鉴定T细胞受体的方法
鉴定t细胞受体的方法
1.相关申请的交叉引用
2.本pct申请要求于2019年7月30日提交的美国临时申请第62/880,492号和于2020年5月22日提交的美国临时申请第63/029,103号的优先权权益,它们各自通过引用以其整体并入本文。
3.通过efs-web电子提交的序列表的引用
4.该电子提交的序列表(名称:4285.009pc02_sl_st25.txt,大小:291,794字节;创建日期:2020年7月28日)的内容通过引用以其整体并入本文。
发明领域
5.本公开提供了鉴定mhc ii类特异性t细胞受体(“tcr”)的方法。
6.发明背景
7.免疫疗法已成为对抗包括癌症在内的多种疾病的关键工具。t细胞疗法处于免疫治疗发展的前沿,并且抗肿瘤t细胞的过继转移已被证明可诱导癌症患者的临床反应。尽管许多t细胞疗法靶向突变的肿瘤抗原,但绝大多数新抗原不是共享的,并且对每个患者都是独一无二的。
8.潜在的非突变抗原数量多于突变抗原多个数量级。阐明来源于共享抗原的t细胞表位可能有助于有效和安全的过继性t细胞疗法的稳健发展,这些疗法很容易为更大的癌症患者群体所用。然而,非突变抗原的绝对数量和hla基因的高多态性可能阻碍了针对非突变抗原的抗肿瘤t细胞反应的特异性的综合分析。
技术实现要素:
9.本公开的某些方面涉及鉴定mhc ii类特异性t细胞受体(tcr)的方法,所述方法包括使t细胞与包含mhc ii类分子和肽的复合物接触;其中所述t细胞表达cd4和一种或多种tcr;其中所述mhc ii类分子包含α链和β链,其中所述mhc ii类分子对cd4的亲和力高于天然存在的mhc ii类分子对cd4的亲和力;并且其中所述mhc ii类特异性tcr特异性结合包含所述mhc ii类分子和所述肽的复合物。
10.在一些方面,mhc ii类分子的β链包含相对于mhc ii类分子的野生型β链具有一个或多个突变的氨基酸序列。在一些方面,mhc ii类分子的α链包含相对于mhc ii类分子的野生型α链具有一个或多个突变的氨基酸序列。在一些方面,一种或多种突变包括取代突变。
11.在一些方面,mhc ii类分子是hla-dp、hla-dq或hla-dr等位基因,或它们的任何组合。在一些方面,(i)hla ii类分子的β链是hla-dp等位基因,(ii)hla ii类分子的α链是hla-dp等位基因,或(iii)(i)和(ii)两者兼而有之。在一些方面,hla ii类分子的β链是dp1、dp2、dp3、dp4、dp5、dp6、dp8或dp9等位基因。
12.在一些方面,mhc ii类分子的β链包含选自由以下组成的组的hla等位基因:dpb1*01、dpb1*02、dpb1*03、dpb1*04、dpb1*05、dpb1*06、dpb1*08、dpb1*09、dpb1*10、dpb1*100、dpb1*101、dpb1*102、dpb1*103、dpb1*104、dpb1*105、dpb1*106、dpb1*107、dpb1*108、dpb1*
109、dpb1*110、dpb1*111、dpb1*112、dpb1*113、dpb1*114、dpb1*115、dpb1*116、dpb1*117、dpb1*118、dpb1*119、dpb1*11、dpb1*120、dpb1*121、dpb1*122、dpb1*123、dpb1*124、dpb1*125、dpb1*126、dpb1*127、dpb1*128、dpb1*129、dpb1*130、dpb1*131、dpb1*132、dpb1*133、dpb1*134、dpb1*135、dpb1*136、dpb1*137、dpb1*138、dpb1*139、dpb1*13、dpb1*140、dpb1*141、dpb1*142、dpb1*143、dpb1*144、dpb1*145、dpb1*146、dpb1*147、dpb1*148、dpb1*149、dpb1*14、dpb1*150、dpb1*151、dpb1*152、dpb1*153、dpb1*154、dpb1*155、dpb1*156、dpb1*157、dpb1*158、dpb1*159、dpb1*15、dpb1*160、dpb1*161、dpb1*162、dpb1*163、dpb1*164、dpb1*165、dpb1*166、dpb1*167、dpb1*168、dpb1*169、dpb1*16、dpb1*170、dpb1*171、dpb1*172、dpb1*173、dpb1*174、dpb1*175、dpb1*176、dpb1*177、dpb1*178、dpb1*179、dpb1*17、dpb1*180、dpb1*181、dpb1*182、dpb1*183、dpb1*184、dpb1*185、dpb1*186、dpb1*187、dpb1*188、dpb1*189、dpb1*18、dpb1*190、dpb1*191、dpb1*192、dpb1*193、dpb1*194、dpb1*195、dpb1*196、dpb1*197、dpb1*198、dpb1*199、dpb1*19、dpb1*200、dpb1*201、dpb1*202、dpb1*203、dpb1*204、dpb1*205、dpb1*206、dpb1*207、dpb1*208、dpb1*209、dpb1*20、dpb1*210、dpb1*211、dpb1*212、dpb1*213、dpb1*214、dpb1*215、dpb1*216、dpb1*217、dpb1*218、dpb1*219、dpb1*21、dpb1*220、dpb1*221、dpb1*222、dpb1*223、dpb1*224、dpb1*225、dpb1*226、dpb1*227、dpb1*228、dpb1*229、dpb1*22、dpb1*230、dpb1*231、dpb1*232、dpb1*233、dpb1*234、dpb1*235、dpb1*236、dpb1*237、dpb1*238、dpb1*239、dpb1*23、dpb1*240、dpb1*241、dpb1*242、dpb1*243、dpb1*244、dpb1*245、dpb1*246、dpb1*247、dpb1*248、dpb1*249、dpb1*24、dpb1*250、dpb1*251、dpb1*252、dpb1*253、dpb1*254、dpb1*255、dpb1*256、dpb1*257、dpb1*258、dpb1*259、dpb1*25、dpb1*260、dpb1*261、dpb1*262、dpb1*263、dpb1*264、dpb1*265、dpb1*266、dpb1*267、dpb1*268、dpb1*269、dpb1*26、dpb1*270、dpb1*271、dpb1*272、dpb1*273、dpb1*274、dpb1*275、dpb1*276、dpb1*277、dpb1*278、dpb1*279、dpb1*27、dpb1*280、dpb1*281、dpb1*282、dpb1*283、dpb1*284、dpb1*285、dpb1*286、dpb1*287、dpb1*288、dpb1*289、dpb1*28、dpb1*290、dpb1*291、dpb1*292、dpb1*293、dpb1*294、dpb1*295、dpb1*296、dpb1*297、dpb1*298、dpb1*299、dpb1*29、dpb1*300、dpb1*301、dpb1*302、dpb1*303、dpb1*304、dpb1*305、dpb1*306、dpb1*307、dpb1*308、dpb1*309、dpb1*30、dpb1*310、dpb1*311、dpb1*312、dpb1*313、dpb1*314、dpb1*315、dpb1*316、dpb1*317、dpb1*318、dpb1*319、dpb1*31、dpb1*320、dpb1*321、dpb1*322、dpb1*323、dpb1*324、dpb1*325、dpb1*326、dpb1*327、dpb1*328、dpb1*329、dpb1*32、dpb1*330、dpb1*331、dpb1*332、dpb1*333、dpb1*334、dpb1*335、dpb1*336、dpb1*337、dpb1*338、dpb1*339、dpb1*33、dpb1*340、dpb1*341、dpb1*342、dpb1*343、dpb1*344、dpb1*345、dpb1*346、dpb1*347、dpb1*348、dpb1*349、dpb1*34、dpb1*350、dpb1*351、dpb1*352、dpb1*353、dpb1*354、dpb1*355、dpb1*356、dpb1*357、dpb1*358、dpb1*359、dpb1*35、dpb1*360、dpb1*361、dpb1*362、dpb1*363、dpb1*364、dpb1*365、dpb1*366、dpb1*367、dpb1*368、dpb1*369、dpb1*36、dpb1*370、dpb1*371、dpb1*372、dpb1*373、dpb1*374、dpb1*375、dpb1*376、dpb1*377、dpb1*378、dpb1*379、dpb1*37、dpb1*380、dpb1*381、dpb1*382、dpb1*383、dpb1*384、dpb1*385、dpb1*386、dpb1*387、dpb1*388、dpb1*389、dpb1*38、dpb1*390、dpb1*391、dpb1*392、dpb1*393、dpb1*394、dpb1*395、dpb1*396、dpb1*397、dpb1*398、dpb1*399、dpb1*39、dpb1*400、dpb1*401、dpb1*402、dpb1*403、dpb1*404、dpb1*405、dpb1*406、dpb1*407、dpb1*408、dpb1*409、dpb1*40、dpb1*410、dpb1*411、
dpb1*412、dpb1*413、dpb1*414、dpb1*415、dpb1*416、dpb1*417、dpb1*418、dpb1*419、dpb1*41、dpb1*420、dpb1*421、dpb1*422、dpb1*423、dpb1*424、dpb1*425、dpb1*426、dpb1*427、dpb1*428、dpb1*429、dpb1*430、dpb1*431、dpb1*432、dpb1*433、dpb1*434、dpb1*435、dpb1*436、dpb1*437、dpb1*438、dpb1*439、dpb1*440、dpb1*441、dpb1*442、dpb1*443、dpb1*444、dpb1*445、dpb1*446、dpb1*447、dpb1*448、dpb1*449、dpb1*44、dpb1*450、dpb1*451、dpb1*452、dpb1*453、dpb1*454、dpb1*455、dpb1*456、dpb1*457、dpb1*458、dpb1*459、dpb1*45、dpb1*460、dpb1*461、dpb1*462、dpb1*463、dpb1*464、dpb1*465、dpb1*466、dpb1*467、dpb1*468、dpb1*469、dpb1*46、dpb1*470、dpb1*471、dpb1*472、dpb1*473、dpb1*474、dpb1*475、dpb1*476、dpb1*477、dpb1*478、dpb1*479、dpb1*47、dpb1*480、dpb1*481、dpb1*482、dpb1*483、dpb1*484、dpb1*485、dpb1*486、dpb1*487、dpb1*488、dpb1*489、dpb1*48、dpb1*490、dpb1*491、dpb1*492、dpb1*493、dpb1*494、dpb1*495、dpb1*496、dpb1*497、dpb1*498、dpb1*499、dpb1*49、dpb1*500、dpb1*501、dpb1*502、dpb1*503、dpb1*504、dpb1*505、dpb1*506、dpb1*507、dpb1*508、dpb1*509、dpb1*50、dpb1*510、dpb1*511、dpb1*512、dpb1*513、dpb1*514、dpb1*515、dpb1*516、dpb1*517、dpb1*518、dpb1*519、dpb1*51、dpb1*520、dpb1*521、dpb1*522、dpb1*523、dpb1*524、dpb1*525、dpb1*526、dpb1*527、dpb1*528、dpb1*529、dpb1*52、dpb1*530、dpb1*531、dpb1*532、dpb1*533、dpb1*534、dpb1*535、dpb1*536、dpb1*537、dpb1*538、dpb1*539、dpb1*53、dpb1*540、dpb1*541、dpb1*542、dpb1*543、dpb1*544、dpb1*545、dpb1*546、dpb1*547、dpb1*548、dpb1*549、dpb1*54、dpb1*550、dpb1*551、dpb1*552、dpb1*553、dpb1*554、dpb1*555、dpb1*556、dpb1*557、dpb1*558、dpb1*559、dpb1*55、dpb1*560、dpb1*561、dpb1*562、dpb1*563、dpb1*564、dpb1*565、dpb1*566、dpb1*567、dpb1*568、dpb1*569、dpb1*56、dpb1*570、dpb1*571、dpb1*572、dpb1*573、dpb1*574、dpb1*575、dpb1*576、dpb1*577、dpb1*578、dpb1*579、dpb1*57、dpb1*580、dpb1*581、dpb1*582、dpb1*583、dpb1*584、dpb1*585、dpb1*586、dpb1*587、dpb1*588、dpb1*589、dpb1*58、dpb1*590、dpb1*591、dpb1*592、dpb1*593、dpb1*594、dpb1*595、dpb1*596、dpb1*597、dpb1*598、dpb1*599、dpb1*59、dpb1*600、dpb1*601、dpb1*602、dpb1*603、dpb1*604、dpb1*605、dpb1*606、dpb1*607、dpb1*608、dpb1*609、dpb1*60、dpb1*610、dpb1*611、dpb1*612、dpb1*613、dpb1*614、dpb1*615、dpb1*616、dpb1*617、dpb1*618、dpb1*619、dpb1*61、dpb1*620、dpb1*621、dpb1*622、dpb1*623、dpb1*624、dpb1*625、dpb1*626、dpb1*627、dpb1*628、dpb1*629、dpb1*62、dpb1*630、dpb1*631、dpb1*632、dpb1*633、dpb1*634、dpb1*635、dpb1*636、dpb1*637、dpb1*638、dpb1*639、dpb1*63、dpb1*640、dpb1*641、dpb1*642、dpb1*643、dpb1*644、dpb1*645、dpb1*646、dpb1*647、dpb1*648、dpb1*649、dpb1*64、dpb1*650、dpb1*651、dpb1*652、dpb1*653、dpb1*654、dpb1*655、dpb1*656、dpb1*657、dpb1*658、dpb1*659、dpb1*65、dpb1*660、dpb1*661、dpb1*662、dpb1*663、dpb1*664、dpb1*665、dpb1*666、dpb1*667、dpb1*668、dpb1*669、dpb1*66、dpb1*670、dpb1*671、dpb1*672、dpb1*673、dpb1*674、dpb1*675、dpb1*676、dpb1*677、dpb1*678、dpb1*679、dpb1*67、dpb1*680、dpb1*681、dpb1*682、dpb1*683、dpb1*684、dpb1*685、dpb1*686、dpb1*687、dpb1*688、dpb1*689、dpb1*68、dpb1*690、dpb1*691、dpb1*692、dpb1*693、dpb1*694、dpb1*695、dpb1*696、dpb1*697、dpb1*698、dpb1*699、dpb1*69、dpb1*700、dpb1*701、dpb1*702、dpb1*703、dpb1*704、dpb1*705、dpb1*706、dpb1*707、dpb1*708、dpb1*709、dpb1*70、dpb1*710、dpb1*711、dpb1*712、dpb1*713、dpb1*714、dpb1*
715、dpb1*716、dpb1*717、dpb1*718、dpb1*719、dpb1*71、dpb1*720、dpb1*721、dpb1*722、dpb1*723、dpb1*724、dpb1*725、dpb1*726、dpb1*727、dpb1*728、dpb1*729、dpb1*72、dpb1*730、dpb1*731、dpb1*732、dpb1*733、dpb1*734、dpb1*735、dpb1*736、dpb1*737、dpb1*738、dpb1*739、dpb1*73、dpb1*740、dpb1*741、dpb1*742、dpb1*743、dpb1*744、dpb1*745、dpb1*746、dpb1*747、dpb1*748、dpb1*749、dpb1*74、dpb1*750、dpb1*751、dpb1*752、dpb1*753、dpb1*754、dpb1*755、dpb1*756、dpb1*757、dpb1*758、dpb1*759、dpb1*75、dpb1*760、dpb1*761、dpb1*762、dpb1*763、dpb1*764、dpb1*765、dpb1*766、dpb1*767、dpb1*768、dpb1*769、dpb1*76、dpb1*770、dpb1*771、dpb1*772、dpb1*773、dpb1*774、dpb1*775、dpb1*776、dpb1*777、dpb1*778、dpb1*779、dpb1*77、dpb1*780、dpb1*781、dpb1*782、dpb1*783、dpb1*784、dpb1*785、dpb1*786、dpb1*787、dpb1*788、dpb1*789、dpb1*78、dpb1*790、dpb1*791、dpb1*792、dpb1*794、dpb1*795、dpb1*796、dpb1*797、dpb1*798、dpb1*799、dpb1*79、dpb1*800、dpb1*801、dpb1*802、dpb1*803、dpb1*804、dpb1*805、dpb1*806、dpb1*807、dpb1*808、dpb1*809、dpb1*80、dpb1*810、dpb1*811、dpb1*812、dpb1*813、dpb1*814、dpb1*815、dpb1*816、dpb1*817、dpb1*818、dpb1*819、dpb1*81、dpb1*820、dpb1*821、dpb1*822、dpb1*823、dpb1*824、dpb1*825、dpb1*826、dpb1*827、dpb1*828、dpb1*829、dpb1*82、dpb1*830、dpb1*831、dpb1*832、dpb1*833、dpb1*834、dpb1*835、dpb1*836、dpb1*837、dpb1*838、dpb1*839、dpb1*83、dpb1*840、dpb1*841、dpb1*842、dpb1*843、dpb1*844、dpb1*845、dpb1*846、dpb1*847、dpb1*848、dpb1*849、dpb1*84、dpb1*850、dpb1*851、dpb1*852、dpb1*853、dpb1*854、dpb1*855、dpb1*856、dpb1*857、dpb1*858、dpb1*859、dpb1*85、dpb1*860、dpb1*861、dpb1*862、dpb1*863、dpb1*864、dpb1*865、dpb1*866、dpb1*867、dpb1*868、dpb1*869、dpb1*86、dpb1*870、dpb1*871、dpb1*872、dpb1*873、dpb1*874、dpb1*875、dpb1*876、dpb1*877、dpb1*878、dpb1*879、dpb1*87、dpb1*880、dpb1*881、dpb1*882、dpb1*883、dpb1*884、dpb1*885、dpb1*886、dpb1*887、dpb1*888、dpb1*889、dpb1*88、dpb1*890、dpb1*891、dpb1*892、dpb1*893、dpb1*894、dpb1*895、dpb1*896、dpb1*897、dpb1*898、dpb1*899、dpb1*89、dpb1*900、dpb1*901、dpb1*902、dpb1*903、dpb1*904、dpb1*905、dpb1*906、dpb1*907、dpb1*908、dpb1*909、dpb1*90、dpb1*910、dpb1*911、dpb1*912、dpb1*913、dpb1*914、dpb1*915、dpb1*916、dpb1*917、dpb1*918、dpb1*919、dpb1*91、dpb1*920、dpb1*921、dpb1*922、dpb1*923、dpb1*924、dpb1*925、dpb1*926、dpb1*927、dpb1*928、dpb1*929、dpb1*92、dpb1*930、dpb1*931、dpb1*932、dpb1*933、dpb1*934、dpb1*935、dpb1*936、dpb1*937、dpb1*938、dpb1*939、dpb1*93、dpb1*940、dpb1*941、dpb1*942、dpb1*943、dpb1*944、dpb1*945、dpb1*946、dpb1*947、dpb1*948、dpb1*949、dpb1*94、dpb1*950、dpb1*951、dpb1*952、dpb1*953、dpb1*954、dpb1*955、dpb1*956、dpb1*957、dpb1*958、dpb1*959、dpb1*95、dpb1*960、dpb1*961、dpb1*962、dpb1*963、dpb1*964、dpb1*965、dpb1*96、dpb1*97、dpb1*98、和dpb1*99等位基因。
13.在一些方面,mhc ii类分子的α链包含hla-dpa1*01、hla-dpa1*02、hla-dpa1*03或hla-dpa1*04等位基因。
14.在一些方面,mhc ii类分子的β链包含在对应于seq id no:1的氨基酸残基112的位置处的除亮氨酸之外的氨基酸。在一些方面,mhc ii类分子的β链包含在对应于seq id no:1的氨基酸残基141的位置处的除缬氨酸之外的氨基酸。
15.本公开的某些方面涉及鉴定mhc ii类特异性t细胞受体(tcr)的方法,所述方法包
括使t细胞与包含mhc ii类分子和肽的复合物接触;其中所述t细胞表达cd4和一种或多种tcr;其中所述mhc ii类分子包含α链和β链,其中所述mhc ii类分子的β链包含(i)在对应于seq id no:1的氨基酸残基112的位置处的除亮氨酸之外的氨基酸,(ii)在对应于seq id no:1的氨基酸残基141的位置处的除缬氨酸之外的氨基酸,或(iii)(i)和(ii)两者兼而有之;并且其中所述mhc ii类特异性tcr特异性结合包含所述mhc ii类分子和所述肽的复合物。
16.在一些方面,mhc ii类分子对cd4的亲和力高于天然存在的mhc ii类分子对cd4的亲和力。
17.在一些方面,在对应于seq id no:1的氨基酸残基112的位置处的除亮氨酸之外的氨基酸包含疏水侧链。在一些方面,在对应于seq id no:1的氨基酸残基112的位置处的除亮氨酸之外的氨基酸选自由以下组成的组:丙氨酸、缬氨酸、异亮氨酸、甲硫氨酸、苯丙氨酸、酪氨酸、和色氨酸。在一些方面,在对应于seq id no:1的氨基酸残基112的位置处的除亮氨酸之外的氨基酸是色氨酸。
18.在一些方面,在对应于seq id no:1的氨基酸残基141的位置处的除缬氨酸之外的氨基酸包含疏水侧链。在一些方面,在对应于seq id no:1的氨基酸残基141的位置处的除缬氨酸之外的氨基酸选自丙氨酸、异亮氨酸、亮氨酸、甲硫氨酸、苯丙氨酸、酪氨酸、和色氨酸。在一些方面,在对应于seq id no:1的氨基酸残基141的位置处的除缬氨酸之外的氨基酸是甲硫氨酸。
19.在一些方面,(i)hla ii类分子的β链是hla-dq等位基因,(ii)hla ii类分子的α链是hla-dq等位基因,或(iii)(i)和(ii)两者兼而有之。在一些方面,hla ii类分子的β链包含dq2、dq3、dq4、dq5或dq6等位基因。在一些方面,mhc ii类分子的β链包含hla-dqb1*02、hla-dqb1*03、hla-dqb1*04、hla-dqb1*05或hla-dqb1*06等位基因。在一些方面,mhc ii类分子的α链包含hla-dqa1*01、hla-dqa1*02、hla-dqa1*03、hla-dqa1*04、hla-dqa1*05或hla-dqa1*06等位基因。
20.在一些方面,mhc ii类分子的β链包含(a)在对应于seq id no:11的氨基酸残基114的位置处的除亮氨酸之外的氨基酸;(b)在对应于seq id no:11的氨基酸残基143的位置处的除缬氨酸之外的氨基酸;以及(c)下列中的至少三项:(i)在对应于seq id no:11的氨基酸残基110的位置处的除天冬酰胺之外的氨基酸,(ii)在对应于seq id no:11的氨基酸残基116的位置处的除异亮氨酸之外的氨基酸,(iii)在对应于seq id no:11的氨基酸残基118的位置处的除丝氨酸之外的氨基酸,和(iv)在对应于seq id no:11的氨基酸残基146的位置处的除脯氨酸之外的氨基酸。
21.在一些方面,mhc ii类分子的β链包含(a)在对应于seq id no:11的氨基酸残基114的位置处的除亮氨酸之外的氨基酸;(b)在对应于seq id no:11的氨基酸残基143的位置处的除缬氨酸之外的氨基酸;(c)在对应于seq id no:11的氨基酸残基110的位置处的除天冬酰胺之外的氨基酸;(d)在对应于seq id no:11的氨基酸残基116的位置处的除异亮氨酸之外的氨基酸;(e)在对应于seq id no:11的氨基酸残基118的位置处的除丝氨酸之外的氨基酸;和(f)在对应于seq id no:11的氨基酸残基146的位置处的除脯氨酸之外的氨基酸。
22.在一些方面,在对应于seq id no:11的氨基酸残基114的位置处的除亮氨酸之外
的氨基酸包含疏水侧链。在一些方面,在对应于seq id no:11的氨基酸残基114的位置处的除亮氨酸之外的氨基酸选自由以下组成的组:丙氨酸、缬氨酸、异亮氨酸、甲硫氨酸、苯丙氨酸、酪氨酸、和色氨酸。在一些方面,在对应于seq id no:11的氨基酸残基114的位置处的除亮氨酸之外的氨基酸是色氨酸。
23.在一些方面,在对应于seq id no:11的氨基酸残基143的位置处的除缬氨酸之外的氨基酸包含疏水侧链。在一些方面,在对应于seq id no:11的氨基酸残基143的位置处的除缬氨酸之外的氨基酸选自丙氨酸、异亮氨酸、亮氨酸、甲硫氨酸、苯丙氨酸、酪氨酸、和色氨酸。在一些方面,在对应于seq id no:11的氨基酸残基143的位置处的除缬氨酸之外的氨基酸是甲硫氨酸。
24.在一些方面,mhc ii类分子的β链包含在对应于seq id no:11的氨基酸残基110的位置处的除天冬酰胺之外的氨基酸。在一些方面,在对应于seq id no:11的氨基酸残基110的位置处的除天冬酰胺之外的氨基酸选自丝氨酸、苏氨酸和谷氨酰胺。在一些方面,在对应于seq id no:11的氨基酸残基110的位置处的除天冬酰胺之外的氨基酸是谷氨酰胺。
25.在一些方面,mhc ii类分子的β链包含在对应于seq id no:11的氨基酸残基116的位置处的除异亮氨酸之外的氨基酸。在一些方面,在对应于seq id no:11的氨基酸残基116的位置处的除异亮氨酸之外的氨基酸选自丙氨酸、缬氨酸、亮氨酸、甲硫氨酸、苯丙氨酸、酪氨酸、和色氨酸。在一些方面,在对应于seq id no:11的氨基酸残基116的位置处的除异亮氨酸之外的氨基酸是缬氨酸。
26.在一些方面,mhc ii类分子的β链包含在对应于seq id no:11的氨基酸残基118的位置处的除丝氨酸之外的氨基酸。在一些方面,在对应于seq id no:11的氨基酸残基118的位置处的除丝氨酸之外的氨基酸选自精氨酸、组氨酸和赖氨酸。在一些方面,在对应于seq id no:11的氨基酸残基118的位置处的除丝氨酸之外的氨基酸是组氨酸。
27.在一些方面,mhc ii类分子的β链包含在对应于seq id no:11的氨基酸残基146的位置处的除脯氨酸之外的氨基酸。在一些方面,在对应于seq id no:11的氨基酸残基146的位置处的除脯氨酸之外的氨基酸选自丝氨酸、苏氨酸、天冬酰胺和谷氨酰胺。在一些方面,在对应于seq id no:11的氨基酸残基146的位置处的除脯氨酸之外的氨基酸是谷氨酰胺。
28.在一些方面,(i)hla ii类分子的β链是hla-dr等位基因,(ii)hla ii类分子的α链是hla-dr等位基因,或(iii)(i)和(ii)两者兼而有之。
29.在一些方面,hla ii类分子的β链包含dr2、dr3、dr4、dr5、dr6、dr7、dr8、dr9、dr10、dr11、dr12、dr13、dr14、dr15或dr16等位基因。在一些方面,mhc ii类分子的β链包含选自由以下组成的组的hla等位基因:drb1*01、drb1*03、drb1*04、drb1*07、drb1*08、drb1*09、drb1*10、drb1*11、drb1*12、drb1*13、drb1*14、drb1*15、和drb1*16。在一些方面,mhc ii类分子的α链包含hla-dra1*01等位基因。
30.在一些方面,β链包含:(a)在对应于seq id no:19的氨基酸残基114的位置处的除亮氨酸之外的氨基酸;(b)在对应于seq id no:19的氨基酸残基143的位置处的除缬氨酸之外的氨基酸;以及(c)下列中的至少两项:(i)在对应于seq id no:19的氨基酸残基118的位置处的除丝氨酸之外的氨基酸,(ii)在对应于seq id no:19的氨基酸残基139的位置处的除赖氨酸之外的氨基酸,(iii)在对应于seq id no:19的氨基酸残基146的位置处的除甘氨酸之外的氨基酸,(iv)在对应于seq id no:19的氨基酸残基157的位置处的除苏氨酸之外
的氨基酸,(v)在对应于seq id no:19的氨基酸残基163的位置处的除苏氨酸之外的氨基酸,和(vi)在对应于seq id no:19的氨基酸残基164的位置处的除缬氨酸之外的氨基酸。
31.在一些方面,所述β链包含:(c)下列中的至少三项:(i)在对应于seq id no:19的氨基酸残基118的位置处的除亮氨酸之外的氨基酸,(ii)在对应于seq id no:19的氨基酸残基139的位置处的除赖氨酸之外的氨基酸;(iii)在对应于seq id no:19的氨基酸残基146的位置处的除甘氨酸之外的氨基酸;(iv)在对应于seq id no:19的氨基酸残基157的位置处的除苏氨酸之外的氨基酸;(v)在对应于seq id no:19的氨基酸残基163的位置处的除苏氨酸之外的氨基酸;和(vi)在对应于seq id no:19的氨基酸残基164的位置处的除缬氨酸之外的氨基酸。
32.在一些方面,所述β链包含:(c)下列中的至少四项:(i)在对应于seq id no:19的氨基酸残基118的位置处的除亮氨酸之外的氨基酸,(ii)在对应于seq id no:19的氨基酸残基139的位置处的除赖氨酸之外的氨基酸;(iii)在对应于seq id no:19的氨基酸残基146的位置处的除甘氨酸之外的氨基酸;(iv)在对应于seq id no:19的氨基酸残基157的位置处的除苏氨酸之外的氨基酸;(v)在对应于seq id no:19的氨基酸残基163的位置处的除苏氨酸之外的氨基酸;和(vi)在对应于seq id no:19的氨基酸残基164的位置处的除缬氨酸之外的氨基酸。
33.在一些方面,所述β链包含:(a)在对应于seq id no:19的氨基酸残基114的位置处的除亮氨酸之外的氨基酸,(b)在对应于seq id no:19的氨基酸残基143的位置处的除缬氨酸之外的氨基酸,(c)在对应于seq id no:19的氨基酸残基118的位置处的除丝氨酸之外的氨基酸,和(d)在对应于seq id no:19的氨基酸残基157的位置处的除苏氨酸之外的氨基酸。
34.在一些方面,β链包含:(a)在对应于seq id no:19的氨基酸残基114的位置处的除亮氨酸之外的氨基酸,(b)在对应于seq id no:19的氨基酸残基143的位置处的除缬氨酸之外的氨基酸,(c)在对应于seq id no:19的氨基酸残基118的位置处的除丝氨酸之外的氨基酸,(d)在对应于seq id no:19的氨基酸残基139的位置处的除赖氨酸之外的氨基酸,(e)在对应于seq id no:19的氨基酸残基146的位置处的除甘氨酸之外的氨基酸,(f)在对应于seq id no:19的氨基酸残基157的位置处的除苏氨酸之外的氨基酸,(g)在对应于seq id no:19的氨基酸残基163的位置处的除苏氨酸之外的氨基酸,和(h)在对应于seq id no:19的氨基酸残基164的位置处的除缬氨酸之外的氨基酸。
35.在一些方面,在对应于seq id no:19的氨基酸残基114的位置处的除亮氨酸之外的氨基酸包含疏水侧链。在一些方面,在对应于seq id no:19的氨基酸残基114的位置处的除亮氨酸之外的氨基酸选自由以下组成的组:丙氨酸、缬氨酸、异亮氨酸、甲硫氨酸、苯丙氨酸、酪氨酸、和色氨酸。在一些方面,在对应于seq id no:19的氨基酸残基114的位置处的除亮氨酸之外的氨基酸是色氨酸。
36.在一些方面,在对应于seq id no:19的氨基酸残基143的位置处的除缬氨酸之外的氨基酸包含疏水侧链。在一些方面,在对应于seq id no:19的氨基酸残基143的位置处的除缬氨酸之外的氨基酸选自丙氨酸、异亮氨酸、亮氨酸、甲硫氨酸、苯丙氨酸、酪氨酸、和色氨酸。在一些方面,在对应于seq id no:19的氨基酸残基143的位置处的除缬氨酸之外的氨基酸是甲硫氨酸。
37.在一些方面,mhc ii类分子的β链包含在对应于seq id no:19的氨基酸残基118的位置处的除丝氨酸之外的氨基酸。在一些方面,在对应于seq id no:19的氨基酸残基118的位置处的除丝氨酸之外的氨基酸选自精氨酸、组氨酸和赖氨酸。在一些方面,在对应于seq id no:19的氨基酸残基118的位置处的除丝氨酸之外的氨基酸是组氨酸。
38.在一些方面,mhc ii类分子的β链包含在对应于seq id no:19的氨基酸残基139的位置处的除赖氨酸之外的氨基酸。在一些方面,在对应于seq id no:19的氨基酸残基139的位置处的除赖氨酸之外的氨基酸选自丝氨酸、苏氨酸和谷氨酰胺。在一些方面,在对应于seq id no:19的氨基酸残基139的位置处的除赖氨酸之外的氨基酸是苏氨酸。
39.在一些方面,mhc ii类分子的β链包含在对应于seq id no:19的氨基酸残基146的位置处的除甘氨酸之外的氨基酸。在一些方面,在对应于seq id no:19的氨基酸残基146的位置处的除甘氨酸之外的氨基酸选自丝氨酸、天冬酰胺、苏氨酸和谷氨酰胺。在一些方面,在对应于seq id no:19的氨基酸残基146的位置处的除甘氨酸之外的氨基酸是谷氨酰胺。
40.在一些方面,mhc ii类分子的β链包含在对应于seq id no:19的氨基酸残基157的位置处的除苏氨酸之外的氨基酸。在一些方面,在对应于seq id no:19的氨基酸残基157的位置处的除苏氨酸之外的氨基酸选自丙氨酸、缬氨酸、异亮氨酸、亮氨酸、甲硫氨酸、苯丙氨酸、酪氨酸、和色氨酸。在一些方面,在对应于seq id no:19的氨基酸残基157的位置处的除苏氨酸之外的氨基酸是异亮氨酸。
41.在一些方面,mhc ii类分子的β链包含在对应于seq id no:19的氨基酸残基163的位置处的除苏氨酸之外的氨基酸。在一些方面,在对应于seq id no:19的氨基酸残基163的位置处的除苏氨酸之外的氨基酸选自丙氨酸、缬氨酸、异亮氨酸、亮氨酸、甲硫氨酸、苯丙氨酸、酪氨酸、和色氨酸。在一些方面,在对应于seq id no:19的氨基酸残基163的位置处的除苏氨酸之外的氨基酸是甲硫氨酸。
42.在一些方面,mhc ii类分子的β链包含在对应于seq id no:19的氨基酸残基164的位置处的除缬氨酸之外的氨基酸。在一些方面,在对应于seq id no:19的氨基酸残基164的位置处的除缬氨酸之外的氨基酸选自丝氨酸、天冬酰胺、苏氨酸和谷氨酰胺。在一些方面,在对应于seq id no:19的氨基酸残基164的位置处的除缬氨酸之外的氨基酸是苏氨酸。
43.在一些方面,所述β链包含:(a)在对应于seq id no:19的氨基酸残基114的位置处的色氨酸,(b)在对应于seq id no:19的氨基酸残基143的位置处的甲硫氨酸,(c)在对应于seq id no:19的氨基酸残基118的位置处的组氨酸,和(d)在对应于seq id no:19的氨基酸残基157的位置处的异亮氨酸。
44.在一些方面,天然存在的mhc ii类分子包含:(a)在对应于seq id no:1的氨基酸残基112或seq id no:11或19的氨基酸残基114的位置处的亮氨酸,(b)在对应于seq id no:1的氨基酸残基141或seq id no:11或19的氨基酸残基143的位置处的缬氨酸,或(c)(a)和(b)两者兼而有之。
45.在一些方面,天然存在的mhc ii类分子包含:(a)在对应于seq id no:1的氨基酸残基112或seq id no:11或19的氨基酸残基114的位置处的亮氨酸,(b)在对应于seq id no:1的氨基酸残基141或seq id no:11或19的氨基酸残基143的位置处的缬氨酸,(c)在对应于seq id no:11的氨基酸残基110的位置处的天冬酰胺,(d)在对应于seq id no:11的氨基酸残基116的位置处的异亮氨酸;(e)在对应于seq id no:11或19的氨基酸残基118的位
置处的丝氨酸;以及(f)在对应于seq id no:11的氨基酸残基146的位置处的脯氨酸,(g)在对应于seq id no:19的氨基酸残基139的位置处的赖氨酸,(h)在对应于seq id no:19的氨基酸残基146的位置处的甘氨酸,(i)在对应于seq id no:19的氨基酸残基157的位置处的苏氨酸,(j)在对应于seq id no:19的氨基酸残基163的位置处的苏氨酸,(k)在对应于seq id no:19的氨基酸残基164的位置处的缬氨酸,或(l)(a)至(k)的任何组合。
46.在一些方面,mhc ii类分子是二聚体。在一些方面,mhc ii类分子是三聚体。在一些方面,mhc ii类分子是四聚体。在一些方面,肽包含蛋白质的片段。在一些方面,蛋白质由患病细胞表达。在一些方面,蛋白质由肿瘤细胞表达。
47.在一些方面,肽包含至少约10个氨基酸。在一些方面,肽包含约10至约100个氨基酸、约10至约90个氨基酸、约10至约80个氨基酸、约10至约70个氨基酸、约10至约60个氨基酸、约10至约50个氨基酸、约10至约40个氨基酸、约10至约30个氨基酸、约10至约25个氨基酸、约10至约20个氨基酸、约10至约15个氨基酸、约15至约100个氨基酸、20至约100个氨基酸、25至约100个氨基酸、30至约100个氨基酸、35至约100个氨基酸、40至约100个氨基酸、50至约100个氨基酸、60至约100个氨基酸、70至约100个氨基酸、80至约100个氨基酸或90至约100个氨基酸。
48.在一些方面,肽包含约10个氨基酸、约11个氨基酸、约12个氨基酸、约13个氨基酸、约14个氨基酸、约15个氨基酸、约16个氨基酸、约17个氨基酸、约18个氨基酸、约19个氨基酸、约20个氨基酸、约25个氨基酸、约30个氨基酸、约35个氨基酸、约40个氨基酸、约45个氨基酸、约50个氨基酸、约55个氨基酸、约60个氨基酸、约65个氨基酸、约70个氨基酸、约75个氨基酸、约80个氨基酸、约85个氨基酸、约90个氨基酸、约95个氨基酸或约100个氨基酸。
49.在一些方面,mhc ii类分子在抗原呈递细胞的表面上表达。
50.在一些方面,t细胞获自人受试者。在一些方面,t细胞是肿瘤浸润淋巴细胞(til)。
51.在一些方面,mhc ii类分子对cd4的亲和力是天然存在的mhc ii类分子对cd4的结合亲和力的至少约2倍、至少约3倍、至少约4倍、至少约5倍、至少约6倍、至少约7倍、至少约8倍、至少约9倍、至少约10倍、至少约15倍、至少约20倍、至少约25倍、至少约30倍、至少约35倍、至少约40倍、至少约45倍、至少约50倍、至少约60倍、至少约70倍、至少约80倍、至少约90倍、至少约100倍、至少约200倍、至少约300倍、至少约400倍、至少约500倍、或至少约100倍。
52.在一些方面,所述方法进一步包括选择由mhc ii类分子结合的t细胞。在一些方面,所述方法进一步包括分离与mhc ii类分子结合的tcr。在一些方面,所述方法进一步包括对tcr进行测序。在一些方面,所述方法进一步包括克隆tcr。在一些方面,所述方法进一步包括在宿主细胞中重组表达tcr。
53.在一些方面,mhc ii类分子以小于约100μm、小于约50μm、小于约20μm或小于约10μm的kd结合cd4。在一些方面,mhc ii类分子以约14μm或更小的kd结合cd4。在一些方面,mhc ii类分子以约8.9μm或更小的kd结合cd4。
附图说明
54.图1a至图1v是说明亲和力成熟的dp4
l112w/v141m
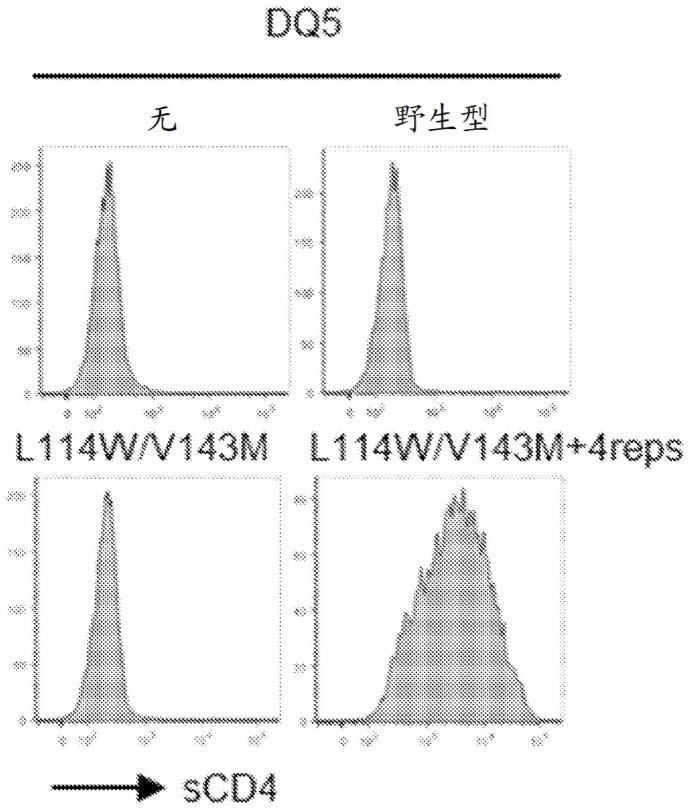
分子展现出增强的cd4结合能力的数据的图形表示。图1a至图1f是直方图,显示了用空白、野生型或含有l112w、v114m、v141m和m158i取代(dp4
l112w/v114m/v141m/m158i
)的突变型dpβ链(dpb1*04:01)转导并用抗ii类mab和可溶
性cd4(scd4)染色的稳定表达野生型dpα链(dpa1*01:03)的hla ii类-空k562细胞的结果。图1g是总结了所有可能的dp4回复突变体对scd4的结合亲和力(mfi;y轴)的条形图,这些突变体与图1a至图1f类似地表达并用scd4染色。如通过稳态分析所量化的,图1h显示了dp4
l112w/v141m
和cd4之间的亲和力。图1i显示了由用分级浓度的dp4/wt1肽冲击的表达野生型dp4或dp4
l112w/v141m
的aapc刺激的dp4/wt1 tcr克隆9转导的jurkat76和jurkat 76/cd4细胞的il-2 epispot测定的结果。图1j至图1w是表示用抗ii类mab和scd4染色的表达dp
l112w/v141m
等位基因(如所示)的k562细胞的直方图。开放直方图代表同型对照染色。*p《0.05,通过学生t检验得到。条形和误差条代表一式三份实验结果的平均值
±
sd。进行至少2次独立实验。图1x至图1aa是直方图,显示了k562细胞表面上的用指示的抗hla ii类抗体检测到的野生型dp4和dp4l112w/v141m分子。没有ii类表达的对照细胞的染色显示为实心灰色。图1ab至图1bh是直方图,显示了指示浓度的表达指示dp4的aapc或用scd4染色的ii类亲本细胞。图1bi显示了指示浓度的表达野生型dp4或dp4
l112w/v141m
的aapc的定量。误差条代表一式三份进行的实验的平均值
±
标准偏差。图1bj是生物层干涉传感图,显示了在一定浓度范围内的生物素化的野生型dp4(配体)与scd4(分析物)的相互作用。图1bk是生物层干涉传感图,显示了在一定浓度范围内的生物素化的dp4
l112w/v141m
(配体)与scd4(分析物)的相互作用。图1bj和1bi中的实验平行进行。所有数据代表两次独立实验。
55.图2a至图2d是dp4
l112w/v141m
和人cd4复合物的模型结构的带状图。图2a至图2b是dpa1*01:03、dpb1*04:01和cd4的三元复合物模型结构的两个方向,如所示的。dpb1*04:01-cd4结合界面围上了虚线方框(图2b)。图2c至图2d提供了野生型dp4(图2c)和dp4
l112w/v141m
(图2d)的cd4结合界面的特写视图。相互作用残基的侧链显示为球棍表示(图2c至图2d)。
56.图3a至图3p是说明dp4
l112w/v141m
二聚体染色在人原代cd4
+
t细胞中表达的同源tcr的数据的图形表示。用dp4/mage-a3
243-258
(r12c9;图3e至图3h)、dp4/wt1
328-348
(克隆9;图3i至图3l)或dp4/ny-eso-1
157-170
(5b8;图3m至图3p)tcr转导原代t细胞并用指示的dp4
l112w/v141m
二聚体染色(图3b至图3d、图3f至图3h、图3j至图3l和图3n至图3p)。
57.图4a至图4d是说明了用dp4
l112w/v141m
二聚体和抗vβ22 mab染色的r12c9转导cd4
+
t细胞的复合染色的散点图。注意,r12c9表示vβ22。图4e至图4h是说明了用dp4
l112w/v141m
二聚体和抗ngfr mab双重染色的克隆9转导的cd4
+
t细胞的复合染色的散点图。注意,克隆9和δngfr基因与p2a融合。
58.图5a至图5p是说明了用5μg/ml常规野生型dp4四聚体和dp4
l112w/v141m
二聚体染色的克隆9转导(图5a至图5h)和5b8转导(图5i至图5p)的原代t细胞的复合染色的散点图。进行至少2次独立实验。
59.图6a至图6f是说明了用dp4
l112w/v141m
二聚体进行全面筛选的结果的条形图,其鉴定了一系列新的dp4限制性肿瘤相关抗原。从6个dp4
+
黑素瘤患者中纯化外周cd4
+
t细胞,并用分别以来源于肿瘤相关抗原的196种不同肽冲击的表达dp4的aapc刺激,并用同源dp4
l112w/v141m
二聚体染色。使用具有最高阳性值的30种肽的结果显示在图6a至图6b中。剩余的166种肽的结果显示在图6c至图6f中。设置每个门以使对照二聚体染色显示《0.2%阳性。阳性二聚体染色定义为染色超过对照二聚体染色3个标准偏差,如虚线所示(》0.6%)。
60.图7a至图7l是来自黑素瘤患者的肽特异性cd4
+
t细胞的dp4
l112w/v141m
二聚体染色的图形表示。从6个dp4
+
黑素瘤患者中纯化原代cd4
+
t细胞,并用分别以来源于肿瘤相关抗原的
196种不同肽冲击的表达dp4的aapc刺激,并用同源dp4
l112w/v141m
二聚体染色,如图6a至图6f所示。显示了dp4
l112w/v141m
二聚体染色的示例。*p《0.05,通过学生t检验得到。n.s.,不显著。进行至少2次独立实验。
61.图8a至图8x是数据的图形表示,说明从dp4
l112w/v141m
二聚体阳性细胞中分离并在人tcr缺陷型cd4
+
t细胞中重构的dp4限制性tcr以dp4限制性和抗原特异性方式起作用。03-ccnd1
219-238
(图8a至图8d)、05-hsd17b12
225-244
和09-hsd17b12
225-244
(图8e至图8j)、05-lgsn
296-315
(图8k至图8n)、03-mage-a2
108-127
和06-mage-a2
108-127
(图8o至图8t)和05-muc5ac
4922-4941
(图8u至图8x)从dp4
l112w/v141m
二聚体阳性细胞克隆,在tcr缺陷型jurkat76/cd4细胞中重构,并用相应的dp4
l112w/v141m
二聚体染色。
62.图9a至图9g是条形图,说明在il-2 elispot测定中由用相应肽冲击的aapc刺激的03-ccnd1
219-238
(图9a)、05-hsd17b12
225-244
(图9b)、09-hsd17b12
225-244
(图9c)、05-lgsn
296-315
(图9d)、03-mage-a2
108-127
(图9e)、06-mage-a2
108-127
(图9f)和05-muc5ac
4922-4941
