对灰翅夜蛾属的控制的制作方法
对灰翅夜蛾属的控制
1.序列表
2.提供ascii文本格式的序列表作为纸质副本的替代,该序列表是根据37c.f.r.
§
1.821提交的,名称为“81889-wo-reg-org-p-1_st25.txt”,大小为308千字节,于2020年5月31日生成并经由efs-web提交。这个序列表特此通过引用以其披露内容并入本说明书中。
技术领域
3.本发明涉及有害生物昆虫控制、蛋白质工程化和植物分子生物学领域。更具体地,本发明涉及使用txp40基因或txp40蛋白或使用txp40蛋白与第二有害生物控制剂组合来控制灰翅夜蛾(spodoptera)目中的有害生物昆虫,特别是草地贪夜蛾(spodoptera frugiperda)(秋粘虫;faw)的方法。本发明还涉及保护作物(特别是玉米)对抗灰翅夜蛾(特别是faw),更特别是对抗cry和/或vip3抗性faw的方法。本发明还提供了对faw具有杀昆虫活性的txp40蛋白毒素及其变体,该蛋白毒素及其变体的表达产生杀昆虫蛋白的核酸,以及制备这些杀昆虫蛋白和相应核酸的方法和使用这些杀昆虫蛋白和相应核酸来控制faw有害生物的方法。本发明还涉及植物,尤其是单子叶植物,特别是玉米植物,其可被faw侵染并且用可表达的txp40基因或用可表达的txp40基因和编码第二杀有害生物剂的基因转化。
背景技术:
4.灰翅夜蛾物种,包括草地贪夜蛾(秋粘虫,faw),是多种行播作物(包括玉米、棉花和大豆)的严重有害生物。faw是巴西和其他南美国家玉米植物(玉蜀黍(zea mays/maize))中最具破坏性的一致有害生物物种,导致产量减少高达57%。在某些年份,faw还会对包括美国在内的其他国家的玉米造成重大经济损失。该昆虫的幼虫最初以叶组织为食,然后进入轮生体深处,对发育中的雄穗造成严重损害。由faw引起的脱叶减少了光合面积,这可能使植物矮化并降低产量。faw幼虫还通过在穗侧面钻孔和以发育中的谷粒为食而损害穗,从而降低谷物质量和产量。
5.历史上,已经使用合成杀昆虫剂实现了对玉米中faw的控制。然而,这样的杀昆虫剂仅对幼虫有效,并且在它们深入到轮生体或穗之前有效。此外,在玉米密集栽培的地区,faw群体已经进化出对几种杀昆虫剂的抗性,如λ-三氯氟氰菊酯、毒死蜱、多杀菌素和氯芬奴隆。
6.其他faw控制选择最近包括使用表达一种或多种来自苏云金芽孢杆菌(bt)细菌的杀昆虫晶体蛋白(cry蛋白)和/或营养期杀昆虫蛋白(vip)的转基因玉米植物。迄今为止,来自bt的最有效的faw活性蛋白是营养期杀昆虫蛋白vip3。只有非常少的来自bt的cry蛋白对faw有活性。具有中度至高度活性的少数cry蛋白包括cry1ab、cry1be、cry1d、cry1f、cry1if、cry1j和cry2a。自大约2007年以来,表达这些cry蛋白和vip3蛋白中一种或多种的转基因玉米事件的商业发布为faw管理提供了新的策略。目前,将含有这些所说的bt事件的产品在faw为有害生物的大部分主要玉米种植区上栽培。例如,将这些产品在巴西的80多个玉米种植区(约12.5百万公顷/年)栽培。快速采用含有具有抗faw活性的bt杀昆虫蛋白的产
品有助于减少对抗faw的杀昆虫喷雾。然而,在一些情况下,bt衍生蛋白在玉米植物中的持续表达对目标faw有害生物群体造成了强烈的选择压力,有利于抗性的进化。例如,在巴西,faw的田间群体已经进化出对转基因玉米产品中表达的cry1f和cry1ab蛋白的抗性。faw还在波多黎各和美国地区发展了对cry1f玉米的抗性。在巴西,在faw群体中也报道了对具有包含cry1蛋白的叠加产物的产品的高频率抗性。相反,没有检测到对vip3a蛋白的抗性。
7.仍然需要鉴定使用杀昆虫剂控制行播作物的昆虫有害生物的新型和有效的方法,所述杀昆虫剂为农民提供经济利益并且是环境可接受的。特别需要的是使用与来自bt的现有杀昆虫蛋白(例如cry蛋白和/或vip3蛋白)具有不同作用模式的蛋白作为减轻抗性发展的方式来控制灰翅夜蛾属物种,特别是faw的方法。此外,通过这些使环境负担最小化的产品(如通过转基因植物)递送昆虫控制剂是最令人希望的。
技术实现要素:
