方案47的方法,其中所述蛋白质未经注释。
实施方案49.实施方案47的方法,其中所述蛋白质具有移码。
实施方案50.实施方案47-49中任一个的方法,其中所述治疗靶标为所述蛋白质的表位。
实施方案51.实施方案47-50中任一个的方法,其中所述肽阵列包含至少10,000个不同的肽。
实施方案52.实施方案47-51中任一个的方法,其中所述肽阵列包含至少100,000个不同的肽。
实施方案53.实施方案47-52中任一个的方法,其中所述肽阵列包含至少330,000个不同的肽。
实施方案54.实施方案47-53中任一个的方法,其中所述肽阵列包含至少500,000个不同的肽。
实施方案55.实施方案51-54中任一个的方法,其中所述肽阵列上的不同的肽为8至35个残基的长度。
实施方案56.实施方案51-54中任一个的方法,其中所述肽阵列上的不同的肽具有范围为2-4nm的平均间距。
实施方案57.实施方案47-56中任一个的方法,其中所述肽阵列上的不同的肽具有范围为3-6nm的平均间距。
实施方案58.实施方案47-57中任一个的方法,其中所述不同的肽包含肽模拟物。
实施方案59.实施方案47-58中任一个的方法,其中所述不同的肽具有随机氨基酸序列。
实施方案60.实施方案47-59中任一个的方法,其中所述不同的肽包含非天然氨基酸。
实施方案61.实施方案47-60中任一个的方法,其中所述自身免疫病症选自1型糖尿病、类风湿性关节炎、多发性硬化、炎性肠病、系统性红斑狼疮、银屑病和硬皮病。
实施方案62.实施方案47-61中任一个的方法,其中所述自身免疫病症为i型糖尿病。
序列表
<110>亚利桑那州评议委员会,亚利桑那州法人团体,代理和代表亚利桑那州立大学
<120>发现治疗靶标的方法
<130>43638-708.601
<140>pct/us2013/058325
<141>2013-09-05
<150>14/014,168
<151>2013-08-29
<150>61/696,873
<151>2012-09-05
<160>17
<170>patentinversion3.5
<210>1
<211>20
<212>prt
<213>人工序列
<220>
<223>人工序列的描述:合成肽
<400>1
lysserphehisglyargvalileglnaspvalvalglygluprotyr
151015
glyglysercys
20
<210>2
<211>20
<212>prt
<213>人工序列
<220>
<223>人工序列的描述:合成肽
<400>2
proalametlyshisarggluprohistrpvalileproglyileile
151015
trpglysercys
20
<210>3
<211>20
<212>prt
<213>人工序列
<220>
<223>人工序列的描述:合成肽
<400>3
gluprometglumethisaspaspargthrmetargproasnglyala
151015
pheglysercys
20
<210>4
<211>12
<212>prt
<213>甲型流感病毒
<400>4
sertrparglyslysileleuargthrglngluser
1510
<210>5
<211>8
<212>prt
<213>人工序列
<220>
<223>人工序列的描述:合成肽
<400>5
valileglnaspvalvalglyglu
15
<210>6
<211>8
<212>prt
<213>人工序列
<220>
<223>人工序列的描述:合成肽
<220>
<221>mod_res
<222>(6)..(6)
<223>val或leu
<400>6
valileglnaspvalxaaglyglu
15
<210>7
<211>8
<212>prt
<213>小鼠(mussp.)
<400>7
valileaspglnvalleuglyglu
15
<210>8
<211>7
<212>prt
<213>人工序列
<220>
<223>人工序列的描述:合成肽
<400>8
phehisglyargvalilegln
15
<210>9
<211>7
<212>prt
<213>人工序列
<220>
<223>人工序列的描述:合成肽
<220>
<221>mod_res
<222>(6)..(6)
<223>ile或thr
<400>9
phehisglyargvalxaagln
15
<210>10
<211>7
<212>prt
<213>小鼠(mussp.)
<400>10
phehisglyargvalthrgln
15
<210>11
<211>17
<212>prt
<213>人工序列
<220>
<223>人工序列的描述:合成肽
<400>11
gluprometglumethisaspaspargthrmetargproasnglyala
151015
phe
<210>12
<211>57
<212>prt
<213>甲型流感病毒
<400>12
gluglulysargglnleulyslysglyleulysserglngluglncys
151015
alaserleuprothrlysvalserargargthrserproalaleulys
202530
ileleuglupromettrpmetaspserasnargthralathrleuarg
354045
alasercysleulyscysprolyslys
5055
<210>13
<211>17
<212>prt
<213>人工序列
<220>
<223>人工序列的描述:合成肽
<400>13
proalametlyshisarggluprohistrpvalileproglyileile
151015
trp
<210>14
<211>20
<212>prt
<213>人工序列
<220>
<223>人工序列的描述:合成肽
<400>14
gluglulysargglnleulyslysglyleulysserglngluglncys
151015
alaglysercys
20
<210>15
<211>20
<212>prt
<213>人工序列
<220>
<223>人工序列的描述:合成肽
<400>15
glncysalaserleuprothrlysvalserargargthrserproala
151015
leuglysercys
20
<210>16
<211>20
<212>prt
<213>人工序列
<220>
<223>人工序列的描述:合成肽
<400>16
proalaleulysileleuglupromettrpmetaspserasnargthr
151015
alaglysercys
20
<210>17
<211>20
<212>prt
<213>人工序列
<220>
<223>人工序列的描述:合成肽
<400>17
serasnargthralathrleuargalasercysleulyscysprolys
151015
lysglysercys
20
- 还没有人留言评论。精彩留言会获得点赞!