(图9g)的il-2 epispot测定的结果。dp4/wt1(克隆9)tcr用作阴性对照。进行至少2次独立实验。*,p《0.05,通过学生t检验得到。条形和误差条代表一式三份实验结果的平均值
±
sd。
63.图10a至图10q是数据的图形表示,显示从dp4
l112w/v141m
二聚体阳性细胞中分离并在人原代cd4
+
t细胞中重构的dp4限制性tcr以dp4限制性和抗原特异性方式起作用。03-ccnd1
219-238
(图10a至图10d和图10o)、03-mage-a2
108-127
和06-mage-a2
108-127
(图10e至图10j和图10p)和05-muc5ac
4922-4941
(图10k至图10n和图10q)被逆转录病毒转导到人原代cd4
+
t细胞中并用相应的dp4
l112w/v141m
二聚体染色(图10a至图10n)。*p《0.05,通过学生t检验得到。n.s.,不显著。进行至少2次独立实验。*,p《0.05,通过学生t检验得到。条形和误差条代表一式三份实验结果的平均值
±
sd。
64.图11a至图11e呈现的数据显示从黑素瘤患者克隆的dp4限制性tcr识别由基于k562的aapc内源加工和呈递的肽。图11a至图11b是显示了在k562来源的aapc细胞中内源性表达的ccdn1(图11a)和mage-a2(图11b)的凝胶色谱的图像。图11c至图11d是显示了用03-ccnd1
219-238
(图11c)或06-mage-a2
108-127
(图11d)逆转录病毒转导并用未进行肽冲击的hla-空或dp4-aapc刺激的人原代t细胞的ifn-γelispot测定的结果的条形图(图11c至图11d)。图11e是显示了用05-muc5ac
4922-4941
tcr逆转录病毒转导并用muc5ac
4914-4949
小基因转导和未进行肽冲击的hla-空或dp4-aapc刺激的人原代t细胞的ifn-γelispot测定的结果的条形图。进行至少2次独立实验。*,p《0.05,通过学生t检验得到。条形和误差条代表一式三份实验结果的平均值
±
sd。
65.图12a至图12e呈现的数据显示06-mage-a2
108-127
tcr以dp4和mage-a2依赖性方式识别黑素瘤细胞系。图12a是显示k562细胞和所示黑素瘤细胞系中内源性mage-a2表达的western印迹图像。图12b至图12e是条形图,显示来自用sk-mel-21(dp4
+
mage-a2-;图12b)或sk-mel-37(dp4
+
mage-a2
+
;图12c)刺激的用06-mage-a2
108-127 tcr转导的原代人t细胞以及用sk-mel-28(dp4-mage-a2
+
;图12d)和me275(dp4-mage-a2
+
;图12e)刺激用dp4转导的情况的ifn-γelispot测定的数据。*,p《0.05,通过学生t检验得到。条形和误差条代表一式三份实验结果的平均值
±
sd。进行至少2次独立实验。
66.图13a至图13q是比较用抗hla ii类mab克隆9-49染色的k562细胞中野生型hladp*04:01及其衍生物的表达水平的直方图。开放直方图代表同型对照染色。
67.图14a至图14f提供了说明经修饰的dq分子的增强的cd4结合能力的数据。图14a是比较dpb1*04:01、dqb1*05:01和dqb1*05:01
l114w/v143m+4reps
的氨基酸序列的表格,其中突变的氨基酸加下划线。图14b和图14c是用scd4染色的稳定表达野生型dq5(dqa1*01:01/dqb1*05:01)、dq5
l114w/v143m
、dq5
l114w/v143m+4reps
、野生型dp4或dp4
l112w/v141m
(如图14a所示)的ii类缺陷型k562细胞的数据的图形表示。图14d显示了类似地用scd4染色的分别表达在四个位置之一具有单个氨基酸反转的dq5
l114w/v143m+4reps
突变体的一系列k562衍生物的cd4结合能力。图14e是列出dpb1*04:01、dqb1*02:01、dqb1*04:02和dqb1*06:01的氨基酸序列的表格,其中替换的氨基酸加下划线。注意,与dqb1*05:01不同,dqb1*02:01、dqb1*04:02和dqb1*06:01在位置116处编码val,类似于dpb1*04:01,其编码位置114处的val。图14f提供了显示β链中的l114w/v143m+3reps替换增强dq2、dq4和dq6与cd4的结合的数据的图形表示。进行至少2次独立实验。*,p《0.05,通过学生t检验得到。条形和误差条代表一式三份实验结果的平均值
±
sd。
68.图15a至图15b是说明亲和力成熟的dq二聚体检测到在人原代cd4+t细胞中表达的同源tcr的图形表示。将dq5(dqa1*01:01-dqb1*05:01)限制性ddx3y特异性tcr(e6)(图15a)和dq6(dqa1*01:02-dqb1*06:02)限制性流感病毒ha特异性tcr(dm2)(图15b)在人原代cd4+t细胞中重构,并分别通过dq5
l114w/v143m+4reps
和dq6
l114w/v143m+3reps
二聚体染色。进行至少2次独立实验。
69.图16a至图16q是说明hla ii类基因的相当表达水平的直方图的图形表示。将hla-dq及其衍生物在k562细胞中重构并用抗hla ii类单克隆抗体染色。使用抗hla ii类单克隆抗体克隆9-49(i3)(dq5和dq6)或抗ii类单克隆抗体克隆(dq2和dq4)检测每个dq2、dq5和dq6等位基因的表面表达。开放直方图代表同型对照染色。
70.图17a至图17f提供了说明修饰的dr分子的增强的cd4结合能力的数据。图17a是比较dpb1*04:01、drb1*01:01和drb1*01:01
l114w/v143m+6reps
的氨基酸序列的表格,其中突变的氨基酸加下划线。图17b和图17c是用野生型dr1(dra1*01:01/drb1*01:01)、dr1
l114w/v143m
、dr1
l114w/v143m+6reps
、野生型dp4或dp4
l112w/v141m
稳定转导并用scd4染色的ii类缺陷型k562细胞的数据的图形表示。图17d至图17e显示了分别表达在六个位置之一具有单个氨基酸反转的dr1
l114w/v143m+6reps
突变体(类似地用scd4染色)(图17d)和携带s118h和t157i连同l114w/v143m的drb1
l114w/v143m+2reps
(图17e)的一系列k562衍生物的cd4结合能力。图17f是列出dpb1*04:01的氨基酸序列的表格,并且将dr3、dr4、dr7、dr10、dr11和dr13的drb1等位基因与drb1
l114w/v143m+6reps
和drb1
l114w/v143m+2reps
的等位基因一起进行比较,其中突变的氨基酸加下划线。图17g至图17l是显示l114w/v143m+2reps突变比l114w/v143m+6reps突变更好地增强dr3、dr4、dr7、dr10、dr11和dr13与cd4的结合的数据的图形表示。进行至少2次独立实验。*,p《0.05,通过学生t检验得到。条形和误差条代表一式三份实验结果的平均值
±
sd。图17m至图17n是生物层干涉传感图,显示了在一定浓度范围内的生物素化的hla-dr1(配体)与可溶性cd4(分析物)的相互作用。野生型dr1(图17m)和dr1
l114w/v143m+2reps
(图17n)的结合实验平行进行,并且对野生型dr1(图17m)没有检测到结合。图17o是显示了通过稳态分析量化的dr1
l114w/v143m+2reps
和cd4之间的亲和力的图。所有数据代表两次独立实验。
71.图18a至图18d是说明亲和力成熟的dr二聚体检测到的同源tcr在人原代cd4+t细胞中表达的图形表示。将dr1限制性tcr(ha1.7和sb95)(图18a)、dr7限制性tcr(sd334)(图
18b)和dr11限制性tcr(f24)(图18c)在原代人t细胞中重构,并且通过相应的dr
l114w/v143m+2reps
二聚体染色。用dr11
l114w/v143m+2reps
二聚体和抗vβ22 mab染色dr11限制性f24转导的cd4
+
t细胞(图18d)。注意,f24表示vβ22。进行至少2次独立实验。
72.图19a至图19d是hla-dr1
l114w/v143m+2reps
和人cd4复合物的模型结构的图。如所示的,图19a提供dra1*01:01、drb1*01:01和cd4的三元复合物模型结构的概览带状模型。图19b至图19d提供野生型dr1(左)和突变型dr1
l114w/v143m+2reps
(右)中的以下四个突变残基的特写视图:l114w和v143m(图19b)、s118h(图19c)和t157i(图19d),如使用球棍表示说明的。
73.图20a至图20ii是说明hla ii类基因的相当表达水平的直方图的图形表示。将hla-dr及其衍生物在k562细胞中重构并用抗hla ii类单克隆抗体染色。使用抗hla ii类单克隆抗体克隆9-49(i3)检测所有dr等位基因的表面表达。开放直方图代表同型对照染色。
74.图21a至图21d是显示比较dp4
l112w/v141m
二聚体和dextramer对内源性trpc1
578-597
特异性cd4
+
t细胞染色的数据的图形表示。通过用肽冲击的和经辐照的dp4
+
人工apc刺激从黑素瘤患者中扩增内源性(非转导)trpc1
578-597
特异性cd4
+
t细胞并用dp4
l112w/v141m trpc1
578-597
二聚体(图21b)或trpc1
578-597
dextramer(图21d)染色。对应的clip多聚体用作对照(图21a和图21c)。
75.图22a至图22f是显示比较dp4
l112w/v141m
二聚体与常规dp4四聚体和dextramer对内源性ny-eso-1
157-170
特异性t细胞染色的数据的图形表示。从4号dp4
+
健康供体中纯化cd4
+
t细胞,并用ny-eso-1
157-170
冲击的和经辐照的dp4
+
人工apc刺激一次。如由三种不同的dp4多聚体(dp4
l112w/v141m
二聚体(图22b)、dp4四聚体(图22d)或dp4 dextramer(图22f))所示,分别对扩增的cd4
+
t细胞进行染色。
76.图23a至图23y是显示经历了用dp4
l112w/v141m
二聚体进行的离体染色的病原体特异性cd4
+
t细胞的数据的图形表示。从五个dp4
+
供体中纯化记忆cd4
+
t细胞,并在没有体外刺激的情况下针对以下病原体相关肽用dp4
l112w/v141m
二聚体进行离体染色:tt
948-968
(图23f至图23j)、hsv-2-ul21
283-302
(图23k至图23o)、flu-ha
527-546
(图23p至图23t)和rsv-gp
162-175
(图23u至图23y)。clip肽用作阴性对照(图23a至图23e)。
77.图24a至图24w是显示从dp4
l112w/v141m
二聚体
+
细胞成功建立的内源性rsv-gp
162-175
特异性cd4
+
t细胞克隆的数据的图形表示。从06号dp4
+
供体中纯化记忆cd4
+
t细胞,并在没有体外刺激的情况下用dp4
l112w/v141m rsv-gp
162-175
二聚体进行离体染色。然后通过有限稀释克隆二聚体
+
cd4
+
t细胞。图24a至图24v是10个二聚体阳性和1个二聚体阴性单细胞克隆的代表性二聚体染色数据的图形表示。将84个克隆中的77个(91.7%)成功地用dp4
l112w/v141m rsv-gp
162-175
二聚体染色。图24w是显示了rsv-gp
162-175
二聚体
+
单细胞克隆中的抗原特异性il-2产生的条形图。
78.图25a至图25s是显示从dp4
l112w/v141m
二聚体
+
细胞成功建立的内源性dp4 tt
948-968
特异性cd4
+
t细胞克隆的数据的图形表示。从04号dp4
+
供体中纯化记忆cd4
+
t细胞,并在没有体外刺激的情况下用dp4
l112w/v141m tt
948-968
二聚体进行离体染色。然后通过有限稀释克隆二聚体
+
cd4
+
t细胞。图25a至图25r是8个二聚体阳性和1个二聚体阴性单细胞克隆的代表性二聚体染色数据的图形表示。29个克隆中的26个(89.7%)成功地用dp4
l112w/v141m tt
948-968
二聚体染色。图25s是显示tt
948-968
二聚体
+
单细胞克隆中的抗原特异性il-2产生的条形图。
79.图26a至图26nn是rsv-gp(图26a至图26p)和tt(图26o至图26nn)二聚体
+
单细胞克
隆的dp4多聚体染色的图形表示。将rsv-gp二聚体
+
单细胞克隆(c6、c12、c26和c39)用dp4
l112w/v141m rsv-gp
162-175
二聚体(图26b、图26d、图26f和图26h)或野生型dp4dextramer(图26j、图26l、图26n和图26p)染色。将tt二聚体
+
单细胞克隆(c2、c4、c6和c9)分别用三种不同的dp4 tt
948-968
多聚体(dp4
l112w/v141m
二聚体(图26r、图26t、图26v和图26x)、野生型dp4四聚体(图26z、图26bb、图26dd和图26ff)和野生型dp4dextramer(图26hh、图26jj、图26ll和图26nn))染色。
80.图27a至图27l是显示dq5
l114w/v143m+4reps
二聚体稳健染色e6转导的cd4
+
t细胞的图形表示。将e6在cd4
+
t细胞中重构,然后用野生型dq5(图27d和27j)、dq5
l114w/v143m
(图27e和27k)和dq5
l114w/v143m+4reps
(图27f和图27l)clip对照二聚体(图4d至图4f)和对ddx3y
171-190
有特异性的二聚体(图27j至图27l)染色。未用tcr转导的对照细胞示于图27a至图27c和图27g至图27i中。
81.图28a至图28h是显示使用亲和力成熟二聚体克隆dq5限制性tcr的图形表示。从dq5.1
+
黑素瘤患者中纯化原代cd4
+
t细胞,并用经辐照的gpc3
138-157
冲击的表达dq5.1的aapc刺激。两周后,用同源gpc3
138-157-dq5
l114w/v143m+4reps
二聚体对刺激的cd4
+
t细胞进行染色(图28a至图28b)。将gpc3特异性tcr在tcr缺陷型jurkat76/cd4细胞中重构,并通过相应的dq5
l114w/v143m+4reps
二聚体(图28c(e6/对照);图28d(e6/gpc3
138-157
);图28e(dq5-06-gpc3
138-157
/对照);和图28f(dq5-06-gpc3
138-157
/gpc3
138-157
))染色。在il-2elispot测定中,将表达gpc3特异性tcr的jurkat 76/cd4细胞由用相应肽冲击的dq5-k562细胞刺激(图28g)。
82.图29a至图29l是显示经历用dr1
l114w/v143m+2reps
二聚体进行的离体染色的流感病毒血球凝集素特异性外周cd4
+
t细胞的图形表示。从两个dr1
+
供体(07号(图29a至图29f)和08号(图29g至图29l))中纯化记忆cd4
+
t细胞,并在没有体外刺激的情况下,用对flu-ha
5-24
(图29b和图29h)、flu-ha
117-136
(图29c和图29i)、flu-ha
232-251
(图29d和图29j)、flu-ha
268-287
(图29e和图29k)和flu-ha
306-318
(图29f和图29l)流感病毒血球凝集素(flu-ha)肽有特异性的dr1
l114w/v143m+2reps
二聚体染色。clip肽用作阴性对照(图29a和图29g)。
83.图30a至图30x是显示dr1
l114w/v143m+6reps
和dr1
l114w/v143m+2reps
二聚体稳健染色的ha1.7转导的cd4
+
t细胞的图形表示。将ha1.7在原代cd4
+
t细胞中重构,然后在没有转导的tcr(图30i至图30l)和用ha1.7 tcr转导(图30m至图30p)的情况下,用野生型dr1(图30i和30m)、dr1
l114w/v143m
(图30j和30n)、dr1
l114w/v143m+6reps
(图30k和30o)和dr1
l114w/v143m+2reps
(图30l和30p)二聚体染色,用clip二聚体作为阴性对照(图30a至图30h)。此外,将ha1.7在原代cd4
+
t细胞中重构,然后用对flu-ha
306-318
有特异性的dr1
l114w/v143m+2reps
二聚体(图30o至图30t)或野生型dr1 dextramer(图30u至图30x)染色。
84.图31a至图31p是显示使用亲和力成熟二聚体克隆dr1限制性tcr的数据的图形表示。从两个dr1
+
黑素瘤患者中纯化原代cd4
+
t细胞,并用经辐照的hsd17b12
225-244
冲击(图31b)和ly6k
99-118
冲击(图31d)的表达dr1的aapc刺激。两周后,用同源dr1
l114w/v143m+2reps
二聚体对刺激的cd4
+
t细胞进行染色(图31a至图31d)。将dr1限制性tcr在原代cd4
+
t细胞中重构,并通过相应的二聚体染色(图31e至图31m)。在il-2 elispot测定中,将表达dr1限制性dr1-07-hsd17b12
225-244
(图31n)和dr1-08-ly6k
99-118
(图31o)tcr的原代cd4
+
t细胞分别由用hsd17b12
225-244
(图31n)和ly6k
99-118
(图31o)肽冲击的dr1-k562细胞刺激。
具体实施方式
85.本公开涉及鉴定mhc ii类特异性tcr的方法,所述方法包括使t细胞与包含mhc ii类分子和肽的复合物接触,其中所述mhc ii类分子对cd4的亲和力高于天然存在的mhc ii类分子对cd4的亲和力。在一些方面,mhc ii类分子包含α链和β链,其中所述mhc ii类分子的β链包含相对于mhc ii类分子的野生型β链具有一个或多个突变的氨基酸序列。
86.i.术语
87.为使本公开可更易于被理解,首先定义某些术语。如本技术中所使用的,除非本文另有明确规定,否则以下术语中的每一个应具有下文所述的含义。另外的定义在整个申请中阐述。
88.应注意术语“一个(种)(a/an)”实体是指一个(种)或多个(种)所述实体;例如”一个(种)核苷酸序列”应理解为表示一个(种)或多个(种)核苷酸序列。因此,术语“一个(种)”、“一个(种)或多个(种)”和“至少一个(种)”在本文中可互换使用。
89.此外,当在本文中使用时,“和/或”应视为特定公开两个指定特征或组分中的每一者,伴有或不伴有另一者。因此,如本文中诸如“a和/或b”的短语中所用的术语“和/或”意图包括“a和b”、“a或b”、“a”(单独)和“b”(单独)。类似地,在短语中使用的术语“和/或”诸如“a、b和/或c”意图涵盖以下方面中的每一者:a、b和c;a、b或c;a或c;a或b;b或c;a和c;a和b;b和c;a(单独);b(单独);和c(单独)。
90.术语“约”在本文中用于意指近似、大致、约或在
……
左右。当术语“约”与数值范围结合使用时,它通过使边界延伸高于和低于数值来修饰范围。一般来说,术语“约”在本文中用于以高于和低于值10%上下(升高或降低)的变化来修饰数值。
91.应了解,每当在本文中以措辞“包含”描述各个方面的情况下,还提供了以“由
……
组成”和/或“基本上由
……
组成”描述的另外类似方面。
92.除非另外定义,否则本文所用的所有技术和科学术语都具有与由本公开所相关领域中的普通技术人员通常理解相同的含义。例如,concise dictionary of biomedicine and molecular biology,juo,pei-show,第2版,2002,crc press;dictionary of cell and molecular biology,第3版,1999,academic press;和oxford dictionary of biochemistry and molecular biology,修订板,2000,oxford university press,给普通技术人员提供本公开中使用的许多术语的通用字典。
93.单位、前缀和符号以它们的国际单位制(si)接受形式表示。数值范围包括界定范围的数值。除非另外指示,否则核苷酸序列以5'至3'定向从左至右书写。氨基酸序列以氨基至羧基定向从左至右书写。本文提供的标题并非是对本公开的各个方面的限制,所述各个方面可通过参考说明书整体而得到。因此,通过参考说明书整体,以下紧接着定义的术语得到更充分定义。
[0094]“施用”是指使用本领域技术人员已知的各种方法和递送系统中的任一种将剂以物理方式引入受试者。用于本文公开的配制品的示例性施用途径包括静脉内、肌内、皮下、腹膜内、脊髓或其他肠胃外施用途径,例如通过注射或输注。如本文所用,短语“肠胃外施用”意指除肠内和局部施用之外的施用方式,通常通过注射,并且包括但不限于静脉内、肌肉内、动脉内、鞘内、淋巴管内、病灶内、囊内、眶内、心内、皮内、腹膜内、经气管、皮下、表皮下、关节内、囊下、蛛网膜下、脊柱内、硬膜外以及胸骨内注射和输注,以及体内电穿孔。在一
些方面,配制品通过非肠胃外途径施用,例如口服。其他非肠胃外途径包括局部、表皮或粘膜施用途径,例如鼻内、阴道、直肠、舌下或局部。施用还可以例如进行一次、多次和/或经一个或多个延长的时间段。
[0095]
如本文所用,术语“t细胞受体”(tcr)是指能够与靶抗原特异性相互作用的异聚细胞表面受体。如本文所用,“tcr”包括但不限于天然存在的和非天然存在的tcr;全长tcr及其抗原结合部分;嵌合tcr;tcr融合构建体;和合成tcr。在人中,tcr在t细胞的表面上表达,并且它们负责t细胞识别和抗原呈递细胞的靶向。抗原呈递细胞(apc)展示与主要组织相容性复合物(mhc)复合(本文也称为与hla分子(例如hla ii类分子)复合)的外来蛋白(抗原)片段。tcr识别并结合肽:hla复合物并募集cd8(对于mhc i类分子)或cd4(对于mhc ii类分子),从而激活tcr。激活的tcr启动下游信号传导和免疫反应,包括epc的破坏。
[0096]
通常,tcr可以包含两条链,α链和β链(或不太常见地γ链和δ链),它们通过二硫键相互连接。每条链都包含可变区(α链可变结构域和β链可变结构域)和恒定区(α链恒定区和β链恒定区)。可变结构域位于细胞膜远侧,并且可变结构域与抗原相互作用。恒定区位于细胞膜近侧。tcr可以进一步包含跨膜区和短的细胞质尾。如本文所用,术语“恒定区”包括跨膜区和细胞质尾以及(当存在时)传统的“恒定区”。
[0097]
可变结构域可以进一步细分为高变区,称为互补决定区(cdr),其中散布着更保守的区域,称为框架区(fr)。每个α链可变结构域和β链可变结构域包含三个cdr和四个fr:fr1、cdr1、fr2、cdr2、fr3、cdr3、fr4。每个可变结构域都包含与抗原相互作用的结合结构域。虽然每条链上的所有三个cdr都参与抗原结合,但cdr3被认为是主要的抗原结合区,而cdr1和cdr2被认为主要识别hla分子。
[0098]
在没有明确说明的情况下,除非上下文另有指示,否则术语“tcr”还包括本文公开的任何tcr的抗原结合片段或抗原结合部分,并且包括单价和二价片段或部分,以及单链tcr。术语“tcr”不限于与t细胞的表面结合的天然存在的tcr。如本文所用,术语“tcr”进一步指本文所述的在除t细胞之外的细胞(例如,如本文所述天然表达或经修饰以表达cd4的细胞)的表面上表达的tcr或本文所述的不含细胞膜的tcr(例如,分离的tcr或可溶性tcr)。
[0099]“抗原结合分子”、“tcr的部分”或“tcr片段”是指tcr的小于整体的任何部分。抗原结合分子可以包括抗原性cdr。
[0100]“抗原”是指任何分子,例如肽,其引起免疫反应或能够被tcr结合。如本文所用,“表位”是指引起免疫反应或能够被tcr结合的多肽的一部分。免疫反应可能涉及抗体的产生或特定免疫活性细胞的激活,或两者兼而有之。本领域技术人员将容易理解任何大分子(包括几乎所有的蛋白质或肽)都可以用作抗原。抗原和/或表位可以内源表达,即通过基因组dna表达,或者可以重组表达。抗原和/或表位可以特异于特定组织,如患病细胞,例如癌细胞,或者它可以被广泛表达。此外,较大分子的片段可以充当抗原。在一方面,抗原是肿瘤抗原。表位可以存在于较长多肽中(例如,蛋白质中),或者表位可以作为较长多肽的片段存在。在一些方面,表位与主要组织相容性复合物(mhc)复合(在本文中也称为与hla分子(例如hla 1类分子)复合)。
[0101]
术语“自体”是指来源于同一个体的任何材料,之后将所述材料重新引入所述个体中。例如,自体t细胞疗法包括向受试者施用从同一受试者分离的t细胞。术语“同种异体”是指来源于一个个体的任何材料,然后将所述材料引入同一物种的另一个个体中。例如,同种
异体t细胞移植包括向受试者施用从除所述受试者之外的供体获得的t细胞。
[0102]
如本文所用,“ccnd1”、“g1/s特异性细胞周期蛋白-d1”、“b细胞淋巴瘤1蛋白”、“bcl-1”或“prad1”是指磷酸化和抑制包括rb1在内的视网膜母细胞瘤(rb)蛋白家族成员且调节g1/s过渡期间的细胞周期的细胞周期蛋白d1-cdk4(dc)复合物的人调节成分。rb1的磷酸化允许转录因子e2f从rb/e2f复合物中解离并随后转录负责g1期进展的e2f靶基因。ccnd1还参与早期g1期的rb1低磷酸化。细胞周期蛋白d-cdk4复合物是各种促有丝分裂和抗有丝分裂信号的主要整合剂。ccnd1也是smad3的底物,以细胞周期依赖性方式磷酸化smad3并阻遏其转录活性。ccnd1也是三元复合物细胞周期蛋白d1/cdk4/cdkn1b的组分,是细胞周期蛋白d-cdk4复合物的核转位和活性所必需的,并且ccnd1在neurod1和ins启动子上以细胞周期依赖性方式与insm1一起展现出转录共阻遏活性。ccnd1的突变、扩增和过表达会改变细胞周期进程,这在多种肿瘤中经常观察到,并可能导致肿瘤发生。
[0103]
如本文所用,ccnd1不仅指全长典范序列,还指其变体和片段。ccnd1的氨基酸序列(seq id no:27)提供于表1a中(uniprotkb
–
p24385)。
[0104]
表1a.ccnd1氨基酸序列
[0105][0106]
如本文所用,“muc5ac”或“粘蛋白5ac”是指胃和呼吸道上皮的人凝胶形成糖蛋白,其通过与随后被粘液纤毛系统去除的吸入微生物和颗粒结合来保护粘膜免受感染和化学损伤。
[0107]
如本文所用,muc5ac不仅指全长典范序列,还指其变体和片段。muc5ac的氨基酸序列(seq id no:28)提供于表1b中(uniprotkb
–
p98088)。
[0108]
表1b.muc5ac氨基酸序列
[0109]
[0110]
[0111][0112]
如本文所用,“mage-a2”、“黑素瘤相关抗原2”或“癌症/睾丸抗原1.2”是指主要由肿瘤细胞表达的人蛋白质。mage-a2通过将hdac3募集到p53/tp53转录位点来降低p53/tp53反式激活功能。mage-a2阻遏p73/tp73活性。在体外,mage-a2促进黑素瘤细胞系的细胞活力。mage-a2在几种类型的多种肿瘤中表达,如黑素瘤、头颈部鳞状细胞癌、肺癌和乳腺癌。然而,在健康组织中,mage-a2仅在睾丸中表达。
[0113]
如本文所用,mage-a2不仅指全长序列,还指其变体和片段。mage-a2的氨基酸序列(seq id no:29)提供于表1c中(uniprotkb
–
p43356)。
[0114]
表1c.mage-a2氨基酸序列
[0115][0116]
如本文所用,术语“hla”是指人白细胞抗原。在人中,hla基因编码主要组织相容性复合物(mhc)蛋白。mhc蛋白在细胞的表面上表达,并参与免疫反应的激活。hla ii类基因编码在专职抗原呈递细胞(apc)的表面上表达的mhc ii类蛋白。专职apc的非限制性示例包括单核细胞、巨噬细胞、树突细胞(dc)和b淋巴细胞。一些内皮细胞和上皮细胞在炎症信号被激活后也表达mhc ii类分子。缺乏功能性mhc ii类分子的人极易感染一系列感染性疾病,并且典型地在年轻时死亡。
[0117]
如本文所用,“hla ii类分子”或“mhc ii类分子”是指编码mhc ii类分子的野生型或变体hla ii类基因的蛋白质产物。因此,“hla ii类分子”和“mhc ii类分子”在本文中可互换使用。典型的mhc ii类分子包含两条蛋白质链:α链和β链。一般来说,天然存在的α链和
β链各自包含将α/β链锚定到细胞表面的跨膜结构域,以及携带抗原并与细胞上表达的tcr和/或cd4相互作用的胞外结构域。
[0118]
mhc ii类α和β链两者均由hla基因复合物编码。hla复合物位于人6号染色体短臂的6p21.3区域内,并包含多于220个具有不同功能的基因。本领域已知,hla基因复合物高度变异,具有超过20,000个hla等位基因和相关等位基因,包括超过250个mhc ii类α链等位基因和5,000个mhc ii类β链等位基因,编码数千种mhc ii类蛋白(参见,例如,hla.alleles.org,上次访问时间为2019年5月20日,通过引用以其整体并入本文)。例如,一种这样的hla-dp等位基因,dp4,是许多种族群体中最常见的等位基因。
[0119]
hla复合物中的三个基因座编码mhc ii类蛋白:hla-dp、hla-dq和hla-dr。hla-do和hla-dm编码与mhc ii类分子缔合的蛋白质并支持其构型和功能。
[0120]
当mhc ii类分子与抗原肽复合时,10-30个氨基酸长的抗原肽结合肽结合槽并在细胞外被呈递给cd4+细胞。α链和β链两者都折叠成两个独立的结构域;α多肽的α-1和α-2,以及β多肽的β-1和β-2。在α-1和β-1结构域之间发现了保持呈递抗原的开放式肽结合槽。在与cd4+t细胞相互作用时,mhc ii类复合物与在t细胞表面上表达的t细胞受体(tcr)相互作用。此外,mhc ii类分子的β链与t细胞表面上表达的cd4弱相互作用(kd》2mm)。典范cd4氨基酸序列(uniprot-p01730)提供于表2中(seq id no:10)。
[0121]
表2.人cd4氨基酸序列
[0122][0123]
术语“自体”是指来源于同一个体的任何材料,之后将所述材料重新引入所述个体中。例如,自体t细胞疗法包括向受试者施用从同一受试者分离的t细胞。术语“同种异体”是指来源于一个个体的任何材料,然后将所述材料引入同一物种的另一个个体中。例如,同种异体t细胞移植包括向受试者施用从除所述受试者之外的供体获得的t细胞。
[0124]“癌症”是指广泛的一组各类疾病,其特征在于体内异常细胞的不受控制的生长。不受调控的细胞分裂和生长导致恶性肿瘤的形成,其侵入邻近组织并且还可以通过淋巴系统或血流转移到身体的远侧部分。“癌症”或“癌组织”可以包括肿瘤。可以通过本发明的方法治疗的癌症的示例包括但不限于免疫系统的癌症,包括淋巴瘤、白血病和其他白细胞恶性肿瘤。在一些方面,本发明的方法可用于减小来源于以下项的肿瘤的肿瘤大小,例如骨癌、肾癌、前列腺癌、乳腺癌、结肠癌、肺癌、皮肤或眼内恶性黑素瘤、胰腺癌、皮肤癌、头颈癌、皮肤或眼内恶性黑素瘤、子宫癌、卵巢癌、直肠癌、肛区癌、胃癌、睾丸癌、子宫癌、输卵管癌、子宫内膜癌、宫颈癌、阴道癌、外阴癌、霍奇金病、非霍奇金淋巴瘤(nhl)、原发纵隔大b细胞淋巴瘤(pmbc)、弥漫性大b细胞淋巴瘤(dlbcl)、滤泡性淋巴瘤(fl)、转化型滤泡性淋巴瘤、脾边缘区淋巴瘤(smzl)、食道癌、小肠癌、内分泌系统癌、甲状腺癌、甲状旁腺癌、肾上腺
癌、软组织肉瘤、尿道癌、阴茎癌、急性或慢性白血病、急性髓性白血病(aml)、慢性髓性白血病、急性成淋巴细胞性白血病(all)(包括非t细胞all)、慢性淋巴细胞性白血病(cll)、儿童实体瘤、淋巴细胞性淋巴瘤、膀胱癌、肾或输尿管癌、肾盂癌、中枢神经系统瘤、原发性cns淋巴瘤、肿瘤血管生成、脊髓轴肿瘤、脑干胶质瘤、垂体腺瘤、卡波西肉瘤、表皮样癌、鳞状细胞癌、t细胞淋巴瘤、包括由石棉诱导的那些的环境诱导癌症、其他b细胞恶性肿瘤及所述癌症的组合。特定的癌症可以对化学疗法或放射疗法有反应,或者癌症可以是难治性的。
[0125]
难治性癌症是指不能适用手术干预的癌症,并且所述癌症最初对化学疗法或放射疗法没有反应,或者随着时间的推移所述癌症变得没有反应。
[0126]
如本文所用,“抗肿瘤作用”是指可以呈现为肿瘤体积减小、肿瘤细胞数量减少、肿瘤细胞增殖减少、转移数量减少、总生存期或无进展生存期的增加、预期寿命的增加或与肿瘤相关的各种生理学症状的改善。抗肿瘤作用也可以指预防肿瘤的发生,例如疫苗。
[0127]
如本文所用,术语“无进展生存期”可缩写为pfs,是指从治疗日期到根据修订的iwg恶性淋巴瘤反应标准的疾病进展日期或任何原因导致的死亡的时间。
[0128]
如本文所用,“疾病进展”或“进展性疾病”可缩写为pd,是指与特定疾病相关的一种或多种症状的恶化。例如,患有癌症的受试者的疾病进展可以包括一个或多个恶性病灶的数量或大小的增加、肿瘤转移和死亡。
[0129]
如本文所用,“反应持续时间”可缩写为dor,是指受试者的第一次客观反应与根据经修订的iwg恶性淋巴瘤反应标准确认的疾病进展日期或死亡之间的时间段。
[0130]
术语“总生存期”可缩写为os,定义为从治疗日期到死亡日期的时间。
[0131]
如本文所用,“细胞因子”是指由一个细胞响应于与特定抗原接触而释放的非抗体蛋白,其中所述细胞因子与第二个细胞相互作用以介导第二个细胞中的反应。细胞因子可以由细胞内源性表达或施用于受试者。免疫细胞(包括巨噬细胞、b细胞、t细胞和肥大细胞)可以释放细胞因子以传播免疫反应。细胞因子可以在受体细胞中诱导各种反应。细胞因子可以包括稳态细胞因子、趋化因子、促炎细胞因子、效应子和急性期蛋白。例如,稳态细胞因子,包括白细胞介素(il)7和il-15,可促进免疫细胞生存和增殖,而促炎细胞因子可促进炎症反应。稳态细胞因子的示例包括但不限于il-2、il-4、il-5、il-7、il-10、il-12p40、il-12p70、il-15和干扰素(ifn)γ。促炎细胞因子的示例包括但不限于il-1a、il-1b、il-6、il-13、il-17a、肿瘤坏死因子(tnf)-α、tnf-β、成纤维细胞生长因子(fgf)2、粒细胞巨噬细胞集落刺激因子(gm-csf)、可溶性细胞间粘附分子1(sicam-1)、可溶性血管粘附分子1(svcam-1)、血管内皮生长因子(vegf)、vegf-c、vegf-d和胎盘生长因子(plgf)。效应子的示例包括但不限于颗粒酶a、颗粒酶b、可溶性fas配体(sfasl)和穿孔素。急性期蛋白的示例包括但不限于c反应蛋白(crp)和血清淀粉样蛋白a(saa)。
[0132]“趋化因子”是一种类型的介导细胞趋化性或定向运动的细胞因子。趋化因子的示例包括但不限于il-8、il-16、嗜酸性粒细胞趋化因子、嗜酸性粒细胞趋化因子-3、巨噬细胞来源的趋化因子(mdc或ccl22)、单核细胞趋化蛋白1(mcp-1或ccl2)、mcp-4、巨噬细胞炎症蛋白1α(mip-1α、mip-1a)、mip-1β(mip-1b)、γ诱导蛋白10(ip-10)以及胸腺和激活调节的趋化因子(tarc或ccl17)。
[0133]
本发明的分析物和细胞因子的其他示例包括但不限于趋化因子(c-c基序)配体(ccl)1、ccl5、单核细胞特异性趋化因子3(mcp3或ccl7)、单核细胞趋化蛋白2(mcp-2或
ccl8)、ccl13、il-1、il-3、il-9、il-11、il-12、il-14、il-17、il-20、il-21、粒细胞集落刺激因子(g-csf)、白血病抑制因子(lif)、制瘤素m(osm)、cd154、淋巴毒素(lt)β、4-1bb配体(4-1bbl)、增殖诱导配体(april)、cd70、cd153、cd178、糖皮质激素诱导tnfr相关配体(gitrl)、肿瘤坏死因子超家族成员14(tnfsf14)、ox40l、tnf和apol相关白细胞表达配体1(tall-1)或tnf相关凋亡诱导配体(trail)。
[0134]
药物或治疗剂的“治疗有效量”、“有效剂量”、“有效量”或“治疗有效剂量”是所述药物的在单独或与另一治疗剂组合使用时,由降低疾病症状的严重程度、增加疾病无症状期的频率和持续时间、或预防由于疾病病痛引起的损伤或伤残证明的,保护受试者免于疾病发作或促进疾病消退的任何量的药物。可以使用本领域技术人员已知的多种方法评估治疗剂促进疾病消退的能力,例如在临床试验期间的人受试者中,在预测人中功效的动物模型系统中,或通过测定剂在体外测定中的活性。
[0135]
如本文所用,术语“感染”是指外来物质对身体的一个或多个组织的任何类型的侵入。术语“感染”包括但不限于由病毒(包括类病毒和朊病毒)、细菌、真菌、寄生虫及其任何组合引起的感染。
[0136]
如本文所用,术语“淋巴细胞”包括自然杀伤(nk)细胞、t细胞或b细胞。nk细胞是一种类型的细胞毒性(cytotoxic/cell toxic)淋巴细胞,代表固有免疫系统的主要组分。nk细胞排斥肿瘤和被病毒感染的细胞。它通过细胞凋亡或程序性细胞死亡过程起作用。它们被称为“自然杀伤物”,因为它们不需要激活即可杀伤细胞。t细胞在细胞介导的免疫中起主要作用(无抗体参与)。t细胞受体(tcr)将t细胞与其他淋巴细胞类型区分开来。胸腺是免疫系统的一个特殊器官,主要负责t细胞的成熟。t细胞有六种类型,即:辅助t细胞(例如cd4+细胞)、细胞毒性t细胞(也称为tc、细胞毒性t淋巴细胞、ctl、t杀伤细胞、溶细胞性t细胞、cd8+t细胞或杀伤t细胞),记忆t细胞((i)干记忆t
scm
细胞,与幼稚细胞一样,是cd45ro-、ccr7+、cd45ra+、cd62l+(l-选择素)、cd27+、cd28+和il-7rα+,但它们还表达大量的cd95、il-2rβ、cxcr3和lfa-1,并显示出记忆细胞独特的许多功能属性);(ii)中央记忆t
cm
细胞表达l-选择素和ccr7,它们分泌il-2,但不分泌ifnγ或il-4,和(iii)效应记忆t
em
细胞,然而,不表达l-选择素或ccr7但产生效应细胞因子,如ifnγ和il-4)、调节性t细胞(treg、抑制性t细胞或cd4+cd25+调节性t细胞)、自然杀伤t细胞(nkt)和γδt细胞。另一方面,b细胞在体液免疫中起主要作用(有抗体参与)。b细胞制造抗体和抗原,并发挥抗原呈递细胞(apc)的作用,并在由抗原相互作用激活后转变为记忆b细胞。在哺乳动物中,未成熟的b细胞在骨髓中形成,它的名字源于此。
[0137]
术语“修饰的”和“突变的”在本文中用于指核苷酸或氨基酸序列时,是指序列相对于野生型序列或特定参考序列的变化。除非另有说明,术语“修饰的”和“突变的”不需要用于制备修饰或突变的序列(例如,修饰的β链序列)的过程中的步骤。相反,这些术语指示修饰或突变的序列相对于参考序列(例如野生型序列)存在变异。例如,包含在对应于seq id no:1的氨基酸残基112的位置处的取代突变的dpβ链不需要野生型dpβ链已经物理改变以得到所述dpβ链;而是当正确排列时,所述dpβ链包含在所述位置(残基112)处的如下氨基酸残基,该氨基酸残基不同于野生型或参考dpβ链中对应位置处的氨基酸残基。
[0138]
如本文所用,术语“任何氨基酸”是指任何已知的氨基酸。氨基酸是包含以下项的有机化合物:(i)胺(-nh2)官能团,(ii)羧基(-cooh)_官能团,和(iii)侧链(r基团),其中所
述侧链特异于每个氨基酸。这包括但不限于任何天然存在的氨基酸以及其任何修饰和变体。约有500种天然存在的氨基酸,其中20种由遗传密码编码。具有带正电的侧链的氨基酸包括精氨酸(arg;r)、组氨酸(his;h)和赖氨酸(lys;k)。具有带负电的侧链的氨基酸包括天冬氨酸(asp;d)和谷氨酸(glu;e)。具有极性不带电侧链的氨基酸包括丝氨酸(ser;s)、苏氨酸(thr;t)、谷氨酰胺(gln;q)和天冬酰胺(asn;n)。具有疏水侧链的氨基酸包括丙氨酸(ala;a)、异亮氨酸(ile;i)、亮氨酸(leu;l)、甲硫氨酸(met;m)、苯丙氨酸(phe;f)、缬氨酸(val;v)、色氨酸(trp;w)、酪氨酸(tyr;y)。色氨酸(trp;w)、酪氨酸(tyr;y)和甲硫氨酸(met;m)也可归类为极性和/或两亲性氨基酸,因为这些氨基酸通常存在于蛋白质或脂质膜的表面。另外的氨基酸包括半胱氨酸(cys;c)、硒代半胱氨酸(sec;u)、甘氨酸(gly;g)和脯氨酸(pro;p)。
[0139]
如本文所用,“在对应于
……
的位置处”用作用以鉴定多核苷酸中的特定氨基酸残基(例如特定氨基酸位置)或多肽中的特定核酸(例如特定核酸位置)的手段。可以通过将讨论中的序列与参考序列正确比对来确定位置。本领域技术人员将容易理解如何与序列比对以确定相对位置。例如,可在线获得各种比对工具,包括但不限于www.ebi.ac.uk(上次访问时间为2019年5月25日)上获得的“clustal omega multiple sequence alignment”。
[0140]
术语“基因工程化的”或“工程化的”是指修饰细胞的基因组的方法,所述方法包括但不限于缺失编码区或非编码区或其部分,或插入编码区或其部分。在一些方面,修饰的细胞是淋巴细胞,例如表达cd4的t细胞或经修饰的细胞,其可以从患者或供体获得。可以修饰细胞以表达外源构建体,例如像本文公开的t细胞受体(tcr),其被掺入到细胞基因组中。在一些方面,细胞被修饰以表达cd4。
[0141]“免疫反应”是指免疫系统的细胞(例如,t淋巴细胞、b淋巴细胞、自然杀伤(nk)细胞、巨噬细胞、嗜酸性粒细胞、肥大细胞、树突细胞和嗜中性粒细胞)和由任何这些细胞或肝脏产生的可溶性大分子(包括ab、细胞因子和补体)的作用,这些可溶性大分子导致选择性靶向、结合、损害、破坏和/或消除脊椎动物体内的入侵病原体、感染病原体的细胞或组织、癌细胞或其他异常细胞、或(在自身免疫或病理学炎症的情况下)正常人体细胞或组织。
[0142]
术语“免疫疗法”是指通过包括诱导、增强、抑制或以其他方式修改免疫反应的方法治疗患有疾病或者处于感染疾病或遭受疾病复发风险下的受试者。免疫疗法的示例包括但不限于t细胞疗法。t细胞疗法可包括过继性t细胞疗法、肿瘤浸润淋巴细胞(til)免疫疗法、自体细胞疗法、工程化自体细胞疗法(eact)和同种异体t细胞移植。
[0143]
本文所述的免疫疗法中使用的细胞可以来自本领域已知的任何来源。例如,可以在体外从造血干细胞群中分化出t细胞,或者可以从受试者中获得t细胞。t细胞可以从例如外周血单核细胞、骨髓、淋巴结组织、脐带血、胸腺组织、感染部位的组织、腹水、胸腔积液、脾组织和肿瘤中获得。此外,t细胞可以来源于本领域可得到的一种或多种t细胞系。t细胞也可以使用任何数量的本领域技术人员已知的技术(如ficoll
tm
分离和/或单采术)由从受试者收集的单位血液中获得。分离t细胞用于t细胞疗法的另外的方法在美国专利公开第2013/0287748号中公开,该专利通过引用以其整体并入本文。免疫疗法还可以包括向受试者施用经修饰的细胞,其中所述经修饰的细胞表达本文公开的cd4和tcr。在一些方面,经修饰的细胞不是t细胞。
[0144]
如本文所用,“患者”包括患有癌症(例如,淋巴瘤或白血病)的任何人。术语“受试
者”和“患者”在本文中可互换使用。
[0145]
术语“肽”、“多肽”和“蛋白质”在可互换使用,并且是指由通过肽键共价连接的氨基酸残基构成的化合物。蛋白质或肽必须含有至少两个氨基酸,并且对可以组成蛋白质或肽的序列的氨基酸的最大数目没有限制。多肽包括包含通过肽键彼此连接的两个或更多个氨基酸的任何肽或蛋白质。如本文所用,所述术语既指例如在本领域中也通常称为肽、寡肽和寡聚体的短链,还指在本领域中通常称为蛋白质的更长链,其中有很多类型。“多肽”包括例如生物活性片段、基本上同源的多肽、寡肽、同二聚体、异二聚体、多肽的变体、经修饰的多肽、衍生物、类似物、融合蛋白等。多肽包括天然肽、重组肽、合成肽或其组合。
[0146]
如本文所用,“刺激”是指由刺激分子与其同源配体结合诱导的初级反应,其中该结合介导信号转导事件。“刺激分子”是t细胞上的分子,例如t细胞受体(tcr)/cd4复合物,其与抗原呈递细胞上存在的同源刺激配体特异性结合。“刺激性配体”是当存在于抗原呈递细胞(例如,aapc、树突细胞、b细胞等)上时可与t细胞上的刺激性分子特异性结合,从而介导t细胞的初级反应,包括但不限于激活、免疫反应的启动、增殖等。刺激性配体包括但不限于装载有肽、抗cd4抗体、超激动剂抗cd2抗体、超激动剂抗cd28抗体和超激动剂抗cd3抗体的mhc ii类分子。
[0147]
术语“调理”和“预调理”在本文中可互换使用,并指示使需要t细胞疗法的患者为合适的病症做好准备。如本文所用的调理包括但不限于在t细胞疗法之前减少内源性淋巴细胞的数量、去除细胞因子沉没、增加一种或多种稳态细胞因子或促炎因子的血清水平、增强在调理后施用的t细胞的效应功能、增强抗原呈递细胞激活和/或可用性、或其任何组合。在一个方面,“调理”包括增加以下一种或多种细胞因子的血清水平,例如白细胞介素7(il-7)、白细胞介素15(il-15)、白细胞介素10(il-10)、白细胞介素5(il-5)、γ-诱导蛋白10(ip-10)、白细胞介素8(il-8)、单核细胞趋化蛋白1(mcp-1)、胎盘生长因子(plgf)、c反应蛋白(crp)、可溶性细胞间粘附分子1(sicam-1)、可溶性血管粘附分子1(svcam-1)、或其任何组合。在另一个方面,“调理”包括增加il-7、il-15、ip-10、mcp-1、plgf、crp或其任何组合的血清水平。
[0148]
对受试者的“治疗(treatment或treating)”是指对受试者进行的任何类型的干预或处理,或向受试者施用活性剂,目的是逆转、减轻、改善、抑制、减缓或预防症状、并发症或病症或者与疾病相关的生化指标的发作、进展、发展、严重程度或复发。在一个方面,“治疗(treatment或treating)”包括部分缓解。在另一个方面,“治疗(treatment或treating)”包括完全缓解。
[0149]
使用可选方案(例如,“或”)应理解为是指可选方案中的一者、两者或其任何组合。如本文所用,不定冠词“一个/种(a/an)”应理解为是指任何所描述或列举的组分中的“一个/种或多个/种”。
[0150]
术语“约”或“基本上包括”是指在本领域普通技术人员确定的特定值或组成的可接受误差范围内的值或组成,其部分取决于如何测量或确定值或组成,即测量系统的局限性。例如,“约”或“基本上包括”可以指按照本领域实践在1个或超过1个标准偏差内。可选地,“约”或“基本上包括”可以指高达10%(即
±
10%)的范围。例如,约3mg可包括2.7mg和3.3mg之间(10%)的任何数量。此外,特别是关于生物系统或过程,这些术语可以指高达一个数量级或高达5倍的值。当在本技术和权利要求中提供特定值或组成时,除非另有说明,
否则应当假定“约”或“基本上包括”的含义在该特定值或组成的可接受的误差范围内。
[0151]
如本文所述,除非另有指示,否则任何浓度范围、百分比范围、比率范围或整数范围应理解为包括所述范围内的任何整数的值,并且在适当时包括其分数(如整数的十分之一和百分之一)。
[0152]
在以下小节中更详细地描述了本发明的各个方面。
[0153]
ii.本公开的方法
[0154]
本公开涉及鉴定mhc ii类特异性tcr的方法,所述方法包括使t细胞与包含(i)具有增强的cd4结合的hla ii类分子和(ii)肽(例如表位)的复合物接触。在某些方面,t细胞表达cd4。在某些方面,t细胞表达一种或多种tcr。在一些方面,mhc ii类特异性tcr特异性结合包含mhc ii类分子和肽的复合物。
[0155]
在一些方面,mhc ii类分子包含α链和β链,其中α链、β链或α链和β链两者包含相对于mhc ii类分子的野生型α链和/或β链具有一个或多个突变的氨基酸序列。在一些方面,α链包含相对于mhc ii类分子的野生型α链具有一个或多个突变的氨基酸序列。在一些方面,β链包含相对于mhc ii类分子的野生型β链具有一个或多个突变的氨基酸序列。在一些方面,α链包含相对于mhc ii类分子的野生型α链具有一个或多个突变的氨基酸序列,并且β链包含相对于mhc ii类分子的野生型β链具有一个或多个突变的氨基酸序列。
[0156]