8.鉴于这些需要,本发明提供了使用txp40杀昆虫蛋白和txp40杀昆虫蛋白的变体控制草地贪夜蛾(秋粘虫;faw)和其他昆虫有害生物的方法。本发明还提供了灰翅夜蛾活性txp40蛋白和与txp40基本相同的变体txp40蛋白。本发明的蛋白质令人惊讶地对faw具有口服毒性,所述faw是对许多类型的杀昆虫蛋白质,特别是对来自bt的cry蛋白质来说难以控制的昆虫有害生物物种。本发明还涉及编码txp40蛋白质或变体txp40蛋白质的重组密码子优化的多核苷酸。
9.本发明还包括含有本发明的重组多核苷酸的表达盒和载体;包括和能够表达这样的多核苷酸的植物或微生物;用这样的多核苷酸转化的植物,例如转基因玉米植物;这样的植物的子代,其含有稳定地掺入和以孟德尔方式遗传的多核苷酸,和/或这样的植物的种子和这样的子代。本发明还包括育种的方法,以将包含本发明的多核苷酸的转基因引入子代植物和各种玉米种质中。
10.本发明还包括含有txp40或其变体的组合物和配制品,这些组合物和配制品能够抑制faw存活、生长和/或繁殖的能力,或能够限制与faw有关的损害或作物植物的损失,例如将txp40或其变体作为组合物或配制品的一部分施用至faw侵染的区域或植物,或施用至预防性处理易感染faw的区域或植物以赋予针对faw有害生物的保护。
11.本发明进一步涉及制造txp40或其变体的方法,以及使用本发明的多核苷酸的方法,例如,在微生物中控制faw或者在转基因植物中赋予免于faw损害的保护。
12.通过学习本发明的以下说明书和非限制性实例,本发明的其他方面和优点对于本领域的技术人员而言将变得清楚。
13.序列表中的序列简述
14.附上的序列表中列出的核苷酸序列使用核苷酸碱基的标准字母缩写(如37c.f.r.
§
1.822中所定义)表示。列出的核苷酸和氨基酸序列定义了具有以所述方式排列的核苷酸和氨基酸单体的分子(即,分别是多核苷酸和多肽)。列出的核苷酸和氨基酸序列还各自定义了多核苷酸或多肽(其包括以所述方式排列的核苷酸和氨基酸单体)的属。考虑到遗传密码的冗余性,应理解包括编码序列的核苷酸序列也描述了与由参考序列组成的多核苷酸编码相同的多肽的多核苷酸的属。还将理解,氨基酸序列描述了编码该多肽的多核苷酸orf的属。
15.仅展示每个核酸序列的一条链,但通过对所呈现的链的任何提及,应理解为包括互补链。由于一级核苷酸序列的互补序列和反向互补序列必须由一级核酸序列披露,因此除非明确指出是其他情况(或者从出现序列的上下文中显然是其他情况),否则该互补序列和反向互补序列是指该核苷酸序列。此外,如本领域所理解的,rna链的核苷酸序列由转录其的dna序列决定(但是核苷酸碱基尿嘧啶(u)取代胸腺嘧啶(t)),rna序列通过编码其的dna序列的任何参考而被包括。在随附的序列表中:
16.seq id no:1是来自发光光状杆菌的txp40-1氨基酸序列。
17.seq id no:2是变体txp40-1氨基酸序列。
18.seq id no:3是来自发光光状杆菌的天然txp40-1核苷酸序列。
19.seq id no:4是大肠杆菌密码子优化的txp40-1核苷酸序列。
20.seq id no:5是玉蜀黍密码子优化的txp40-1核苷酸序列。
21.seq id no:6是来自发光杆菌属的h1txp40氨基酸序列。
22.seq id no:7是来自发光杆菌属的h2txp40氨基酸序列。
23.seq id no:8是来自发光杆菌属的h3txp40氨基酸序列。
24.seq id no:9是来自发光杆菌属的h5txp40氨基酸序列。
25.seq id no:10是来自嗜线虫致病杆菌的txp40氨基酸序列。
26.seq id no:11是大肠杆菌密码子优化的txp40-1 k31a变体核苷酸序列。
27.seq id no:12是大肠杆菌密码子优化的txp40-1 k48a变体核苷酸序列。
28.seq id no:13是大肠杆菌密码子优化的txp40-1 k49a变体核苷酸序列。
29.seq id no:14是大肠杆菌密码子优化的txp40-1 k73a变体核苷酸序列。
30.seq id no:15是大肠杆菌密码子优化的txp40-1 k75a变体核苷酸序列。
31.seq id no:16是大肠杆菌密码子优化的txp40-1 k103a变体核苷酸序列。