在一些方面,一种或多种突变包括取代突变。在一些方面,一种或多种突变包括缺失突变。在一些方面,一种或多种突变包括插入突变。在一些方面,一种或多种突变包括用一种或多种异源氨基酸取代单个氨基酸。在一些方面,一种或多种突变包括用不同氨基酸取代单个氨基酸。在一些方面,一个或多个突变包括用2个不同氨基酸、3个不同氨基酸、4个不同氨基酸、5个不同氨基酸或多于5个不同氨基酸取代单个氨基酸。
[0157]
在一些方面,mhc ii类分子是二聚体。在一些方面,mhc ii类分子是三聚体。在一些方面,mhc ii类分子是四聚体。
[0158]
本公开的某些方面涉及富集从人受试者获得的靶标t细胞群的方法。在一些方面,所述方法包括使t细胞与本文公开的hla ii类分子接触。在一些方面,所述方法包括使t细胞与本文公开的细胞(例如)apc接触。在一些方面,在接触之后,相对于接触之前能够结合hla ii类分子的t细胞的数量,富集的t细胞群包含更多数量的能够结合hla ii类分子的t细胞。
[0159]
本公开的一些方面涉及选择能够靶向患病细胞(例如肿瘤细胞)的t细胞的方法。在一些方面,所述方法包括使分离的t细胞群在体外与包含本文公开的mhc ii类分子和多肽的片段的复合物接触,所述多肽为例如由患病细胞表达的抗原,例如肿瘤表达的多肽,例如,表位。
[0160]
在一些方面,本文公开的方法中使用的t细胞获自人受试者。从人受试者获得的t细胞可以是本文公开的任何t细胞。在一些方面,从人受试者获得的t细胞是肿瘤浸润淋巴细胞(til)。
[0161]
在一些方面,所述方法进一步包括选择由mhc ii类分子结合的t细胞。在一些方面,所述方法进一步包括向人受试者施用富集的t细胞。在一些方面,受试者在接受如本文所述的t细胞之前进行预调理。
[0162]
在一些方面,所述方法进一步包括分离与mhc ii类分子结合的tcr。在一些方面,
所述方法进一步包括对tcr进行测序。在一些方面,所述方法进一步包括克隆tcr。在一些方面,所述方法进一步包括在宿主细胞中重组表达tcr或其修饰变体。在一些方面,宿主细胞是免疫细胞,例如t细胞。在一些方面,所述方法进一步包括向受试者施用宿主细胞。在一些方面,受试者患有癌症,并且包含tcr的宿主细胞治疗受试者的癌症。
[0163]
ii.a.mhc ii类分子
[0164]
人白细胞抗原(hla)系统(人的主要组织相容性复合物[mhc])是免疫系统的重要组成部分,并且由位于6号染色体上的基因控制。它编码在t细胞上专门将抗原肽呈递给tcr的细胞表面分子。(另外参见overview of the immune system)。呈递抗原(ag)的mhc分子分为两大类:i类mhc分子和ii类mhc分子。
[0165]
ii类mhc分子作为跨膜糖蛋白存在于专职抗原呈递细胞(apc)的表面。完整的ii类分子由α链和β链组成。hla复合物中的三个基因座编码mhc ii类蛋白:hla-dp、hla-dq和hla-dr。表达cd4分子的t细胞与ii类mhc分子发生反应。这些淋巴细胞通常具有效应子功能和辅助功能,并激活反应以消除感染细胞内病原体的自身细胞或破坏细胞外寄生虫并帮助其他t细胞,如cd8 t细胞。因为只有专职apc表达ii类mhc分子,所以只有这些细胞为cd4 t细胞呈递抗原(cd4分别与mhc ii类分子的α链和β链的α-2和β-2结构域的非多态性部分结合)。
[0166]
在一些方面,hla ii类α和β链选自hla-dp、hla-dq和hla-dr等位基因。在某些方面,hla ii类β链是hla-dp等位基因。在某些方面,hla ii类α链是hla-dp等位基因。在某些方面,hla ii类β链是hla-dq等位基因。在某些方面,hla ii类α链是hla-dq等位基因。在某些方面,hla ii类β链是hla-dr等位基因。在某些方面,hla ii类α链是hla-dr等位基因。
[0167]
ii.a.1.hla-dp分子
[0168]
许多hla-dp等位基因在本领域中是已知的,并且任何已知的等位基因都可以用在本公开的方法中。hla-dpα链和β链等位基因的示例如表3所示。hla等位基因的更新清单可在hla.alleles.org/(上次访问时间为2019年2月27日)上获得。
[0169]
表3:dpβ链和α链氨基酸和核苷酸序列。
[0170]
[0171]
[0172]
[0173][0174]
ii.a.1.a.hla-dpβ链
[0175]
在某些方面,hla ii类分子包含dpβ链,其中所述dpβ链包含在对应于seq id no:1的氨基酸残基112的位置处的除亮氨酸之外的氨基酸。任何除亮氨酸之外的氨基酸可以存在于对应于seq id no:1的氨基酸残基112的位置处。在一些方面,除亮氨酸之外的氨基酸是包含疏水侧链的氨基酸。在某些方面,在对应于seq id no:1的氨基酸残基112的位置处的除亮氨酸之外的氨基酸是选自丙氨酸、缬氨酸、异亮氨酸、甲硫氨酸、苯丙氨酸、酪氨酸、和色氨酸的氨基酸。在某些方面,在对应于seq id no:1的氨基酸残基112的位置处的除亮氨酸之外的氨基酸是丙氨酸。在某些方面,在对应于seq id no:1的氨基酸残基112的位置处的除亮氨酸之外的氨基酸是缬氨酸。在某些方面,在对应于seq id no:1的氨基酸残基112的位置处的除亮氨酸之外的氨基酸是异亮氨酸。在某些方面,在对应于seq id no:1的氨基酸残基112的位置处的除亮氨酸之外的氨基酸是甲硫氨酸。在某些方面,在对应于seq id no:1的氨基酸残基112的位置处的除亮氨酸之外的氨基酸是苯丙氨酸。在某些方面,在对应于seq id no:1的氨基酸残基112的位置处的除亮氨酸之外的氨基酸是酪氨酸。在某些方面,在对应于seq id no:1的氨基酸残基112的位置处的除亮氨酸之外的氨基酸是色氨酸。
[0176]
在一些实施方案中,在对应于seq id no:1的氨基酸残基112的位置处的除亮氨酸之外的氨基酸由多于一个氨基酸组成,例如,两个氨基酸、三个氨基酸、四个氨基酸、五个氨基酸或者更多。在一些方面,多于一个氨基酸中的至少一个包含疏水侧链。在某些方面,在对应于seq id no:1的氨基酸残基112的位置处的除亮氨酸之外的氨基酸由一系列,例如至少2个、至少3个、至少4个或至少5个氨基酸组成,其中所述系列氨基酸中的每一个都包含疏水侧链。
[0177]
在某些方面,hla ii类分子包含dpβ链,其中所述dpβ链包含在对应于seq id no:1的氨基酸残基141的位置处的除缬氨酸之外的氨基酸。任何除缬氨酸之外的氨基酸都可存在于对应于seq id no:1的氨基酸残基141的位置处。在一些方面,除缬氨酸之外的氨基酸是包含疏水侧链的氨基酸。在某些方面,在对应于seq id no:1的氨基酸残基141的位置处
的除缬氨酸之外的氨基酸是选自丙氨酸、异亮氨酸、亮氨酸、甲硫氨酸、苯丙氨酸、酪氨酸、和色氨酸的氨基酸。在某些方面,在对应于seq id no:1的氨基酸残基141的位置处的除缬氨酸之外的氨基酸是丙氨酸。在某些方面,在对应于seq id no:1的氨基酸残基141的位置处的除缬氨酸之外的氨基酸是异亮氨酸。在某些方面,在对应于seq id no:1的氨基酸残基141的位置处的除缬氨酸之外的氨基酸是亮氨酸。在某些方面,在对应于seq id no:1的氨基酸残基141的位置处的除缬氨酸之外的氨基酸是甲硫氨酸。在某些方面,在对应于seq id no:1的氨基酸残基141的位置处的除缬氨酸之外的氨基酸是苯丙氨酸。在某些方面,在对应于seq id no:1的氨基酸残基141的位置处的除缬氨酸之外的氨基酸是酪氨酸。在某些方面,在对应于seq id no:1的氨基酸残基141的位置处的除缬氨酸之外的氨基酸是色氨酸。
[0178]
在一些方面,在对应于seq id no:1的氨基酸残基141的位置处的除缬氨酸之外的氨基酸由多于一个氨基酸组成,例如,两个氨基酸、三个氨基酸、四个氨基酸、五个氨基酸或者更多。在一些方面,多于一个氨基酸中的至少一个包含疏水侧链。在某些方面,在对应于seq id no:1的氨基酸残基141的位置处的除缬氨酸之外的氨基酸由一系列,例如至少2个、至少3个、至少4个或至少5个氨基酸组成,其中所述系列氨基酸中的每一个都包含疏水侧链。
[0179]
在本公开的某些方面,mhc ii类分子包含相对于野生型dpβ链含有多于一个取代突变的dpβ链。在某些方面,dpβ链相对于野生型dpβ链包含至少两个突变、至少三个突变、至少四个突变、至少五个突变、至少六个突变、至少七个突变、至少八个突变、至少九个突变、或至少十个突变。
[0180]
在某些方面,dpβ链包含在对应于seq id no:1的氨基酸残基112的位置处的除亮氨酸之外的氨基酸和在对应于seq id no:1的氨基酸残基141的位置处的除缬氨酸之外的氨基酸。在一些方面,(i)在对应于seq id no:1的氨基酸残基112的位置处的除亮氨酸之外的氨基酸,(ii)在对应于seq id no:1的氨基酸残基141的位置处的除缬氨酸之外的氨基酸,或者在对应于seq id no:1的氨基酸残基112位置处的除亮氨酸之外的氨基酸和在对应于seq id no:1的氨基酸残基141的位置处的除缬氨酸之外的氨基酸中的每一个是包含疏水侧链的氨基酸。在一些方面,(i)在对应于seq id no:1的氨基酸残基112的位置处的除亮氨酸之外的氨基酸选自丙氨酸、缬氨酸、异亮氨酸、甲硫氨酸、苯丙氨酸、酪氨酸和色氨酸;并且(ii)在对应于seq id no:1的氨基酸残基141的位置处的除缬氨酸之外的氨基酸选自丙氨酸、异亮氨酸、亮氨酸、甲硫氨酸、苯丙氨酸、酪氨酸和色氨酸。
[0181]
在一些方面,(i)在对应于seq id no:1的氨基酸残基112的位置处的除亮氨酸之外的氨基酸是色氨酸;并且(ii)在对应于seq id no:1的氨基酸残基141的位置处的除缬氨酸之外的氨基酸选自丙氨酸、异亮氨酸、亮氨酸、甲硫氨酸、苯丙氨酸、酪氨酸和色氨酸。在一些方面,(i)在对应于seq id no:1的氨基酸残基112的位置处的除亮氨酸之外的氨基酸选自丙氨酸、缬氨酸、异亮氨酸、甲硫氨酸、苯丙氨酸、酪氨酸和色氨酸;并且(ii)在对应于seq id no:1的氨基酸残基141的位置处的除缬氨酸之外的氨基酸是甲硫氨酸。在一些方面,(i)在对应于seq id no:1的氨基酸残基112的位置处的除亮氨酸之外的氨基酸是色氨酸;并且(ii)在对应于seq id no:1的氨基酸残基141的位置处的除缬氨酸之外的氨基酸是甲硫氨酸。
[0182]
在某些方面,dpβ链进一步包含在对应于seq id no:1的氨基酸残基114的位置处
的除缬氨酸之外的氨基酸。在一些方面,在对应于seq id no:1的氨基酸残基114的位置处的除缬氨酸之外的氨基酸选自丙氨酸、异亮氨酸、亮氨酸、甲硫氨酸、苯丙氨酸、酪氨酸、和色氨酸。在某些方面,在对应于seq id no:1的氨基酸残基114的位置处的除缬氨酸之外的氨基酸是甲硫氨酸。
[0183]
在某些方面,dpβ链进一步包含在对应于seq id no:1的氨基酸残基158的位置处的除甲硫氨酸之外的氨基酸。在一些方面,在对应于seq id no:1的氨基酸残基158的位置处的除甲硫氨酸之外的氨基酸选自丙氨酸、缬氨酸、异亮氨酸、甲硫氨酸、苯丙氨酸、酪氨酸、和色氨酸。在某些方面,在对应于seq id no:1的氨基酸残基158的位置处的除甲硫氨酸之外的氨基酸是异亮氨酸。
[0184]
在一些方面,dpβ链包含(i)在对应于seq id no:1的氨基酸残基112的位置处的除亮氨酸之外的氨基酸,和(ii)在对应于seq id no:1的氨基酸残基114的位置处的除缬氨酸之外的氨基酸。在一些方面,dpβ链包含(i)在对应于seq id no:1的氨基酸残基112的位置处的除亮氨酸之外的氨基酸,和(ii)在对应于seq id no:1的氨基酸残基158的位置处的除甲硫氨酸之外的氨基酸。
[0185]
在一些方面,dpβ链包含(i)在对应于seq id no:1的氨基酸残基112的位置处的色氨酸,和(ii)在对应于seq id no:1的氨基酸残基114的位置处的甲硫氨酸。在一些方面,dpβ链包含(i)在对应于seq id no:1的氨基酸残基112的位置处的色氨酸,和(ii)在对应于seq id no:1的氨基酸残基158的位置处的异亮氨酸。
[0186]
在一些方面,dpβ链包含(i)在对应于seq id no:1的氨基酸残基141的位置处的除缬氨酸之外的氨基酸,和(ii)在对应于seq id no:1的氨基酸残基114的位置处的除缬氨酸之外的氨基酸。在一些方面,dpβ链包含(i)在对应于seq id no:1的氨基酸残基141的位置处的除缬氨酸之外的氨基酸,和(ii)在对应于seq id no:1的氨基酸残基158的位置处的除甲硫氨酸之外的氨基酸。
[0187]
在一些方面,dpβ链包含(i)在对应于seq id no:1的氨基酸残基141的位置处的甲硫氨酸,和(ii)在对应于seq id no:1的氨基酸残基114的位置处的除缬氨酸之外的氨基酸。在一些方面,dpβ链包含(i)在对应于seq id no:1的氨基酸残基141的位置处的甲硫氨酸,和(ii)在对应于seq id no:1的氨基酸残基158的位置处的异亮氨酸。
[0188]
在一些方面,dpβ链包含(i)在对应于seq id no:1的氨基酸残基112的位置处的除亮氨酸之外的氨基酸,(ii)在对应于seq id no:1的氨基酸残基141的位置处的除缬氨酸之外的氨基酸,(iii)在对应于seq id no:1的氨基酸残基114的位置处的除缬氨酸之外的氨基酸,和(iv)在对应于seq id no:1的氨基酸残基158的位置处的除甲硫氨酸之外的氨基酸。
[0189]
在一些方面,dpβ链包含(i)在对应于seq id no:1的氨基酸残基112的位置处的色氨酸,(ii)在对应于seq id no:1的氨基酸残基141的位置处的甲硫氨酸,(iii)在对应于seq id no:1的氨基酸残基114的位置处的甲硫氨酸,和(iv)在对应于seq id no:1的氨基酸残基158的位置处的异亮氨酸。
[0190]
在一些方面,dpβ链包含在对应于seq id no:1的氨基酸残基114的位置处的缬氨酸。在一些方面,dpβ链包含在对应于seq id no:1的氨基酸残基158的位置处的甲硫氨酸。在一些方面,dpβ链包含(i)在对应于seq id no:1的氨基酸残基112的位置处的除亮氨酸之
外的氨基酸,(ii)在对应于seq id no:1的氨基酸残基141的位置处的除缬氨酸之外的氨基酸,和(iii)在对应于seq id no:1的氨基酸残基114的位置处的缬氨酸。在一些方面,dpβ链包含(i)在对应于seq id no:1的氨基酸残基112的位置处的除亮氨酸之外的氨基酸,(ii)在对应于seq id no:1的氨基酸残基141的位置处的除缬氨酸之外的氨基酸,和(iii)在对应于seq id no:1的对应于氨基酸残基158的位置处的甲硫氨酸。在一些方面,dpβ链包含(i)在对应于seq id no:1的氨基酸残基112的位置处的除亮氨酸之外的氨基酸,(ii)在对应于seq id no:1的氨基酸残基141的位置处的除缬氨酸之外的氨基酸,(iii)在对应于seq id no:1的氨基酸残基114的位置处的缬氨酸,和(iv)在对应于seq id no:1的对应于氨基酸残基158的位置处的甲硫氨酸。
[0191]
在一些方面,dpβ链包含(i)在对应于seq id no:1的氨基酸残基112的位置处的色氨酸,(ii)在对应于seq id no:1的氨基酸残基141的位置处的甲硫氨酸,和(iii)在对应于seq id no:1的氨基酸残基114的位置处的缬氨酸。在一些方面,dpβ链包含(i)在对应于seq id no:1的氨基酸残基112的位置处的色氨酸,(ii)在对应于seq id no:1的氨基酸残基141的位置处的甲硫氨酸,和(iii)在对应于seq id no:1的对应于氨基酸残基158的位置处的甲硫氨酸。在一些方面,dpβ链包含(i)在对应于seq id no:1的氨基酸残基112的位置处的色氨酸,(ii)在对应于seq id no:1的氨基酸残基141的位置处的甲硫氨酸,(iii)在对应于seq id no:1的氨基酸残基114的位置处的缬氨酸,和(iv)在对应于seq id no:1的氨基酸残基158的位置处的甲硫氨酸。
[0192]
在一些方面,dpβ链包含(i)在对应于seq id no:1的氨基酸残基112的位置处的色氨酸,(ii)在对应于seq id no:1的氨基酸残基141的位置处的甲硫氨酸,(iii)在对应于seq id no:1的氨基酸残基114的位置处的甲硫氨酸,和(iv)在对应于seq id no:1的氨基酸残基158的位置处的异亮氨酸。
[0193]
在某些方面,与参考hla ii类分子相比,本文所述的dpβ链对cd4蛋白具有增加的亲和力。在一些方面,参考hla ii类分子是具有野生型dpβ链的hla ii类分子。在一些方面,参考hla ii类分子是具有dpβ链的hla ii类分子,所述dpβ链包含(i)在对应于seq id no:1的氨基酸残基112的位置处的亮氨酸和/或(ii)在对应于seq id no:1的氨基酸残基141的位置处的缬氨酸。
[0194]
在一些方面,增加的对cd4的亲和力是参考hla ii类分子对cd4的亲和力的至少约1.5倍、至少约2倍、至少约3倍、至少约4倍、至少约5倍、至少约6倍、至少约7倍、至少约8倍、至少约9倍、至少约10倍、至少约15倍、至少约20倍、至少约25倍、至少约30倍、至少约35倍、至少约40倍、至少约45倍、至少约50倍、至少约75倍、至少约100倍、至少约200倍、至少约300倍、至少约400倍、至少约500倍、至少约1000倍、至少约1500倍、至少约2000倍、至少约2500倍、至少约3000倍、至少约3500倍、至少约4000倍、至少约4500倍、或至少约4000倍。
[0195]
在一些方面,增加的对cd4的亲和力是参考hla ii类分子对cd4的亲和力的至少约1.5倍到至少约5000倍、1.5倍到至少约4000倍、1.5倍到至少约3000倍、1.5倍到至少约2000倍、1.5倍到至少约1000倍、10倍到至少约5000倍、10倍到至少约4000倍、10倍到至少约3000倍、10倍到至少约2000倍、10倍到至少约1000倍、10倍到至少约900倍、10倍到至少约800倍、10倍到至少约700倍、10倍到至少约600倍、10倍到至少约500倍、10倍到至少约400倍、10倍到至少约300倍、10倍到至少约200倍、10倍到至少约100倍、100倍到至少约5000倍、100倍到
至少约4000倍、100倍到至少约3000倍、100倍到至少约2000倍、100倍到至少约1000倍、100倍到至少约900倍、100倍到至少约800倍、100倍到至少约700倍、100倍到至少约600倍、100倍到至少约500倍、100倍到至少约400倍、100倍到至少约300倍、或100倍到至少约200倍。
[0196]
在某些方面,dpβ链包含选自以下的等位基因:dpb1*01、dpb1*02、dpb1*03、dpb1*04、dpb1*05、dpb1*06、dpb1*08、dpb1*09、dpb1*10、dpb1*100、dpb1*101、dpb1*102、dpb1*103、dpb1*104、dpb1*105、dpb1*106、dpb1*107、dpb1*108、dpb1*109、dpb1*11、dpb1*110、dpb1*111、dpb1*112、dpb1*113、dpb1*114、dpb1*115、dpb1*116、dpb1*117、dpb1*118、dpb1*119、dpb1*120、dpb1*121、dpb1*122、dpb1*123、dpb1*124、dpb1*125、dpb1*126、dpb1*127、dpb1*128、dpb1*129、dpb1*13、dpb1*130、dpb1*131、dpb1*132、dpb1*133、dpb1*134、dpb1*135、dpb1*136、dpb1*137、dpb1*138、dpb1*139、dpb1*14、dpb1*140、dpb1*141、dpb1*142、dpb1*143、dpb1*144、dpb1*145、dpb1*146、dpb1*147、dpb1*148、dpb1*149、dpb1*15、dpb1*150、dpb1*151、dpb1*152、dpb1*153、dpb1*154、dpb1*155、dpb1*156、dpb1*157、dpb1*158、dpb1*159、dpb1*16、dpb1*160、dpb1*161、dpb1*162、dpb1*163、dpb1*164、dpb1*165、dpb1*166、dpb1*167、dpb1*168、dpb1*169、dpb1*17、dpb1*170、dpb1*171、dpb1*172、dpb1*173、dpb1*174、dpb1*175、dpb1*176、dpb1*177、dpb1*178、dpb1*179、dpb1*18、dpb1*180、dpb1*181、dpb1*182、dpb1*183、dpb1*184、dpb1*185、dpb1*186、dpb1*187、dpb1*188、dpb1*189、dpb1*19、dpb1*190、dpb1*191、dpb1*192、dpb1*193、dpb1*194、dpb1*195、dpb1*196、dpb1*197、dpb1*198、dpb1*199、dpb1*20、dpb1*200、dpb1*201、dpb1*202、dpb1*203、dpb1*204、dpb1*205、dpb1*206、dpb1*207、dpb1*208、dpb1*209、dpb1*21、dpb1*210、dpb1*211、dpb1*212、dpb1*213、dpb1*214、dpb1*215、dpb1*216、dpb1*217、dpb1*218、dpb1*219、dpb1*22、dpb1*220、dpb1*221、dpb1*222、dpb1*223、dpb1*224、dpb1*225、dpb1*226、dpb1*227、dpb1*228、dpb1*229、dpb1*23、dpb1*230、dpb1*231、dpb1*232、dpb1*233、dpb1*234、dpb1*235、dpb1*236、dpb1*237、dpb1*238、dpb1*239、dpb1*24、dpb1*240、dpb1*241、dpb1*242、dpb1*243、dpb1*244、dpb1*245、dpb1*246、dpb1*247、dpb1*248、dpb1*249、dpb1*25、dpb1*250、dpb1*251、dpb1*252、dpb1*253、dpb1*254、dpb1*255、dpb1*256、dpb1*257、dpb1*258、dpb1*259、dpb1*26、dpb1*260、dpb1*261、dpb1*262、dpb1*263、dpb1*264、dpb1*265、dpb1*266、dpb1*267、dpb1*268、dpb1*269、dpb1*27、dpb1*270、dpb1*271、dpb1*272、dpb1*273、dpb1*274、dpb1*275、dpb1*276、dpb1*277、dpb1*278、dpb1*279、dpb1*28、dpb1*280、dpb1*281、dpb1*282、dpb1*283、dpb1*284、dpb1*285、dpb1*286、dpb1*287、dpb1*288、dpb1*289、dpb1*29、dpb1*290、dpb1*291、dpb1*292、dpb1*293、dpb1*294、dpb1*295、dpb1*296、dpb1*297、dpb1*298、dpb1*299、dpb1*30、dpb1*300、dpb1*301、dpb1*302、dpb1*303、dpb1*304、dpb1*305、dpb1*306、dpb1*307、dpb1*308、dpb1*309、dpb1*31、dpb1*310、dpb1*311、dpb1*312、dpb1*313、dpb1*314、dpb1*315、dpb1*316、dpb1*317、dpb1*318、dpb1*319、dpb1*32、dpb1*320、dpb1*321、dpb1*322、dpb1*323、dpb1*324、dpb1*325、dpb1*326、dpb1*327、dpb1*328、dpb1*329、dpb1*33、dpb1*330、dpb1*331、dpb1*332、dpb1*333、dpb1*334、dpb1*335、dpb1*336、dpb1*337、dpb1*338、dpb1*339、dpb1*34、dpb1*340、dpb1*341、dpb1*342、dpb1*343、dpb1*344、dpb1*345、dpb1*346、dpb1*347、dpb1*348、dpb1*349、dpb1*35、dpb1*350、dpb1*351、dpb1*352、dpb1*353、dpb1*354、dpb1*355、dpb1*356、dpb1*357、dpb1*358、dpb1*359、dpb1*36、dpb1*360、dpb1*361、dpb1*362、dpb1*363、dpb1*364、dpb1*365、dpb1*366、dpb1*
367、dpb1*368、dpb1*369、dpb1*37、dpb1*370、dpb1*371、dpb1*372、dpb1*373、dpb1*374、dpb1*375、dpb1*376、dpb1*377、dpb1*378、dpb1*379、dpb1*38、dpb1*380、dpb1*381、dpb1*382、dpb1*383、dpb1*384、dpb1*385、dpb1*386、dpb1*387、dpb1*388、dpb1*389、dpb1*39、dpb1*390、dpb1*391、dpb1*392、dpb1*393、dpb1*394、dpb1*395、dpb1*396、dpb1*397、dpb1*398、dpb1*399、dpb1*40、dpb1*400、dpb1*401、dpb1*402、dpb1*403、dpb1*404、dpb1*405、dpb1*406、dpb1*407、dpb1*408、dpb1*409、dpb1*41、dpb1*410、dpb1*411、dpb1*412、dpb1*413、dpb1*414、dpb1*415、dpb1*416、dpb1*417、dpb1*418、dpb1*419、dpb1*420、dpb1*421、dpb1*422、dpb1*423、dpb1*424、dpb1*425、dpb1*426、dpb1*427、dpb1*428、dpb1*429、dpb1*430、dpb1*431、dpb1*432、dpb1*433、dpb1*434、dpb1*435、dpb1*436、dpb1*437、dpb1*438、dpb1*439、dpb1*44、dpb1*440、dpb1*441、dpb1*442、dpb1*443、dpb1*444、dpb1*445、dpb1*446、dpb1*447、dpb1*448、dpb1*449、dpb1*45、dpb1*450、dpb1*451、dpb1*452、dpb1*453、dpb1*454、dpb1*455、dpb1*456、dpb1*457、dpb1*458、dpb1*459、dpb1*46、dpb1*460、dpb1*461、dpb1*462、dpb1*463、dpb1*464、dpb1*465、dpb1*466、dpb1*467、dpb1*468、dpb1*469、dpb1*47、dpb1*470、dpb1*471、dpb1*472、dpb1*473、dpb1*474、dpb1*475、dpb1*476、dpb1*477、dpb1*478、dpb1*479、dpb1*48、dpb1*480、dpb1*481、dpb1*482、dpb1*483、dpb1*484、dpb1*485、dpb1*486、dpb1*487、dpb1*488、dpb1*489、dpb1*49、dpb1*490、dpb1*491、dpb1*492、dpb1*493、dpb1*494、dpb1*495、dpb1*496、dpb1*497、dpb1*498、dpb1*499、dpb1*50、dpb1*500、dpb1*501、dpb1*502、dpb1*503、dpb1*504、dpb1*505、dpb1*506、dpb1*507、dpb1*508、dpb1*509、dpb1*51、dpb1*510、dpb1*511、dpb1*512、dpb1*513、dpb1*514、dpb1*515、dpb1*516、dpb1*517、dpb1*518、dpb1*519、dpb1*52、dpb1*520、dpb1*521、dpb1*522、dpb1*523、dpb1*524、dpb1*525、dpb1*526、dpb1*527、dpb1*528、dpb1*529、dpb1*53、dpb1*530、dpb1*531、dpb1*532、dpb1*533、dpb1*534、dpb1*535、dpb1*536、dpb1*537、dpb1*538、dpb1*539、dpb1*54、dpb1*540、dpb1*541、dpb1*542、dpb1*543、dpb1*544、dpb1*545、dpb1*546、dpb1*547、dpb1*548、dpb1*549、dpb1*55、dpb1*550、dpb1*551、dpb1*552、dpb1*553、dpb1*554、dpb1*555、dpb1*556、dpb1*557、dpb1*558、dpb1*559、dpb1*56、dpb1*560、dpb1*561、dpb1*562、dpb1*563、dpb1*564、dpb1*565、dpb1*566、dpb1*567、dpb1*568、dpb1*569、dpb1*57、dpb1*570、dpb1*571、dpb1*572、dpb1*573、dpb1*574、dpb1*575、dpb1*576、dpb1*577、dpb1*578、dpb1*579、dpb1*58、dpb1*580、dpb1*581、dpb1*582、dpb1*583、dpb1*584、dpb1*585、dpb1*586、dpb1*587、dpb1*588、dpb1*589、dpb1*59、dpb1*590、dpb1*591、dpb1*592、dpb1*593、dpb1*594、dpb1*595、dpb1*596、dpb1*597、dpb1*598、dpb1*599、dpb1*60、dpb1*600、dpb1*601、dpb1*602、dpb1*603、dpb1*604、dpb1*605、dpb1*606、dpb1*607、dpb1*608、dpb1*609、dpb1*61、dpb1*610、dpb1*611、dpb1*612、dpb1*613、dpb1*614、dpb1*615、dpb1*616、dpb1*617、dpb1*618、dpb1*619、dpb1*62、dpb1*620、dpb1*621、dpb1*622、dpb1*623、dpb1*624、dpb1*625、dpb1*626、dpb1*627、dpb1*628、dpb1*629、dpb1*63、dpb1*630、dpb1*631、dpb1*632、dpb1*633、dpb1*634、dpb1*635、dpb1*636、dpb1*637、dpb1*638、dpb1*639、dpb1*64、dpb1*640、dpb1*641、dpb1*642、dpb1*643、dpb1*644、dpb1*645、dpb1*646、dpb1*647、dpb1*648、dpb1*649、dpb1*65、dpb1*650、dpb1*651、dpb1*652、dpb1*653、dpb1*654、dpb1*655、dpb1*656、dpb1*657、dpb1*658、dpb1*659、dpb1*66、dpb1*660、dpb1*661、dpb1*662、dpb1*663、dpb1*664、dpb1*665、dpb1*666、dpb1*667、dpb1*668、dpb1*669、dpb1*67、
dpb1*670、dpb1*671、dpb1*672、dpb1*673、dpb1*674、dpb1*675、dpb1*676、dpb1*677、dpb1*678、dpb1*679、dpb1*68、dpb1*680、dpb1*681、dpb1*682、dpb1*683、dpb1*684、dpb1*685、dpb1*686、dpb1*687、dpb1*688、dpb1*689、dpb1*69、dpb1*690、dpb1*691、dpb1*692、dpb1*693、dpb1*694、dpb1*695、dpb1*696、dpb1*697、dpb1*698、dpb1*699、dpb1*70、dpb1*700、dpb1*701、dpb1*702、dpb1*703、dpb1*704、dpb1*705、dpb1*706、dpb1*707、dpb1*708、dpb1*709、dpb1*71、dpb1*710、dpb1*711、dpb1*712、dpb1*713、dpb1*714、dpb1*715、dpb1*716、dpb1*717、dpb1*718、dpb1*719、dpb1*72、dpb1*720、dpb1*721、dpb1*722、dpb1*723、dpb1*724、dpb1*725、dpb1*726、dpb1*727、dpb1*728、dpb1*729、dpb1*73、dpb1*730、dpb1*731、dpb1*732、dpb1*733、dpb1*734、dpb1*735、dpb1*736、dpb1*737、dpb1*738、dpb1*739、dpb1*74、dpb1*740、dpb1*741、dpb1*742、dpb1*743、dpb1*744、dpb1*745、dpb1*746、dpb1*747、dpb1*748、dpb1*749、dpb1*75、dpb1*750、dpb1*751、dpb1*752、dpb1*753、dpb1*754、dpb1*755、dpb1*756、dpb1*757、dpb1*758、dpb1*759、dpb1*76、dpb1*760、dpb1*761、dpb1*762、dpb1*763、dpb1*764、dpb1*765、dpb1*766、dpb1*767、dpb1*768、dpb1*769、dpb1*77、dpb1*770、dpb1*771、dpb1*772、dpb1*773、dpb1*774、dpb1*775、dpb1*776、dpb1*777、dpb1*778、dpb1*779、dpb1*78、dpb1*780、dpb1*781、dpb1*782、dpb1*783、dpb1*784、dpb1*785、dpb1*786、dpb1*787、dpb1*788、dpb1*789、dpb1*79、dpb1*790、dpb1*791、dpb1*792、dpb1*794、dpb1*795、dpb1*796、dpb1*797、dpb1*798、dpb1*799、dpb1*80、dpb1*800、dpb1*801、dpb1*802、dpb1*803、dpb1*804、dpb1*805、dpb1*806、dpb1*807、dpb1*808、dpb1*809、dpb1*81、dpb1*810、dpb1*811、dpb1*812、dpb1*813、dpb1*814、dpb1*815、dpb1*816、dpb1*817、dpb1*818、dpb1*819、dpb1*82、dpb1*820、dpb1*821、dpb1*822、dpb1*823、dpb1*824、dpb1*825、dpb1*826、dpb1*827、dpb1*828、dpb1*829、dpb1*83、dpb1*830、dpb1*831、dpb1*832、dpb1*833、dpb1*834、dpb1*835、dpb1*836、dpb1*837、dpb1*838、dpb1*839、dpb1*84、dpb1*840、dpb1*841、dpb1*842、dpb1*843、dpb1*844、dpb1*845、dpb1*846、dpb1*847、dpb1*848、dpb1*849、dpb1*85、dpb1*850、dpb1*851、dpb1*852、dpb1*853、dpb1*854、dpb1*855、dpb1*856、dpb1*857、dpb1*858、dpb1*859、dpb1*86、dpb1*860、dpb1*861、dpb1*862、dpb1*863、dpb1*864、dpb1*865、dpb1*866、dpb1*867、dpb1*868、dpb1*869、dpb1*87、dpb1*870、dpb1*871、dpb1*872、dpb1*873、dpb1*874、dpb1*875、dpb1*876、dpb1*877、dpb1*878、dpb1*879、dpb1*88、dpb1*880、dpb1*881、dpb1*882、dpb1*883、dpb1*884、dpb1*885、dpb1*886、dpb1*887、dpb1*888、dpb1*889、dpb1*89、dpb1*890、dpb1*891、dpb1*892、dpb1*893、dpb1*894、dpb1*895、dpb1*896、dpb1*897、dpb1*898、dpb1*899、dpb1*90、dpb1*900、dpb1*901、dpb1*902、dpb1*903、dpb1*904、dpb1*905、dpb1*906、dpb1*907、dpb1*908、dpb1*909、dpb1*91、dpb1*910、dpb1*911、dpb1*912、dpb1*913、dpb1*914、dpb1*915、dpb1*916、dpb1*917、dpb1*918、dpb1*919、dpb1*92、dpb1*920、dpb1*921、dpb1*922、dpb1*923、dpb1*924、dpb1*925、dpb1*926、dpb1*927、dpb1*928、dpb1*929、dpb1*93、dpb1*930、dpb1*931、dpb1*932、dpb1*933、dpb1*934、dpb1*935、dpb1*936、dpb1*937、dpb1*938、dpb1*939、dpb1*94、dpb1*940、dpb1*941、dpb1*942、dpb1*943、dpb1*944、dpb1*945、dpb1*946、dpb1*947、dpb1*948、dpb1*949、dpb1*95、dpb1*950、dpb1*951、dpb1*952、dpb1*953、dpb1*954、dpb1*955、dpb1*956、dpb1*957、dpb1*958、dpb1*959、dpb1*96、dpb1*960、dpb1*961、dpb1*962、dpb1*963、dpb1*964、dpb1*965、dpb1*97、dpb1*98、和dpb1*99。在一些方面,dpβ链包含hla-dpb1*01、hla-dpb1*
02、hla-dpb1*03、hla-dpb1*04、hla-dpb1*05、hla-dpb1*06、hla-dpb1*08、或hla-dpb1*09等位基因。在某些方面,dpβ链包含hla-dpb1*04等位基因。在特定方面,dpβ链包含hla-dpb1*04:01等位基因。
[0197]
在某些方面,mhc ii类分子包含dpβ链,所述dpβ链包含与seq id no:3具有至少约70%、至少约75%、至少约80%、至少约85%、至少约90%、至少约95%、至少约96%、至少约97%、至少约98%、至少约99%或约100%序列同一性的氨基酸序列,其中所述dpβ链包含在对应于seq id no:1的氨基酸残基112的位置处的色氨酸,并且其中所述dpβ链包含在对应于seq id no:1的氨基酸残基141的位置处的甲硫氨酸。在某些方面,mhc ii类分子包含dpβ链,所述dpβ链包含与seq id no:3具有至少约70%、至少约75%、至少约80%、至少约85%、至少约90%、至少约95%、至少约96%、至少约97%、至少约98%、至少约99%或约100%序列同一性的氨基酸序列,其中所述dpβ链包含(i)在对应于seq id no:1的氨基酸残基112的位置处的色氨酸,(ii)在对应于seq id no:1的氨基酸残基141的位置处的甲硫氨酸,(iii)在对应于seq id no:1的氨基酸残基114的位置处的缬氨酸,和(iv)在对应于seq id no:1的对应于氨基酸残基158的位置处的甲硫氨酸。在某些方面,mhc ii类分子包含dpβ链,所述dpβ链包含seq id no:3所示的氨基酸序列。
[0198]
ii.a.1.a.hla-dpα链
[0199]
在本公开的一些方面,mhc ii类分子还包含α链。在一些方面,α链是野生型α链。在一些方面,α链是dpα链。可以在本公开的组合物和方法中使用任何dpα链。在一些方面,dpα链包含hla-dpa1*01、hla-dpa1*02、hla-dpa1*03、或hla-dpa1*04等位基因。在某些方面,dpα链包含hla-dpa1*01等位基因。在某些方面,dpα链包含hla-dpa1*02等位基因。在某些方面,dpα链包含hla-dpa1*03等位基因。在某些方面,dpα链包含hla-dpa1*04等位基因。
[0200]
在某些方面,dpα链选自dpa1*01:03:01:01、dpa1*01:03:01:02、dpa1*01:03:01:03、dpa1*01:03:01:04、dpa1*01:03:01:05、dpa1*01:03:01:06、dpa1*01:03:01:07、dpa1*01:03:01:08、dpa1*01:03:01:09、dpa1*01:03:01:10、dpa1*01:03:01:11、dpa1*01:03:01:12、dpa1*01:03:01:13、dpa1*01:03:01:14、dpa1*01:03:01:15、dpa1*01:03:01:16、dpa1*01:03:01:17、dpa1*01:03:01:18q、dpa1*01:03:01:19、dpa1*01:03:01:20、dpa1*01:03:01:21、dpa1*01:03:01:22、dpa1*01:03:01:23、dpa1*01:03:02、dpa1*01:03:03、dpa1*01:03:04、dpa1*01:03:05、dpa1*01:03:06、dpa1*01:03:07、dpa1*01:03:08、dpa1*01:03:09、dpa1*01:04、dpa1*01:05、dpa1*01:06:01、dpa1*01:06:02、dpa1*01:07、dpa1*01:08、dpa1*01:09、dpa1*01:10、dpa1*01:11、dpa1*01:12、dpa1*01:13、dpa1*01:14、dpa1*01:15、dpa1*01:16、dpa1*01:17、dpa1*01:18、dpa1*01:19、dpa1*02:01:01:01、dpa1*02:01:01:02、dpa1*02:01:01:03、dpa1*02:01:01:04、dpa1*02:01:01:05、dpa1*02:01:01:06、dpa1*02:01:01:07、dpa1*02:01:01:08、dpa1*02:01:01:09、dpa1*02:01:01:10、dpa1*02:01:01:11、dpa1*02:01:02:01、dpa1*02:01:02:02、dpa1*02:01:03、dpa1*02:01:04、dpa1*02:01:05、dpa1*02:01:06、d pa1*02:01:07、dpa1*02:01:08:01、dpa1*02:01:08:02、dpa1*02:02:02:01、dpa1*02:02:02:02、dpa1*02:02:02:03、dpa1*02:02:02:04、dpa1*02:02:02:05、dpa1*02:02:03、dpa1*02:02:04、dpa1*02:02:05、dpa1*02:02:06、dpa1*02:03、dpa1*02:04、dpa1*02:05、dpa1*02:06、dpa1*02:07:01:01、dpa1*02:07:01:02、dpa1*02:07:01:03、dpa1*02:08、dpa1*02:09、dpa1*02:10、dpa1*02:11、dpa1*02:12、dpa1*02:13n、dpa1*02:
14、dpa1*02:15、dpa1*02:16、dpa1*03:01:01:01、dpa1*03:01:01:02、dpa1*03:01:01:03、dpa1*03:01:01:04、dpa1*03:01:01:05、dpa1*03:01:02、dpa1*03:02、dpa1*03:03、dpa1*03:04、dpa1*04:01:01:01、dpa1*04:01:01:02、dpa1*04:01:01:03、dpa1*04:02、或其任何组合。
[0201]
在某些方面,mhc ii类分子包含dpα链,所述dpα链包含与seq id no:6具有至少约70%、至少约75%、至少约80%、至少约85%、至少约90%、至少约95%、至少约96%、至少约97%、至少约98%、至少约99%或约100%序列同一性的氨基酸序列。在某些方面,mhc类ii分子包含dpα链,所述dpα链包含与seq id no:8具有至少约70%、至少约75%、至少约80%、至少约85%、至少约90%、至少约95%、至少约96%、至少约97%、至少约98%、至少约99%或约100%序列同一性的氨基酸序列。在某些方面,mhc ii类分子包含dpα链,所述dpα链包含seq id no:6所示的氨基酸序列。在某些方面,mhc ii类分子包含dpα链,所述dpα链包含seq id no:8所示的氨基酸序列。
[0202]
ii.a.2.hla-dq分子
[0203]
许多hla-dq等位基因在本领域中是已知的,并且任何已知的等位基因都可以用在本公开中。hla-dqα链和β链等位基因的示例如表4所示。hla等位基因的更新清单可在hla.alleles.org/(上次访问时间为2019年7月10)上获得。
[0204]
表4:dqβ链和α链氨基酸和核苷酸序列。
[0205]
[0206][0207]
ii.a.2.a.hla-dqβ链
[0208]
在某些方面,hla ii类分子包含dqβ链,其中所述dqβ链包含在对应于seq id no:11的氨基酸残基114的位置处的除亮氨酸之外的氨基酸。任何除亮氨酸之外的氨基酸都可以存在于对应于seq id no:11的氨基酸残基114的位置处。在一些方面,除亮氨酸之外的氨基酸是包含疏水侧链的氨基酸。在某些方面,在对应于seq id no:11的氨基酸残基114的位置处的除亮氨酸之外的氨基酸是选自丙氨酸、缬氨酸、异亮氨酸、甲硫氨酸、苯丙氨酸、酪氨酸、和色氨酸的氨基酸。在某些方面,在对应于seq id no:11的氨基酸残基114的位置处的除亮氨酸之外的氨基酸是丙氨酸。在某些方面,在对应于seq id no:11的氨基酸残基114的位置处的除亮氨酸之外的氨基酸是缬氨酸。在某些方面,在对应于seq id no:11的氨基酸
残基114的位置处的除亮氨酸之外的氨基酸是异亮氨酸。在某些方面,在对应于seq id no:11的氨基酸残基114的位置处的除亮氨酸之外的氨基酸是甲硫氨酸。在某些方面,在对应于seq id no:11的氨基酸残基114的位置处的除亮氨酸之外的氨基酸是苯丙氨酸。在某些方面,在对应于seq id no:11的氨基酸残基114的位置处的除亮氨酸之外的氨基酸是酪氨酸。在某些方面,在对应于seq id no:11的氨基酸残基114的位置处的除亮氨酸之外的氨基酸是色氨酸。
[0209]
在一些实施方案中,在对应于seq id no:11的氨基酸残基114的位置处的除亮氨酸之外的氨基酸由多于一个氨基酸组成,例如,两个氨基酸、三个氨基酸、四个氨基酸、五个氨基酸或者更多。在一些方面,多于一个氨基酸中的至少一个包含疏水侧链。在某些方面,在对应于seq id no:11的氨基酸残基114的位置处的除亮氨酸之外的氨基酸由一系列,例如至少2个、至少3个、至少4个或至少5个氨基酸组成,其中所述系列氨基酸中的每一个都包含疏水侧链。
[0210]
在某些方面,hla ii类分子包含dqβ链,其中所述dqβ链包含在对应于seq id no:11的氨基酸残基143的位置处的除缬氨酸之外的氨基酸。任何除缬氨酸之外的氨基酸都可存在于对应于seq id no:11的氨基酸残基143的位置处。在一些方面,除缬氨酸之外的氨基酸是包含疏水侧链的氨基酸。在某些方面,在对应于seq id no:11的氨基酸残基143的位置处的除缬氨酸之外的氨基酸是选自丙氨酸、异亮氨酸、亮氨酸、甲硫氨酸、苯丙氨酸、酪氨酸、和色氨酸的氨基酸。在某些方面,在对应于seq id no:11的氨基酸残基143的位置处的除缬氨酸之外的氨基酸是丙氨酸。在某些方面,在对应于seq id no:11的氨基酸残基143的位置处的除缬氨酸之外的氨基酸是异亮氨酸。在某些方面,在对应于seq id no:11的氨基酸残基143的位置处的除缬氨酸之外的氨基酸是亮氨酸。在某些方面,在对应于seq id no:11的氨基酸残基143的位置处的除缬氨酸之外的氨基酸是甲硫氨酸。在某些方面,在对应于seq id no:11的氨基酸残基143的位置处的除缬氨酸之外的氨基酸是苯丙氨酸。在某些方面,在对应于seq id no:11的氨基酸残基143的位置处的除缬氨酸之外的氨基酸是酪氨酸。在某些方面,在对应于seq id no:11的氨基酸残基143的位置处的除缬氨酸之外的氨基酸是色氨酸。
[0211]
在一些方面,在对应于seq id no:11的氨基酸残基143的位置处的除缬氨酸之外的氨基酸由多于一个氨基酸组成,例如,两个氨基酸、三个氨基酸、四个氨基酸、五个氨基酸或者更多。在一些方面,多于一个氨基酸中的至少一个包含疏水侧链。在某些方面,在对应于seq id no:11的氨基酸残基143的位置处的除缬氨酸之外的氨基酸由一系列,例如至少2个、至少3个、至少4个或至少5个氨基酸组成,其中所述系列氨基酸中的每一个都包含疏水侧链。
[0212]
在某些方面,hla ii类分子包含dqβ链,其中所述dqβ链包含在对应于seq id no:11的氨基酸残基110的位置处的除天冬酰胺之外的氨基酸。任何除天冬酰胺之外的氨基酸都可以存在于对应于seq id no:11的氨基酸残基110的位置处。在一些方面,除天冬酰胺之外的氨基酸是包含极性不带电侧链的氨基酸。在某些方面,在对应于seq id no:11的氨基酸残基110的位置处的除天冬酰胺之外的氨基酸是选自丝氨酸、苏氨酸和谷氨酰胺的氨基酸。在某些方面,在对应于seq id no:11的氨基酸残基110的位置处的除天冬酰胺之外的氨基酸是丝氨酸。在某些方面,在对应于seq id no:11的氨基酸残基110的位置处的除天冬酰
胺之外的氨基酸是苏氨酸。在某些方面,在对应于seq id no:11的氨基酸残基110的位置处的除天冬酰胺之外的氨基酸是谷氨酰胺。
[0213]
在一些方面,在对应于seq id no:11的氨基酸残基110的位置处的除天冬酰胺之外的氨基酸由多于一个氨基酸组成,例如,两个氨基酸、三个氨基酸、四个氨基酸、五个氨基酸或者更多。在一些方面,多于一个氨基酸中的至少一个包含极性不带电侧链。在某些方面,在对应于seq id no:11的氨基酸残基110的位置处的除天冬酰胺之外的氨基酸由一系列,例如至少2个、至少3个、至少4个或至少5个氨基酸组成,其中所述系列氨基酸中的每一个都包含极性不带电侧链。
[0214]
在某些方面,hla ii类分子包含dqβ链,其中所述dqβ链包含在对应于seq id no:11的氨基酸残基116的位置处的除异亮氨酸之外的氨基酸。任何除异亮氨酸之外的氨基酸都可以存在于对应于seq id no:11的氨基酸残基116的位置处。在一些方面,除异亮氨酸之外的氨基酸是包含疏水侧链的氨基酸。在某些方面,在对应于seq id no:11的氨基酸残基116的位置处的除异亮氨酸之外的氨基酸是选自丙氨酸、缬氨酸、亮氨酸、甲硫氨酸、苯丙氨酸、酪氨酸、和色氨酸的氨基酸。在某些方面,在对应于seq id no:11的氨基酸残基116的位置处的除异亮氨酸之外的氨基酸是丙氨酸。在某些方面,在对应于seq id no:11的氨基酸残基116的位置处的除异亮氨酸之外的氨基酸是缬氨酸。在某些方面,在对应于seq id no:11的氨基酸残基116的位置处的除异亮氨酸之外的氨基酸是亮氨酸。在某些方面,在对应于seq id no:11的氨基酸残基116的位置处的除异亮氨酸之外的氨基酸是甲硫氨酸。在某些方面,在对应于seq id no:11的氨基酸残基116的位置处的除异亮氨酸之外的氨基酸是苯丙氨酸。在某些方面,在对应于seq id no:11的氨基酸残基116的位置处的除异亮氨酸之外的氨基酸是酪氨酸。在某些方面,在对应于seq id no:11的氨基酸残基116的位置处的除异亮氨酸之外的氨基酸是色氨酸。
[0215]
在一些方面,在对应于seq id no:11的氨基酸残基116的位置处的除异亮氨酸之外的氨基酸由多于一个氨基酸组成,例如,两个氨基酸、三个氨基酸、四个氨基酸、五个氨基酸或者更多。在一些方面,多于一个氨基酸中的至少一个包含疏水侧链。在某些方面,在对应于seq id no:11的氨基酸残基116的位置处的除异亮氨酸之外的氨基酸由一系列,例如至少2个、至少3个、至少4个或至少5个氨基酸组成,其中所述系列氨基酸中的每一个都包含疏水侧链。
[0216]
在某些方面,hla ii类分子包含dqβ链,其中所述dqβ链包含在对应于seq id no:11的氨基酸残基118的位置处的除丝氨酸之外的氨基酸。任何除丝氨酸之外的氨基酸都可以存在于对应于seq id no:11的氨基酸残基118的位置处。在一些方面,除丝氨酸之外的氨基酸是包含带电侧链的氨基酸。在某些方面,在对应于seq id no:11的氨基酸残基118的位置处的除丝氨酸之外的氨基酸是选自精氨酸、组氨酸和赖氨酸的氨基酸。在某些方面,在对应于seq id no:11的氨基酸残基118的位置处的除丝氨酸之外的氨基酸是精氨酸。在某些方面,在对应于seq id no:11的氨基酸残基118的位置处的除丝氨酸之外的氨基酸是组氨酸。在某些方面,在对应于seq id no:11的氨基酸残基118的位置处的除丝氨酸之外的氨基酸是赖氨酸。
[0217]
在一些方面,在对应于seq id no:11的氨基酸残基118的位置处的除丝氨酸之外的氨基酸由多于一个氨基酸组成,例如,两个氨基酸、三个氨基酸、四个氨基酸、五个氨基酸
或者更多。在一些方面,多于一个氨基酸中的至少一个包含带电侧链。在某些方面,在对应于seq id no:11的氨基酸残基118的位置处的除丝氨酸之外的氨基酸由一系列,例如至少2个、至少3个、至少4个或至少5个氨基酸组成,其中所述系列氨基酸中的每一个都包含带电侧链。
[0218]
在某些方面,hla ii类分子包含dqβ链,其中所述dqβ链包含在对应于seq id no:11的氨基酸残基146的位置处的除脯氨酸之外的氨基酸。任何除脯氨酸之外的氨基酸都可以存在于对应于seq id no:11的氨基酸残基146的位置处。在一些方面,除脯氨酸之外的氨基酸是包含极性不带电侧链的氨基酸。在某些方面,在对应于seq id no:11的氨基酸残基146的位置处的除脯氨酸之外的氨基酸是选自丝氨酸、苏氨酸、天冬酰胺和谷氨酰胺的氨基酸。在某些方面,在对应于seq id no:11的氨基酸残基146的位置处的除脯氨酸之外的氨基酸是丝氨酸。在某些方面,在对应于seq id no:11的氨基酸残基146的位置处的除脯氨酸之外的氨基酸是苏氨酸。在某些方面,在对应于seq id no:11的氨基酸残基146的位置处的除脯氨酸之外的氨基酸是天冬酰胺。在某些方面,在对应于seq id no:11的氨基酸残基146的位置处的除脯氨酸之外的氨基酸是谷氨酰胺。
[0219]
在一些方面,在对应于seq id no:11的氨基酸残基146的位置处的除脯氨酸之外的氨基酸由多于一个氨基酸组成,例如,两个氨基酸、三个氨基酸、四个氨基酸、五个氨基酸或者更多。在一些方面,多于一个氨基酸中的至少一个包含极性不带电侧链。在某些方面,在对应于seq id no:11的氨基酸残基146的位置处的除脯氨酸之外的氨基酸由一系列,例如至少2个、至少3个、至少4个或至少5个氨基酸组成,其中所述系列氨基酸中的每一个都包含极性不带电侧链。
[0220]
在本公开的某些方面,mhc ii类分子包含相对于野生型dqβ链含有多于一个取代突变的dqβ链。在某些方面,dqβ链相对于野生型dqβ链包含至少两个突变、至少三个突变、至少四个突变、至少五个突变、至少六个突变、至少七个突变、至少八个突变、至少九个突变、或至少十个突变。
[0221]
在某些方面,dqβ链包含在对应于seq id no:11的氨基酸残基114的位置处的除亮氨酸之外的氨基酸和在对应于seq id no:11的氨基酸残基143的位置处的除缬氨酸之外的氨基酸。在某些方面,dqβ链包含在对应于seq id no:11的氨基酸残基114的位置处的除亮氨酸之外的氨基酸;在对应于seq id no:11的氨基酸残基143的位置处的除缬氨酸之外的氨基酸;以及下列中的至少三项:(i)在对应于seq id no:11的氨基酸残基110的位置处的除天冬酰胺之外的氨基酸,(ii)在对应于seq id no:11的氨基酸残基116的位置处的除异亮氨酸之外的氨基酸,(iii)在对应于seq id no:11的氨基酸残基118的位置处的除丝氨酸之外的氨基酸,和(iv)在对应于seq id no:11的氨基酸残基146的位置处的除脯氨酸之外的氨基酸。
[0222]
在一些方面,dqβ链包含(i)在对应于seq id no:11的氨基酸残基114的位置处的除亮氨酸之外的氨基酸;(ii)在对应于seq id no:11的氨基酸残基143的位置处的除缬氨酸之外的氨基酸;(iii)在对应于seq id no:11的氨基酸残基110的位置处的除天冬酰胺之外的氨基酸;(iv)在对应于seq id no:11的氨基酸残基116的位置处的除异亮氨酸之外的氨基酸;(v)在对应于seq id no:11的氨基酸残基118的位置处的除丝氨酸之外的氨基酸;和(vi)在对应于seq id no:11的氨基酸残基146的位置处的除脯氨酸之外的氨基酸。
id no:11的氨基酸残基146的位置处的脯氨酸。
[0227]
在一些方面,增加的对cd4的亲和力是参考hla ii类分子对cd4的亲和力的至少约1.5倍、至少约2倍、至少约3倍、至少约4倍、至少约5倍、至少约6倍、至少约7倍、至少约8倍、至少约9倍、至少约10倍、至少约15倍、至少约20倍、至少约25倍、至少约30倍、至少约35倍、至少约40倍、至少约45倍、至少约50倍、至少约75倍、至少约100倍、至少约200倍、至少约300倍、至少约400倍、至少约500倍、至少约1000倍、至少约1500倍、至少约2000倍、至少约2500倍、至少约3000倍、至少约3500倍、至少约4000倍、至少约4500倍、或至少约4000倍。
[0228]
在一些方面,增加的对cd4的亲和力是参考hla ii类分子对cd4的亲和力的至少约1.5倍到至少约5000倍、1.5倍到至少约4000倍、1.5倍到至少约3000倍、1.5倍到至少约2000倍、1.5倍到至少约1000倍、10倍到至少约5000倍、10倍到至少约4000倍、10倍到至少约3000倍、10倍到至少约2000倍、10倍到至少约1000倍、10倍到至少约900倍、10倍到至少约800倍、10倍到至少约700倍、10倍到至少约600倍、10倍到至少约500倍、10倍到至少约400倍、10倍到至少约300倍、10倍到至少约200倍、10倍到至少约100倍、100倍到至少约5000倍、100倍到至少约4000倍、100倍到至少约3000倍、100倍到至少约2000倍、100倍到至少约1000倍、100倍到至少约900倍、100倍到至少约800倍、100倍到至少约700倍、100倍到至少约600倍、100倍到至少约500倍、100倍到至少约400倍、100倍到至少约300倍、或100倍到至少约200倍。
[0229]
在某些方面,dqβ链包含选自以下的等位基因:hla-dqb1*02、hla-dqb1*03、hla-dqb1*04、hla-dqb1*05和hla-dqb1*06等位基因。在某些方面,dqβ链包含hla-dqb1*05等位基因。在特定方面,dqβ链包含hla-dqb1*05:01等位基因。
[0230]
在某些方面,dqβ链包含选自以下的等位基因:dqb1*02:01:01、dqb1*02:01:02、dqb1*02:01:03、dqb1*02:01:04、dqb1*02:01:05、dqb1*02:01:06、dqb1*02:01:07、dqb1*02:01:08、dqb1*02:01:09、dqb1*02:01:10、dqb1*02:01:11、dqb1*02:01:12、dqb1*02:01:13、dqb1*02:01:14、dqb1*02:01:15、dqb1*02:01:16、dqb1*02:01:17、dqb1*02:01:18、dqb1*02:01:19、dqb1*02:01:20、dqb1*02:01:21、dqb1*02:01:22、dqb1*02:01:23、dqb1*02:01:24、dqb1*02:01:25、dqb1*02:01:26、dqb1*02:01:27、dqb1*02:01:28、dqb1*02:01:29、dqb1*02:01:30、dqb1*02:01:31、dqb1*02:02:01:01、dqb1*02:02:01:02、dqb1*02:02:01:03、dqb1*02:02:01:04、dqb1*02:02:02、dqb1*02:02:03、dqb1*02:02:04、dqb1*02:02:05、dqb1*02:02:06、dqb1*02:02:07、dqb1*02:02:08、dqb1*02:02:09、dqb1*02:03:01、dqb1*02:03:02、dqb1*02:04、dqb1*02:05、dqb1*02:06、dqb1*02:07:01、dqb1*02:07:02、dqb1*02:08、dqb1*02:09、dqb1*02:10、dqb1*02:100、dqb1*02:101、dqb1*02:102、dqb1*02:103、dqb1*02:104、dqb1*02:105、dqb1*02:106、dqb1*02:107、dqb1*02:108、dqb1*02:109、dqb1*02:11、dqb1*02:110、dqb1*02:111、dqb1*02:112、dqb1*02:113、dqb1*02:114、dqb1*02:115、dqb1*02:116、dqb1*02:117、dqb1*02:118、dqb1*02:119、dqb1*02:12、dqb1*02:120、dqb1*02:121、dqb1*02:122、dqb1*02:123、dqb1*02:124、dqb1*02:125、dqb1*02:126、dqb1*02:127、dqb1*02:128、dqb1*02:129n、dqb1*02:13、dqb1*02:130、dqb1*02:131、dqb1*02:132n、dqb1*02:133、dqb1*02:134n、dqb1*02:135、dqb1*02:136、dqb1*02:137、dqb1*02:138、dqb1*02:139、dqb1*02:140、dqb1*02:141、dqb1*02:142、dqb1*02:14:01、dqb1*02:14:02、dqb1*02:15、dqb1*02:16、dqb1*02:17、dqb1*02:18n、dqb1*02:19、dqb1*02:20n、dqb1*02:21、dqb1*02:22、dqb1*02:23、dqb1*02:24、dqb1*02:25、dqb1*02:26、dqb1*02:27、dqb1*
02:28、dqb1*02:29、dqb1*02:30、dqb1*02:31、dqb1*02:32、dqb1*02:33、dqb1*02:34、dqb1*02:35、dqb1*02:36、dqb1*02:37、dqb1*02:38、dqb1*02:39、dqb1*02:40、dqb1*02:41、dqb1*02:42、dqb1*02:43、dqb1*02:44、dqb1*02:45、dqb1*02:46、dqb1*02:47、dqb1*02:48、dqb1*02:49、dqb1*02:50、dqb1*02:51、dqb1*02:52、dqb1*02:53q、dqb1*02:54、dqb1*02:55、dqb1*02:56、dqb1*02:57、dqb1*02:58n、dqb1*02:59、dqb1*02:60、dqb1*02:61、dqb1*02:62、dqb1*02:63、dqb1*02:64、dqb1*02:65、dqb1*02:66、dqb1*02:67nx、dqb1*02:68、dqb1*02:69、dqb1*02:70、dqb1*02:71、dqb1*02:72、dqb1*02:73、dqb1*02:74、dqb1*02:75、dqb1*02:76、dqb1*02:77、dqb1*02:78、dqb1*02:79、dqb1*02:80、dqb1*02:81、dqb1*02:82、dqb1*02:83、dqb1*02:84、dqb1*02:85、dqb1*02:86、dqb1*02:87、dqb1*02:88、dqb1*02:89:01、dqb1*02:89:02、dqb1*02:90、dqb1*02:91、dqb1*02:92、dqb1*02:93、dqb1*02:94、dqb1*02:95、dqb1*02:96n、dqb1*02:97、dqb1*02:98、dqb1*02:99、dqb1*03:01:01:01、dqb1*03:01:01:02、dqb1*03:01:01:03、dqb1*03:01:01:04、dqb1*03:01:01:05、dqb1*03:01:01:06、dqb1*03:01:01:07、dqb1*03:01:01:08、dqb1*03:01:01:09、dqb1*03:01:01:10、dqb1*03:01:01:11、dqb1*03:01:01:12、dqb1*03:01:01:14、dqb1*03:01:01:15、dqb1*03:01:01:16、dqb1*03:01:01:17、dqb1*03:01:01:18、dqb1*03:01:01:19、dqb1*03:01:01:20、dqb1*03:01:02、dqb1*03:01:03、dqb1*03:01:04、dqb1*03:01:05、dqb1*03:01:06、dqb1*03:01:07、dqb1*03:01:08、dqb1*03:01:09、dqb1*03:01:10、dqb1*03:01:11、dqb1*03:01:12、dqb1*03:01:13、dqb1*03:01:14、dqb1*03:01:15、dqb1*03:01:16、dqb1*03:01:17、dqb1*03:01:18、dqb1*03:01:19、dqb1*03:01:20、dqb1*03:01:21、dqb1*03:01:22、dqb1*03:01:23、dqb1*03:01:24、dqb1*03:01:25、dqb1*03:01:26、dqb1*03:01:27、dqb1*03:01:28、dqb1*03:01:29、dqb1*03:01:30、dqb1*03:01:31、dqb1*03:01:32、dqb1*03:01:33、dqb1*03:01:34、dqb1*03:01:35、dqb1*03:01:36、dqb1*03:01:37、dqb1*03:01:38、dqb1*03:01:39、dqb1*03:01:40、dqb1*03:01:41、dqb1*03:01:42、dqb1*03:01:43、dqb1*03:01:44、dqb1*03:01:45、dqb1*03:01:46、dqb1*03:02:01:01、dqb1*03:02:01:02、dqb1*03:02:01:03、dqb1*03:02:01:04、dqb1*03:02:01:05、dqb1*03:02:01:06、dqb1*03:02:01:07、dqb1*03:02:01:08、dqb1*03:02:02、dqb1*03:02:03、dqb1*03:02:04、dqb1*03:02:05、dqb1*03:02:06、dqb1*03:02:07、dqb1*03:02:08、dqb1*03:02:09、dqb1*03:02:10、dqb1*03:02:11、dqb1*03:02:12、dqb1*03:02:13、dqb1*03:02:14、dqb1*03:02:15、dqb1*03:02:16、dqb1*03:02:17、dqb1*03:02:18、dqb1*03:02:19、dqb1*03:02:20、dqb1*03:02:21、dqb1*03:02:22、dqb1*03:02:23、dqb1*03:02:24、dqb1*03:02:25、dqb1*03:02:26、dqb1*03:02:27、dqb1*03:02:28、dqb1*03:02:29、dqb1*03:02:30、dqb1*03:03:02:01、dqb1*03:03:02:02、dqb1*03:03:02:03、dqb1*03:03:02:04、dqb1*03:03:02:05、dqb1*03:03:03、dqb1*03:03:04、dqb1*03:03:05、dqb1*03:03:06、dqb1*03:03:07、dqb1*03:03:08、dqb1*03:03:09、dqb1*03:03:10、dqb1*03:03:11、dqb1*03:03:12、dqb1*03:03:13、dqb1*03:03:14、dqb1*03:03:15、dqb1*03:03:16、dqb1*03:03:17、dqb1*03:03:18、dqb1*03:03:19、dqb1*03:03:20、dqb1*03:03:21、dqb1*03:04:01、dqb1*03:04:02、dqb1*03:04:03、dqb1*03:04:04、dqb1*03:05:01、dqb1*03:05:02、dqb1*03:05:03、dqb1*03:05:04、dqb1*03:06、dqb1*03:07、dqb1*03:08、dqb1*03:09、dqb1*03:100、dqb1*03:101、dqb1*03:102、dqb1*03:103、dqb1*03:104、dqb1*03:105、dqb1*03:106、dqb1*03:107、dqb1*03:108、dqb1*03:109、dqb1*
03:10:01、dqb1*03:10:02:01、dqb1*03:10:02:02、dqb1*03:11、dqb1*03:110、dqb1*03:111、dqb1*03:112、dqb1*03:113、dqb1*03:114、dqb1*03:115、dqb1*03:116、dqb1*03:117、dqb1*03:118n、dqb1*03:119、dqb1*03:12、dqb1*03:120、dqb1*03:121、dqb1*03:122、dqb1*03:123、dqb1*03:124、dqb1*03:125、dqb1*03:126、dqb1*03:127、dqb1*03:128、dqb1*03:129、dqb1*03:13、dqb1*03:130、dqb1*03:131、dqb1*03:132、dqb1*03:133、dqb1*03:134、dqb1*03:135、dqb1*03:136、dqb1*03:137、dqb1*03:138、dqb1*03:139、dqb1*03:140、dqb1*03:141、dqb1*03:142、dqb1*03:143、dqb1*03:144、dqb1*03:145、dqb1*03:146、dqb1*03:147、dqb1*03:148、dqb1*03:149、dqb1*03:14:01、dqb1*03:14:02、dqb1*03:15、dqb1*03:150、dqb1*03:151、dqb1*03:152、dqb1*03:153、dqb1*03:154、dqb1*03:155、dqb1*03:156、dqb1*03:157、dqb1*03:158、dqb1*03:159、dqb1*03:16、dqb1*03:160、dqb1*03:161、dqb1*03:162、dqb1*03:163、dqb1*03:164、dqb1*03:165、dqb1*03:166、dqb1*03:167、dqb1*03:168、dqb1*03:169、dqb1*03:170、dqb1*03:171、dqb1*03:172、dqb1*03:173、dqb1*03:174、dqb1*03:175、dqb1*03:176、dqb1*03:177、dqb1*03:178、dqb1*03:179、dqb1*03:17:01、dqb1*03:17:02、dqb1*03:18、dqb1*03:180、dqb1*03:181、dqb1*03:182、dqb1*03:183、dqb1*03:184、dqb1*03:185、dqb1*03:186、dqb1*03:187、dqb1*03:188、dqb1*03:189、dqb1*03:190、dqb1*03:191、dqb1*03:192、dqb1*03:193、dqb1*03:194、dqb1*03:195、dqb1*03:196、dqb1*03:197q、dqb1*03:198:01、dqb1*03:198:02、dqb1*03:199、dqb1*03:19:01、dqb1*03:19:02、dqb1*03:19:03、dqb1*03:19:04、dqb1*03:20、dqb1*03:200、dqb1*03:201、dqb1*03:202、dqb1*03:203、dqb1*03:204、dqb1*03:205、dqb1*03:206、dqb1*03:207、dqb1*03:208、dqb1*03:209、dqb1*03:21、dqb1*03:210、dqb1*03:211、dqb1*03:212、dqb1*03:213nx、dqb1*03:214、dqb1*03:215、dqb1*03:216、dqb1*03:217、dqb1*03:218、dqb1*03:219、dqb1*03:220、dqb1*03:221、dqb1*03:222、dqb1*03:223、dqb1*03:224、dqb1*03:225、dqb1*03:226、dqb1*03:227、dqb1*03:228、dqb1*03:229、dqb1*03:22:01、dqb1*03:22:02、dqb1*03:230、dqb1*03:231、dqb1*03:232、dqb1*03:233、dqb1*03:234、dqb1*03:235、dqb1*03:236、dqb1*03:237n、dqb1*03:238、dqb1*03:239、dqb1*03:23:01、dqb1*03:23:02、dqb1*03:23:03、dqb1*03:24、dqb1*03:240、dqb1*03:241、dqb1*03:242、dqb1*03:243、dqb1*03:244、dqb1*03:245、dqb1*03:246、dqb1*03:247、dqb1*03:248、dqb1*03:249、dqb1*03:250、dqb1*03:251、dqb1*03:252、dqb1*03:253、dqb1*03:254、dqb1*03:255、dqb1*03:256、dqb1*03:257、dqb1*03:258、dqb1*03:259、dqb1*03:25:01、dqb1*03:25:02、dqb1*03:26、dqb1*03:260、dqb1*03:261、dqb1*03:262、dqb1*03:263、dqb1*03:264、dqb1*03:265、dqb1*03:266、dqb1*03:267、dqb1*03:268、dqb1*03:269n、dqb1*03:27、dqb1*03:270、dqb1*03:271、dqb1*03:272、dqb1*03:273、dqb1*03:274、dqb1*03:275、dqb1*03:277、dqb1*03:278、dqb1*03:279、dqb1*03:28、dqb1*03:280、dqb1*03:281、dqb1*03:282n、dqb1*03:283、dqb1*03:284、dqb1*03:285、dqb1*03:286、dqb1*03:287、dqb1*03:288、dqb1*03:289、dqb1*03:29、dqb1*03:290、dqb1*03:291、dqb1*03:292、dqb1*03:293、dqb1*03:294、dqb1*03:295、dqb1*03:296、dqb1*03:297、dqb1*03:298、dqb1*03:299、dqb1*03:30、dqb1*03:300、dqb1*03:301、dqb1*03:302、dqb1*03:303n、dqb1*03:304、dqb1*03:305、dqb1*03:306、dqb1*03:307、dqb1*03:308、dqb1*03:309、dqb1*03:31、dqb1*03:310n、dqb1*03:311、dqb1*03:312、dqb1*03:313、dqb1*03:314、dqb1*03:315、dqb1*03:316、dqb1*03:317:01、dqb1*03:317:02、
dqb1*03:318、dqb1*03:319、dqb1*03:32、dqb1*03:320、dqb1*03:321、dqb1*03:322、dqb1*03:323、dqb1*03:324、dqb1*03:326、dqb1*03:327、dqb1*03:328、dqb1*03:329、dqb1*03:33、dqb1*03:330、dqb1*03:331、dqb1*03:332、dqb1*03:333、dqb1*03:334n4bp、dqb1*03:335、dqb1*03:336、dqb1*03:337、dqb1*03:338n、dqb1*03:339n、dqb1*03:34、dqb1*03:340n、dqb1*03:341、dqb1*03:342、dqb1*03:343、dqb1*03:344、dqb1*03:345、dqb1*03:346、dqb1*03:347、dqb1*03:348、dqb1*03:349、dqb1*03:35、dqb1*03:350、dqb1*03:351、dqb1*03:352、dqb1*03:353、dqb1*03:354n、dqb1*03:355、dqb1*03:356nx、dqb1*03:357n、dqb1*03:358n、dqb1*03:36、dqb1*03:37、dqb1*03:38:01、dqb1*03:38:02、dqb1*03:39、dqb1*03:40、dqb1*03:41、dqb1*03:42、dqb1*03:43、dqb1*03:44、dqb1*03:45、dqb1*03:46、dqb1*03:47、dqb1*03:48、dqb1*03:49、dqb1*03:50、dqb1*03:51、dqb1*03:52、dqb1*03:53、dqb1*03:54、dqb1*03:55、dqb1*03:56、dqb1*03:57、dqb1*03:58、dqb1*03:59、dqb1*03:60、dqb1*03:61、dqb1*03:62、dqb1*03:63、dqb1*03:64、dqb1*03:65、dqb1*03:66n、dqb1*03:67、dqb1*03:68、dqb1*03:69、dqb1*03:70、dqb1*03:71、dqb1*03:72、dqb1*03:73、dqb1*03:74、dqb1*03:75、dqb1*03:76、dqb1*03:77、dqb1*03:78、dqb1*03:79、dqb1*03:80、dqb1*03:81、dqb1*03:82、dqb1*03:83、dqb1*03:84n、dqb1*03:85、dqb1*03:86、dqb1*03:87、dqb1*03:88、dqb1*03:89、dqb1*03:90n、dqb1*03:91q、dqb1*03:92、dqb1*03:93、dqb1*03:94、dqb1*03:95n、dqb1*03:96、dqb1*03:97、dqb1*03:98、dqb1*03:99q、dqb1*04:01:01:01、dqb1*04:01:01:02、dqb1*04:01:02、dqb1*04:01:03、dqb1*04:01:04、dqb1*04:01:05、dqb1*04:02:01:01、dqb1*04:02:01:04、dqb1*04:02:01:05、dqb1*04:02:01:06、dqb1*04:02:01:07、dqb1*04:02:01:08、dqb1*04:02:01:09、dqb1*04:02:01:10、dqb1*04:02:02、dqb1*04:02:03、dqb1*04:02:04、dqb1*04:02:05、dqb1*04:02:06、dqb1*04:02:07、dqb1*04:02:08、dqb1*04:02:09、dqb1*04:02:10、dqb1*04:02:11、dqb1*04:02:12、dqb1*04:02:13、dqb1*04:02:14、dqb1*04:02:15、dqb1*04:02:16、dqb1*04:02:17、dqb1*04:02:18、dqb1*04:03:01、dqb1*04:03:02、dqb1*04:03:03、dqb1*04:04、dqb1*04:05、dqb1*04:06、dqb1*04:07、dqb1*04:08、dqb1*04:09、dqb1*04:10、dqb1*04:11、dqb1*04:12、dqb1*04:13、dqb1*04:14、dqb1*04:15、dqb1*04:16、dqb1*04:17、dqb1*04:18、dqb1*04:19、dqb1*04:20、dqb1*04:21、dqb1*04:22、dqb1*04:23、dqb1*04:24、dqb1*04:25n、dqb1*04:26、dqb1*04:27、dqb1*04:28、dqb1*04:29、dqb1*04:30、dqb1*04:31、dqb1*04:32、dqb1*04:33、dqb1*04:34、dqb1*04:35、dqb1*04:36n、dqb1*04:37、dqb1*04:38、dqb1*04:39、dqb1*04:40、dqb1*04:41n、dqb1*04:42、dqb1*04:43、dqb1*04:44、dqb1*04:45、dqb1*04:46n、dqb1*04:47、dqb1*04:48、dqb1*04:49、dqb1*04:50、dqb1*04:51、dqb1*04:52、dqb1*04:53、dqb1*04:54、dqb1*04:55、dqb1*04:56、dqb1*04:57、dqb1*04:58、dqb1*04:59n、dqb1*04:60、dqb1*04:61、dqb1*04:62、dqb1*05:01:01:01、dqb1*05:01:01:02、dqb1*05:01:01:03、dqb1*05:01:01:04、dqb1*05:01:01:05、dqb1*05:01:02、dqb1*05:01:03、dqb1*05:01:04、dqb1*05:01:05、dqb1*05:01:06、dqb1*05:01:07、dqb1*05:01:08、dqb1*05:01:09、dqb1*05:01:10、dqb1*05:01:11、dqb1*05:01:12、dqb1*05:01:13、dqb1*05:01:14、dqb1*05:01:15、dqb1*05:01:16、dqb1*05:01:17、dqb1*05:01:18、dqb1*05:01:19、dqb1*05:01:20、dqb1*05:01:21、dqb1*05:01:22、dqb1*05:01:23、dqb1*05:01:24:01、dqb1*05:01:24:02、dqb1*05:01:25、dqb1*05:01:26、dqb1*05:01:27、dqb1*05:01:28、dqb1*05:01:29、dqb1*05:01:30、dqb1*05:01:31、
dqb1*05:01:32、dqb1*05:01:33、dqb1*05:01:34、dqb1*05:02:01:01、dqb1*05:02:01:02、dqb1*05:02:01:03、dqb1*05:02:01:04、dqb1*05:02:01:05、dqb1*05:02:01:06、dqb1*05:02:02、dqb1*05:02:03、dqb1*05:02:04、dqb1*05:02:05、dqb1*05:02:06、dqb1*05:02:07、dqb1*05:02:08、dqb1*05:02:09、dqb1*05:02:10、dqb1*05:02:11、dqb1*05:02:12、dqb1*05:02:13、dqb1*05:02:14、dqb1*05:02:15、dqb1*05:02:16、dqb1*05:02:17、dqb1*05:02:18、dqb1*05:02:19、dqb1*05:03:01:01、dqb1*05:03:01:02、dqb1*05:03:01:03、dqb1*05:03:02、dqb1*05:03:03、dqb1*05:03:04、dqb1*05:03:05、dqb1*05:03:06、dqb1*05:03:07、dqb1*05:03:08、dqb1*05:03:09、dqb1*05:03:10、dqb1*05:03:11、dqb1*05:03:12、dqb1*05:03:13、dqb1*05:03:14、dqb1*05:03:15、dqb1*05:03:16、dqb1*05:03:17、dqb1*05:03:18、dqb1*05:03:19、dqb1*05:03:20、dqb1*05:04、dqb1*05:05:01、dqb1*05:05:02、dqb1*05:06:01、dqb1*05:06:02、dqb1*05:07、dqb1*05:08、dqb1*05:09、dqb1*05:10、dqb1*05:100、dqb1*05:101、dqb1*05:102、dqb1*05:103、dqb1*05:104、dqb1*05:105、dqb1*05:106、dqb1*05:107、dqb1*05:108、dqb1*05:109、dqb1*05:110n、dqb1*05:111、dqb1*05:112、dqb1*05:113、dqb1*05:114、dqb1*05:115、dqb1*05:116、dqb1*05:117、dqb1*05:118、dqb1*05:119、dqb1*05:11:01、dqb1*05:11:02、dqb1*05:12、dqb1*05:120、dqb1*05:121、dqb1*05:122、dqb1*05:123、dqb1*05:124、dqb1*05:125、dqb1*05:126、dqb1*05:127、dqb1*05:128n、dqb1*05:129、dqb1*05:13、dqb1*05:130、dqb1*05:131、dqb1*05:132q、dqb1*05:133、dqb1*05:134、dqb1*05:135、dqb1*05:136、dqb1*05:137、dqb1*05:138、dqb1*05:139、dqb1*05:14、dqb1*05:140、dqb1*05:141、dqb1*05:142、dqb1*05:143、dqb1*05:144、dqb1*05:145、dqb1*05:146、dqb1*05:147、dqb1*05:148、dqb1*05:149、dqb1*05:15、dqb1*05:150、dqb1*05:151、dqb1*05:152、dqb1*05:153、dqb1*05:154、dqb1*05:155、dqb1*05:156、dqb1*05:157、dqb1*05:158、dqb1*05:159、dqb1*05:16、dqb1*05:160、dqb1*05:161、dqb1*05:162、dqb1*05:163、dqb1*05:164、dqb1*05:165、dqb1*05:166、dqb1*05:167、dqb1*05:168、dqb1*05:169、dqb1*05:17、dqb1*05:170、dqb1*05:171、dqb1*05:172、dqb1*05:173、dqb1*05:174、dqb1*05:175、dqb1*05:176、dqb1*05:177、dqb1*05:178、dqb1*05:179、dqb1*05:18、dqb1*05:180、dqb1*05:181、dqb1*05:182、dqb1*05:183、dqb1*05:184、dqb1*05:185n、dqb1*05:186、dqb1*05:187、dqb1*05:188、dqb1*05:189、dqb1*05:19、dqb1*05:190、dqb1*05:191、dqb1*05:192、dqb1*05:193、dqb1*05:194、dqb1*05:195、dqb1*05:196、dqb1*05:197、dqb1*05:198、dqb1*05:199、dqb1*05:20、dqb1*05:200、dqb1*05:201、dqb1*05:202、dqb1*05:203、dqb1*05:204、dqb1*05:205、dqb1*05:206n、dqb1*05:207、dqb1*05:208n5bp、dqb1*05:209、dqb1*05:21、dqb1*05:210、dqb1*05:211、dqb1*05:212、dqb1*05:213、dqb1*05:214、dqb1*05:215n、dqb1*05:216、dqb1*05:217、dqb1*05:22、dqb1*05:23、dqb1*05:24、dqb1*05:25、dqb1*05:26、dqb1*05:27、dqb1*05:28、dqb1*05:29、dqb1*05:30、dqb1*05:31、dqb1*05:32、dqb1*05:33、dqb1*05:34、dqb1*05:35、dqb1*05:36、dqb1*05:37、dqb1*05:38、dqb1*05:39、dqb1*05:40、dqb1*05:41n、dqb1*05:42、dqb1*05:43:01、dqb1*05:43:02、dqb1*05:44、dqb1*05:45、dqb1*05:46、dqb1*05:47、dqb1*05:48、dqb1*05:49、dqb1*05:50、dqb1*05:51、dqb1*05:52、dqb1*05:53、dqb1*05:54、dqb1*05:55、dqb1*05:56、dqb1*05:57、dqb1*05:58、dqb1*05:59、dqb1*05:60、dqb1*05:61、dqb1*05:62、dqb1*05:63、dqb1*05:64、dqb1*05:65、dqb1*05:66:01、dqb1*05:66:02、dqb1*05:67、dqb1*05:68、dqb1*05:69、dqb1*
05:70、dqb1*05:71、dqb1*05:72、dqb1*05:73、dqb1*05:74、dqb1*05:75、dqb1*05:76、dqb1*05:77、dqb1*05:78、dqb1*05:79、dqb1*05:80、dqb1*05:81、dqb1*05:82、dqb1*05:83、dqb1*05:84、dqb1*05:85、dqb1*05:86、dqb1*05:87q、dqb1*05:88、dqb1*05:89:01、dqb1*05:89:02、dqb1*05:90n、dqb1*05:91、dqb1*05:92、dqb1*05:93、dqb1*05:94、dqb1*05:95、dqb1*05:96、dqb1*05:97、dqb1*05:98、dqb1*05:99、dqb1*06:01:01:01、dqb1*06:01:01:02、dqb1*06:01:02、dqb1*06:01:03、dqb1*06:01:04、dqb1*06:01:05、dqb1*06:01:06、dqb1*06:01:07、dqb1*06:01:08、dqb1*06:01:09、dqb1*06:01:10、dqb1*06:01:11、dqb1*06:01:12、dqb1*06:01:13、dqb1*06:01:14、dqb1*06:01:15、dqb1*06:01:16、dqb1*06:01:17、dqb1*06:01:18、dqb1*06:01:19、dqb1*06:01:20、dqb1*06:01:21、dqb1*06:02:01:01、dqb1*06:02:01:02、dqb1*06:02:01:03、dqb1*06:02:01:04、dqb1*06:02:02、dqb1*06:02:03、dqb1*06:02:04、dqb1*06:02:05、dqb1*06:02:06、dqb1*06:02:07、dqb1*06:02:08、dqb1*06:02:09、dqb1*06:02:10、dqb1*06:02:11、dqb1*06:02:12、dqb1*06:02:13、dqb1*06:02:14、dqb1*06:02:15、dqb1*06:02:16、dqb1*06:02:17、dqb1*06:02:18、dqb1*06:02:19、dqb1*06:02:20、dqb1*06:02:21、dqb1*06:02:22、dqb1*06:02:23、dqb1*06:02:24、dqb1*06:02:25、dqb1*06:02:26、dqb1*06:02:27、dqb1*06:02:28、dqb1*06:02:29、dqb1*06:02:30、dqb1*06:02:31、dqb1*06:02:32、dqb1*06:02:33、dqb1*06:02:34、dqb1*06:02:35、dqb1*06:02:36、dqb1*06:02:37、dqb1*06:02:38、dqb1*06:03:01:01、dqb1*06:03:01:02、dqb1*06:03:01:03、dqb1*06:03:02、dqb1*06:03:03、dqb1*06:03:04、dqb1*06:03:05、dqb1*06:03:06、dqb1*06:03:07、dqb1*06:03:08、dqb1*06:03:09、dqb1*06:03:10、dqb1*06:03:11、dqb1*06:03:12、dqb1*06:03:13、dqb1*06:03:14、dqb1*06:03:15、dqb1*06:03:16、dqb1*06:03:17、dqb1*06:03:18、dqb1*06:03:19、dqb1*06:03:20、dqb1*06:03:21、dqb1*06:03:22、dqb1*06:03:23、dqb1*06:03:24、dqb1*06:03:25、dqb1*06:03:26、dqb1*06:03:27、dqb1*06:03:28、dqb1*06:03:29、dqb1*06:03:30、dqb1*06:03:31、dqb1*06:03:32、dqb1*06:03:33、dqb1*06:03:34、dqb1*06:03:35、dqb1*06:04:01、dqb1*06:04:02、dqb1*06:04:03、dqb1*06:04:04、dqb1*06:04:05、dqb1*06:04:06、dqb1*06:04:07、dqb1*06:04:08、dqb1*06:04:09、dqb1*06:04:10、dqb1*06:04:11、dqb1*06:04:12、dqb1*06:05:01、dqb1*06:05:02、dqb1*06:06、dqb1*06:07:01、dqb1*06:07:02、dqb1*06:08:01、dqb1*06:08:02、dqb1*06:08:03、dqb1*06:09:01:01、dqb1*06:09:01:02、dqb1*06:09:02、dqb1*06:09:03、dqb1*06:09:04、dqb1*06:09:05、dqb1*06:09:06、dqb1*06:09:07、dqb1*06:09:08、dqb1*06:09:09、dqb1*06:09:10、dqb1*06:10、dqb1*06:100、dqb1*06:101、dqb1*06:102n、dqb1*06:103、dqb1*06:104、dqb1*06:105、dqb1*06:106、dqb1*06:107、dqb1*06:108、dqb1*06:109、dqb1*06:110、dqb1*06:111、dqb1*06:112n、dqb1*06:113、dqb1*06:114、dqb1*06:115、dqb1*06:116、dqb1*06:117、dqb1*06:118:01、dqb1*06:118:02、dqb1*06:118:03、dqb1*06:119、dqb1*06:11:01、dqb1*06:11:02、dqb1*06:11:03、dqb1*06:11:04、dqb1*06:12、dqb1*06:120、dqb1*06:121、dqb1*06:122、dqb1*06:123、dqb1*06:124、dqb1*06:125、dqb1*06:126、dqb1*06:127、dqb1*06:128、dqb1*06:129、dqb1*06:130、dqb1*06:131、dqb1*06:132、dqb1*06:133、dqb1*06:134、dqb1*06:135、dqb1*06:136、dqb1*06:137、dqb1*06:138、dqb1*06:139、dqb1*06:13:01、dqb1*06:13:02、dqb1*06:13:03、dqb1*06:140、dqb1*06:141、dqb1*06:142、dqb1*06:143、dqb1*06:144n、dqb1*06:145、dqb1*06:146:
01、dqb1*06:146:02、dqb1*06:147、dqb1*06:148、dqb1*06:149、dqb1*06:14:01、dqb1*06:14:02、dqb1*06:14:03、dqb1*06:150、dqb1*06:151、dqb1*06:152、dqb1*06:153:01、dqb1*06:153:02、dqb1*06:154、dqb1*06:155、dqb1*06:156、dqb1*06:157、dqb1*06:158n、dqb1*06:159、dqb1*06:15:01、dqb1*06:15:02、dqb1*06:16、dqb1*06:160、dqb1*06:161、dqb1*06:162、dqb1*06:163、dqb1*06:164、dqb1*06:165、dqb1*06:166、dqb1*06:167、dqb1*06:168、dqb1*06:169、dqb1*06:17、dqb1*06:170、dqb1*06:171、dqb1*06:172、dqb1*06:173、dqb1*06:174、dqb1*06:175、dqb1*06:176、dqb1*06:177、dqb1*06:178、dqb1*06:179n、dqb1*06:180、dqb1*06:181、dqb1*06:182、dqb1*06:183、dqb1*06:184、dqb1*06:185、dqb1*06:186、dqb1*06:187、dqb1*06:188、dqb1*06:189、dqb1*06:18:01、dqb1*06:18:02、dqb1*06:190:01、dqb1*06:190:02、dqb1*06:191、dqb1*06:192、dqb1*06:193n、dqb1*06:194、dqb1*06:195、dqb1*06:196、dqb1*06:197、dqb1*06:198、dqb1*06:199、dqb1*06:19:01、dqb1*06:19:02、dqb1*06:20、dqb1*06:200、dqb1*06:201、dqb1*06:202、dqb1*06:203、dqb1*06:204、dqb1*06:205、dqb1*06:206:01、dqb1*06:206:02、dqb1*06:207、dqb1*06:208、dqb1*06:209、dqb1*06:21、dqb1*06:210、dqb1*06:211、dqb1*06:212、dqb1*06:213、dqb1*06:214、dqb1*06:215、dqb1*06:216n、dqb1*06:217、dqb1*06:218、dqb1*06:219、dqb1*06:221、dqb1*06:222、dqb1*06:223、dqb1*06:224、dqb1*06:225、dqb1*06:226、dqb1*06:227、dqb1*06:228、dqb1*06:229、dqb1*06:22:01、dqb1*06:22:02、dqb1*06:22:03、dqb1*06:23、dqb1*06:230、dqb1*06:231、dqb1*06:232、dqb1*06:233、dqb1*06:234、dqb1*06:235、dqb1*06:236、dqb1*06:237、dqb1*06:238、dqb1*06:239、dqb1*06:24、dqb1*06:240、dqb1*06:241、dqb1*06:242、dqb1*06:243、dqb1*06:244、dqb1*06:245、dqb1*06:246、dqb1*06:247、dqb1*06:248、dqb1*06:249、dqb1*06:25、dqb1*06:250、dqb1*06:251、dqb1*06:252n、dqb1*06:253、dqb1*06:254、dqb1*06:255、dqb1*06:256、dqb1*06:257、dqb1*06:258、dqb1*06:259、dqb1*06:260、dqb1*06:261、dqb1*06:262、dqb1*06:263、dqb1*06:264、dqb1*06:265、dqb1*06:266、dqb1*06:267、dqb1*06:268、dqb1*06:269、dqb1*06:26n、dqb1*06:270:01、dqb1*06:270:02、dqb1*06:271、dqb1*06:272、dqb1*06:273、dqb1*06:274、dqb1*06:275、dqb1*06:276、dqb1*06:277、dqb1*06:278、dqb1*06:279、dqb1*06:27:01、dqb1*06:27:02、dqb1*06:28、dqb1*06:280、dqb1*06:281、dqb1*06:282、dqb1*06:283、dqb1*06:284、dqb1*06:285、dqb1*06:286、dqb1*06:287、dqb1*06:288、dqb1*06:289、dqb1*06:29、dqb1*06:290、dqb1*06:291、dqb1*06:292、dqb1*06:293、dqb1*06:294、dqb1*06:295、dqb1*06:296、dqb1*06:297、dqb1*06:298、dqb1*06:299、dqb1*06:30、dqb1*06:300、dqb1*06:301、dqb1*06:302、dqb1*06:303n、dqb1*06:304n、dqb1*06:305、dqb1*06:306n、dqb1*06:307、dqb1*06:308n、dqb1*06:309、dqb1*06:31、dqb1*06:310、dqb1*06:311、dqb1*06:312、dqb1*06:313、dqb1*06:314、dqb1*06:315、dqb1*06:316、dqb1*06:317n、dqb1*06:318、dqb1*06:319、dqb1*06:320、dqb1*06:321、dqb1*06:322、dqb1*06:323、dqb1*06:324、dqb1*06:325、dqb1*06:326、dqb1*06:32:01、dqb1*06:32:02、dqb1*06:33、dqb1*06:34、dqb1*06:35、dqb1*06:36、dqb1*06:37、dqb1*06:38、dqb1*06:39、dqb1*06:40、dqb1*06:41、dqb1*06:42、dqb1*06:43、dqb1*06:44、dqb1*06:45、dqb1*06:46、dqb1*06:47、dqb1*06:48:01、dqb1*06:48:02、dqb1*06:49、dqb1*06:50、dqb1*06:51:01、dqb1*06:51:02、dqb1*06:52、dqb1*06:53:01、dqb1*06:53:02、dqb1*06:54n、dqb1*06:55、dqb1*06:56、dqb1*06:57、
dqb1*06:58、dqb1*06:59、dqb1*06:60、dqb1*06:61、dqb1*06:62、dqb1*06:63、dqb1*06:64、dqb1*06:65、dqb1*06:66、dqb1*06:67、dqb1*06:68、dqb1*06:69:01、dqb1*06:69:02、dqb1*06:70、dqb1*06:71、dqb1*06:72、dqb1*06:73、dqb1*06:74、dqb1*06:75nx、dqb1*06:76、dqb1*06:77n、dqb1*06:78、dqb1*06:79:01、dqb1*06:79:02、dqb1*06:80、dqb1*06:81、dqb1*06:82、dqb1*06:83、dqb1*06:84、dqb1*06:85、dqb1*06:86、dqb1*06:87、dqb1*06:88、dqb1*06:89、dqb1*06:90、dqb1*06:91、dqb1*06:92:01、dqb1*06:92:02、dqb1*06:93、dqb1*06:94、dqb1*06:95、dqb1*06:96:01、dqb1*06:96:02、dqb1*06:97、dqb1*06:98、dqb1*06:99:01、dqb1*06:99:02、及其任何组合。
[0231]
在某些方面,mhc ii类分子包含dqβ链,所述dqβ链包含与seq id no:13具有至少约70%、至少约75%、至少约80%、至少约85%、至少约90%、至少约95%、至少约96%、至少约97%、至少约98%、至少约99%或约100%序列同一性的氨基酸序列,其中所述dqβ链包含在对应于seq id no:11的氨基酸残基114的位置处的色氨酸,并且其中所述dqβ链包含在对应于seq id no:11的氨基酸残基143的位置处的甲硫氨酸。在某些方面,mhc ii类分子包含dqβ链,所述dqβ链包含与seq id no:13具有至少约70%、至少约75%、至少约80%、至少约85%、至少约90%、至少约95%、至少约96%、至少约97%、至少约98%、至少约99%或约100%序列同一性的氨基酸序列,其中所述dqβ链包含(i)在对应于seq id no:11的氨基酸残基114的位置处的色氨酸,(ii)在对应于seq id no:11的氨基酸残基143的位置处的甲硫氨酸,(iii)在对应于seq id no:11的氨基酸残基110的位置处的谷氨酰胺;(iv)在对应于seq id no:11的氨基酸残基116的位置处的缬氨酸;(v)在对应于seq id no:11的氨基酸残基118的位置处的组氨酸;和(vi)在对应于seq id no:11的氨基酸残基146的位置处的谷氨酰胺。在某些方面,mhc ii类分子包含dqβ链,所述dqβ链包含seq id no:13所示的氨基酸序列。
[0232]
ii.a.2.b.hla-dqα链
[0233]
在本公开的一些方面,mhc ii类分子还包含α链。在一些方面,α链是野生型α链。在一些方面,α链是dqα链。可以在本公开的组合物和方法中使用任何dqα链。在一些方面,dqα链包含hla-dqa1*01、hla-dqa1*02、hla-dqa1*03、hla-dqa1*04、hla-dqa1*05、或hla-dqa1*06等位基因。在某些方面,dqα链包含hla-dqa1*01等位基因。在某些方面,dqα链包含hla-dqa1*02等位基因。在某些方面,dqα链包含hla-dqa1*03等位基因。在某些方面,dqα链包含hla-dqa1*04等位基因。在某些方面,dqα链包含hla-dqa1*05等位基因。在某些方面,dqα链包含hla-dqa1*06等位基因。
[0234]
在某些方面,dqα链选自dqa1*01:01:01:01、dqa1*01:01:01:02、dqa1*01:01:01:03、dqa1*01:01:01:05、dqa1*01:01:01:06、dqa1*01:01:02、dqa1*01:01:03、dqa1*01:01:04、dqa1*01:01:05、dqa1*01:02:01:01、dqa1*01:02:01:02、dqa1*01:02:01:03、dqa1*01:02:01:04、dqa1*01:02:01:05、dqa1*01:02:01:06、dqa1*01:02:01:07、dqa1*01:02:01:08、dqa1*01:02:01:09、dqa1*01:02:01:10、dqa1*01:02:01:11、dqa1*01:02:01:12、dqa1*01:02:02:01、dqa1*01:02:02:02、dqa1*01:02:02:03、dqa1*01:02:02:04、dqa1*01:02:03、dqa1*01:02:04、dqa1*01:03:01:01、dqa1*01:03:01:02、dqa1*01:03:01:03、dqa1*01:03:01:04、dqa1*01:03:01:05、dqa1*01:03:01:06、dqa1*01:03:01:07、dqa1*01:03:01:08、dqa1*01:03:01:09、dqa1*01:04:01:01、dqa1*01:04:01:02、dqa1*01:04:01:03、dqa1*01:
04:01:04、dqa1*01:04:02、dqa1*01:05:01、dqa1*01:05:02、dqa1*01:06、dqa1*01:07q、dqa1*01:08、dqa1*01:09、dqa1*01:10、dqa1*01:11、dqa1*01:12、dqa1*01:13、dqa1*01:14、dqa1*01:15n、dqa1*01:16n、dqa1*01:17、dqa1*01:18、dqa1*01:19、dqa1*01:20、dqa1*01:21、dqa1*01:22、dqa1*01:23、dqa1*01:24、dqa1*01:25、dqa1*01:26、dqa1*02:01:01:01、dqa1*02:01:01:02、dqa1*02:01:02、dqa1*02:02n、dqa1*02:03、dqa1*03:01:01、dqa1*03:01:03、dqa1*03:02:01:01、dqa1*03:02:01:02、dqa1*03:03:01:01、dqa1*03:03:01:02、dqa1*03:03:01:03、dqa1*03:03:01:04、dqa1*03:03:01:05、dqa1*03:03:01:06、dqa1*03:03:01:07、dqa1*03:03:02、dqa1*03:04、dqa1*03:05、dqa1*03:06、dqa1*03:07、dqa1*04:01:01:01、dqa1*04:01:01:02、dqa1*04:01:01:03、dqa1*04:01:01:04、dqa1*04:01:01:05、dqa1*04:01:01:06、dqa1*04:01:01:07、dqa1*04:01:01:08、dqa1*04:01:02:01、dqa1*04:01:02:02、dqa1*04:01:03、dqa1*04:02、dqa1*04:03n、dqa1*04:04、dqa1*04:05、dqa1*05:01:01:01、dqa1*05:01:01:02、dqa1*05:01:01:03、dqa1*05:01:01:04、dqa1*05:01:02、dqa1*05:01:04、dqa1*05:01:05、dqa1*05:01:06、dqa1*05:02、dqa1*05:03:01:01、dqa1*05:03:01:02、dqa1*05:04、dqa1*05:05:01:01、dqa1*05:05:01:02、dqa1*05:05:01:03、dqa1*05:05:01:04、dqa1*05:05:01:05、dqa1*05:05:01:06、dqa1*05:05:01:07、dqa1*05:05:01:08、dqa1*05:05:01:09、dqa1*05:05:01:10、dqa1*05:05:01:11、dqa1*05:05:01:12、dqa1*05:05:01:13、dqa1*05:05:01:14、dqa1*05:05:01:15、dqa1*05:05:01:16、dqa1*05:05:01:17、dqa1*05:05:01:18、dqa1*05:05:01:19、dqa1*05:05:01:20、dqa1*05:06:01:01、dqa1*05:06:01:02、dqa1*05:07、dqa1*05:08、dqa1*05:09、dqa1*05:10、dqa1*05:11、dqa1*05:12、dqa1*05:13、dqa1*05:14、dqa1*05:15n、dqa1*06:01:01:01、dqa1*06:01:01:02、dqa1*06:01:01:03、dqa1*06:01:01:04、dqa1*06:01:02、dqa1*06:02、及其任何组合。
[0235]
在某些方面,mhc ii类分子包含dqα链,所述dqα链包含与seq id no:16具有至少约70%、至少约75%、至少约80%、至少约85%、至少约90%、至少约95%、至少约96%、至少约97%、至少约98%、至少约99%或约100%序列同一性的氨基酸序列。在某些方面,mhc ii类分子包含dqα链,所述dqα链包含与seq id no:18具有至少约70%、至少约75%、至少约80%、至少约85%、至少约90%、至少约95%、至少约96%、至少约97%、至少约98%、至少约99%或约100%序列同一性的氨基酸序列。在某些方面,mhc ii类分子包含dqα链,所述dqα链包含seq id no:16所示的氨基酸序列。在某些方面,mhc ii类分子包含dqα链,所述dqα链包含seq id no:18所示的氨基酸序列。
[0236]
ii.a.3.hla-dr分子
[0237]
许多hla-dr等位基因在本领域中是已知的,并且任何已知的等位基因都可以用在本公开中。hla-drα链和β链等位基因的示例如表5所示。hla等位基因的更新清单可在hla.alleles.org/(上次访问时间为2019年7月10)上获得。
[0238]
表5:drβ链和α链氨基酸和核苷酸序列。
[0239]
[0240]
[0241]
[0242][0243]
ii.a.3.a.hla-drβ链
[0244]
在某些方面,hla ii类分子包含drβ链,其中所述drβ链包含在对应于seq id no:19的氨基酸残基114的位置处的除亮氨酸之外的氨基酸。任何除亮氨酸之外的氨基酸都可以存在于对应于seq id no:19的氨基酸残基114的位置处。在一些方面,除亮氨酸之外的氨
基酸是包含疏水侧链的氨基酸。在某些方面,在对应于seq id no:19的氨基酸残基114的位置处的除亮氨酸之外的氨基酸是选自丙氨酸、缬氨酸、异亮氨酸、甲硫氨酸、苯丙氨酸、酪氨酸、和色氨酸的氨基酸。在某些方面,在对应于seq id no:19的氨基酸残基114的位置处的除亮氨酸之外的氨基酸是丙氨酸。在某些方面,在对应于seq id no:19的氨基酸残基114的位置处的除亮氨酸之外的氨基酸是缬氨酸。在某些方面,在对应于seq id no:19的氨基酸残基114的位置处的除亮氨酸之外的氨基酸是异亮氨酸。在某些方面,在对应于seq id no:19的氨基酸残基114的位置处的除亮氨酸之外的氨基酸是甲硫氨酸。在某些方面,在对应于seq id no:19的氨基酸残基114的位置处的除亮氨酸之外的氨基酸是苯丙氨酸。在某些方面,在对应于seq id no:19的氨基酸残基114的位置处的除亮氨酸之外的氨基酸是酪氨酸。在某些方面,在对应于seq id no:19的氨基酸残基114的位置处的除亮氨酸之外的氨基酸是色氨酸。
[0245]
在一些实施方案中,在对应于seq id no:19的氨基酸残基114的位置处的除亮氨酸之外的氨基酸由多于一个氨基酸组成,例如,两个氨基酸、三个氨基酸、四个氨基酸、五个氨基酸或者更多。在一些方面,多于一个氨基酸中的至少一个包含疏水侧链。在某些方面,在对应于seq id no:19的氨基酸残基114的位置处的除亮氨酸之外的氨基酸由一系列,例如至少2个、至少3个、至少4个或至少5个氨基酸组成,其中所述系列氨基酸中的每一个都包含疏水侧链。
[0246]
在某些方面,hla ii类分子包含drβ链,其中所述drβ链包含在对应于seq id no:19的氨基酸残基143的位置处的除缬氨酸之外的氨基酸。任何除缬氨酸之外的氨基酸都可存在于对应于seq id no:19的氨基酸残基143的位置处。在一些方面,除缬氨酸之外的氨基酸是包含疏水侧链的氨基酸。在某些方面,在对应于seq id no:19的氨基酸残基143的位置处的除缬氨酸之外的氨基酸是选自丙氨酸、异亮氨酸、亮氨酸、甲硫氨酸、苯丙氨酸、酪氨酸、和色氨酸的氨基酸。在某些方面,在对应于seq id no:19的氨基酸残基143的位置处的除缬氨酸之外的氨基酸是丙氨酸。在某些方面,在对应于seq id no:19的氨基酸残基143的位置处的除缬氨酸之外的氨基酸是异亮氨酸。在某些方面,在对应于seq id no:19的氨基酸残基143的位置处的除缬氨酸之外的氨基酸是亮氨酸。在某些方面,在对应于seq id no:19的氨基酸残基143的位置处的除缬氨酸之外的氨基酸是甲硫氨酸。在某些方面,在对应于seq id no:19的氨基酸残基143的位置处的除缬氨酸之外的氨基酸是苯丙氨酸。在某些方面,在对应于seq id no:19的氨基酸残基143的位置处的除缬氨酸之外的氨基酸是酪氨酸。在某些方面,在对应于seq id no:19的氨基酸残基143的位置处的除缬氨酸之外的氨基酸是色氨酸。
[0247]
在一些方面,在对应于seq id no:19的氨基酸残基143的位置处的除缬氨酸之外的氨基酸由多于一个氨基酸组成,例如,两个氨基酸、三个氨基酸、四个氨基酸、五个氨基酸或者更多。在一些方面,多于一个氨基酸中的至少一个包含疏水侧链。在某些方面,在对应于seq id no:19的氨基酸残基143的位置处的除缬氨酸之外的氨基酸由一系列,例如至少2个、至少3个、至少4个或至少5个氨基酸组成,其中所述系列氨基酸中的每一个都包含疏水侧链。
[0248]
在某些方面,hla ii类分子包含drβ链,其中所述drβ链包含在对应于seq id no:19的氨基酸残基118的位置处的除丝氨酸之外的氨基酸。任何除丝氨酸之外的氨基酸都可
以存在于对应于seq id no:19的氨基酸残基118的位置处。在一些方面,除丝氨酸之外的氨基酸是包含带电侧链的氨基酸。在某些方面,在对应于seq id no:19的氨基酸残基118的位置处的除丝氨酸之外的氨基酸是选自精氨酸、组氨酸和赖氨酸的氨基酸。在某些方面,在对应于seq id no:19的氨基酸残基118的位置处的除丝氨酸之外的氨基酸是精氨酸。在某些方面,在对应于seq id no:19的氨基酸残基118的位置处的除丝氨酸之外的氨基酸是组氨酸。在某些方面,在对应于seq id no:19的氨基酸残基118的位置处的除丝氨酸之外的氨基酸是赖氨酸。
[0249]
在一些方面,在对应于seq id no:19的氨基酸残基118的位置处的除丝氨酸之外的氨基酸由多于一个氨基酸组成,例如,两个氨基酸、三个氨基酸、四个氨基酸、五个氨基酸或者更多。在一些方面,多于一个氨基酸中的至少一个包含带电侧链。在某些方面,在对应于seq id no:19的氨基酸残基118的位置处的除丝氨酸之外的氨基酸由一系列,例如至少2个、至少3个、至少4个或至少5个氨基酸组成,其中所述系列氨基酸中的每一个都包含带电侧链。
[0250]
在某些方面,hla ii类分子包含drβ链,其中所述drβ链包含在对应于seq id no:19的氨基酸残基157的位置处的除苏氨酸之外的氨基酸。任何除苏氨酸之外的氨基酸都可以存在于对应于seq id no:19的氨基酸残基157的位置处。在一些方面,除苏氨酸之外的氨基酸是包含疏水侧链的氨基酸。在某些方面,在对应于seq id no:19的氨基酸残基157的位置处的除苏氨酸之外的氨基酸是选自丙氨酸、缬氨酸、异亮氨酸、亮氨酸、甲硫氨酸、苯丙氨酸、酪氨酸、和色氨酸的氨基酸。在某些方面,在对应于seq id no:19的氨基酸残基157的位置处的除苏氨酸之外的氨基酸是丙氨酸。在某些方面,在对应于seq id no:19的氨基酸残基157的位置处的除苏氨酸之外的氨基酸是缬氨酸。在某些方面,在对应于seq id no:19的氨基酸残基157的位置处的除苏氨酸之外的氨基酸是异亮氨酸。在某些方面,在对应于seq id no:19的氨基酸残基157的位置处的除苏氨酸之外的氨基酸是亮氨酸。在某些方面,在对应于seq id no:19的氨基酸残基157的位置处的除苏氨酸之外的氨基酸是甲硫氨酸。在某些方面,在对应于seq id no:19的氨基酸残基157的位置处的除苏氨酸之外的氨基酸是苯丙氨酸。在某些方面,在对应于seq id no:19的氨基酸残基157的位置处的除苏氨酸之外的氨基酸是酪氨酸。在某些方面,在对应于seq id no:19的氨基酸残基157的位置处的除苏氨酸之外的氨基酸是色氨酸。
[0251]
在一些方面,在对应于seq id no:19的氨基酸残基157的位置处的除苏氨酸之外的氨基酸由多于一个氨基酸组成,例如,两个氨基酸、三个氨基酸、四个氨基酸、五个氨基酸或者更多。在一些方面,多于一个氨基酸中的至少一个包含疏水侧链。在某些方面,在对应于seq id no:19的氨基酸残基157的位置处的除苏氨酸之外的氨基酸由一系列,例如至少2个、至少3个、至少4个或至少5个氨基酸组成,其中所述系列氨基酸中的每一个都包含疏水侧链。
[0252]
在某些方面,hla ii类分子包含drβ链,其中所述drβ链包含在对应于seq id no:19的氨基酸残基139的位置处的除赖氨酸之外的氨基酸。任何除赖氨酸之外的氨基酸都可以存在于对应于seq id no:19的氨基酸残基139的位置处。在一些方面,除赖氨酸之外的氨基酸是包含极性不带电侧链的氨基酸。在某些方面,在对应于seq id no:19的氨基酸残基139的位置处的除赖氨酸之外的氨基酸是选自丝氨酸、苏氨酸和谷氨酰胺的氨基酸。在某些
方面,在对应于seq id no:19的氨基酸残基139的位置处的除赖氨酸之外的氨基酸是丝氨酸。在某些方面,在对应于seq id no:19的氨基酸残基139的位置处的除赖氨酸之外的氨基酸是苏氨酸。在某些方面,在对应于seq id no:19的氨基酸残基139的位置处的除赖氨酸之外的氨基酸是谷氨酰胺。
[0253]
在一些方面,在对应于seq id no:19的氨基酸残基139的位置处的除赖氨酸之外的氨基酸由多于一个氨基酸组成,例如,两个氨基酸、三个氨基酸、四个氨基酸、五个氨基酸或者更多。在一些方面,多于一个氨基酸中的至少一个包含极性不带电侧链。在某些方面,在对应于seq id no:19的氨基酸残基139的位置处的除赖氨酸之外的氨基酸由一系列,例如至少2个、至少3个、至少4个或至少5个氨基酸组成,其中所述系列氨基酸中的每一个都包含极性不带电侧链。
[0254]
在某些方面,hla ii类分子包含drβ链,其中所述drβ链包含在对应于seq id no:19的氨基酸残基146的位置处的除甘氨酸之外的氨基酸。任何除甘氨酸之外的氨基酸都可以存在于对应于seq id no:19的氨基酸残基146的位置处。在一些方面,除甘氨酸之外的氨基酸是包含极性不带电侧链的氨基酸。在某些方面,在对应于seq id no:19的氨基酸残基146的位置处的除甘氨酸之外的氨基酸是选自丝氨酸、天冬酰胺、苏氨酸和谷氨酰胺的氨基酸。在某些方面,在对应于seq id no:19的氨基酸残基146的位置处的除甘氨酸之外的氨基酸是丝氨酸。在某些方面,在对应于seq id no:19的氨基酸残基146的位置处的除甘氨酸之外的氨基酸是天冬酰胺。在某些方面,在对应于seq id no:19的氨基酸残基146的位置处的除甘氨酸之外的氨基酸是苏氨酸。在某些方面,在对应于seq id no:19的氨基酸残基146的位置处的除甘氨酸之外的氨基酸是谷氨酰胺。
[0255]
在一些方面,在对应于seq id no:19的氨基酸残基146的位置处的除甘氨酸之外的氨基酸由多于一个氨基酸组成,例如,两个氨基酸、三个氨基酸、四个氨基酸、五个氨基酸或者更多。在一些方面,多于一个氨基酸中的至少一个包含极性不带电侧链。在某些方面,在对应于seq id no:19的氨基酸残基146的位置处的除甘氨酸之外的氨基酸由一系列,例如至少2个、至少3个、至少4个或至少5个氨基酸组成,其中所述系列氨基酸中的每一个都包含极性不带电侧链。
[0256]
在某些方面,hla ii类分子包含drβ链,其中所述drβ链包含在对应于seq id no:19的氨基酸残基163的位置处的除苏氨酸之外的氨基酸。任何除苏氨酸之外的氨基酸都可以存在于对应于seq id no:19的氨基酸残基163的位置处。在一些方面,除苏氨酸之外的氨基酸是包含疏水侧链的氨基酸。在某些方面,在对应于seq id no:19的氨基酸残基163的位置处的除苏氨酸之外的氨基酸是选自丙氨酸、缬氨酸、异亮氨酸、亮氨酸、甲硫氨酸、苯丙氨酸、酪氨酸、和色氨酸的氨基酸。在某些方面,在对应于seq id no:19的氨基酸残基163的位置处的除苏氨酸之外的氨基酸是丙氨酸。在某些方面,在对应于seq id no:19的氨基酸残基163的位置处的除苏氨酸之外的氨基酸是缬氨酸。在某些方面,在对应于seq id no:19的氨基酸残基163的位置处的除苏氨酸之外的氨基酸是异亮氨酸。在某些方面,在对应于seq id no:19的氨基酸残基163的位置处的除苏氨酸之外的氨基酸是亮氨酸。在某些方面,在对应于seq id no:19的氨基酸残基163的位置处的除苏氨酸之外的氨基酸是甲硫氨酸。在某些方面,在对应于seq id no:19的氨基酸残基163的位置处的除苏氨酸之外的氨基酸是苯丙氨酸。在某些方面,在对应于seq id no:19的氨基酸残基163的位置处的除苏氨酸之外的
氨基酸是酪氨酸。在某些方面,在对应于seq id no:19的氨基酸残基163的位置处的除苏氨酸之外的氨基酸是色氨酸。
[0257]
在一些方面,在对应于seq id no:19的氨基酸残基163的位置处的除苏氨酸之外的氨基酸由多于一个氨基酸组成,例如,两个氨基酸、三个氨基酸、四个氨基酸、五个氨基酸或者更多。在一些方面,多于一个氨基酸中的至少一个包含疏水侧链。在某些方面,在对应于seq id no:19的氨基酸残基163的位置处的除苏氨酸之外的氨基酸由一系列,例如至少2个、至少3个、至少4个或至少5个氨基酸组成,其中所述系列氨基酸中的每一个都包含疏水侧链。
[0258]
在某些方面,hla ii类分子包含drβ链,其中所述drβ链包含在对应于seq id no:19的氨基酸残基164的位置处的除缬氨酸之外的氨基酸。任何除缬氨酸之外的氨基酸都可存在于对应于seq id no:19的氨基酸残基164的位置处。在一些方面,除缬氨酸之外的氨基酸是包含极性不带电侧链的氨基酸。在某些方面,在对应于seq id no:19的氨基酸残基164的位置处的除缬氨酸之外的氨基酸是选自丝氨酸、天冬酰胺、苏氨酸和谷氨酰胺的氨基酸。在某些方面,在对应于seq id no:19的氨基酸残基164的位置处的除缬氨酸之外的氨基酸是丝氨酸。在某些方面,在对应于seq id no:19的氨基酸残基164的位置处的除缬氨酸之外的氨基酸是天冬酰胺。在某些方面,在对应于seq id no:19的氨基酸残基164的位置处的除缬氨酸之外的氨基酸是苏氨酸。在某些方面,在对应于seq id no:19的氨基酸残基164的位置处的除缬氨酸之外的氨基酸是谷氨酰胺。
[0259]
在一些方面,在对应于seq id no:19的氨基酸残基164的位置处的除缬氨酸之外的氨基酸由多于一个氨基酸组成,例如,两个氨基酸、三个氨基酸、四个氨基酸、五个氨基酸或者更多。在一些方面,多于一个氨基酸中的至少一个包含极性不带电侧链。在某些方面,在对应于seq id no:19的氨基酸残基164的位置处的除缬氨酸之外的氨基酸由一系列,例如至少2个、至少3个、至少4个或至少5个氨基酸组成,其中所述系列氨基酸中的每一个都包含极性不带电侧链。
[0260]
在本公开的某些方面,mhc ii类分子包含相对于野生型drβ链含有多于一个取代突变的drβ链。在某些方面,drβ链相对于野生型drβ链包含至少两个突变、至少三个突变、至少四个突变、至少五个突变、至少六个突变、至少七个突变、至少八个突变、至少九个突变、或至少十个突变。
[0261]
在某些方面,drβ链包含在对应于seq id no:19的氨基酸残基114的位置处的除亮氨酸之外的氨基酸和在对应于seq id no:19的氨基酸残基143的位置处的除缬氨酸之外的氨基酸。
[0262]
在某些方面,drβ链包含(a)在对应于seq id no:19的氨基酸残基114的位置处的除亮氨酸之外的氨基酸,(b)在对应于seq id no:19的氨基酸残基143的位置处的除缬氨酸之外的氨基酸,(c)在对应于seq id no:19的氨基酸残基118的位置处的除丝氨酸之外的氨基酸,和(d)在对应于seq id no:19的氨基酸残基157的位置处的除苏氨酸之外的氨基酸。
[0263]
在某些方面,drβ链包含在对应于seq id no:19的氨基酸残基114的位置处的除亮氨酸之外的氨基酸;在对应于seq id no:19的氨基酸残基143的位置处的除缬氨酸之外的氨基酸;以及下列中的至少两项:(i)在对应于seq id no:19的氨基酸残基118的位置处的除丝氨酸之外的氨基酸,(ii)在对应于seq id no:19的氨基酸残基139的位置处的除赖氨
酸之外的氨基酸,(iii)在对应于seq id no:19的氨基酸残基146的位置处的除甘氨酸之外的氨基酸,(iv)在对应于seq id no:19的氨基酸残基157的位置处的除苏氨酸之外的氨基酸,(v)在对应于seq id no:19的氨基酸残基163的位置处的除苏氨酸之外的氨基酸,和(vi)在对应于seq id no:19的氨基酸残基164的位置处的除缬氨酸之外的氨基酸。
[0264]
在某些方面,drβ链包含在对应于seq id no:19的氨基酸残基114的位置处的除亮氨酸之外的氨基酸;在对应于seq id no:19的氨基酸残基143的位置处的除缬氨酸之外的氨基酸;以及下列中的至少三项:(i)在对应于seq id no:19的氨基酸残基118的位置处的除丝氨酸之外的氨基酸,(ii)在对应于seq id no:19的氨基酸残基139的位置处的除赖氨酸之外的氨基酸,(iii)在对应于seq id no:19的氨基酸残基146的位置处的除甘氨酸之外的氨基酸,(iv)在对应于seq id no:19的氨基酸残基157的位置处的除苏氨酸之外的氨基酸,(v)在对应于seq id no:19的氨基酸残基163的位置处的除苏氨酸之外的氨基酸,和(vi)在对应于seq id no:19的氨基酸残基164的位置处的除缬氨酸之外的氨基酸。
[0265]
在某些方面,drβ链包含在对应于seq id no:19的氨基酸残基114的位置处的除亮氨酸之外的氨基酸;在对应于seq id no:19的氨基酸残基143的位置处的除缬氨酸之外的氨基酸;以及下列中的至少四项:(i)在对应于seq id no:19的氨基酸残基118的位置处的除丝氨酸之外的氨基酸,(ii)在对应于seq id no:19的氨基酸残基139的位置处的除赖氨酸之外的氨基酸,(iii)在对应于seq id no:19的氨基酸残基146的位置处的除甘氨酸之外的氨基酸,(iv)在对应于seq id no:19的氨基酸残基157的位置处的除苏氨酸之外的氨基酸,(v)在对应于seq id no:19的氨基酸残基163的位置处的除苏氨酸之外的氨基酸,和(vi)在对应于seq id no:19的氨基酸残基164的位置处的除缬氨酸之外的氨基酸。
[0266]
在某些方面,drβ链包含(a)在对应于seq id no:19的氨基酸残基114的位置处的除亮氨酸之外的氨基酸,(b)在对应于seq id no:19的氨基酸残基143的位置处的除缬氨酸之外的氨基酸,(c)在对应于seq id no:19的氨基酸残基118的位置处的除丝氨酸之外的氨基酸,和(d)在对应于seq id no:19的氨基酸残基157的位置处的除苏氨酸之外的氨基酸;以及下列中的至少一项:(i)在对应于seq id no:19的氨基酸残基139的位置处的除赖氨酸之外的氨基酸,(ii)在对应于seq id no:19的氨基酸残基146的位置处的除甘氨酸之外的氨基酸,(iii)在对应于seq id no:19的氨基酸残基163的位置处的除苏氨酸之外的氨基酸,和(iv)在对应于seq id no:19的氨基酸残基164的位置处的除缬氨酸之外的氨基酸。
[0267]
在某些方面,drβ链包含(a)在对应于seq id no:19的氨基酸残基114的位置处的除亮氨酸之外的氨基酸,(b)在对应于seq id no:19的氨基酸残基143的位置处的除缬氨酸之外的氨基酸,(c)在对应于seq id no:19的氨基酸残基118的位置处的除丝氨酸之外的氨基酸,和(d)在对应于seq id no:19的氨基酸残基157的位置处的除苏氨酸之外的氨基酸;以及下列中的至少两项:(i)在对应于seq id no:19的氨基酸残基139的位置处的除赖氨酸之外的氨基酸,(ii)在对应于seq id no:19的氨基酸残基146的位置处的除甘氨酸之外的氨基酸,(iii)在对应于seq id no:19的氨基酸残基163的位置处的除苏氨酸之外的氨基酸,和(iv)在对应于seq id no:19的氨基酸残基164的位置处的除缬氨酸之外的氨基酸。
[0268]
在某些方面,drβ链包含(a)在对应于seq id no:19的氨基酸残基114的位置处的除亮氨酸之外的氨基酸,(b)在对应于seq id no:19的氨基酸残基143的位置处的除缬氨酸之外的氨基酸,(c)在对应于seq id no:19的氨基酸残基118的位置处的除丝氨酸之外的氨
基酸,和(d)在对应于seq id no:19的氨基酸残基157的位置处的除苏氨酸之外的氨基酸;以及下列中的至少三项:(i)在对应于seq id no:19的氨基酸残基139的位置处的除赖氨酸之外的氨基酸,(ii)在对应于seq id no:19的氨基酸残基146的位置处的除甘氨酸之外的氨基酸,(iii)在对应于seq id no:19的氨基酸残基163的位置处的除苏氨酸之外的氨基酸,和(iv)在对应于seq id no:19的氨基酸残基164的位置处的除缬氨酸之外的氨基酸。
[0269]
在某些方面,drβ链包含(a)在对应于seq id no:19的氨基酸残基114的位置处的除亮氨酸之外的氨基酸,(b)在对应于seq id no:19的氨基酸残基143的位置处的除缬氨酸之外的氨基酸,(c)在对应于seq id no:19的氨基酸残基118的位置处的除丝氨酸之外的氨基酸,(d)在对应于seq id no:19的氨基酸残基139的位置处的除赖氨酸之外的氨基酸,(e)在对应于seq id no:19的氨基酸残基146的位置处的除甘氨酸之外的氨基酸,(f)在对应于seq id no:19的氨基酸残基157的位置处的除苏氨酸之外的氨基酸,(g)在对应于seq id no:19的氨基酸残基163的位置处的除苏氨酸之外的氨基酸,和(h)在对应于seq id no:19的氨基酸残基164的位置处的除缬氨酸之外的氨基酸。
[0270]
在某些方面,drβ链包含(a)在对应于seq id no:19的氨基酸残基114的位置处的色氨酸,(b)在对应于seq id no:19的氨基酸残基143的位置处的甲硫氨酸,(c)在对应于seq id no:19的氨基酸残基118的位置处的组氨酸,和(d)在对应于seq id no:19的氨基酸残基157的位置处的异亮氨酸。
[0271]
在一些方面,(i)在对应于seq id no:19的氨基酸残基114的位置处的除亮氨酸之外的氨基酸,(ii)在对应于seq id no:19的氨基酸残基143的位置处的除缬氨酸之外的氨基酸,或者在对应于seq id no:19的氨基酸残基114的位置处的除亮氨酸之外的氨基酸和在对应于seq id no:19的氨基酸残基143的位置处的除缬氨酸之外的氨基酸中的每一个是包含疏水侧链的氨基酸。
[0272]
在一些方面,(i)在对应于seq id no:19的氨基酸残基114的位置处的除亮氨酸之外的氨基酸选自丙氨酸、缬氨酸、异亮氨酸、甲硫氨酸、苯丙氨酸、酪氨酸和色氨酸;(ii)在对应于seq id no:19的氨基酸残基143的位置处的除缬氨酸之外的氨基酸选自丙氨酸、异亮氨酸、亮氨酸、甲硫氨酸、苯丙氨酸、酪氨酸和色氨酸;(iii)在对应于seq id no:19的氨基酸残基118的位置处的除丝氨酸之外的氨基酸选自精氨酸、组氨酸和赖氨酸;和/或(iv)在对应于seq id no:19的氨基酸残基157的位置处的除苏氨酸之外的氨基酸选自丙氨酸、缬氨酸、异亮氨酸、亮氨酸、甲硫氨酸、苯丙氨酸、酪氨酸和色氨酸。
[0273]
在一些方面,(i)在对应于seq id no:19的氨基酸残基114的位置处的除亮氨酸之外的氨基酸选自丙氨酸、缬氨酸、异亮氨酸、甲硫氨酸、苯丙氨酸、酪氨酸和色氨酸;(ii)在对应于seq id no:19的氨基酸残基143的位置处的除缬氨酸之外的氨基酸选自丙氨酸、异亮氨酸、亮氨酸、甲硫氨酸、苯丙氨酸、酪氨酸和色氨酸;(iii)在对应于seq id no:19的氨基酸残基118的位置处的除丝氨酸之外的氨基酸选自精氨酸、组氨酸和赖氨酸;(iv)在对应于seq id no:19的氨基酸残基157的位置处的除苏氨酸之外的氨基酸选自丙氨酸、缬氨酸、异亮氨酸、亮氨酸、甲硫氨酸、苯丙氨酸、酪氨酸和色氨酸;(v)在对应于seq id no:19的氨基酸残基139的位置处的除赖氨酸之外的氨基酸选自丝氨酸、苏氨酸和谷氨酰胺;(vi)在对应于seq id no:19的氨基酸残基146的位置处的除甘氨酸之外的氨基酸选自丝氨酸、天冬酰胺、苏氨酸和谷氨酰胺;(vii)在对应于seq id no:19的氨基酸残基163的位置处的除苏
氨酸之外的氨基酸选自丙氨酸、缬氨酸、异亮氨酸、亮氨酸、甲硫氨酸、苯丙氨酸、酪氨酸和色氨酸;和/或(viii)在对应于seq id no:19的氨基酸残基164的位置处的除缬氨酸之外的氨基酸选自丝氨酸、天冬酰胺、苏氨酸和谷氨酰胺。
[0274]
在某些方面,与参考hla ii类分子相比,本文所述的drβ链对cd4蛋白具有增加的亲和力。在一些方面,参考hla ii类分子是具有野生型drβ链的hla ii类分子。在一些方面,参考hla ii类分子是具有drβ链的hla ii类分子,所述drβ链包含(i)在对应于seq id no:19的氨基酸残基114的位置处的亮氨酸和/或(ii)在对应于seq id no:19的氨基酸残基143的位置处的缬氨酸。在一些方面,参考hla ii类分子是具有drβ链的hla ii类分子,所述drβ链包含(i)在对应于seq id no:19的氨基酸残基114的位置处的亮氨酸,(ii)在对应于seq id no:19的氨基酸残基143的位置处的缬氨酸,(iii)在对应于seq id no:19的氨基酸残基118的位置处的丝氨酸,和(iv)在对应于seq id no:19的氨基酸残基157的位置处的苏氨酸。
[0275]
在一些方面,增加的对cd4的亲和力是参考hla ii类分子对cd4的亲和力的至少约1.5倍、至少约2倍、至少约3倍、至少约4倍、至少约5倍、至少约6倍、至少约7倍、至少约8倍、至少约9倍、至少约10倍、至少约15倍、至少约20倍、至少约25倍、至少约30倍、至少约35倍、至少约40倍、至少约45倍、至少约50倍、至少约75倍、至少约100倍、至少约200倍、至少约300倍、至少约400倍、至少约500倍、至少约1000倍、至少约1500倍、至少约2000倍、至少约2500倍、至少约3000倍、至少约3500倍、至少约4000倍、至少约4500倍、或至少约4000倍。
[0276]
在一些方面,增加的对cd4的亲和力是参考hla ii类分子对cd4的亲和力的至少约1.5倍到至少约5000倍、1.5倍到至少约4000倍、1.5倍到至少约3000倍、1.5倍到至少约2000倍、1.5倍到至少约1000倍、10倍到至少约5000倍、10倍到至少约4000倍、10倍到至少约3000倍、10倍到至少约2000倍、10倍到至少约1000倍、10倍到至少约900倍、10倍到至少约800倍、10倍到至少约700倍、10倍到至少约600倍、10倍到至少约500倍、10倍到至少约400倍、10倍到至少约300倍、10倍到至少约200倍、10倍到至少约100倍、100倍到至少约5000倍、100倍到至少约4000倍、100倍到至少约3000倍、100倍到至少约2000倍、100倍到至少约1000倍、100倍到至少约900倍、100倍到至少约800倍、100倍到至少约700倍、100倍到至少约600倍、100倍到至少约500倍、100倍到至少约400倍、100倍到至少约300倍、或100倍到至少约200倍。
[0277]
在某些方面,drβ链包含选自以下的等位基因:hla-drb1*01、hla-drb1*03、hla-drb1*04、hla-drb1*06、hla-drb1*07、hla-drb1*08、hla-drb1*09、hla-drb1*10、hla-drb1*11、hla-drb1*12、hla-drb1*13、hla-drb1*14、hla-drb1*15、或hla-drb1*16等位基因。在一些方面,drβ链包含hla-drb1*01等位基因。在特定方面,drβ链包含hla-drb1*01:01等位基因。
[0278]
在某些方面,drβ链包含选自以下的等位基因:drb1*01:01:01、drb1*01:01:02、drb1*01:01:03、drb1*01:01:04、drb1*01:01:05、drb1*01:01:06、drb1*01:01:07、drb1*01:01:08、drb1*01:01:09、drb1*01:01:10、drb1*01:01:11、drb1*01:01:12、drb1*01:01:13、drb1*01:01:14、drb1*01:01:15、drb1*01:01:16、drb1*01:01:17、drb1*01:01:18、drb1*01:01:19、drb1*01:01:20、drb1*01:01:21、drb1*01:01:22、drb1*01:01:23、drb1*01:01:24、drb1*01:01:25、drb1*01:01:26、drb1*01:01:27、drb1*01:01:28、drb1*01:01:29、drb1*01:01:30、drb1*01:01:31、drb1*01:01:32、drb1*01:01:33、drb1*01:02:01:01、