32.seq id no:17是大肠杆菌密码子优化的txp40-1 k111a变体核苷酸序列。
33.seq id no:18是大肠杆菌密码子优化的txp40-1 k119a变体核苷酸序列。
34.seq id no:19是大肠杆菌密码子优化的txp40-1 k133a变体核苷酸序列。
35.seq id no:20是大肠杆菌密码子优化的txp40-1 k143a变体核苷酸序列。
36.seq id no:21是大肠杆菌密码子优化的txp40-1 k170a变体核苷酸序列。
37.seq id no:22是大肠杆菌密码子优化的txp40-1 k191a变体核苷酸序列。
38.seq id no:23是大肠杆菌密码子优化的txp40-1 k200a变体核苷酸序列。
39.seq id no:24是大肠杆菌密码子优化的txp40-1 k209a变体核苷酸序列。
40.seq id no:25是大肠杆菌密码子优化的txp40-1 k210a变体核苷酸序列。
41.seq id no:26是大肠杆菌密码子优化的txp40-1 k213a变体核苷酸序列。
42.seq id no:27是大肠杆菌密码子优化的txp40-1 k221a变体核苷酸序列。
43.seq id no:28是大肠杆菌密码子优化的txp40-1 k238a变体核苷酸序列。
44.seq id no:29是大肠杆菌密码子优化的txp40-1 k247a变体核苷酸序列。
45.seq id no:30是大肠杆菌密码子优化的txp40-1 k250a变体核苷酸序列。
46.seq id no:31是大肠杆菌密码子优化的txp40-1 k263a变体核苷酸序列。
47.seq id no:32是大肠杆菌密码子优化的txp40-1 k265a变体核苷酸序列。
48.seq id no:33是大肠杆菌密码子优化的txp40-1 k271a变体核苷酸序列。
49.seq id no:34是大肠杆菌密码子优化的txp40-1 k275a变体核苷酸序列。
50.seq id no:35是大肠杆菌密码子优化的txp40-1 k284a变体核苷酸序列。
51.seq id no:36是大肠杆菌密码子优化的txp40-1 k296a变体核苷酸序列。
52.seq id no:37是大肠杆菌密码子优化的txp40-1 k309a变体核苷酸序列。
53.seq id no:38是大肠杆菌密码子优化的txp40-1 k333a变体核苷酸序列。
54.seq id no:39是大肠杆菌密码子优化的txp40-1 r10a变体核苷酸序列。
55.seq id no:40是大肠杆菌密码子优化的txp40-1 r26a变体核苷酸序列。
56.seq id no:41是大肠杆菌密码子优化的txp40-1 r46a变体核苷酸序列。
57.seq id no:42是大肠杆菌密码子优化的txp40-1 r167a变体核苷酸序列。
58.seq id no:43是大肠杆菌密码子优化的txp40-1 r186a变体核苷酸序列。
59.seq id no:44是大肠杆菌密码子优化的txp40-1 r187a变体核苷酸序列。
60.seq id no:45是大肠杆菌密码子优化的txp40-1 r208a变体核苷酸序列。
61.seq id no:46是大肠杆菌密码子优化的txp40-1 r217a变体核苷酸序列。
62.seq id no:47是大肠杆菌密码子优化的txp40-1 r226a变体核苷酸序列。
63.seq id no:48是大肠杆菌密码子优化的txp40-1 r240a变体核苷酸序列。
64.seq id no:49是大肠杆菌密码子优化的txp40-1 r252a变体核苷酸序列。
65.seq id no:50是大肠杆菌密码子优化的txp40-1 r305a变体核苷酸序列。
66.seq id no:51是大肠杆菌密码子优化的txp40-1 r311a变体核苷酸序列。
67.seq id no:52是大肠杆菌密码子优化的txp40-1 k31a/k49a变体核苷酸序列。
68.seq id no:53是大肠杆菌密码子优化的txp40-1 k31a/k333a变体核苷酸序列。