drb1*01:02:01:02、drb1*01:02:02、drb1*01:02:03、drb1*01:02:04、drb1*01:02:05、drb1*01:02:06、drb1*01:02:07、drb1*01:02:08、drb1*01:02:09、drb1*01:02:10、drb1*01:02:11、drb1*01:02:12、drb1*01:02:13、drb1*01:03:01、drb1*01:03:02、drb1*01:03:03、drb1*01:03:04、drb1*01:04、drb1*01:05、drb1*01:06、drb1*01:07、drb1*01:08、drb1*01:09、drb1*01:10、drb1*01:100、drb1*01:11:01、drb1*01:11:02、drb1*01:12、drb1*01:13、drb1*01:14、drb1*01:15、drb1*01:16、drb1*01:17、drb1*01:18:01、drb1*01:18:02、drb1*01:19、drb1*01:20:01、drb1*01:20:02、drb1*01:21、drb1*01:22、drb1*01:23、drb1*01:24:01、drb1*01:24:02、drb1*01:25、drb1*01:26、drb1*01:27、drb1*01:28、drb1*01:29:01、drb1*01:29:02、drb1*01:30、drb1*01:31、drb1*01:32、drb1*01:33n、drb1*01:34、drb1*01:35、drb1*01:36、drb1*01:37、drb1*01:38、drb1*01:39n、drb1*01:40n、drb1*01:41、drb1*01:42、drb1*01:43、drb1*01:44:01、drb1*01:44:02、drb1*01:45、drb1*01:46、drb1*01:47、drb1*01:48、drb1*01:49、drb1*01:50、drb1*01:51、drb1*01:52n、drb1*01:53、drb1*01:54、drb1*01:55、drb1*01:56、drb1*01:57、drb1*01:58、drb1*01:59、drb1*01:60、drb1*01:61、drb1*01:62n、drb1*01:63、drb1*01:64、drb1*01:65:01、drb1*01:65:02、drb1*01:66、drb1*01:67、drb1*01:68n、drb1*01:69、drb1*01:70、drb1*01:71、drb1*01:72、drb1*01:73、drb1*01:74、drb1*01:75、drb1*01:76、drb1*01:77、drb1*01:78、drb1*01:79、drb1*01:80、drb1*01:81、drb1*01:82、drb1*01:83、drb1*01:84、drb1*01:85、drb1*01:86、drb1*01:87、drb1*01:88、drb1*01:89、drb1*01:90、drb1*01:91q、drb1*01:92、drb1*01:93、drb1*01:94、drb1*01:95、drb1*01:96、drb1*01:97、drb1*01:98、drb1*01:99、drb1*03:01:01:01、drb1*03:01:01:02、drb1*03:01:01:03、drb1*03:01:02、drb1*03:01:03、drb1*03:01:04、drb1*03:01:05、drb1*03:01:06、drb1*03:01:07、drb1*03:01:08、drb1*03:01:09、drb1*03:01:10、drb1*03:01:11、drb1*03:01:12、drb1*03:01:13、drb1*03:01:14、drb1*03:01:15、drb1*03:01:16、drb1*03:01:17、drb1*03:01:18、drb1*03:01:19、drb1*03:01:20、drb1*03:01:21、drb1*03:01:22、drb1*03:01:23、drb1*03:01:24、drb1*03:01:25、drb1*03:01:26、drb1*03:01:27、drb1*03:01:28、drb1*03:02:01、drb1*03:02:02、drb1*03:02:03、drb1*03:03、drb1*03:04:01、drb1*03:04:02、drb1*03:05:01、drb1*03:05:02、drb1*03:05:03、drb1*03:06、drb1*03:07:01、drb1*03:07:02、drb1*03:08、drb1*03:09、drb1*03:10、drb1*03:100:01、drb1*03:100:02、drb1*03:101、drb1*03:102、drb1*03:103、drb1*03:104、drb1*03:105、drb1*03:106、drb1*03:107、drb1*03:108、drb1*03:109、drb1*03:110、drb1*03:111、drb1*03:112、drb1*03:113、drb1*03:114、drb1*03:115、drb1*03:116、drb1*03:117、drb1*03:118、drb1*03:119、drb1*03:11:01、drb1*03:12、drb1*03:120、drb1*03:121、drb1*03:122、drb1*03:123、drb1*03:124、drb1*03:125、drb1*03:126、drb1*03:127、drb1*03:128、drb1*03:129、drb1*03:130、drb1*03:131、drb1*03:132、drb1*03:133、drb1*03:134、drb1*03:135、drb1*03:136、drb1*03:137、drb1*03:138、drb1*03:139、drb1*03:13:01、drb1*03:13:02、drb1*03:14、drb1*03:140、drb1*03:141、drb1*03:142、drb1*03:143、drb1*03:144、drb1*03:145、drb1*03:146、drb1*03:147、drb1*03:148、drb1*03:149、drb1*03:150、drb1*03:151、drb1*03:152、drb1*03:153、drb1*03:154、drb1*03:155、drb1*03:156n、drb1*03:157、drb1*03:158、drb1*03:15:01、drb1*03:15:02、drb1*03:16、drb1*03:17、drb1*03:18、drb1*03:19、drb1*03:20、drb1*03:21、drb1*
03:22、drb1*03:23、drb1*03:24、drb1*03:25:01、drb1*03:25:02、drb1*03:26、drb1*03:27、drb1*03:28、drb1*03:29、drb1*03:30、drb1*03:31、drb1*03:32、drb1*03:33、drb1*03:34、drb1*03:35、drb1*03:36、drb1*03:37、drb1*03:38、drb1*03:39、drb1*03:40、drb1*03:41:01、drb1*03:41:02、drb1*03:42、drb1*03:43、drb1*03:44、drb1*03:45、drb1*03:46、drb1*03:47、drb1*03:48、drb1*03:49、drb1*03:50、drb1*03:51、drb1*03:52、drb1*03:53、drb1*03:54、drb1*03:55、drb1*03:56、drb1*03:57、drb1*03:58、drb1*03:59、drb1*03:60、drb1*03:61、drb1*03:62、drb1*03:63、drb1*03:64、drb1*03:65、drb1*03:66、drb1*03:67n、drb1*03:68n、drb1*03:69、drb1*03:70、drb1*03:71:01、drb1*03:71:02、drb1*03:72、drb1*03:73、drb1*03:74、drb1*03:75、drb1*03:76、drb1*03:77、drb1*03:78、drb1*03:79、drb1*03:80、drb1*03:81、drb1*03:82、drb1*03:83、drb1*03:84、drb1*03:85、drb1*03:86、drb1*03:87、drb1*03:88、drb1*03:89、drb1*03:90、drb1*03:91、drb1*03:92、drb1*03:93、drb1*03:94、drb1*03:95、drb1*03:96、drb1*03:97、drb1*03:98、drb1*03:99、drb1*04:01:01:01、drb1*04:01:01:02、drb1*04:01:01:03、drb1*04:01:02、drb1*04:01:03、drb1*04:01:04、drb1*04:01:05、drb1*04:01:06、drb1*04:01:07、drb1*04:01:08、drb1*04:01:09、drb1*04:01:10、drb1*04:01:11、drb1*04:01:12、drb1*04:01:13、drb1*04:01:14、drb1*04:01:15、drb1*04:01:16、drb1*04:01:17、drb1*04:01:18、drb1*04:01:19、drb1*04:01:20、drb1*04:01:21、drb1*04:02:01、drb1*04:02:02、drb1*04:02:03、drb1*04:02:04、drb1*04:02:05、drb1*04:02:06、drb1*04:03:01:01、drb1*04:03:01:02、drb1*04:03:02、drb1*04:03:03、drb1*04:03:04、drb1*04:03:05、drb1*04:03:06、drb1*04:03:07、drb1*04:03:08、drb1*04:03:09、drb1*04:03:10、drb1*04:03:11、drb1*04:03:12、drb1*04:03:13、drb1*04:03:14、drb1*04:03:15、drb1*04:04:01、drb1*04:04:02、drb1*04:04:03、drb1*04:04:04、drb1*04:04:05、drb1*04:04:06、drb1*04:04:07、drb1*04:04:08、drb1*04:04:09、drb1*04:04:10、drb1*04:04:11、drb1*04:04:12、drb1*04:04:13、drb1*04:04:14、drb1*04:04:15、drb1*04:05:01:01、drb1*04:05:01:02、drb1*04:05:01:03、drb1*04:05:02、drb1*04:05:03、drb1*04:05:04、drb1*04:05:05、drb1*04:05:06、drb1*04:05:07、drb1*04:05:08、drb1*04:05:09、drb1*04:05:10、drb1*04:05:11、drb1*04:05:13、drb1*04:05:14、drb1*04:05:15、drb1*04:05:16、drb1*04:05:17、drb1*04:05:18、drb1*04:05:19、drb1*04:05:20、drb1*04:06:01、drb1*04:06:02、drb1*04:06:03、drb1*04:06:04、drb1*04:06:05、drb1*04:06:06、drb1*04:06:07、drb1*04:07:01:01、drb1*04:07:01:02、drb1*04:07:02、drb1*04:07:03、drb1*04:07:04、drb1*04:07:05、drb1*04:07:06、drb1*04:08:01、drb1*04:08:02、drb1*04:08:03、drb1*04:08:04、drb1*04:09、drb1*04:100、drb1*04:101、drb1*04:102、drb1*04:103、drb1*04:104、drb1*04:105:01、drb1*04:105:02、drb1*04:106、drb1*04:107、drb1*04:108、drb1*04:109、drb1*04:10:01、drb1*04:10:02、drb1*04:10:03、drb1*04:110、drb1*04:111、drb1*04:112、drb1*04:113、drb1*04:114、drb1*04:115、drb1*04:116、drb1*04:117、drb1*04:118、drb1*04:119n、drb1*04:11:01、drb1*04:11:02、drb1*04:11:03、drb1*04:11:04、drb1*04:11:05、drb1*04:12、drb1*04:120n、drb1*04:121、drb1*04:122、drb1*04:123、drb1*04:124、drb1*04:125、drb1*04:126、drb1*04:127、drb1*04:128、drb1*04:129、drb1*04:13、drb1*04:130、drb1*04:131:01、drb1*04:131:02、drb1*04:132、drb1*04:133、drb1*04:134、drb1*04:135、drb1*04:136、
drb1*04:137、drb1*04:138、drb1*04:139、drb1*04:14、drb1*04:140、drb1*04:141、drb1*04:142n、drb1*04:143、drb1*04:144、drb1*04:145、drb1*04:146、drb1*04:147、drb1*04:148、drb1*04:149、drb1*04:15、drb1*04:150、drb1*04:151、drb1*04:152、drb1*04:153、drb1*04:154、drb1*04:155、drb1*04:156、drb1*04:157n、drb1*04:158n、drb1*04:159、drb1*04:16、drb1*04:160、drb1*04:161、drb1*04:162、drb1*04:163、drb1*04:164、drb1*04:165、drb1*04:166、drb1*04:167、drb1*04:168、drb1*04:169、drb1*04:170、drb1*04:171、drb1*04:172、drb1*04:173、drb1*04:174、drb1*04:175、drb1*04:176、drb1*04:177、drb1*04:178n、drb1*04:179、drb1*04:17:01、drb1*04:17:02、drb1*04:18、drb1*04:180、drb1*04:181、drb1*04:182、drb1*04:183、drb1*04:184、drb1*04:185、drb1*04:186n、drb1*04:187、drb1*04:188、drb1*04:189、drb1*04:19、drb1*04:190、drb1*04:191、drb1*04:192、drb1*04:193、drb1*04:194、drb1*04:195、drb1*04:196、drb1*04:197、drb1*04:198、drb1*04:199、drb1*04:20、drb1*04:200、drb1*04:201、drb1*04:202、drb1*04:203、drb1*04:204、drb1*04:205、drb1*04:206、drb1*04:207、drb1*04:208、drb1*04:209、drb1*04:21、drb1*04:210、drb1*04:211、drb1*04:212n、drb1*04:213、drb1*04:214n、drb1*04:215、drb1*04:216、drb1*04:217、drb1*04:218、drb1*04:219、drb1*04:22、drb1*04:220、drb1*04:221、drb1*04:222、drb1*04:223、drb1*04:224、drb1*04:225、drb1*04:226:01、drb1*04:226:02、drb1*04:227、drb1*04:228、drb1*04:229、drb1*04:23、drb1*04:230、drb1*04:231、drb1*04:232、drb1*04:233、drb1*04:234、drb1*04:235、drb1*04:236、drb1*04:237、drb1*04:238、drb1*04:239、drb1*04:24、drb1*04:240、drb1*04:241、drb1*04:242、drb1*04:243、drb1*04:244、drb1*04:245、drb1*04:246、drb1*04:247n、drb1*04:248、drb1*04:249、drb1*04:25、drb1*04:250、drb1*04:251、drb1*04:252、drb1*04:253、drb1*04:254、drb1*04:255、drb1*04:256、drb1*04:257、drb1*04:258、drb1*04:259、drb1*04:26、drb1*04:260、drb1*04:261、drb1*04:262、drb1*04:263、drb1*04:264n、drb1*04:265、drb1*04:266n、drb1*04:267n、drb1*04:268、drb1*04:269、drb1*04:27、drb1*04:270、drb1*04:271、drb1*04:272、drb1*04:28、drb1*04:29、drb1*04:30、drb1*04:31、drb1*04:32、drb1*04:33、drb1*04:34、drb1*04:35、drb1*04:36、drb1*04:37、drb1*04:38、drb1*04:39、drb1*04:40、drb1*04:41、drb1*04:42、drb1*04:43、drb1*04:44:01、drb1*04:44:02、drb1*04:45、drb1*04:46、drb1*04:47、drb1*04:48、drb1*04:49、drb1*04:50、drb1*04:51、drb1*04:52、drb1*04:53:01、drb1*04:53:02、drb1*04:54、drb1*04:55、drb1*04:56:01、drb1*04:56:02、drb1*04:57、drb1*04:58、drb1*04:59、drb1*04:60、drb1*04:61、drb1*04:62、drb1*04:63、drb1*04:64、drb1*04:65、drb1*04:66、drb1*04:67、drb1*04:68、drb1*04:69、drb1*04:70、drb1*04:71、drb1*04:72:01、drb1*04:72:02、drb1*04:73:01、drb1*04:73:02、drb1*04:74、drb1*04:75、drb1*04:76、drb1*04:77、drb1*04:78、drb1*04:79、drb1*04:80、drb1*04:81n、drb1*04:82、drb1*04:83、drb1*04:84、drb1*04:85、drb1*04:86、drb1*04:87、drb1*04:88、drb1*04:89、drb1*04:90、drb1*04:91、drb1*04:92、drb1*04:93、drb1*04:94:01n、drb1*04:95:01、drb1*04:95:02、drb1*04:96、drb1*04:97、drb1*04:98:01、drb1*04:98:02、drb1*04:99、drb1*07:01:01:01、drb1*07:01:01:02、drb1*07:01:01:03、drb1*07:01:01:04、drb1*07:01:02、drb1*07:01:03、drb1*07:01:04、drb1*07:01:05、drb1*07:01:06、drb1*07:01:07、drb1*07:01:08、drb1*07:01:09、drb1*07:01:10、drb1*
07:01:11、drb1*07:01:12、drb1*07:01:13、drb1*07:01:14、drb1*07:01:15、drb1*07:01:16、drb1*07:01:17、drb1*07:01:18、drb1*07:01:19、drb1*07:01:20、drb1*07:01:21、drb1*07:01:22、drb1*07:03、drb1*07:04、drb1*07:05、drb1*07:06、drb1*07:07、drb1*07:08、drb1*07:09、drb1*07:100、drb1*07:101n、drb1*07:10n、drb1*07:11、drb1*07:12、drb1*07:13、drb1*07:14、drb1*07:15、drb1*07:16、drb1*07:17、drb1*07:18、drb1*07:19、drb1*07:20、drb1*07:21、drb1*07:22、drb1*07:23、drb1*07:24、drb1*07:25、drb1*07:26n、drb1*07:27、drb1*07:28、drb1*07:29、drb1*07:30、drb1*07:31、drb1*07:32、drb1*07:33、drb1*07:34、drb1*07:35、drb1*07:36、drb1*07:37、drb1*07:38、drb1*07:39、drb1*07:40、drb1*07:41、drb1*07:42、drb1*07:43、drb1*07:44、drb1*07:45、drb1*07:46、drb1*07:47、drb1*07:48、drb1*07:49、drb1*07:50、drb1*07:51、drb1*07:52、drb1*07:53、drb1*07:54、drb1*07:55、drb1*07:56、drb1*07:57、drb1*07:58n、drb1*07:59、drb1*07:60、drb1*07:61、drb1*07:62、drb1*07:63、drb1*07:64、drb1*07:65、drb1*07:66、drb1*07:67、drb1*07:68n、drb1*07:69、drb1*07:70、drb1*07:71、drb1*07:72、drb1*07:73、drb1*07:74、drb1*07:75、drb1*07:76、drb1*07:77、drb1*07:78、drb1*07:79、drb1*07:80、drb1*07:81、drb1*07:82、drb1*07:83、drb1*07:84、drb1*07:85、drb1*07:86、drb1*07:87n、drb1*07:88、drb1*07:89、drb1*07:90、drb1*07:91、drb1*07:92、drb1*07:93、drb1*07:94、drb1*07:95、drb1*07:96、drb1*07:97、drb1*07:98、drb1*07:99、drb1*08:01:01、drb1*08:01:02、drb1*08:01:04、drb1*08:01:05、drb1*08:01:06、drb1*08:01:07、drb1*08:02:01:01、drb1*08:02:01:02、drb1*08:02:02、drb1*08:02:03、drb1*08:02:04、drb1*08:03:02:01、drb1*08:03:02:02、drb1*08:03:03、drb1*08:03:04、drb1*08:03:05、drb1*08:03:06、drb1*08:03:07、drb1*08:03:08、drb1*08:03:09、drb1*08:04:01、drb1*08:04:02、drb1*08:04:03、drb1*08:04:04、drb1*08:04:05、drb1*08:04:06、drb1*08:04:07、drb1*08:05、drb1*08:06、drb1*08:07、drb1*08:08、drb1*08:09、drb1*08:10、drb1*08:11、drb1*08:12、drb1*08:13、drb1*08:14、drb1*08:15、drb1*08:16、drb1*08:17、drb1*08:18、drb1*08:19、drb1*08:20、drb1*08:21、drb1*08:22、drb1*08:23、drb1*08:24:01、drb1*08:24:02、drb1*08:25、drb1*08:26、drb1*08:27、drb1*08:28、drb1*08:29、drb1*08:30:01、drb1*08:30:02、drb1*08:30:03、drb1*08:31、drb1*08:32、drb1*08:33、drb1*08:34、drb1*08:35、drb1*08:36:01、drb1*08:36:02、drb1*08:37、drb1*08:38、drb1*08:39、drb1*08:40、drb1*08:41、drb1*08:42、drb1*08:43、drb1*08:44、drb1*08:45:01、drb1*08:45:02、drb1*08:46、drb1*08:47、drb1*08:48、drb1*08:49、drb1*08:50、drb1*08:51、drb1*08:52、drb1*08:53、drb1*08:54、drb1*08:55、drb1*08:56、drb1*08:57、drb1*08:58、drb1*08:59、drb1*08:60n、drb1*08:61、drb1*08:62、drb1*08:63、drb1*08:64、drb1*08:65、drb1*08:66、drb1*08:67、drb1*08:68:01、drb1*08:68:02、drb1*08:69、drb1*08:70、drb1*08:71、drb1*08:72、drb1*08:73、drb1*08:74、drb1*08:75、drb1*08:76、drb1*08:77、drb1*08:78n、drb1*08:79、drb1*08:80、drb1*08:81、drb1*08:82、drb1*08:83、drb1*08:84、drb1*08:85、drb1*08:86、drb1*08:87、drb1*08:88、drb1*08:89n、drb1*08:90、drb1*09:01:02:01、drb1*09:01:02:02、drb1*09:01:03、drb1*09:01:04、drb1*09:01:05、drb1*09:01:06、drb1*09:01:07、drb1*09:01:08、drb1*09:01:09、drb1*09:01:10、drb1*09:01:11、drb1*09:02:01、drb1*09:02:02、drb1*09:03、drb1*09:04、drb1*09:05、drb1*09:06、drb1*09:07、drb1*09:
08、drb1*09:09、drb1*09:10、drb1*09:11、drb1*09:12、drb1*09:13、drb1*09:14、drb1*09:15、drb1*09:16、drb1*09:17、drb1*09:18、drb1*09:19、drb1*09:20、drb1*09:21、drb1*09:22、drb1*09:23、drb1*09:24、drb1*09:25、drb1*09:26、drb1*09:27、drb1*09:28、drb1*09:29、drb1*09:30、drb1*09:31、drb1*09:32、drb1*09:33、drb1*09:34、drb1*09:35、drb1*09:36、drb1*09:37n、drb1*09:38、drb1*09:39、drb1*09:40、drb1*10:01:01:01、drb1*10:01:01:02、drb1*10:01:01:03、drb1*10:01:02、drb1*10:01:03、drb1*10:01:04、drb1*10:01:05、drb1*10:01:06、drb1*10:01:07、drb1*10:01:08、drb1*10:01:09、drb1*10:01:10、drb1*10:01:11、drb1*10:01:12、drb1*10:02、drb1*10:03、drb1*10:04、drb1*10:05、drb1*10:06、drb1*10:07、drb1*10:08、drb1*10:09、drb1*10:10、drb1*10:11、drb1*10:12、drb1*10:13、drb1*10:14、drb1*10:15、drb1*10:16、drb1*10:17、drb1*10:18、drb1*10:19、drb1*10:20、drb1*10:21、drb1*10:22、drb1*10:23、drb1*10:24、drb1*10:25、drb1*10:26、drb1*10:27、drb1*10:28、drb1*10:29、drb1*10:30、drb1*10:31、drb1*10:32、drb1*10:33、drb1*11:01:01:01、drb1*11:01:01:02、drb1*11:01:01:03、drb1*11:01:01:04、drb1*11:01:02、drb1*11:01:03、drb1*11:01:04、drb1*11:01:05、drb1*11:01:06、drb1*11:01:07、drb1*11:01:08、drb1*11:01:09、drb1*11:01:10、drb1*11:01:11、drb1*11:01:12、drb1*11:01:13、drb1*11:01:14、drb1*11:01:15、drb1*11:01:16、drb1*11:01:17、drb1*11:01:18、drb1*11:01:19、drb1*11:01:20、drb1*11:01:21、drb1*11:01:22、drb1*11:01:23、drb1*11:01:24、drb1*11:01:25、drb1*11:01:26、drb1*11:01:27、drb1*11:01:28、drb1*11:01:29、drb1*11:01:30、drb1*11:01:31、drb1*11:01:32、drb1*11:01:33、drb1*11:02:01、drb1*11:02:02、drb1*11:02:03、drb1*11:02:04、drb1*11:02:05、drb1*11:03:01、drb1*11:03:02、drb1*11:03:03、drb1*11:03:04、drb1*11:04:01、drb1*11:04:02、drb1*11:04:03、drb1*11:04:04、drb1*11:04:05、drb1*11:04:06、drb1*11:04:07、drb1*11:04:08、drb1*11:04:09、drb1*11:04:10、drb1*11:04:11、drb1*11:04:12、drb1*11:04:13、drb1*11:04:14、drb1*11:04:15、drb1*11:04:16、drb1*11:04:17、drb1*11:04:18、drb1*11:05、drb1*11:06:01、drb1*11:06:02、drb1*11:06:03、drb1*11:07:01、drb1*11:07:02、drb1*11:08:01、drb1*11:08:02、drb1*11:08:03、drb1*11:09、drb1*11:100、drb1*11:101:01、drb1*11:101:02、drb1*11:102:01、drb1*11:102:02、drb1*11:103:01、drb1*11:103:02、drb1*11:104、drb1*11:105、drb1*11:106、drb1*11:107、drb1*11:108、drb1*11:109、drb1*11:10:01、drb1*11:10:02、drb1*11:110、drb1*11:111、drb1*11:112、drb1*11:113、drb1*11:114、drb1*11:115、drb1*11:116、drb1*11:117:01、drb1*11:117:02、drb1*11:118、drb1*11:119、drb1*11:11:01、drb1*11:11:03、drb1*11:120、drb1*11:121、drb1*11:122、drb1*11:123、drb1*11:124:01、drb1*11:124:02、drb1*11:125、drb1*11:126、drb1*11:127、drb1*11:128、drb1*11:129、drb1*11:12:01、drb1*11:12:02、drb1*11:12:03、drb1*11:130、drb1*11:131、drb1*11:132、drb1*11:133、drb1*11:134、drb1*11:135、drb1*11:136、drb1*11:137、drb1*11:138、drb1*11:139、drb1*11:13:01、drb1*11:13:02、drb1*11:140、drb1*11:141、drb1*11:142、drb1*11:143、drb1*11:144、drb1*11:145、drb1*11:146、drb1*11:147:01、drb1*11:147:02、drb1*11:148、drb1*11:149、drb1*11:14:01、drb1*11:14:02、drb1*11:15、drb1*11:150、drb1*11:151、drb1*11:152、drb1*11:153、drb1*11:154、drb1*11:155、drb1*11:156、drb1*11:157、drb1*11:158、drb1*11:159、drb1*11:16、drb1*
11:160、drb1*11:161、drb1*11:162、drb1*11:163、drb1*11:164、drb1*11:165:01、drb1*11:165:02、drb1*11:166、drb1*11:167、drb1*11:168、drb1*11:169n、drb1*11:17、drb1*11:170、drb1*11:171、drb1*11:172、drb1*11:173、drb1*11:174、drb1*11:175、drb1*11:176、drb1*11:177、drb1*11:178、drb1*11:179、drb1*11:18、drb1*11:180、drb1*11:181、drb1*11:182、drb1*11:183、drb1*11:184、drb1*11:185、drb1*11:186、drb1*11:187、drb1*11:188、drb1*11:189、drb1*11:190、drb1*11:191、drb1*11:192、drb1*11:193:01、drb1*11:193:02、drb1*11:194、drb1*11:195、drb1*11:196、drb1*11:197、drb1*11:198、drb1*11:199、drb1*11:19:01、drb1*11:19:02、drb1*11:19:03、drb1*11:20、drb1*11:200、drb1*11:201、drb1*11:202、drb1*11:203、drb1*11:204、drb1*11:205、drb1*11:206、drb1*11:207、drb1*11:208、drb1*11:209、drb1*11:21、drb1*11:210、drb1*11:211、drb1*11:212、drb1*11:213、drb1*11:214、drb1*11:215、drb1*11:216、drb1*11:217n、drb1*11:218、drb1*11:219、drb1*11:22、drb1*11:220、drb1*11:221、drb1*11:222、drb1*11:223、drb1*11:224、drb1*11:225、drb1*11:226、drb1*11:227、drb1*11:228、drb1*11:229、drb1*11:230、drb1*11:231、drb1*11:232、drb1*11:233、drb1*11:234、drb1*11:235、drb1*11:236、drb1*11:237、drb1*11:238、drb1*11:239、drb1*11:23:01、drb1*11:23:02、drb1*11:240、drb1*11:241、drb1*11:242、drb1*11:243、drb1*11:244、drb1*11:245、drb1*11:246n、drb1*11:247、drb1*11:248q、drb1*11:249、drb1*11:24:01、drb1*11:24:02、drb1*11:25、drb1*11:250n、drb1*11:251、drb1*11:252、drb1*11:253、drb1*11:254、drb1*11:26、drb1*11:27:01、drb1*11:27:02、drb1*11:27:03、drb1*11:28:01、drb1*11:28:02、drb1*11:29:01、drb1*11:29:02、drb1*11:30、drb1*11:31、drb1*11:32、drb1*11:33、drb1*11:34、drb1*11:35、drb1*11:36、drb1*11:37:01、drb1*11:37:02、drb1*11:38、drb1*11:39、drb1*11:40、drb1*11:41、drb1*11:42:01、drb1*11:42:02、drb1*11:43、drb1*11:44、drb1*11:45、drb1*11:46:01、drb1*11:46:02、drb1*11:47、drb1*11:48、drb1*11:49:01、drb1*11:49:02、drb1*11:50、drb1*11:51、drb1*11:52、drb1*11:53、drb1*11:54:01、drb1*11:54:02、drb1*11:55、drb1*11:56、drb1*11:57、drb1*11:58:01、drb1*11:58:02、drb1*11:59、drb1*11:60、drb1*11:61、drb1*11:62:01、drb1*11:62:02、drb1*11:63:01、drb1*11:63:02、drb1*11:64、drb1*11:65:01、drb1*11:65:02、drb1*11:66:01、drb1*11:66:02、drb1*11:67、drb1*11:68、drb1*11:69、drb1*11:70、drb1*11:72、drb1*11:73、drb1*11:74:01、drb1*11:74:02、drb1*11:75、drb1*11:76、drb1*11:77、drb1*11:78、drb1*11:79、drb1*11:80、drb1*11:81、drb1*11:82、drb1*11:83、drb1*11:84:01、drb1*11:84:02、drb1*11:84:03、drb1*11:85、drb1*11:86、drb1*11:87、drb1*11:88、drb1*11:89、drb1*11:90、drb1*11:91、drb1*11:92、drb1*11:93、drb1*11:94、drb1*11:95、drb1*11:96、drb1*11:97、drb1*11:98、drb1*11:99、drb1*12:01:01:01、drb1*12:01:01:02、drb1*12:01:01:03、drb1*12:01:01:04、drb1*12:01:01:05、drb1*12:01:01:06、drb1*12:01:02、drb1*12:01:03、drb1*12:01:04、drb1*12:01:05、drb1*12:01:06、drb1*12:01:07、drb1*12:01:08、drb1*12:01:09、drb1*12:02:01:01、drb1*12:02:01:02、drb1*12:02:01:03、drb1*12:02:01:04、drb1*12:02:02、drb1*12:02:03、drb1*12:02:04、drb1*12:02:05、drb1*12:02:06、drb1*12:02:07、drb1*12:02:08、drb1*12:02:09、drb1*12:03:02、drb1*12:03:03、drb1*12:04、drb1*12:05、drb1*12:06、drb1*12:07、drb1*12:08、drb1*12:09、drb1*12:10、drb1*12:11、drb1*12:
12、drb1*12:13、drb1*12:14、drb1*12:15、drb1*12:16:01、drb1*12:16:02、drb1*12:16:03、drb1*12:17、drb1*12:18、drb1*12:19、drb1*12:20、drb1*12:21、drb1*12:22、drb1*12:23、drb1*12:24n、drb1*12:25、drb1*12:26、drb1*12:27、drb1*12:28、drb1*12:29、drb1*12:30、drb1*12:31n、drb1*12:32、drb1*12:33、drb1*12:34、drb1*12:35、drb1*12:36、drb1*12:37、drb1*12:38、drb1*12:39、drb1*12:40、drb1*12:41、drb1*12:42、drb1*12:43、drb1*12:44、drb1*12:45、drb1*12:46、drb1*12:47、drb1*12:48、drb1*12:49、drb1*12:50、drb1*12:51、drb1*12:52、drb1*12:53、drb1*12:54、drb1*12:55、drb1*12:56、drb1*12:57、drb1*12:58、drb1*12:59、drb1*12:60n、drb1*12:61、drb1*12:62、drb1*12:63、drb1*12:64、drb1*12:65、drb1*12:66、drb1*12:67、drb1*12:68、drb1*12:69、drb1*12:70、drb1*12:71、drb1*12:72n、drb1*12:73、drb1*12:74n、drb1*12:75、drb1*13:01:01:01、drb1*13:01:01:02、drb1*13:01:02、drb1*13:01:03、drb1*13:01:04、drb1*13:01:05、drb1*13:01:06、drb1*13:01:07、drb1*13:01:08、drb1*13:01:09、drb1*13:01:10、drb1*13:01:11、drb1*13:01:12、drb1*13:01:13、drb1*13:01:14、drb1*13:01:15、drb1*13:01:16、drb1*13:01:17、drb1*13:01:18、drb1*13:01:19、drb1*13:01:20、drb1*13:01:21、drb1*13:01:22、drb1*13:01:23、drb1*13:01:24、drb1*13:01:25、drb1*13:01:26、drb1*13:02:01:01、drb1*13:02:01:02、drb1*13:02:01:03、drb1*13:02:02、drb1*13:02:03、drb1*13:02:04、drb1*13:02:05、drb1*13:02:06、drb1*13:02:07、drb1*13:02:08、drb1*13:02:09、drb1*13:02:10、drb1*13:02:11、drb1*13:02:12、drb1*13:02:13、drb1*13:02:14、drb1*13:02:15、drb1*13:02:16、drb1*13:02:17、drb1*13:03:01、drb1*13:03:02、drb1*13:03:03、drb1*13:03:04、drb1*13:03:05、drb1*13:03:06、drb1*13:03:07、drb1*13:03:08、drb1*13:03:09、drb1*13:04、drb1*13:05:01、drb1*13:05:02、drb1*13:05:03、drb1*13:06、drb1*13:07:01、drb1*13:07:02、drb1*13:08、drb1*13:09、drb1*13:10、drb1*13:100、drb1*13:101、drb1*13:102、drb1*13:103、drb1*13:104、drb1*13:105、drb1*13:106、drb1*13:107、drb1*13:108、drb1*13:109、drb1*13:110、drb1*13:111、drb1*13:112、drb1*13:113n、drb1*13:114、drb1*13:115、drb1*13:116、drb1*13:117、drb1*13:118、drb1*13:119、drb1*13:11:01、drb1*13:11:02、drb1*13:120、drb1*13:121、drb1*13:122、drb1*13:123、drb1*13:124、drb1*13:125、drb1*13:126、drb1*13:127、drb1*13:128、drb1*13:129、drb1*13:12:01、drb1*13:12:02、drb1*13:12:03、drb1*13:12:04、drb1*13:13、drb1*13:130、drb1*13:131、drb1*13:132、drb1*13:133、drb1*13:134、drb1*13:135、drb1*13:136、drb1*13:137n、drb1*13:138、drb1*13:139、drb1*13:140、drb1*13:141、drb1*13:142n、drb1*13:143、drb1*13:144、drb1*13:145、drb1*13:146、drb1*13:147、drb1*13:148、drb1*13:149、drb1*13:14:01、drb1*13:14:02、drb1*13:14:03、drb1*13:15、drb1*13:150、drb1*13:151、drb1*13:152、drb1*13:153、drb1*13:154、drb1*13:155、drb1*13:156、drb1*13:157、drb1*13:158、drb1*13:159、drb1*13:16、drb1*13:160、drb1*13:161、drb1*13:162、drb1*13:163、drb1*13:164、drb1*13:165、drb1*13:166、drb1*13:167、drb1*13:168、drb1*13:169、drb1*13:17、drb1*13:170、drb1*13:171:01、drb1*13:171:02、drb1*13:172、drb1*13:173、drb1*13:174、drb1*13:175、drb1*13:176、drb1*13:177、drb1*13:178、drb1*13:179、drb1*13:18、drb1*13:180、drb1*13:181、drb1*13:182、drb1*13:183、drb1*13:184、drb1*13:185n、drb1*13:186、drb1*13:187、drb1*13:188、drb1*13:189、drb1*13:19、drb1*13:190、
drb1*13:191、drb1*13:192、drb1*13:193、drb1*13:194、drb1*13:195、drb1*13:196、drb1*13:197、drb1*13:198、drb1*13:199、drb1*13:20、drb1*13:200n、drb1*13:201、drb1*13:202、drb1*13:203、drb1*13:204、drb1*13:205、drb1*13:206、drb1*13:207、drb1*13:208、drb1*13:209、drb1*13:210、drb1*13:211、drb1*13:212、drb1*13:213、drb1*13:214、drb1*13:215、drb1*13:216、drb1*13:217、drb1*13:218、drb1*13:219、drb1*13:21:01、drb1*13:21:02、drb1*13:220、drb1*13:221、drb1*13:222、drb1*13:223、drb1*13:224、drb1*13:225、drb1*13:226、drb1*13:227、drb1*13:228、drb1*13:229、drb1*13:22:01、drb1*13:22:02、drb1*13:230、drb1*13:231、drb1*13:232、drb1*13:233、drb1*13:234、drb1*13:235、drb1*13:236、drb1*13:237、drb1*13:238、drb1*13:239、drb1*13:23:01、drb1*13:23:02、drb1*13:24、drb1*13:240、drb1*13:241、drb1*13:242:01、drb1*13:242:02、drb1*13:243、drb1*13:244、drb1*13:245、drb1*13:246、drb1*13:247、drb1*13:248、drb1*13:249n、drb1*13:25、drb1*13:250、drb1*13:251、drb1*13:252n、drb1*13:253、drb1*13:254、drb1*13:255n、drb1*13:256、drb1*13:257、drb1*13:258、drb1*13:259、drb1*13:260、drb1*13:261、drb1*13:262、drb1*13:263、drb1*13:264、drb1*13:265、drb1*13:266、drb1*13:267、drb1*13:268n、drb1*13:269、drb1*13:26:01、drb1*13:26:02、drb1*13:27、drb1*13:270、drb1*13:271、drb1*13:272、drb1*13:273、drb1*13:274、drb1*13:275、drb1*13:276、drb1*13:277、drb1*13:278q、drb1*13:279、drb1*13:28:01、drb1*13:28:02、drb1*13:29、drb1*13:30、drb1*13:31、drb1*13:32、drb1*13:33:01、drb1*13:33:02、drb1*13:33:03、drb1*13:34、drb1*13:35、drb1*13:36、drb1*13:37、drb1*13:38、drb1*13:39、drb1*13:40、drb1*13:41、drb1*13:42、drb1*13:43、drb1*13:44、drb1*13:45、drb1*13:46、drb1*13:47、drb1*13:48、drb1*13:49、drb1*13:50:01、drb1*13:50:02、drb1*13:50:03、drb1*13:51、drb1*13:52、drb1*13:53、drb1*13:54、drb1*13:55、drb1*13:56、drb1*13:57、drb1*13:58、drb1*13:59、drb1*13:60、drb1*13:61:01、drb1*13:61:02、drb1*13:62、drb1*13:63、drb1*13:64、drb1*13:65、drb1*13:66:01、drb1*13:66:02、drb1*13:67、drb1*13:68、drb1*13:69、drb1*13:70、drb1*13:71、drb1*13:72、drb1*13:73、drb1*13:74、drb1*13:75、drb1*13:76、drb1*13:77、drb1*13:78、drb1*13:79、drb1*13:80、drb1*13:81、drb1*13:82、drb1*13:83、drb1*13:84、drb1*13:85、drb1*13:86、drb1*13:87、drb1*13:88、drb1*13:89:01、drb1*13:89:02、drb1*13:90、drb1*13:91、drb1*13:92、drb1*13:93、drb1*13:94:01、drb1*13:94:02、drb1*13:95、drb1*13:96:01、drb1*13:96:02、drb1*13:97:01、drb1*13:97:02、drb1*13:98、drb1*13:99、drb1*14:01:01、drb1*14:01:02、drb1*14:01:03、drb1*14:01:04、drb1*14:02:01:01、drb1*14:02:01:02、drb1*14:02:02、drb1*14:02:03、drb1*14:02:04、drb1*14:02:05、drb1*14:02:06、drb1*14:02:07、drb1*14:03:01、drb1*14:03:02、drb1*14:04:01、drb1*14:04:02、drb1*14:04:03、drb1*14:04:04、drb1*14:04:05、drb1*14:04:06、drb1*14:05:01:01、drb1*14:05:01:02、drb1*14:05:02、drb1*14:05:03、drb1*14:05:04、drb1*14:06:01、drb1*14:06:02、drb1*14:06:03、drb1*14:06:04、drb1*14:07:01、drb1*14:07:02、drb1*14:08、drb1*14:09、drb1*14:10、drb1*14:100、drb1*14:101、drb1*14:102、drb1*14:103、drb1*14:104、drb1*14:105、drb1*14:106、drb1*14:107、drb1*14:108、drb1*14:109、drb1*14:11、drb1*14:110、drb1*14:111、drb1*14:112、drb1*14:113、drb1*14:114、drb1*14:115、drb1*14:116、drb1*14:117、drb1*14:118、drb1*14:119、drb1*