69.seq id no:54是大肠杆菌密码子优化的txp40-1 k31a/r208a变体核苷酸序列。
70.seq id no:55是大肠杆菌密码子优化的txp40-1 r208a/k333a变体核苷酸序列。
71.seq id no:56是大肠杆菌密码子优化的txp40-1 k49a/r208a变体核苷酸序列。
72.seq id no:57是大肠杆菌密码子优化的txp40-1 k49a/k333a变体核苷酸序列。
73.seq id no:58是大肠杆菌密码子优化的txp40-1 k31a/k49a/k333a变体核苷酸序列。
74.seq id no:59是大肠杆菌密码子优化的txp40-1 k31a/k49a/r208a/k333a变体核苷酸序列。
75.seq id no:60是大肠杆菌密码子优化的txp40-1 k73a/k213a变体核苷酸序列。
76.seq id no:61是大肠杆菌密码子优化的txp40-1 k103a/k213a变体核苷酸序列。
77.seq id no:62是大肠杆菌密码子优化的txp40-1 k213a/k284a变体核苷酸序列。
78.seq id no:63是大肠杆菌密码子优化的txp40-1 k31a/k49a/k213a变体核苷酸序列。
79.seq id no:64是大肠杆菌密码子优化的txp40-1 k119a/k213a变体核苷酸序列。
80.seq id no:65是大肠杆菌密码子优化的txp40-1 k213a/r167a变体核苷酸序列。
81.seq id no:66是大肠杆菌密码子优化的txp40-1 k119a/r167a变体核苷酸序列。
82.seq id no:67是大肠杆菌密码子优化的txp40-1 s11y/m86i/k119a/k213t变体核苷酸序列。
83.seq id no:68是大肠杆菌密码子优化的txp40-1 s11y/k119a/k213a变体核苷酸
序列。
84.seq id no:69是大肠杆菌密码子优化的txp40-1 k119a/f169v/k213a变体核苷酸序列。
85.seq id no:70是大肠杆菌密码子优化的txp40-1 k119a/p134l/k213a变体核苷酸序列。
86.seq id no:71是大肠杆菌密码子优化的txp40-1 i101l/k103t/k119a/k213a/n257t变体核苷酸序列。
87.seq id no:72是大肠杆菌密码子优化的txp40-1 h62q/d99v/k119a/t165i/r167c/k213a变体核苷酸序列。
88.seq id no:73是大肠杆菌密码子优化的txp40-1 k119a/k213a/l316v变体核苷酸序列。
89.seq id no:74是txp40-1 k31a变体氨基酸序列。
90.seq id no:75是txp40-1 k48a变体氨基酸序列。
91.seq id no:76是txp40-1 k49a变体氨基酸序列。
92.seq id no:77是txp40-1 k73a变体氨基酸序列。
93.seq id no:78是txp40-1 k75a变体氨基酸序列。
94.seq id no:79是txp40-1 k103a变体氨基酸序列。
95.seq id no:80是txp40-1 k111a变体氨基酸序列。
96.seq id no:81是txp40-1 k119a变体氨基酸序列。
97.seq id no:82是txp40-1 k133a变体氨基酸序列。
98.seq id no:83是txp40-1 k143a变体核苷酸序列。
99.seq id no:84是txp40-1 k170a变体氨基酸序列。
100.seq id no:85是txp40-1 k191a变体氨基酸序列。
101.seq id no:86是txp40-1 k200a变体氨基酸序列。
102.seq id no:87是txp40-1 k209a变体氨基酸序列。
103.seq id no:88是txp40-1 k210a变体氨基酸序列。
104.seq id no:89是txp40-1 k213a变体氨基酸序列。
105.seq id no:90是txp40-1 k221a变体氨基酸序列。
106.seq id no:91是txp40-1 k238a变体氨基酸序列。
107.seq id no:92是txp40-1 k247a变体氨基酸序列。
108.seq id no:93是txp40-1 k250a变体氨基酸序列。
109.seq id no:94是txp40-1 k263a变体氨基酸序列。
110.seq id no:95是txp40-1 k265a变体氨基酸序列。
111.seq id no:96是txp40-1 k271a变体氨基酸序列。
112.seq id no:97是txp40-1 k275a变体氨基酸序列。
113.seq id no:98是txp40-1 k284a变体氨基酸序列。
114.seq id no:99是txp40-1 k296a变体氨基酸序列。
115.seq id no:100是txp40-1 k309a变体氨基酸序列。
116.seq id no:101是txp40-1 k333a变体氨基酸序列。
117.seq id no:102是txp40-1 r10a变体氨基酸序列。
118.seq id no:103是txp40-1 r26a变体氨基酸序列。
119.seq id no:104是txp40-1 r46a变体氨基酸序列。
120.seq id no:105是txp40-1 r167a变体氨基酸序列。
121.seq id no:106是txp40-1 r186a变体氨基酸序列。
122.seq id no:107是txp40-1 r187a变体氨基酸序列。
123.seq id no:108是txp40-1 r208a变体氨基酸序列。
124.seq id no:109是txp40-1 r217a变体氨基酸序列。
125.seq id no:110是txp40-1 r226a变体氨基酸序列。
126.seq id no:111是txp40-1 r240a变体氨基酸序列。
127.seq id no:112是txp40-1 r252a变体氨基酸序列。
128.seq id no:113是txp40-1 r305a变体氨基酸序列。
129.seq id no:114是txp40-1 r311a变体氨基酸序列。
130.seq id no:115是txp40-1 k31a/k49a变体氨基酸序列。
131.seq id no:116是txp40-1 k31a/k333a变体氨基酸序列。
132.seq id no:117是txp40-1 k31a/r208a变体氨基酸序列。
133.seq id no:118是txp40-1 r208a/k333a变体氨基酸序列。
134.seq id no:119是txp40-1 k49a/r208a变体氨基酸序列。
135.seq id no:120是txp40-1 k49a/k333a变体氨基酸序列。
136.seq id no:121是txp40-1 k31a/k49a/k333a变体氨基酸序列。
137.seq id no:122是txp40-1 k31a/k49a/r208a/k333a变体氨基酸序列。
138.seq id no:123是txp40-1 k73a/k213a变体氨基酸序列。
139.seq id no:124是txp40-1 k103a/k213a变体氨基酸序列。
140.seq id no:125是txp40-1 k213a/k284a变体氨基酸序列。
141.seq id no:126是txp40-1 k31a/k49a/k213a变体氨基酸序列。
142.seq id no:127是txp40-1 k119a/k213a变体氨基酸序列。
143.seq id no:128是txp40-1 k213a/r167a变体氨基酸序列。
144.seq id no:129是txp40-1 k119a/r167a变体氨基酸序列。
145.seq id no:130是txp40-1 s11y/m86i/k119a/k213t变体氨基酸序列。
146.seq id no:131是txp40-1 s11y/k119a/k213a变体氨基酸序列。
147.seq id no:132是txp40-1 k119a/f169v/k213a变体氨基酸序列。