14:120、drb1*14:121、drb1*14:122、drb1*14:123、drb1*14:124、drb1*14:125、drb1*14:126:01、drb1*14:126:02、drb1*14:127:01、drb1*14:127:02、drb1*14:128、drb1*14:129、drb1*14:12:01、drb1*14:12:02、drb1*14:13、drb1*14:130、drb1*14:131、drb1*14:132、drb1*14:133、drb1*14:134、drb1*14:135、drb1*14:136、drb1*14:137n、drb1*14:138、drb1*14:139、drb1*14:14、drb1*14:140、drb1*14:141、drb1*14:142、drb1*14:143、drb1*14:144、drb1*14:145、drb1*14:146、drb1*14:147、drb1*14:148、drb1*14:149、drb1*14:15、drb1*14:150、drb1*14:151、drb1*14:152n、drb1*14:153、drb1*14:154、drb1*14:155、drb1*14:156、drb1*14:157、drb1*14:158、drb1*14:159、drb1*14:16、drb1*14:160、drb1*14:161、drb1*14:162、drb1*14:163、drb1*14:164、drb1*14:165、drb1*14:166n、drb1*14:167、drb1*14:168、drb1*14:169、drb1*14:17、drb1*14:170、drb1*14:171、drb1*14:172、drb1*14:173、drb1*14:174、drb1*14:175、drb1*14:176、drb1*14:177、drb1*14:178、drb1*14:179、drb1*14:18、drb1*14:180、drb1*14:181、drb1*14:182、drb1*14:183、drb1*14:184、drb1*14:185、drb1*14:186、drb1*14:187、drb1*14:188n、drb1*14:189、drb1*14:19、drb1*14:190、drb1*14:191、drb1*14:192、drb1*14:193、drb1*14:194、drb1*14:195n、drb1*14:196、drb1*14:197n、drb1*14:198、drb1*14:199、drb1*14:20、drb1*14:200、drb1*14:201、drb1*14:202、drb1*14:203、drb1*14:204、drb1*14:205、drb1*14:206、drb1*14:207、drb1*14:208、drb1*14:209、drb1*14:21、drb1*14:210q、drb1*14:211、drb1*14:22、drb1*14:23:01、drb1*14:23:02、drb1*14:23:03、drb1*14:23:04、drb1*14:24、drb1*14:25:01、drb1*14:25:02、drb1*14:26、drb1*14:27:01、drb1*14:27:02、drb1*14:28、drb1*14:29、drb1*14:30、drb1*14:31、drb1*14:32:01、drb1*14:32:02、drb1*14:32:03、drb1*14:33、drb1*14:34、drb1*14:35、drb1*14:36、drb1*14:37、drb1*14:38:01、drb1*14:38:02、drb1*14:39、drb1*14:40、drb1*14:41、drb1*14:42、drb1*14:43、drb1*14:44:01、drb1*14:44:02、drb1*14:44:03、drb1*14:45、drb1*14:46、drb1*14:47、drb1*14:48、drb1*14:49、drb1*14:50、drb1*14:51、drb1*14:52、drb1*14:53、drb1*14:54:01:01、drb1*14:54:01:02、drb1*14:54:01:03、drb1*14:54:01:04、drb1*14:54:02、drb1*14:54:03、drb1*14:54:04、drb1*14:54:05、drb1*14:54:06、drb1*14:54:07、drb1*14:55、drb1*14:56、drb1*14:57、drb1*14:58、drb1*14:59、drb1*14:60、drb1*14:61、drb1*14:62、drb1*14:63、drb1*14:64、drb1*14:65、drb1*14:67、drb1*14:68:01、drb1*14:68:02、drb1*14:69、drb1*14:70、drb1*14:71、drb1*14:72、drb1*14:73、drb1*14:74、drb1*14:75、drb1*14:76、drb1*14:77、drb1*14:78、drb1*14:79、drb1*14:80、drb1*14:81、drb1*14:82、drb1*14:83、drb1*14:84、drb1*14:85、drb1*14:86、drb1*14:87、drb1*14:88、drb1*14:89、drb1*14:90、drb1*14:91、drb1*14:92n、drb1*14:93、drb1*14:94、drb1*14:95、drb1*14:96、drb1*14:97、drb1*14:98、drb1*14:99、drb1*15:01:01:01、drb1*15:01:01:02、drb1*15:01:01:03、drb1*15:01:01:04、drb1*15:01:01:05、drb1*15:01:02、drb1*15:01:03、drb1*15:01:04、drb1*15:01:05、drb1*15:01:06、drb1*15:01:07、drb1*15:01:08、drb1*15:01:09、drb1*15:01:10、drb1*15:01:11、drb1*15:01:12、drb1*15:01:13、drb1*15:01:14、drb1*15:01:15、drb1*15:01:16、drb1*15:01:17、drb1*15:01:18、drb1*15:01:19、drb1*15:01:20、drb1*15:01:21、drb1*15:01:22、drb1*15:01:23、drb1*15:01:24、drb1*15:01:25、drb1*15:01:26、drb1*15:01:27、drb1*15:01:28、drb1*15:01:29、drb1*15:01:30、drb1*15:01:31、drb1*
15:01:32、drb1*15:01:33、drb1*15:01:34、drb1*15:01:35、drb1*15:01:36、drb1*15:01:37、drb1*15:01:38、drb1*15:01:39、drb1*15:01:40、drb1*15:01:41、drb1*15:02:01:01、drb1*15:02:01:02、drb1*15:02:01:03、drb1*15:02:02、drb1*15:02:03、drb1*15:02:04、drb1*15:02:05、drb1*15:02:06、drb1*15:02:07、drb1*15:02:08、drb1*15:02:09、drb1*15:02:10、drb1*15:02:11、drb1*15:02:12、drb1*15:02:13、drb1*15:02:14、drb1*15:02:15、drb1*15:02:16、drb1*15:02:17、drb1*15:02:18、drb1*15:02:19、drb1*15:03:01:01、drb1*15:03:01:02、drb1*15:03:01:03、drb1*15:03:02、drb1*15:03:03、drb1*15:03:04、drb1*15:04、drb1*15:05、drb1*15:06:01、drb1*15:06:02、drb1*15:06:03、drb1*15:06:04、drb1*15:07:01、drb1*15:07:02、drb1*15:07:03、drb1*15:08、drb1*15:09、drb1*15:10、drb1*15:100、drb1*15:101、drb1*15:102、drb1*15:103、drb1*15:104:01、drb1*15:104:02、drb1*15:104:03、drb1*15:105:01、drb1*15:105:02、drb1*15:106、drb1*15:107、drb1*15:108、drb1*15:109、drb1*15:110、drb1*15:111、drb1*15:112、drb1*15:113n、drb1*15:114、drb1*15:115n、drb1*15:116、drb1*15:117、drb1*15:118、drb1*15:119、drb1*15:11:01、drb1*15:11:02、drb1*15:12、drb1*15:120、drb1*15:121、drb1*15:122、drb1*15:123、drb1*15:124、drb1*15:125、drb1*15:126、drb1*15:127、drb1*15:128、drb1*15:129n、drb1*15:13、drb1*15:130、drb1*15:131、drb1*15:132、drb1*15:133、drb1*15:134n、drb1*15:135、drb1*15:136、drb1*15:137n、drb1*15:138n、drb1*15:139、drb1*15:14、drb1*15:140、drb1*15:141、drb1*15:142、drb1*15:143、drb1*15:144、drb1*15:145、drb1*15:146、drb1*15:147、drb1*15:148n、drb1*15:149、drb1*15:150、drb1*15:151、drb1*15:152、drb1*15:153、drb1*15:154n、drb1*15:155、drb1*15:156、drb1*15:157、drb1*15:158、drb1*15:159n、drb1*15:15:01、drb1*15:15:02、drb1*15:15:03、drb1*15:16、drb1*15:160、drb1*15:161、drb1*15:162、drb1*15:163n、drb1*15:164q、drb1*15:165、drb1*15:166、drb1*15:167、drb1*15:168、drb1*15:169、drb1*15:170、drb1*15:17n、drb1*15:18、drb1*15:19、drb1*15:20、drb1*15:21、drb1*15:22、drb1*15:23、drb1*15:24、drb1*15:25、drb1*15:26、drb1*15:27、drb1*15:28、drb1*15:29、drb1*15:30、drb1*15:31:01、drb1*15:31:02、drb1*15:32、drb1*15:33、drb1*15:34、drb1*15:35、drb1*15:36、drb1*15:37:01、drb1*15:37:02、drb1*15:38、drb1*15:39、drb1*15:40、drb1*15:41、drb1*15:42、drb1*15:43、drb1*15:44、drb1*15:45、drb1*15:46、drb1*15:47、drb1*15:48、drb1*15:49、drb1*15:50n、drb1*15:51、drb1*15:52、drb1*15:53、drb1*15:54、drb1*15:55、drb1*15:56、drb1*15:57、drb1*15:58、drb1*15:59、drb1*15:60、drb1*15:61、drb1*15:62、drb1*15:63、drb1*15:64、drb1*15:65、drb1*15:66:01、drb1*15:66:02、drb1*15:67、drb1*15:68、drb1*15:69、drb1*15:70、drb1*15:71、drb1*15:72、drb1*15:73、drb1*15:74、drb1*15:75、drb1*15:76、drb1*15:77、drb1*15:78、drb1*15:79、drb1*15:80n、drb1*15:81、drb1*15:82、drb1*15:83、drb1*15:84、drb1*15:85、drb1*15:86、drb1*15:87、drb1*15:88、drb1*15:89、drb1*15:90、drb1*15:91、drb1*15:92、drb1*15:93、drb1*15:94、drb1*15:95、drb1*15:96、drb1*15:97、drb1*15:98、drb1*15:99、drb1*16:01:01、drb1*16:01:02、drb1*16:01:03、drb1*16:01:04、drb1*16:01:05、drb1*16:01:06、drb1*16:01:07、drb1*16:01:08、drb1*16:01:09、drb1*16:01:10、drb1*16:01:11、drb1*16:01:12、drb1*16:01:13、drb1*16:01:14、drb1*16:01:15、drb1*16:01:16、drb1*16:02:01:01、drb1*16:02:01:02、drb1*
16:02:01:03、drb1*16:02:02、drb1*16:02:03、drb1*16:02:04、drb1*16:02:05、drb1*16:02:06、drb1*16:02:07、drb1*16:02:08、drb1*16:03、drb1*16:04:01、drb1*16:04:02、drb1*16:05:01、drb1*16:05:02、drb1*16:07、drb1*16:08、drb1*16:09:01、drb1*16:09:02、drb1*16:10:01、drb1*16:10:02、drb1*16:11、drb1*16:12、drb1*16:13n、drb1*16:14、drb1*16:15、drb1*16:16、drb1*16:17、drb1*16:18、drb1*16:19、drb1*16:20、drb1*16:21n、drb1*16:22、drb1*16:23、drb1*16:24、drb1*16:25、drb1*16:26、drb1*16:27、drb1*16:28、drb1*16:29、drb1*16:30、drb1*16:31、drb1*16:32、drb1*16:33、drb1*16:34、drb1*16:35、drb1*16:36、drb1*16:37、drb1*16:38:01、drb1*16:38:02、drb1*16:39、drb1*16:40、drb1*16:41n、drb1*16:42、drb1*16:43、drb1*16:44、drb1*16:45、drb1*16:46、drb1*16:47、drb1*16:48、drb1*16:49、drb1*16:50、drb1*16:51、drb1*16:52、drb1*16:53、drb1*16:54、drb1*16:55n、drb1*16:56、及其任何组合。
[0279]
在某些方面,mhc ii类分子包含drβ链,所述drβ链包含与seq id no:21具有至少约70%、至少约75%、至少约80%、至少约85%、至少约90%、至少约95%、至少约96%、至少约97%、至少约98%、至少约99%或约100%序列同一性的氨基酸序列,其中所述drβ链包含(i)在对应于seq id no:19的氨基酸残基114的位置处的色氨酸,(ii)在对应于seq id no:19的氨基酸残基143的位置处的甲硫氨酸,(iii)在对应于seq id no:19的氨基酸残基118的位置处的组氨酸;以及(vi)在对应于seq id no:19的氨基酸残基157的位置处的异亮氨酸。在某些方面,mhc ii类分子包含drβ链,所述drβ链包含与seq id no:21具有至少约70%、至少约75%、至少约80%、至少约85%、至少约90%、至少约95%、至少约96%、至少约97%、至少约98%、至少约99%或约100%序列同一性的氨基酸序列,其中所述drβ链包含(i)在对应于seq id no:19的氨基酸残基114的位置处的色氨酸,(ii)在对应于seq id no:19的氨基酸残基143的位置处的甲硫氨酸,(iii)在对应于seq id no:19的氨基酸残基118的位置处的组氨酸;(iv)在对应于seq id no:19的氨基酸残基157的位置处的异亮氨酸;(v)在对应于seq id no:19的氨基酸残基139的位置处的苏氨酸;(vi)在对应于seq id no:19的氨基酸残基146的位置处的谷氨酰胺;(vii)在对应于seq id no:19的氨基酸残基163的位置处的甲硫氨酸;和(vii)在对应于seq id no:19的氨基酸残基164的位置处的苏氨酸。在某些方面,mhc ii类分子包含drβ链,所述drβ链包含seq id no:21所示的氨基酸序列。
[0280]
ii.a.3.b.hla-drα链
[0281]
在本公开的一些方面,mhc ii类分子还包含α链。在一些方面,α链是野生型α链。在一些方面,α链是drα链。可以在本公开的组合物和方法中使用任何drα链。在一些方面,drα链包含hla-dra1*01等位基因。
[0282]
在某些方面,drα链选自dra*01:01:01:01、dra*01:01:01:02、dra*01:01:01:03、dra*01:01:02、dra*01:02:01、dra*01:02:02、dra*01:02:03、及其任何组合。
[0283]
在某些方面,mhc ii类分子包含drα链,所述drα链包含与seq id no:24具有至少约70%、至少约75%、至少约80%、至少约85%、至少约90%、至少约95%、至少约96%、至少约97%、至少约98%、至少约99%或约100%序列同一性的氨基酸序列。在某些方面,mhc ii类分子包含drα链,所述drα链包含与seq id no:26具有至少约70%、至少约75%、至少约80%、至少约85%、至少约90%、至少约95%、至少约96%、至少约97%、至少约98%、至少约
99%或约100%序列同一性的氨基酸序列。在某些方面,mhc ii类分子包含drα链,所述drα链包含seq id no:24所示的氨基酸序列。在某些方面,mhc ii类分子包含drα链,所述drα链包含seq id no:26所示的氨基酸序列。
[0284]
ii.a.4.信号肽
[0285]
在一些方面,β链和/或α链进一步包含信号肽。本领域已知的任何信号肽均可用在本文公开的组合物和方法中。在一些方面,β链信号肽与α信号肽相同。在一些方面,β链信号肽不同于α信号肽。
[0286]
在一些方面,信号肽来源于天然信号肽。在一些方面,信号肽来源于天然存在的dpβ链信号肽。在一些方面,信号肽包含天然存在的dpβ链信号肽。在一些方面,信号肽来源于天然存在的dpα链信号肽。在一些方面,信号肽包含天然存在的dpα链信号肽。
[0287]
在一些方面,信号肽来源于天然存在的dqβ链信号肽。在一些方面,信号肽包含天然存在的dqβ链信号肽。在一些方面,信号肽来源于天然存在的dqα链信号肽。在一些方面,信号肽包含天然存在的dqα链信号肽。
[0288]
在一些方面,信号肽来源于天然存在的drβ链信号肽。在一些方面,信号肽包含天然存在的drβ链信号肽。在一些方面,信号肽来源于天然存在的drα链信号肽。在一些方面,信号肽包含天然存在的drα链信号肽。
[0289]
在一些方面,信号肽来源于丝心蛋白轻链(fibl)信号肽。在一些方面,信号肽包含seq id no:9。在一些方面,信号肽是合成的。
[0290]
ii.a.5.跨膜结构域
[0291]
在一些方面,β链和/或α链进一步包含跨膜结构域。跨膜结构域可以是任何长度和任何来源。在一些方面,跨膜结构域的长度为至少约1个到至少约50个氨基酸。在一些方面,跨膜结构域来源于天然存在的跨膜结构域。在一些方面,跨膜结构域包含天然存在的跨膜结构域。在一些方面,跨膜结构域来源于天然存在的hla跨膜结构域。在一些方面,跨膜结构域包含天然存在的hla跨膜结构域。
[0292]
在一些方面,跨膜结构域来源于天然存在的dpβ链跨膜结构域。在一些方面,跨膜结构域包含天然存在的dpβ链跨膜结构域。在一些方面,跨膜结构域来源于天然存在的dpα链跨膜结构域。在一些方面,跨膜结构域包含天然存在的dpα链跨膜结构域。
[0293]
在一些方面,跨膜结构域来源于天然存在的dqβ链跨膜结构域。在一些方面,跨膜结构域包含天然存在的dqβ链跨膜结构域。在一些方面,跨膜结构域来源于天然存在的dqα链跨膜结构域。在一些方面,跨膜结构域包含天然存在的dqα链跨膜结构域。
[0294]
在一些方面,跨膜结构域来源于天然存在的drβ链跨膜结构域。在一些方面,跨膜结构域包含天然存在的drβ链跨膜结构域。在一些方面,跨膜结构域来源于天然存在的drα链跨膜结构域。在一些方面,跨膜结构域包含天然存在的drα链跨膜结构域。
[0295]
ii.a.6.亮氨酸拉链
[0296]
在一些方面,β链和/或α链进一步包含一个或多个亮氨酸拉链(lzip)序列。本领域已知的任何lzip序列均可用在本文公开的组合物和方法中。在一些方面,β链和/或α链包含酸性lzip(αlzip)、碱性lzip(βlzip)或两者。在一些方面,一个或多个lzip序列来源于天然存在的lzip序列。在一些方面,一个或多个lzip序列包含天然存在的lzip序列。在一些方面,一个或多个lzip序列是合成的。在某些方面,一个或多个lzip序列包含seq id no:4、7、
14、17、22或25所示的lzip序列。
[0297]
ii.a.7.接头
[0298]
在一些方面,可用于本公开的β链和/或α链进一步包含接头。本领域已知的任何接头均可用在本文公开的组合物和方法中。在某些方面,接头包括gly/ser接头。在一些方面,接头包含选自glyser、gly2ser、gly3ser和gly4ser的氨基酸序列。在一些方面,接头位于α链或β链的胞外结构域的n端。在一些方面,接头位于α链或β链的胞外结构域的c端。在一些方面,接头位于α链或β链的胞外结构域与跨膜结构域之间。在一些方面,接头位于α链或β链的胞外结构域与一个或多个lzip序列之间。在一些方面,接头位于α链或β链的胞外结构域与信号肽之间。
[0299]
任何长度的接头都可以用在本文公开的组合物和方法中。在一些方面,接头的长度为至少一个氨基酸。在一些方面,接头的长度为至少约1个到至少约100个、至少约1个到至少约90个、至少约1个到至少约80个、至少约1个到至少约70个、至少约1个到至少约60个、至少约1个到至少约50个、至少约1个到至少约40个、至少约1个到至少约30个、至少约1个到至少约20个、至少约1个到至少约15个、至少约1个到至少约14个、至少约1个到至少约13个、至少约1个到至少约12个、至少约1个到至少约11个、至少约1个到至少约10个、至少约1个到至少约9个、至少约1个到至少约8个、至少约1个到至少约7个、至少约1个到至少约6个、至少约1个到至少约5个、至少约1个到至少约4个、至少约1个到至少约3个氨基酸。
[0300]
在一些方面,接头的长度为至少约1个、至少约2个、至少约3个、至少约4个、至少约5个、至少约6个、至少约7个、至少约8个、至少约9个、至少约10个、至少约11个、至少约12个、至少约13个、至少约14个、至少约15个、至少约20个、至少约30个、至少约40个、至少约50个、至少约60个、至少约70个、至少约80个、至少约90个、至少约100个氨基酸。在一些方面,接头的长度为约3个氨基酸。在一些方面,接头的长度为约4个氨基酸。在一些方面,接头的长度为约5个氨基酸。
[0301]
ii.b.细胞
[0302]
在本公开的某些方面,本公开的方法中使用的mhc ii类分子与细胞的膜连接或缔合。因此,本公开的一些方面涉及鉴定mhc ii类特异性tcr的方法,所述方法包括使t细胞与细胞接触,其中所述细胞包含复合物,所述复合物包含本文公开的mhc ii类分子和肽(例如表位)。在某些方面,mhc ii类分子的β链与细胞的膜连接或缔合。在某些方面,mhc ii类分子的α链与细胞的膜连接或缔合。在某些方面,mhc ii类分子的α链和β链与细胞的膜连接或缔合。
[0303]
任何细胞都可以用于本文所述的方法中。在某些方面,细胞是哺乳动物细胞。在一些方面,细胞是昆虫细胞。在一些方面,细胞来源于健康细胞,例如健康成纤维细胞。在一些方面,细胞来源于肿瘤细胞。可用于本公开的细胞的非限制性示例包括k562细胞、t2细胞、hek293细胞、hek293t细胞、a375细胞、sk-mel-28细胞、me275细胞、cos细胞、成纤维细胞、肿瘤细胞或其任何组合。在某些方面,细胞是在hasan等人,adv.genet.eng.4(3):130(2015)(通过引用以其整体并入本文)中公开的任何细胞。
[0304]
在某些方面,细胞是专职apc。在某些方面,细胞是巨噬细胞、b细胞、树突细胞或其任何组合。
[0305]
在某些方面,细胞缺乏一种或多种mhc ii类等位基因的内源性表达。在一些方面,
细胞缺乏hla-dp等位基因的内源性表达。在一些方面,细胞缺乏hla-dpα链等位基因的内源性表达。在一些方面,细胞缺乏hla-dpβ链等位基因的内源性表达。
[0306]
ii.c.可溶性mhc ii类分子
[0307]
在某些方面,本文公开的方法中使用的mhc ii类分子不与细胞膜缔合,例如,mhc ii类分子是可溶形式。如本文所用,可溶性mhc ii类分子包括本文所述的不与细胞膜缔合的任何mhc ii类分子或其部分。在某些方面,mhc ii类分子或其部分不与任何膜结合。在一些方面,mhc ii类分子或其部分与惰性颗粒结合。在一些方面,mhc ii类分子或其部分结合到细胞外囊泡的膜上。在一些方面,mhc ii类分子结合到人造膜或人造表面上,例如阵列板的表面上。
[0308]
本领域已知的任何惰性颗粒均可用在本公开的组合物和方法中。在一些方面,惰性颗粒是珠。在一些方面,珠是玻璃珠、乳胶珠、金属珠或其任何组合。在一些方面,惰性颗粒是纳米粒子(np)。本领域已知的任何np均可用在本公开的组合物和方法中。在某些方面,纳米粒子选自聚乙二醇化氧化铁、壳聚糖、葡聚糖、明胶、藻酸盐、脂质体、淀粉、支化聚合物、碳基载体、聚乳酸、聚(氰基)丙烯酸酯、聚乙烯亚胺、嵌段共聚物、聚己内酯、spions、uspions、cd/zn-硒化物或二氧化硅纳米粒子。在特定方面,纳米粒子是聚乙二醇化氧化铁纳米粒子。可用在本文公开的组合物和方法中的纳米粒子的非限制性示例包括在de jong和borm,int.j.nanomedicine 3(2):133-49(2008)和umeshappa等人,nat.commun.10(1):2150(2019年5月14日)(各自通过引用以其整体并入本文)中阐述的那些。
[0309]
在一些方面,mhc ii类分子包括全长mhc ii类分子的片段,其中α链的跨膜结构域和/或β链的跨膜结构域的一个或多个氨基酸缺失。在一些方面,mhc ii类分子包含α链的胞外结构域(例如,如seq id no:6所示)和/或β链的胞外结构域(例如,如seq id no:1或3所示)。在一些方面,mhc ii类分子包含α链的胞外结构域(例如,如seq id no:16所示)和/或β链的胞外结构域(例如,如seq id no:11或13所示)。在一些方面,mhc ii类分子包含α链的胞外结构域(例如,如seq id no:24所示)和/或β链的胞外结构域(例如,如seq id no:19或21所示)。
[0310]
在某些方面,mhc ii类分子包含dpα链,所述dpα链包含与seq id no:6具有至少约80%、至少约85%、至少约90%、至少约95%、至少约96%、至少约97%、至少约98%或至少约99%序列同一性的氨基酸序列。在一些方面,mhc ii类分子包含dpα链,所述dpα链包含seq id no:6所示的氨基酸序列。
[0311]
在某些方面,mhc ii类分子包含dqα链,所述dqα链包含与seq id no:16具有至少约80%、至少约85%、至少约90%、至少约95%、至少约96%、至少约97%、至少约98%或至少约99%序列同一性的氨基酸序列。在一些方面,mhc ii类分子包含dqα链,所述dqα链包含seq id no:16所示的氨基酸序列。
[0312]
在某些方面,mhc ii类分子包含drα链,所述drα链包含与seq id no:24具有至少约80%、至少约85%、至少约90%、至少约95%、至少约96%、至少约97%、至少约98%或至少约99%序列同一性的氨基酸序列。在一些方面,mhc ii类分子包含drα链,所述drα链包含seq id no:24所示的氨基酸序列。
[0313]
在某些方面,mhc ii类分子包含dpβ链,所述dpβ链包含与seq id no:1具有至少约80%、至少约85%、至少约90%、至少约95%、至少约96%、至少约97%、至少约98%或至少
约99%序列同一性的氨基酸序列。在一些方面,mhc ii类分子包含dpβ链,所述dpβ链包含seq id no:1所示的氨基酸序列。在某些方面,mhc ii类分子包含dpβ链,所述dpβ链包含与seq id no:3具有至少约80%、至少约85%、至少约90%、至少约95%、至少约96%、至少约97%、至少约98%、或至少约99%序列同一性的氨基酸序列。在一些方面,mhc ii类分子包含dpβ链,所述dpβ链包含seq id no:3所示的氨基酸序列。在某些方面,mhc ii类分子包含dpβ链,所述dpβ链包含与seq id no:4具有至少约80%、至少约85%、至少约90%、至少约95%、至少约96%、至少约97%、至少约98%、或至少约99%序列同一性的氨基酸序列。在一些方面,mhc ii类分子包含dpβ链,所述dpβ链包含seq id no:4所示的氨基酸序列。在某些方面,mhc ii类分子包含dpβ链,所述dpβ链包含与seq id no:5具有至少约80%、至少约85%、至少约90%、至少约95%、至少约96%、至少约97%、至少约98%、或至少约99%序列同一性的氨基酸序列。在一些方面,mhc ii类分子包含dpβ链,所述dpβ链包含seq id no:5所示的氨基酸序列。
[0314]
在某些方面,mhc ii类分子包含dqβ链,所述dqβ链包含与seq id no:11具有至少约80%、至少约85%、至少约90%、至少约95%、至少约96%、至少约97%、至少约98%或至少约99%序列同一性的氨基酸序列。在一些方面,mhc ii类分子包含dqβ链,所述dqβ链包含seq id no:11所示的氨基酸序列。在某些方面,mhc ii类分子包含dqβ链,所述dqβ链包含与seq id no:13具有至少约80%、至少约85%、至少约90%、至少约95%、至少约96%、至少约97%、至少约98%或至少约99%序列同一性的氨基酸序列。在一些方面,mhc ii类分子包含dqβ链,所述dqβ链包含seq id no:13所示的氨基酸序列。在某些方面,mhc ii类分子包含dqβ链,所述dqβ链包含与seq id no:14具有至少约80%、至少约85%、至少约90%、至少约95%、至少约96%、至少约97%、至少约98%或至少约99%序列同一性的氨基酸序列。在一些方面,mhc ii类分子包含dqβ链,所述dqβ链包含seq id no:14所示的氨基酸序列。在某些方面,mhc ii类分子包含dqβ链,所述dqβ链包含与seq id no:15具有至少约80%、至少约85%、至少约90%、至少约95%、至少约96%、至少约97%、至少约98%或至少约99%序列同一性的氨基酸序列。在一些方面,mhc ii类分子包含dqβ链,所述dqβ链包含seq id no:15所示的氨基酸序列。
[0315]
在某些方面,mhc ii类分子包含drβ链,所述drβ链包含与seq id no:19具有至少约80%、至少约85%、至少约90%、至少约95%、至少约96%、至少约97%、至少约98%或至少约99%序列同一性的氨基酸序列。在一些方面,mhc ii类分子包含drβ链,所述drβ链包含seq id no:19所示的氨基酸序列。在某些方面,mhc ii类分子包含drβ链,所述drβ链包含与seq id no:21具有至少约80%、至少约85%、至少约90%、至少约95%、至少约96%、至少约97%、至少约98%或至少约99%序列同一性的氨基酸序列。在一些方面,mhc ii类分子包含drβ链,所述drβ链包含seq id no:21所示的氨基酸序列。在某些方面,mhc ii类分子包含drβ链,所述drβ链包含与seq id no:22具有至少约80%、至少约85%、至少约90%、至少约95%、至少约96%、至少约97%、至少约98%或至少约99%序列同一性的氨基酸序列。在一些方面,mhc ii类分子包含drβ链,所述drβ链包含seq id no:22所示的氨基酸序列。在某些方面,mhc ii类分子包含drβ链,所述drβ链包含与seq id no:23具有至少约80%、至少约85%、至少约90%、至少约95%、至少约96%、至少约97%、至少约98%或至少约99%序列同一性的氨基酸序列。在一些方面,mhc ii类分子包含drβ链,所述drβ链包含seq id no:23所
示的氨基酸序列。
[0316]
ii.d.核酸分子和载体
[0317]
本公开的某些方面涉及编码本文公开的mhc ii类分子的核酸分子。在一些方面,核酸分子编码本文公开的mhc ii类β链。在某些方面,编码mhc ii类β链的核酸分子包含与seq id no:2、12或20所示的序列具有至少约50%、至少约55%、至少约60%、至少约65%、至少约70%、至少约75%、至少约80%、至少约85%、至少约90%、至少约95%、至少约96%、至少约97%、至少约98%、或至少约99%序列同一性的核苷酸序列。
[0318]
在一些方面,核酸分子编码本文公开的mhc ii类α链。在某些方面,编码mhc ii类α链的核酸分子包含与seq id no:7、17或25所示的序列具有至少约50%、至少约55%、至少约60%、至少约65%、至少约70%、至少约75%、至少约80%、至少约85%、至少约90%、至少约95%、至少约96%、至少约97%、至少约98%、或至少约99%序列同一性的核苷酸序列。
[0319]
在一些方面,核酸分子编码本文公开的mhc ii类α链和本文公开的mhc ii类β链两者。在一些方面,编码mhc ii类α链的序列在与编码mhc ii类β链的序列相同的启动子的控制下。在一些方面,编码mhc ii类α链的序列在第一启动子的控制下,并且编码mhc ii类β链的序列在第二启动子的控制下。
[0320]
在一些方面,本公开涉及编码本文公开的mhc ii类β链的第一核酸分子和编码本文公开的mhc ii类α链的第二核酸分子。
[0321]
本公开的某些方面涉及包含本文公开的核酸分子的一种载体或一组载体。在一些方面,载体是病毒载体。在一些方面,载体是病毒颗粒或病毒。在一些方面,载体是哺乳动物载体。在一些方面,载体是细菌载体。
[0322]
在某些方面,载体是逆转录病毒载体。在一些方面,载体是腺病毒载体、慢病毒、仙台病毒、杆状病毒载体、爱泼斯坦巴尔病毒载体、乳多空病毒载体、痘苗病毒载体、单纯疱疹病毒载体或腺相关病毒(aav)载体。在特定方面,载体是aav载体。在一些方面,载体是慢病毒。在特定方面,载体是腺病毒载体。在一些方面,载体是仙台病毒。在一些方面,载体是杂合载体。可以在本公开中使用的杂合载体的示例可以在huang和kamihira,biotechnol.adv.31(2):208-23(2103)(通过引用以其整体并入本文)中找到。
[0323]
ii.e.治疗肿瘤的方法
[0324]
在某些方面,本文公开的方法还包括治疗有需要的受试者的癌症。在一些方面,所述方法进一步包括向有需要的受试者施用使用本文公开的方法鉴定的tcr,其中所述受试者患有癌症。在一些方面,所述方法包括向受试者施用细胞,其中所述细胞包含使用本文公开的方法鉴定的tcr。在一些方面,细胞是t细胞。
[0325]
在一些方面,癌症选自黑素瘤、骨癌、肾癌、前列腺癌、乳腺癌、结肠癌、肺癌、皮肤或眼内恶性黑素瘤、胰腺癌、皮肤癌、头颈癌、子宫癌、卵巢癌、直肠癌、肛区癌、胃癌、睾丸癌、子宫癌、输卵管癌、子宫内膜癌、宫颈癌、阴道癌、外阴癌、霍奇金病、非霍奇金淋巴瘤(nhl)、原发纵隔大b细胞淋巴瘤(pmbc)、弥漫性大b细胞淋巴瘤(dlbcl)、滤泡性淋巴瘤(fl)、转化型滤泡性淋巴瘤、脾边缘区淋巴瘤(smzl)、食道癌、小肠癌、内分泌系统癌、甲状腺癌、甲状旁腺癌、肾上腺癌、软组织肉瘤、尿道癌、阴茎癌、急性或慢性白血病、急性髓性白血病(aml)、慢性髓性白血病、急性成淋巴细胞性白血病(all)(包括非t细胞all)、慢性淋巴细胞性白血病(cll)、儿童实体瘤、淋巴细胞性淋巴瘤、膀胱癌、肾或输尿管癌、肾盂癌、中枢
神经系统(cns)瘤、原发性cns淋巴瘤、肿瘤血管生成、脊髓轴肿瘤、脑干胶质瘤、垂体腺瘤、卡波西肉瘤、表皮样癌、鳞状细胞癌、t细胞淋巴瘤、包括由石棉诱导的那些的环境诱导癌症、其他b细胞恶性肿瘤及这些癌症的任何组合。在一些方面,癌症是黑素瘤。
[0326]
在一些方面,癌症是复发性的。在一些方面,癌症是难治性的。在一些方面,癌症是晚期的。在一些方面,癌症是转移性的。
[0327]
在一些方面,本文公开的方法治疗受试者的癌症。在一些方面,本文公开的方法降低癌症的一种或多种症状的严重性。在一些方面,本文公开的方法减少来源于所述癌症的肿瘤的大小或数量。在一些方面,相对于未提供本文公开的方法的受试者,本文公开的方法增加了受试者的总生存期。在一些方面,相对于未提供本文公开的方法的受试者,本文公开的方法增加了受试者的无进展生存期。在一些方面,本文公开的方法导致受试者的部分反应。在一些方面,本文公开的方法导致受试者的完全反应。
[0328]
本公开的某些方面涉及治疗有需要的受试者的感染的方法,所述方法包括向受试者施用本文公开的hla ii类分子、本文公开的核酸分子、本文公开的载体或本文公开的细胞。可以使用本文公开的组合物和方法治疗的感染的非限制性示例包括病毒(包括类病毒和朊病毒)、细菌、真菌、寄生虫或其任何组合的感染。在一些方面,病毒是疱疹病毒、hiv、乳多空病毒、麻疹病毒、风疹病毒、人乳头瘤病毒(hpv)、人t淋巴细胞病毒1、爱泼斯坦-巴尔病毒、甲型肝炎病毒、乙型肝炎病毒、丙型肝炎病毒、流感病毒、诺如病毒及其任何组合。在一些方面,细菌选自链球菌、葡萄球菌和大肠杆菌。在一些方面,细菌感染选自布鲁氏菌病、弯曲杆菌感染、猫抓病、霍乱、大肠杆菌感染、淋病、克雷伯氏菌感染、肠杆菌感染、沙雷氏菌感染、军团菌感染、脑膜炎球菌感染、百日咳、鼠疫、假单胞菌感染、沙门氏菌感染、志贺氏菌病、伤寒、土拉菌病、炭疽病、白喉、肠球菌感染、丹毒丝菌病(erysipelothricosis)、李斯特菌病、奴卡菌病、肺炎球菌感染、葡萄球菌感染、链球菌感染及其任何组合。在一些实施方案中,寄生虫感染选自蛲虫病、滴虫病、弓形虫病、贾第鞭毛虫病、隐孢子虫病、疟疾、钩虫病、癣菌病、绦虫病、吸虫病及其任何组合。在一些方面,真菌感染选自念珠菌属、糠秕马拉色菌、皮肤癣菌(例如,表皮癣菌属、小孢子菌属和毛癣菌属)或其任何组合。
[0329]
在一些方面,本文公开的方法包括治疗有需要的受试者的癌症或感染,所述方法包括向受试者施用本文所述的细胞,其中所述细胞包含本文公开的mhc ii类分子、本文公开的核酸分子、本文公开的载体或其任何组合。
[0330]
在一些方面,细胞获自受试者。在一些方面,细胞获自除受试者之外的供体。
[0331]
本文描述的所有各个方面、方面和选项可以以任何和所有变化形式组合。
[0332]
本说明书中提及的所有出版物、专利和专利申请在此通过引用并入,就如每个单独的出版物、专利或专利申请被具体地和单独地指出通过引用并入本文的程度一样。
[0333]
已经一般性地描述了本公开,通过参考本文提供的实施例可以获得进一步的理解。这些实施例仅用于说明的目的,而不具有限制性。
[0334]
实施例
[0335]
实施例1
–
亲和力成熟hla-dp分子的产生
[0336]
细胞
[0337]
通过密度梯度离心获得外周单核细胞(ficoll-paque plus,ge healthcare life sciences,marlborough,ma)。k562细胞系是一种红白血病细胞系,具有缺陷型hla i/ii类
表达。先前已经报道了分别表达作为单个hla等位基因的各种hla ii类基因连同cd80和cd83的基于k562的人工apc(aapc)(butler等人,plos one 7,e30229(2012))。jurkat 76细胞系是一种t细胞白血病细胞系,缺乏内源性tcr、cd4和cd8表达。jurkat 76/cd4细胞通过逆转录病毒转导人cd4基因生成。a375、sk-mel-21、sk-mel-28、sk-mel-37和me275是黑素瘤细胞系。使hek293t细胞和黑素瘤细胞系在补充有10%fbs和50μg/ml庆大霉素(thermo fisher scientific,waltham,ma)的dmem中生长。将k562和jurkat 76细胞系在补充有10%fbs和50μg/ml庆大霉素的rpmi 1640中培养。
[0338]
肽
[0339]
合成肽购自genscript(piscataway,nj),并以50μg/ml溶解在dmso中。肽序列如表6所示。
[0340]
表6:合成肽序列
[0341]
[0342]
[0343]
[0344]
[0345]
[0346]
[0347][0348]
基因
[0349]
使用smarter race 5'/3'试剂盒(takara bio,shiga,japan)通过cdna末端的5'-快速扩增(race)pcr克隆新的tcr基因,并如前所述进行测序。将所有基因都克隆到pmx逆转录病毒载体中,并使用基于293gpg和pg13细胞的逆转录病毒系统转导到细胞系中。
[0350]
抗体
[0351]
将以下抗体用于流式细胞术分析:pe缀合的抗ii类(9-49(i3))、apc-cy7缀合的抗cd4(rpa-t4,biolegend,san diego,ca)
44
、fitc缀合的抗ngfr(me20.4,biolegend,san diego,ca)、pe缀合的抗his标签(ad1.1.10,abcam,cambridge,ma)和fitc缀合的抗vβ22
(immu 546,beckman coulter,brea,ca)。根据制造商的说明,使用pe缀合的链霉亲和素(thermo fisher scientific,waltham,ma),将生物素化的dp4/ny-eso1
157-170
和dp4/wt1
329-348
单体多聚化。用活/死可固定近红外死细胞染色试剂盒465(live/dead fixable near-ir dead cell stain kit 465)(thermo fisher scientific,waltham,ma)区分死细胞。用canto ii或lsrfortessa x-20(bd biosciences,franklin lakes,nj)分析染色的细胞。使用facs aria ii(bd biosciences,franklin lakes,nj)进行细胞分选。使用flowjo软件(tree star,ashland,or)进行数据分析。
[0352]
将以下抗体用于免疫印迹分析:抗β-肌动蛋白(c4,santa cruz biotechnology,santa cruz,ca)、兔多克隆抗mage-a2(abcam,cambridge,ma)、抗ccnd1(epr2241,abcam,cambridge,ma)、hrp缀合的山羊抗小鼠igg(h+l)二抗(promega,fitchburg,wi)和hrp缀合的抗兔igg(h+l)二抗(promega,fitchburg,wi)。
[0353]
tcr转导至原代t细胞中
[0354]
分别使用pan t细胞分离试剂盒(miltenyi biotec,bergisch gladbach,germany)和cd4
+
t细胞分离试剂盒(miltenyi biotec,bergisch gladbach,germany)纯化cd3
+
和cd4
+
t细胞。使用以20:1的e:t比用200gy辐照的aapc/mokt3刺激纯化的t细胞。从第二天开始,通过连续3天在32℃下以1,000
×
g离心1小时或使用retronectin涂层板(takara bio,shiga,japan),用克隆的tcr基因逆转录病毒转导激活的t细胞。在第二天,将100iu/ml il-2和10ng/ml il-15添加到tcr转导的t细胞中。每2-3天补足培养基。
[0355]
用可溶性cd4染色
[0356]