148.seq id no:133是txp40-1 k119/p134l/k213a变体氨基酸序列。
149.seq id no:134是txp40-1 i101l/k103t/k119a/k213a/n257t变体氨基酸序列。
150.seq id no:135是txp40-1 h62q/d99v/k119a/t165i/r167c/k213a变体氨基酸序列。
151.seq id no:136是k119a/k213a/l316v变体氨基酸序列。
具体实施方式
152.本说明不旨在是可以实施本发明的所有不同方式或可以添加到本发明中的所有
特征的详细目录。例如,关于一个实施例所说明的特征可以并入其他实施例中,并且关于一个特定实施例所说明的特征可以从那个实施例删除。因此,本发明考虑了,在本发明的一些实施例中,可以排除或省略本文陈述的任何特征或特征的组合。此外,鉴于本披露内容,本文提出的不同实施例的众多变化以及附加对于本领域技术人员将是显而易见的,这不脱离本发明。因此,以下说明旨在阐述本发明的一些特定实施例,并且并没有穷尽地叙述其所有排列、组合和变化。
153.除非另外定义,本文所用的所有技术和科学术语均具有与本发明所属领域的普通技术人员通常所理解的相同的含义。在本文的发明的说明中使用的术语是仅出于描述特定实施例的目的,且并不旨在限制本发明。
154.本文引用的所有的公开、专利申请、专利以及其他参考文件对于引用中提及的有关句子和/或段落的传授内容通过引用以其全文并入。
155.在此提供的核苷酸序列以5'至3'方向从左至右表示,并且使用代表核苷酸碱基的标准代码表示,如37cfr
§§
1.821-1.825和世界知识产权组织(wipo)标准st.25中所述,例如:腺嘌呤(a)、胞嘧啶(c)、胸腺嘧啶(t)、以及鸟嘌呤(g)。
156.氨基酸同样是使用wipo标准st.25来指示,例如:丙氨酸(ala;a)、精氨酸(arg;r)、天冬酰胺(asn;n)、天冬氨酸(asp;d)、半胱氨酸(cys;c)、谷氨酰胺(gln;q)、谷氨酸(glu;e)、甘氨酸(gly;g)、组氨酸(his;h)、异亮氨酸(ile;1)、亮氨酸(leu;l)、赖氨酸(lys;k)、甲硫氨酸(met;m)、苯丙氨酸(phe;f)、脯氨酸(pro;p)、丝氨酸(ser;s)、苏氨酸(thr;t)、色氨酸(trp;w)、酪氨酸(tyr;y)、以及缬氨酸(val;v)。
157.除非上下文另外指示,明确地预期的是本文所述的本发明的不同特征可以按任何组合使用。而且,本发明还考虑到在本发明的一些实施例中,本文陈述的任何特征或特征的组合可以被排除或省略。举例说明,如果本说明书陈述组合物包含组分a、b和c,明确地预期a、b或c的任何一种或其组合可单一地或以任何组合被省略和放弃。
158.定义
159.为了清晰起见,定义了在本说明书中所使用的某些术语并且将其呈现如下:
160.如在本发明的说明书和所附的权利要求中所使用的,单数形式“一个”、“一种”和“所述(the)”旨在也包括复数形式,除非上下文清楚地另外指示。
161.如在此使用的,“和/或”是指并且涵盖一个或多个相关的列出项的任何及全部可能组合,连同当以可替代性(“或”)解释时组合的缺少。
162.如在此使用的,术语“约”当是指一个可测量的值如剂量或时间段等时意在涵盖
±
20、
±
10、
±
5、
±
1、
±
0.5、或甚至
±
0.1的指定量的变化。如在此使用的,短语如“在约x和y之间”意指“在约x和约y之间”,并且短语如“从约x至y”意指“从约x至约y”。
163.如在此使用的,短语如“从x至y”应解释为包括x和y,除非上下文另外指示。
164.本发明的杀昆虫蛋白的“活性”意指杀昆虫蛋白作为口服活性的昆虫控制剂发挥作用,具有毒性作用、和/或能够干扰或阻止昆虫摄食,这可能引起或者可能不引起昆虫的死亡。当本发明的杀昆虫蛋白被递送至昆虫时,这种结果典型地是该昆虫的死亡,或者该昆虫不以该杀昆虫蛋白可被该昆虫可用的来源为食。“杀有害生物”被定义为有毒的生物活性,其能够控制有害生物(如昆虫、线虫、真菌、细菌或病毒),优选地通过杀死或破坏它们来进行控制。“杀昆虫”被定义为有毒的生物活性,其能够控制昆虫,优选地通过杀死它们来进