可溶性cd4(scd4)基因是通过gs接头将人cd4胞外结构域与6xhis标签融合生成。用scd4基因逆转录病毒转导hek293t细胞,并收获含有scd4单体的培养上清液。将scd4用pe标记的抗6xhis标签mab(ad1.1.10,abcam,cambridge,ma)二聚化并使用。将表达hla ii类的k562细胞在山羊血清存在下在室温下用二聚化scd4染色30分钟。分别表达各种ii类基因的k562来源细胞中的表面hla ii类表达如图13a至图13q所示。
[0357]
多位点定向dpb1*04:01突变型cdna文库的构建与筛选
[0358]
通过使用pcr和以下引物组将多位点定向随机突变插入dpb1*04:01 cdna:对于l112和v114,正向:5'-caccacaacnnncttnnntgccacgtg-3'(seq id no:30)和反向:5'-cacgtggcannnaagnnngttgtggtg-3'(seq id no:31);对于v141,正向:5'-acagctggggtcnnntccaccaacctg-3'(seq id no:32)和反向:5'-caggttggtggannngaccccagctgt-3'(seq id no:33);对于l156和m158,正向:5'-cagatcnnngtgnnnctggaaatgacc-3'(seq id no:34)和反向:5'-ggtcatttccagnnncacnnngatctg-3'(seq id no:35)。n代表任何核苷酸。将得到的pcr片段相互融合以构建突变型全长dpb1*04:01 cdna表达文库,该文库在位置l112、v114、v141、l156和m158处携带随机突变。用包装细胞系293gpg产生的重组逆转录病毒以低于30%的转导效率感染稳定表达dpa1*01:03基因的k562细胞。将感染的k562细胞用可溶性cd4二聚体染色,并使用流式细胞术细胞分选仪收集二聚体阳性细胞。从收集的细胞中克隆突变型dpb1*04:01基因,并与上述野生型dpa1*01:03基因一起通过逆转录病毒转导到k562细胞中。
[0359]
hla ii类单体和二聚体的生成
[0360]
将野生型ii类α基因的胞外结构域通过gggs接头与酸性亮氨酸拉链融合,随后通过gs接头与6xhis标签融合(参见seq id no:8)。将携带突变的ii类β基因的胞外结构域(参见seq id no:3)通过gggs接头(参见seq id no:4)与碱性亮氨酸拉链类似地连接。使用基于293gpg细胞的逆转录病毒系统用α和β基因转染hek293t细胞,并在补充有10%fbs和50μg/ml庆大霉素的dmem中培养。对于dp4二聚体染色,使稳定分泌可溶性dp4
l112w/v141m
蛋白的hek293t细胞生长至汇合,并将培养基更换为无血清293 sfm ii培养基(thermo fisher scientific,waltham,ma)。48小时后,收获条件培养物并使用amicon超滤器(截留分子量(mwco)10kda)(milliporesigma,burlington,ma)浓缩。然后在37℃下将含有可溶性hla ii类的上清液与100μg/ml目标肽混合20-24小时,以进行体外肽交换。将未进行肽交换的单体用作对照。使用镍涂层板(xpressbio,frederick,md)和抗his标签生物素化mab(ad1.1.10,r&d systems,minneapolis,mn)通过特异性elisa测量单体的浓度。使用pe缀合的抗his mab(ad1.1.10,abcam,cambridge,ma)以2:1的摩尔比在4℃下将可溶性hla ii类单体二聚化1.5小时以进行染色。
[0361]
dp4限制性抗原特异性cd4
+
t细胞的刺激
[0362]
使用cd4
+
t细胞分离试剂盒(miltenyi biotec,bergisch gladbach,germany)纯化cd4
+
t细胞。将纯化的t细胞用通过dp4限制性肽以10μg/ml冲击的表达dp4的aapc刺激,并以20:1的e:t比率在200gy下辐照。四十八小时后,将10iu/ml il-2和10ng/ml il-15添加到cd4
+
t细胞中。每2-3天补足补充有il-2(10iu/ml)和il-15(10ng/ml)的培养基。刺激2周后,对t细胞进行dp4
l112w/v141m
二聚体染色。
[0363]
hla ii类二聚体和四聚体染色
[0364]
将用外源性tcr基因转导的原代t细胞和jurkat 76/cd4 t细胞在37℃下用50nm达沙替尼(lc laboratories,woburn,ma)预处理30分钟,并在室温下用5-15μg/ml ii类二聚体染色4-5小时。洗涤后,用apc-cy7缀合的抗cd4 mab、fitc缀合的抗ngfr mab和pe缀合的抗vβ22 mab对细胞表面分子进行复染。
[0365]
elispot测定
[0366]
如先前报道的,进行细胞因子elispot测定(参见例如yamashita等人,nat.commun.8:15244(2017);和anczurowski等人,sci.rep.8:4804(2018))。
[0367]
免疫印迹
[0368]
如先前报道的,进行免疫印迹分析(参见例如,yamashita等人,nat.commun.8:15244(2017);和anczurowski等人,sci.rep.8:4804(2018))。
[0369]
蛋白质建模
[0370]
使用用于四元结构预测的swiss-model工作区基于pdb id 3s5l和3t0e的结构预测hla-dp4和人cd4复合物模型结构。
[0371]
统计分析
[0372]
使用graphpad prism 6.0软件(graphpad software,san diego,ca)进行统计分析。使用非配对的双尾学生t检验来进行两样本比较。没有使用统计方法来预先确定样本量。在实验或结果评估期间,研究人员对分配不是盲的。实验不是随机化的。
[0373]
生物层干涉传感图
[0374]
后接gs接头和10x组氨酸(his)标签的人cd4的胞外结构域(np_000607.1的残基
26-440)在人细胞系a375中稳定表达(seq id nos:262-263;表7)。用talon金属亲和树脂(takara bio,shiga,japan)从上清液中纯化重组的10x his标记的cd4蛋白。使用具有10kda mwco的amicon ultra-15离心柱(milliporesigma,burlington,ma)浓缩洗脱的蛋白质。使用10kda mwco迷你透析器(mini dialyzer)(thermo fisher scientific,waltham,ma)将缓冲液交换为hbs-ep(ge healthcare life sciences,marlborough,ma)。如sds-page所证实的,重组cd4蛋白的纯度始终》90%。
[0375]
重组dp4蛋白由dpa1*01:03的胞外结构域和野生型dpb1*04:01或l112w/v141m突变体组成。dpa1*01:03后接酸性亮氨酸拉链、gs接头和10x组氨酸标签,而野生型和突变型dpb1后接碱性亮氨酸拉链、gs接头和生物素化序列(glndifeaqkiewhe;seq id no:265)。dpa和dpb基因均在a375-bira细胞中稳定表达,将这些细胞用编码5'端前导序列和3'端er保留kdel基序的密码子优化的bira基因进行转导。用talon金属亲和树脂(takara bio,shiga,japan)从上清液中纯化重组的dp4蛋白。使用具有10kda mwco的vivaspin 500离心柱(ge healthcare life sciences,marlborough,ma)浓缩洗脱的蛋白质,并复原至在pbs中的工作体积。
[0376]
通过octet red系统(fortebio,fremont,ca)测量野生型dp4和dp4
l112w/v141m
与cd4的结合。使用96孔optiplate(perkin elmer,waltham,ma)在25℃下进行实验,样品体积为200μl,并在1,000rpm下持续摇动。将生物素化的重组dp4加载到链霉亲和素涂覆的生物传感器(fortebio,fremont,ca)上直至饱和,随后在hbs-ep缓冲液中进行基线测量。通过以下方式来测量缔合:将加载的传感器与滴定浓度的重组cd4(0.8125至26μm)一起孵育400秒,之后在单独的hbs-ep缓冲液中解离300秒。使用graphpad prism 7.0中的单位点特异性结合模型拟合稳态分析
[0377]
表7.可溶性10x his标记的cd4核酸序列
[0378][0379]
实施例2
–
dpβ链的l112w/v141m取代增强dp与cd4的结合
[0380]
生成了在l112、v114、v141、l156和m158(分别对应于dr1β链的l114、v116、v143、l158和m160)处携带随机突变的dpb1*04:01(dp4β)基因的cdna表达文库,并在ii类缺陷型k562细胞中与野生型dpa1*01:03(dpα)基因一起共表达所述文库。在使用可溶性cd4蛋白(scd4)进行两轮筛选后,分离出具有增强的cd4结合的细胞群,从中分子克隆了携带l112w、v114m、v141m和m158i取代的突变型dp4β基因。当在k562细胞中异位表达时,由野生型dpα链和携带l112w、v114m、v141m和m158i取代(dp4
l112w/v114m/v141m/m158i
)的克隆的突变型dp4β链组成的突变型dp4分子确实显示与野生型dp4分子相比增强的与scd4的结合,从而排除了增强的cd4结合是筛选过程的人工产物的可能性(图1a至图1f)。
[0381]
为了确定四种突变中的哪一种对增强cd4结合至关重要,进行了回复诱变研究。将所有可能的回复dp4突变体都在ii类阴性k562细胞上重构并用scd4染色。l112w和v141m两者而非v114m或m158i单取代分别增强了dp4与scd4的结合(图1g)。重要的是,l112w/v141m双突变(dp4
l112w/v141m
)协同增强了dp4/cd4结合(图1g)。有趣的是,v114m和m158i单替换显现都对dp4
l112w/v141m
突变实现的增强结合具有负面影响(图1g)。以前的研究估计cd4和hla ii类之间的kd值》2mm。使用生物层干涉(bli)结合测定,测量了dpr
l112/v141m
对cd4的亲和力。虽然在野生型dp4和cd4之间未检测到结合,但dp4
l112w/v141m
以8.9μm
±
1.1的kd与cd4结合(图1h和图1x至图1bk)。该值表示结合亲和力至少提高了200倍。此外,观察到的cd4和dp4
l112w/v141m
之间的亲和力高于人cd8和hla i类之间的亲和力(约200μm),并且与小鼠cd8和小鼠mhc i类之间的亲和力(约10μm)相当。为确认dp4
l112w/v141m
和cd4之间增强的结合导致增强的cd4
+
t细胞反应,使用dp4/wt1 tcr(克隆9)转导的cd4-和cd4
+
jurkat 76 t细胞作为应答细胞,对表达作为单个ii类等位基因的野生型dp4或dp4
l112w/v141m
的人工apc(aapc)的免疫刺激能力进行了比较。正如所料,携带dp4
l112w/v141m
的aapc表现出以cd4依赖性方式的增强的t细胞刺激活性(图1i)。
[0382]
接下来分析cd4的其他dp等位基因以确定l112w/v141m突变是否也增强结合。尽管野生型dp2、dp5或dp8均不与cd4结合,但当将l112w/v141m双突变引入这些分子的dpβ链中时,所有三种分子都与cd4强烈结合(图1i至图1w)。基于先前报道构建的结构模型(图2a至图2d)揭示,在dp4
l112w/v141m-cd4复合物中,两个l112w/v141m突变明显在cd4的k35、q40和t45的位置诱导疏水效应。这些结果表明,l112w/v141m突变可以增强至少所有4个所测试的dp等位基因的cd4结合。
[0383]
实施例3
–
亲和力成熟的dp4
l112w/v141m
多聚体特异性染色同源tcr
[0384]
为了确定dp4β的l112w/v141m双突变对dp4多聚体染色的影响,产生了可溶性dp4
l112w/v141m
单体,然后将其与抗his标签mab进行二聚化。将原代t细胞分别用对mage-a3(克隆r12c9)、wt1(克隆9)和ny-eso-1(克隆5b8)有特异性的三种不同的dp4限制性tcr分别转导,然后用同源dp4
l112w/v141m二
聚体染色。如图3a至图3p所示,每种dp4
l112w/v141m
二聚体特异性染色表达同源tcr的cd4
+
t细胞。分别用抗vβ22 mab和抗ngfr mab连同相应的dp4
l112w/v141m
二聚体对r12c9和克隆9转导的t细胞进行复合染色,证实几乎所有tcr转导的cd4
+
t细胞都成功地用相应的dp4
l112w/v141m
二聚体染色(图4a至图4h)。与常规的野生型dp4四聚体相比,我们的新的dp4
l112w/v141m
二聚体对dp4/wt1和dp4/ny-eso-1t细胞两者的染色优于常规野生型dp4四聚体(图5a至图5p)。值得注意的是,即使在可用的最高浓度下,常规的野生型dp4/ny-eso-1四聚体也无法染色同源t细胞(数据未显示)。
[0385]
实施例4
–
dp4
l112w/v141m
二聚体技术稳健且通用
[0386]
为了证明dp4
l112w/v141m
多聚体染色的稳健性和多功能性,针对来源于一系列肿瘤相关抗原的潜在dp4限制性肽的体外免疫原性进行了全面筛选(表6)。使用肽预测算法(netmhc2 ver.2.2)预测了196种dp4限制性和肿瘤相关抗原来源的20-mer肽并进行化学合成(表6)。外周抗原特异性cd4
+
t细胞的频率一般很低;因此,将从6名dp4
+
黑素瘤患者分离的原代cd4
+
t细胞仅用dp4-aapc刺激一次,分别用196种肽冲击并用同源dp4
l112w/v141m
二聚体染色。为了避免潜在的体外引发,使用了弱刺激条件。如图6a至图6f所示,103种预测的dp4肽至少在体外具有免疫原性。
[0387]
为了验证二聚体染色结果,我们自二聚体阳性t细胞克隆了对ccnd1
219-238
、hsd17b12
225-244
、lgsn
296-315
、mage-a2
108-127
和muc5ac
4922-4941
有特异性的七种dp4限制性tcr基因(图7a至图7l和表8)。当在人cd4
+
tcr缺陷型t细胞中进行克隆型重构时,所有这些tcr都被同源dp4
l112w/v141m
二聚体成功染色(图8a至图8x),并以dp4限制和抗原特异性方式发挥功能(图9a至图9g)。
[0388]
在原代t细胞中分别表达的四种tcr中,三种tcr,即03-ccnd1
219-238
、06-mage-a2
108-127
和05-muc5ac
4922-4941
,能够识别内源性加工并且由dp4呈递的同源肽(图10a至图10q和图11a至图11e)。重要的是,06-mage-a2
108-127
转导的原代t细胞能够以dp4和mage-a2依赖
性方式识别黑素瘤细胞系(图12a至图12e)。
[0389]
表8:dp4限制性tcr
[0390][0391][0392]
与cd8相比,cd4作为辅助受体的作用和功能尚未完全阐明。这种信息缺乏主要是因为cd4与ii类之间的结合异常弱,这极大地限制了对cd4与ii类之间缔合作用的研究。在这项研究中,通过增强的cd4结合分离了亲和力成熟形式的hla-dp4,即dp4
l112w/v141m
,并开发了新的dp4
l112w/v141m
二聚体技术,该技术引入了检测dp4限制性抗原特异性cd4
+
t细胞的稳健性和严格性。
[0393]
使用这种dp4
l112w/v141m
二聚体技术,在体外全面研究了dp4限制性抗肿瘤t细胞反应,并鉴定了多种dp4限制性免疫原性肽和同源tcr基因。hla-dp4是许多种族群体中最普遍的hla等位基因,属于dp
84gly
群。与其他ii类分子不同,dp
84gly
分子如dp4组成性地呈递来源于内源性来源的肽,而不管恒定链和hla-dm表达如何。通过ii类改善内源性肽的呈递与癌症患者的生存期改善相关。值得注意的是,首次人体ii类限制性tcr基因疗法确实靶向dp4限制性mage-a3肽(参见例如yao等人,j.immunother.39:191-201(2016))。dp
84gly
基因型(如
dp2和dp4)充当抗中性粒细胞胞质自身抗体相关血管炎的风险等位基因。dp4分子可以组成性地呈递来源于内源性肿瘤相关抗原的肽,它可能比其他ii类分子诱导更多的临床相关抗肿瘤反应,充当保护性ii类等位基因。
[0394]
为了鉴定亲和力成熟的ii类分子,本实施例详细说明了β链而非α链中的多个突变,因为β链比α链具有更直接的与cd4的相互作用。α链和/或β链的另外的突变可能会进一步增强ii类和cd4之间的结合。然而,使用这种具有过度cd4结合能力的可溶性ii类分子可能会导致cd4
+
t细胞的非特异性染色,从而具有不利影响。
[0395]
总之,cd4
+
t细胞在自身免疫性疾病的发展以及对病原体感染和癌症的防护中发挥着关键作用。本文所述的新型hla ii类多聚体技术可以更好地促进对hla-dp等位基因的hla ii类限制性cd4
+
t细胞反应的研究。
[0396]
实施例5
–
亲和力成熟hla-dq分子的产生
[0397]
细胞
[0398]
通过密度梯度离心获得外周单核细胞(ficoll-paque plus,ge healthcare life sciences,marlborough,ma)。k562细胞系是一种红白血病细胞系,具有缺陷型hla i/ii类表达。a375是黑素瘤细胞系。使hek293t细胞和a375细胞在补充有10%fbs和50μg/ml庆大霉素(thermo fisher scientific,waltham,ma)的dmem中生长。将k562和jurkat 76细胞系在补充有10%fbs和50μg/ml庆大霉素的rpmi1640中培养。
[0399]
肽
[0400]
合成肽购自genscript(piscataway,nj),并以50μg/ml溶解在dmso中。
[0401]
抗体
[0402]
将以下抗体用于流式细胞术分析:pe缀合的抗ii类(9-49(i3),beckman coulter,brea,ca;t
ü
39)、apc-cy7缀合的抗cd4(rpa-t4,biolegend,san diego,ca)和pe缀合的抗his标签(ad1.1.10,abcam,cambridge,ma)。用活/死可固定近红外死细胞染色试剂盒465(thermo fisher scientific,waltham,ma)区分死细胞。用canto ii或lsrfortessa x-20(bd biosciences,franklin lakes,nj)分析染色的细胞。使用facs aria ii(bd biosciences,franklin lakes,nj)进行细胞分选。使用flowjo软件(tree star,ashland,or)进行数据分析。
[0403]
tcr转导至原代t细胞中
[0404]
分别使用pan t细胞分离试剂盒(miltenyi biotec,bergisch gladbach,germany)和cd4
+
t细胞分离试剂盒(miltenyi biotec,bergisch gladbach,germany)纯化cd3
+
和cd4
+
t细胞。使用以20:1的e:t比用200gy辐照的aapc/mokt3刺激纯化的t细胞。从第二天开始,通过连续3天在32℃下以1,000
×
g离心1小时或使用retronectin涂层板(takara bio,shiga,japan),用克隆的tcr基因逆转录病毒转导激活的t细胞。在第二天,将100iu/ml il-2和10ng/ml il-15添加到tcr转导的t细胞中。每2-3天补足培养基。
[0405]
用可溶性cd4染色
[0406]
可溶性cd4(scd4)基因是通过gs接头将人cd4胞外结构域与6xhis标签融合生成。用scd4基因逆转录病毒转导hek293t细胞,并收获含有scd4单体的培养上清液。将scd4用pe标记的抗6xhis标签mab(ad1.1.10,abcam,cambridge,ma)二聚化并使用。将表达hla ii类的k562细胞在山羊血清存在下在室温下用二聚化scd4染色30分钟。分别表达各种ii类基因
的k562来源细胞中的表面hla ii类表达如图16a至图16q所示。
[0407]
hla ii类单体和二聚体的生成
[0408]
将野生型ii类α基因的胞外结构域通过gggs接头与酸性亮氨酸拉链融合,随后通过gs接头与6xhis标签融合(参见seq id no:18)。将携带突变的ii类β基因的胞外结构域(参见seq id no:13)通过gggs接头(参见seq id no:14)与碱性亮氨酸拉链类似地连接。使用基于293gpg细胞的逆转录病毒系统用α和β基因转染hek293t细胞和a375细胞,并在补充有10%fbs和50μg/ml庆大霉素的dmem中培养。对于二聚体染色,使稳定分泌可溶性dq5
l114w/v143m+4reps
(除l114w/v143m外,还具有n110q/i116v/s118h/p146n替换(4reps))和dq6
l114w/v143m+3reps
(除l114w/v143m外,还具有n110q/s118h/p146n替换(3reps))蛋白的a375细胞生长至汇合,并在48小时后收获培养基。然后在37℃下将含有可溶性hla ii类的上清液与100μg/ml目标肽混合20-24小时,以进行体外肽交换。将未进行肽交换的单体用作对照。使用镍涂层板(xpressbio,frederick,md)和抗his标签生物素化mab(ad1.1.10,r&d systems,minneapolis,mn)通过特异性elisa测量单体的浓度。使用pe缀合的抗his mab(ad1.1.10,abcam,cambridge,ma)以2:1的摩尔比在4℃下将可溶性hla ii类单体二聚化1.5小时以进行染色。
[0409]
hla ii类二聚体染色
[0410]
将用外源性tcr基因转导的原代t细胞在37℃下用50nm达沙替尼(lc laboratories,woburn,ma)预处理30分钟,并在室温下用5-15μg/ml ii类二聚体染色4-5小时。洗涤后,用apc-cy7缀合的抗cd4 mab对细胞表面分子进行复染。
[0411]
统计分析
[0412]
使用graphpad prism 6.0软件(graphpad software,san diego,ca)进行统计分析。使用非配对的双尾学生t检验来进行两样本比较。没有使用统计方法来预先确定样本量。在实验或结果评估期间,研究人员对分配不是盲的。实验不是随机化的。
[0413]
实施例6
–
具有增强的cd4结合能力的dq分子
[0414]
通过引入l114w/v143m突变生成亲和力增强的dq分子,以确定这些取代是否可以改善hla-dq分子如dq5(dqa1*01:01-dqb1*05:01)与cd4的结合。除了位置114和143之外,dqb1*05:01还编码在位置110、116、118和146处的四种不同的氨基酸。因此,我们产生了表达dq5
l114w/v143m+4reps
的k562细胞,除了l114w/v143m(图14a)外,它还具有n110q/i116v/s118h/p146n替换(4reps),并用scd4对这些细胞进行染色。表达dq5
l114w/v143m+4reps
但不表达dq5
l114w/v143m
、dq5
4reps
或野生型dq5的k562细胞表现出增强的cd4结合(图14b至图14c)。重要的是,分别表达在四个位置之一具有单个氨基酸反转的各种dq5
l114w/v143m+4reps
突变体的一系列k562细胞缺乏增强的cd4结合能力(图14d)。这些结果表明,n110q、i116v、s118h和p146n处的四个另外的替换对于观察到的增强的dq5:cd4结合中l114w/v143m突变的有效性至关重要。
[0415]
dqβ链如dqb1*02:01、04:02和06:01编码在位置110、118和146处而非位置116处的不同的氨基酸(图14e)。与dqb1*05:01不同,dqb1*02:01、04:02和06:01编码在位置116处的val,类似于dpb1*04:01,其编码位置114处的val。所有dq2
l114w/v143m+3reps
、dq4
l114w/v143m+3reps
和dq6
l114w/v143m+3reps
突变体(其β链携带n110q、s118h和p146n替换(3reps)连同l114w/v143m)显示增强的cd4结合活性(图14f)。
[0416]
实施例7
–
亲和力成熟的dq二聚体特异性和稳健地染色同源tcr
[0417]
评估了携带实施例2中描述的突变的亲和力成熟的dq二聚体鉴定抗原特异性cd4
+
t细胞的能力。dq5
l114w/v143m+4reps
和dq6
l114w/v143m+3reps
二聚体分别成功染色dq5限制性ddx3y特异性tcr(e6)和dq6限制性流感病毒特异性tcr(dm2)(图15a至图15b)。
[0418]
为了鉴定亲和力成熟的ii类分子,本实施例详细说明了β链而非α链中的多个突变,因为β链比α链具有更直接的与cd4的相互作用。α链和/或β链的另外的突变可能会进一步增强ii类和cd4之间的结合。然而,使用这种具有过度cd4结合能力的可溶性ii类分子可能会导致cd4
+
t细胞的非特异性染色,从而具有不利影响。
[0419]
总之,cd4
+
t细胞在自身免疫性疾病的发展以及对病原体感染和癌症的防护中发挥着关键作用。本文所述的新型hla ii类多聚体技术可以更好地促进对hla-dq等位基因的hla ii类限制性cd4
+
t细胞反应的研究。
[0420]
实施例8
–
亲和力成熟hla-dr分子的产生
[0421]
细胞
[0422]
通过密度梯度离心获得外周单核细胞(ficoll-paque plus,ge healthcare life sciences,marlborough,ma)。k562细胞系是一种红白血病细胞系,具有缺陷型hla i/ii类表达。先前已经报道了分别表达作为单个hla等位基因的各种hla ii类基因连同cd80和cd83的基于k562的人工apc(aapc)(butler等人,plos one 7,e30229(2012))。使hek293t细胞在补充有10%fbs和50μg/ml庆大霉素(thermo fisher scientific,waltham,ma)的dmem中生长。将k562细胞在补充有10%fbs和50μg/ml庆大霉素的rpmi 1640中培养。
[0423]
肽
[0424]
合成肽购自genscript(piscataway,nj),并以50μg/ml溶解在dmso中。
[0425]
抗体
[0426]
将以下抗体用于流式细胞术分析:pe缀合的抗ii类(9-49(i3))、apc-cy7缀合的抗cd4(rpa-t4,biolegend,san diego,ca)
44
、pe缀合的抗his标签(ad1.1.10,abcam,cambridge,ma)和fitc缀合的抗vβ22(immu 546,beckman coulter,brea,ca)。用活/死可固定近红外死细胞染色试剂盒465(thermo fisher scientific,waltham,ma)区分死细胞。用canto ii或lsrfortessa x-20(bd biosciences,franklin lakes,nj)分析染色的细胞。使用facs aria ii(bd biosciences,franklin lakes,nj)进行细胞分选。使用flowjo软件(tree star,ashland,or)进行数据分析。
[0427]
tcr转导至原代t细胞中
[0428]
使用cd4
+
t细胞分离试剂盒(miltenyi biotec,bergisch gladbach,germany)纯化cd4
+
t细胞。使用以20:1的e:t比用200gy辐照的aapc/mokt3刺激纯化的t细胞。从第二天开始,通过连续3天在32℃下以1,000
×
g离心1小时或使用retronectin涂层板(takara bio,shiga,japan),用克隆的tcr基因逆转录病毒转导激活的t细胞。在第二天,将100iu/ml il-2和10ng/ml il-15添加到tcr转导的t细胞中。每2-3天补足培养基。
[0429]
用可溶性cd4染色
[0430]
可溶性cd4(scd4)基因是通过gs接头将人cd4胞外结构域与6xhis标签融合生成。用scd4基因逆转录病毒转导hek293t细胞,并收获含有scd4单体的培养上清液。将scd4用pe标记的抗6xhis标签mab(ad1.1.10,abcam,cambridge,ma)二聚化并使用。将表达hla ii类
bio,shiga,japan)从上清液中纯化重组dr1蛋白。使用具有10kda mwco的vivaspin 500离心柱(ge healthcare life sciences,marlborough,ma)浓缩洗脱的蛋白质,并复原至在pbs中的工作体积。
[0442]
通过octet red系统(fortebio,fremont,ca)测量野生型dr1和dr1
l114w/v143m+2reps
与cd4的结合。使用96孔optiplate(perkin elmer,waltham,ma)在25℃下进行实验,样品体积为200μl,并在1,000rpm下持续摇动。将生物素化的重组dr1加载到链霉亲和素涂覆的生物传感器(fortebio,fremont,ca)上直至饱和,随后在hbs-ep缓冲液中进行基线测量。通过以下方式来测量缔合:将加载的传感器与滴定浓度的重组cd4(0.8125至26μm)一起孵育400秒,之后在单独的hbs-ep缓冲液中解离300秒。使用graphpad prism 7.0中的单位点特异性结合模型拟合稳态分析。
[0443]
实施例9
–
具有增强的cd4结合能力的dr分子
[0444]
通过引入l114w/v143m突变生成亲和力增强的dr分子,以确定这些取代是否可以改善hla-dr分子如dr1等位基因(dra1*01:01-drb1*01:01)与cd4的结合。除了位置114和143之外,drb1*01:01还编码在位置118、139、146、157、163和164处的六种不同的氨基酸(图17a)。与dr1
l114w/v143m
和野生型dr1相比,dr1
l114w/v143m+6reps
显示出增强的cd4结合(图17b和图17c)。在s118h或t157i而非其他4个位置处有单个氨基酸反转的dr1
l114w/v143m+6reps
来源突变体文库显示cd4结合能力降低,表明s118h和t157i突变都是关键的(图17d)。
[0445]
实际上,在β链中具有l114w/v143m+s118h/t157i替换(2reps)的dr1
l114w/v143m+2reps
的cd4结合能力与dr1
l114w/v143m+6reps
的cd4结合能力相当(图17e)。这些结果表明,s118h和t157i处的两个另外的替换对于l114w/v143m突变在改善dr1与cd4结合方面的功能至关重要。
[0446]
除了位置114和143之外,drβ链如drb1*03:01、04:01、07:01、10:01、11:01和13:01还编码在位置118、139、146、157、163和164处的不同的氨基酸(图17f)。有趣的是,比较dr1
l114w/v143m+2reps
和dr1
l114w/v143m+6reps
突变体之间的cd4结合活性表明,与dr1不同,对于dr3、dr4、dr7、dr10、dr11和dr13,l114w/v143m+2reps突变与l114w/v143m+6reps突变相比能够改善cd4结合(图17g至图17l)。
[0447]
使用生物层干涉(bli)结合测定,测量了野生型dr1和dr1
l114w/v143m+2reps
对cd4的亲和力。虽然在野生型dr1和cd4之间未检测到结合(图17m),但dr1
l114w/v143m+2reps
以14μm
±
2.3的kd与cd4结合(图17n至图17o)。
[0448]
实施例10
–
亲和力成熟的dr二聚体特异性和稳健地染色同源tcr
[0449]
评估了携带实施例2中描述的突变的亲和力成熟的dr二聚体鉴定抗原特异性cd4
+
t细胞的能力。dr1
l114w/v143m+2reps
、dr7
l114w/v143m+2reps
和dr11
l114w/v143m+2reps
二聚体分别特异性染色dr1限制性tcr ha1.7和sb95,dr7限制性tcr sd334,和dr11限制性tcr f24(图18a至图18c)。用抗vβ22 mab连同相应的dr11
l114w/v143m+2reps
二聚体对f24转导的cd4
+
t细胞进行复合染色,证实几乎所有tcr转导的cd4
+
t细胞都成功地用相应的dr11
l114w/v143m+2reps
二聚体染色(图18d)。
[0450]
由cd4和dr1
l114w/v143m+2reps
组成的复合物的结构模型也显示了l114w/v143m替换的潜在疏水作用(图19a至图19b)。此外,在α-链的p96和β-链的s118h之间观察到疏水堆积(图19c),并且发现t157i替换定位于v119、f112、i127、v129、l147和t157周围的β-折叠中(图
19d)。α链和/或β链的另外的突变可能会进一步增强ii类和cd4之间的结合。然而,使用这种具有过度cd4结合能力的可溶性ii类分子可能会导致cd4
+
t细胞的非特异性染色,从而具有不利影响。
[0451]
为了鉴定亲和力成熟的ii类分子,本实施例详细说明了β链而非α链中的多个突变,因为β链比α链具有更直接的与cd4的相互作用。α链和/或β链的另外的突变可能会进一步增强ii类和cd4之间的结合。然而,使用这种具有过度cd4结合能力的可溶性ii类分子可能会导致cd4
+
t细胞的非特异性染色,从而具有不利影响。
[0452]
总之,cd4
+
t细胞在自身免疫性疾病的发展以及对病原体感染和癌症的防护中发挥着关键作用。本文所述的新型hla ii类多聚体技术可以更好地促进对hla-dr等位基因的hla ii类限制性cd4
+
t细胞反应的研究。
[0453]
实施例11
[0454]
分析了内源性(未转导)抗原特异性cd4
+
t细胞的dp4多聚体染色。新型dp4
l112w/v141m
二聚体对内源性trpc1
578-597
特异性cd4
+
t细胞的阳性染色(图21a至图21b)比常规dp4 dextramer(图21c至图21d)更强。与常规四聚体(图22c至图22d)相比,dp4
l112w/v141m
二聚体显示出显著改善的内源性(未转导)ny-eso-1
157-170
特异性cd4
+
t细胞染色(图22a至图22b;表9)或dextramer染色(图22e至图22f)。
[0455]
表9:dp4限制性tcr
[0456]
[0457][0458]
接下来,在没有体外刺激的情况下,用对一系列病原体相关肽有特异性的dp4
l112w/v141m
二聚体对记忆cd4
+
t细胞进行离体染色。对于破伤风毒素
948-968
(tt
948-968
)、单纯疱疹病毒2型-ul21
283-302
(hsv-2-ul21
283-302
)和呼吸道合胞病毒糖蛋白
162-175
(rsv-gp
162-175
),cd4
+
t细胞的一个小亚群被dp4
l112w/v141m
二聚体阳性染色(图23a至图23y)。接下来,我们通过rsv-gp
162-175
(图24a至图24v)和tt
948-968二
聚体
+
cd4
+
t细胞的限制稀释来建立内源性(未转导)单细胞克隆(图25a至图25r)。这些t细胞克隆显示以抗原特异性方式产生il-2(图24w和图24s)。从dp4
l112w/v141m
rsv-gp和tt二聚体
+
单细胞克隆中分离出多个tcrαβ对,包括一个优势
对(表9)。在图24a至图24w和图25a至图25s中,通过rsv-gp
162-175
和tt
948-968
二聚体
+
细胞的限制稀释建立单细胞克隆。当将这些rsv-gp和tt二聚体
+
单细胞克隆分别用三种不同的dp4多聚体(dp4
l112w/v141m
二聚体、野生型dp4四聚体或野生型dp4 dextramer)染色时,dp4
l112w/v141m
二聚体比常规的野生型dp4 rsv-gp dextramer和野生型dp4 tt四聚体和dextramer(图26a至图26nn)显示出更好的rsv-gp-(c12和c39)和tt特异性克隆(c2和c9)染色。
[0459]
产生野生型dq5和dq5
l114w/v143m
二聚体(表10)和dq5
l114w/v143m+4reps
二聚体,并比较它们对tcr转导的cd4
+
t细胞的染色。野生型dq5二聚体不能检测到e6转导的cd4
+
t细胞。dq5
l114w/v143m
二聚体仅显示对e6转导的cd4
+
t细胞的弱染色,而相比之下,dq5
l114w/v143m+4reps
二聚体反而显示出稳健染色(图27a至图27l)。为了验证dq5
l114w/v143m+4reps
二聚体染色,我们从以肽特异性方式体外扩增的二聚体
+
cd4
+
t细胞中克隆了对gpc3
138-157
有特异性的dq5限制性tcr基因。当在人cd4
+
tcr缺陷型t细胞中进行克隆型重构时,tcr被同源dq5
l114w/v143m+4reps
二聚体成功染色,并以dq5限制性和抗原特异性方式发挥作用(图28a至图28g)。
[0460]
表10:tcr序列
[0461][0462]
在没有体外刺激的情况下,用对流感病毒血球凝集素(flu-ha)肽有特异性的dr1
l114w/v143m+2reps
二聚体对记忆cd4
+
t细胞进行离体染色。针对flu-ha
117-136-和flu-ha
306-318
,cd4
+
t细胞的一个小亚群被dr1
l114w/v143m+2reps
二聚体阳性染色(图29a至图29l)。产生野生型dr1、dr1l114
w/v143m
和dr1
l114w/v143m+6reps
二聚体和dr1
l114w/v143m+2reps
二聚体,并比较它们对tcr转导的cd4
+
t细胞的染色。野生型dr1和dr1
l114w/v143m
二聚体在cd4
+
t细胞上检测到非常少的同源tcr(ha1.7),而dr1
l114w/v143m+2reps
和dr1
l114w/v143m+6reps
二聚体显示出相似的稳健染色。重要的是,dr1
l114w/v143m+2reps
二聚体对ha1.7转导的cd4
+
t细胞的染色比野生型dr1 dextramer更稳健且分离更好(图30a至图30x)。为了验证dr1
l114w/v143m+2reps
二聚体染色,从以肽特异性方式体外扩增的二聚体
+
cd4
+
t细胞中克隆对hsd17b12
225-244
和ly6k
99-118
有特异性的dr1限制性tcr基因。当在原代cd4
+
t细胞中进行克隆型重构时,两种tcr(表11)都被同源dr1
l114w/v143m+2reps
二聚体成功染色,并以dr1限制和抗原特异性方式发挥作用(图31a至图31o)。
[0463]
表11:tcr序列
[0464][0465]
方法
[0466]
细胞
[0467]
通过密度梯度离心获得外周单核细胞。先前已经报道了分别表达作为单个hla等位基因的各种hla ii类基因连同cd80和cd83的基于k562的人工抗原呈递细胞(aapc)(参见
butler,m.o.等人,plos one 7,e30229(2012))。jurkat 76细胞系是一种t细胞白血病细胞系,缺乏内源性tcr、cd4和cd8表达(参见.heemskerk,m.h.等人,blood 102,3530-3540(2003))。jurkat 76/cd4细胞通过逆转录病毒转导人cd4基因生成。a375细胞是黑素瘤细胞系。使hek293t细胞和a375细胞在补充有10%fbs和50μg/ml庆大霉素的dmem中生长。将jurkat 76细胞系在补充有10%fbs和50μg/ml庆大霉素的rpmi 1640中培养。
[0468]
肽/抗体
[0469]
将合成肽以50mg/ml溶解在dmso中。将以下抗体用于流式细胞术分析:apc-cy7缀合的抗cd4(rpa-t4,biolegend,san diego,ca;参见wooldridge,l.等人,eur j immunol 36,1847-1855(2006))和pe缀合的抗his标签(ad1.1.10,abcam,cambridge,ma)。使用活/死可固定aqua死细胞染色试剂盒(live/dead fixable aqua dead cell stain kit)区分死细胞。用facscanto ii或lsrfortessa x-20分析染色细胞。使用facsaria ii进行细胞分选。使用flowjo软件(版本9.9.6)进行数据分析。
[0470]
基因
[0471]
通过cdna末端的5'-快速扩增(race)pcr克隆新的tcr基因,并如前所述进行测序(参见,例如,nakatsugawa,m.等人,sci rep 6,23821(2016);nakatsugawa,m.等人,j immunol 194,3487-3500(2015);ochi,t.等人,cancer immunol res 3,1070-1081(2015);每篇文献均通过引用以其整体并入本文)。将所有基因克隆到pmx逆转录病毒载体中,并使用基于293gpg和pg13细胞的逆转录病毒系统转导到细胞系中(参见,例如,hirano,n.等人,blood 107,1528-1536(2006);butler,mo等人,clin cancer res 13,1857-1867(2007);hirano,n.等人,clin cancer res 12,2967-2975(2006);每篇文献均通过引用以其整体并入本文)。
[0472]
hla ii类单体和二聚体的生成
[0473]
使用基于293gpg细胞的逆转录病毒系统用α和β基因转染hek293t细胞(参见hirano,n.等人,blood 107,1528-1536(2006);butler,m.o.等人,clin cancer res 13,1857-1867(2007);hirano,n.等人,blood 108,2662-2668(2006))并在补充有10%fbs和50μg/ml庆大霉素的dmem中培养。对于dp4二聚体染色,使稳定分泌可溶性dp4
l112w/v141m
蛋白的hek293t细胞生长至汇合,并将培养基更换为无血清293sfm ii培养基(thermo fisher scientific,waltham,ma)。
[0474]
使用基于293gpg细胞的逆转录病毒系统用α和β基因转染a375细胞(参见,例如,hirano,n.等人,blood 107,1528-1536(2006);butler,mo等人clin cancer res 13,1857-1867(2007);和hirano,n.等人,blood 108,2662-2668(2006);每篇文献均通过引用以其整体并入本文)并将其在补充有10%fbs和50μg/ml庆大霉素的dmem中培养。
[0475]
48小时后,收获条件培养物并使用amicon超滤器(截留分子量(mwco)10kda)(milliporesigma,burlington,ma)浓缩。然后在37℃下将含有可溶性hla ii类的上清液与100μg/ml目标肽混合20-24小时,以进行体外肽交换。使用镍涂覆板和抗his标签生物素化mab通过特异性elisa测量单体浓度。使用pe缀合的抗his mab以2:1的摩尔比在4℃下将可溶性hla ii类单体二聚化1.5小时以进行染色。
[0476]
dp4限制性抗原特异性cd4
+
t细胞的刺激
[0477]
将cd4
+
t细胞纯化并且用通过dp4限制性肽以10μg/ml冲击的表达dp4的aapc刺激,
并以20:1的e:t比率在200gy下辐照。48小时后,将10iu/ml il-2和10ng/ml il-15添加到cd4
+
t细胞中。每2-3天补足补充有il-2(10iu/ml)和il-15(10ng/ml)的培养基。刺激2周后,对t细胞进行dp4
l112w/v141m
二聚体染色。
[0478]
dq5限制性抗原特异性cd4
+
t细胞的刺激
[0479]
将cd4
+
t细胞纯化并且然后用通过gpc3
138-157
以10μg/ml冲击的表达dq5的aapc刺激,并以20:1的e:t比率在200gy下辐照。48小时后,将10iu/ml il-2和10ng/ml il-15添加到cd4
+
t细胞中。每2-3天补足补充有il-2(10iu/ml)和il-15(10ng/ml)的培养基。两周后,对t细胞进行dq5
l114w/v143m+4reps
二聚体染色。
[0480]
dr1限制性抗原特异性cd4
+
t细胞的刺激
[0481]
将cd4
+
t细胞纯化并且然后用通过dr1限制性肽以10μg/ml冲击的表达dr1的aapc刺激,并以20:1的e:t比率在200gy下辐照。48小时后,将10iu/ml il-2和10ng/ml il-15添加到cd4
+
t细胞中。每2-3天补足补充有il-2(10iu/ml)和il-15(10ng/ml)的培养基。两周后,对t细胞进行dr1
l114w/v143m+2reps
二聚体染色。
[0482]
hla ii类二聚体、四聚体和dextramer染色
[0483]
在多聚体染色分析中比较dp4四聚体和dextramer以及dr1dextramer。
[0484]
将用抗原特异性tcr基因转导的原代cd4
+
t细胞和jurkat76/cd4 t细胞在37℃下用50nm达沙替尼预处理30分钟,并在室温下用5-15μg/ml ii类二聚体染色4-5小时。洗涤后,用apc-cy7缀合的抗cd4 mab对细胞表面分子进行复染。
[0485]
来自黑素瘤患者的pbmc的未刺激cd4
+
t细胞的二聚体染色
[0486]
将100万个cd4
+
t细胞纯化并在37℃下用50nm达沙替尼预处理30分钟。将细胞在室温下用5-15μg/ml ii类二聚体染色4-5小时。洗涤后,用apc-cy7缀合的抗cd4 mab对细胞表面分子进行复染。通过流式细胞术确定二聚体
+
细胞的绝对计数。
[0487]
dp4二聚体
+
t细胞的扩增和单t细胞克隆的建立。
[0488]
为了扩增dp4
l112w/v141m
二聚体
+
t细胞,如上所述,将cd4
+
t细胞进行刺激并用dp4
l112w/v141m
二聚体染色。将二聚体
+
细胞通过使用抗pe磁珠进行分选,并通过使用以5-20:1的e:t比率在200gy下辐照的人工apc/mokt3进行扩增(参见butler,m.o.等人,plos one 7,e30229(2012))。每2-3天补足补充有il-2(10iu/ml)和il-15(10ng/ml)的培养基。两到三周后,对t细胞进行dp4
l112w/v141m
二聚体染色。如前所述,通过有限稀释产生dp4
l112w/v141m
二聚体
+
单细胞克隆(参见su,l.f.等人,immunity 38,373-383(2013))。简而言之,在没有达沙替尼预处理的情况下,使用dp4 l112w/v141m
二聚体纯化和染色记忆cd4
+
t细胞。对二聚体
+
细胞进行分选,然后在96孔板中用在20gy下辐照的来自多个同种异体供体的5μg/ml pha-p和pbmc刺激。用il-2(100iu/ml)和il-15(10ng/ml)刺激1周后补充和补足培养基。两周后,将单细胞克隆用dp4
l112w/v141m
二聚体染色。
[0489]
elispot测定
[0490]
如先前报道的,进行细胞因子elispot测定(参见例如yamashita,y.等人,nat commun 8,15244(2017);和anczurowski,m.等人,sci rep 8,4804(2018));每篇文献均通过引用以其整体并入本文)。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1