行控制。“杀有害生物剂”是具有杀有害生物活性的药剂。“杀昆虫剂”是具有杀昆虫活性的药剂。
[0165]“与
……
相关联/可操作地连接”指物理上或功能上相关的两种核酸。例如,启动子或调节dna序列被说成“与”对rna或蛋白质进行编码的dna序列“相关联”,条件是将这两条序列可操作地连接,或设定为使该调节性dna序列将影响该编码或结构dna序列的表达水平。
[0166]
如本文所使用的术语“嵌合构建体”或“嵌合基因”或“嵌合多核苷酸”或“嵌合核酸”(或类似术语)是指如下构建体或分子,该构建体或分子包含被组装进单个核酸分子中的不同来源的两个或更多个多核苷酸。术语“嵌合构建体”、“嵌合基因”、“嵌合多核苷酸”或“嵌合核酸”是指如下任何构建体或分子,该构建体或分子含有但不限于(1)多核苷酸(例如,dna),包括在自然界中没有被发现在一起的调节多核苷酸和编码多核苷酸(即,构建体中的至少一个多核苷酸相对于它的其他多核苷酸中的至少一个是异源的),或(2)编码不是天然毗邻的蛋白部分的多核苷酸,或(3)不是天然毗邻的启动子部分。另外,嵌合构建体、嵌合基因、嵌合多核苷酸或嵌合核酸可以包含衍生自不同来源的调节多核苷酸和编码多核苷酸,或包含衍生自相同来源、但以与在自然界中所发现的不同的方式进行布置的调节多核苷酸和编码多核苷酸。在本发明的一些实施例中,嵌合构建体、嵌合基因、嵌合多核苷酸或嵌合核酸包含表达盒,该表达盒包含在调节多核苷酸的控制下、特别地在植物或细菌中具有功能性的调节多核苷酸的控制下的本发明的多核苷酸。
[0167]“编码序列”是转录成rna(如mrna、rrna、trna、snrna、正义rna或反义rna)的核酸序列。优选地,rna进而在生物中被翻译以产生蛋白质。
[0168]
如本文所使用的,“密码子优化的”序列意指如下核苷酸序列,其中这些密码子被选择以反映宿主细胞或生物可以具有的特定的密码子偏好性。这典型地是以这样一种方式来完成,该方式是为了保持由待优化的核苷酸序列所编码的多肽的氨基酸序列。在某些实施例中,重组dna构建体的dna序列包括已经针对该构建体有待在其中进行表达的细胞(例如,动物、植物、或真菌细胞)进行了密码子优化的序列。例如,有待在植物细胞中表达的构建体可以使其全部或部分序列(例如,第一基因抑制元件或基因表达元件)进行密码子优化用于在植物中表达。参见例如,美国专利号6,121,014,通过引用并入本文。
[0169]“控制”昆虫意指通过毒性作用抑制了昆虫有害生物存活、生长、摄食、和/或繁殖的能力,或限制与昆虫有关的损害或作物植物中的损失。“控制”昆虫可以是或可以不是意指杀死昆虫,尽管其优选意指杀死昆虫。
[0170]
术语“包含(comprises或comprising)”当用于本说明书中时指示所说明的特征、整数、步骤、操作、要素、或组分的存在,但并不排除一个或多个其他特征、整数、步骤、操作、要素、组分、或其组的存在或添加。
[0171]
如本文所使用的,过渡短语“基本上由
……
组成”(以及语法变体)意指,权利要求书的范围有待被解读为涵盖权利要求书中所列举的指定材料或步骤以及不实质上改变所要求的发明的一个或多个基本和新颖特征的那些。因此,当用于本发明的权利要求中时,术语“基本上由
……
组成”并不旨在被解释为等同于“包含(comprising)”。
[0172]
在本发明的上下文中,“对应于(corresponding to或corresponds to)”意指当变体或同系物cry蛋白的氨基酸序列与彼此比对时,“对应于”在该变体或同系物蛋白中某些
枚举的位置的氨基酸是与参考蛋白中的这些位置比对的那些,但在相对于本发明的特定参考氨基酸序列而言的这些精确的数字位置中是不必要的。例如,如果seq id no:1是参考序列并且与seq id no:10比对的话,seq id no:10的gly51“对应于”seq id no:1的gly42,或者例如,seq id no:10的val59“对应于”seq id no:1的ile50。
[0173]
如在此使用的,术语“cry蛋白”意指苏云金芽孢杆菌晶体δ-内毒素类型的杀昆虫蛋白。术语“cry蛋白”可以指原毒素形式或其任何生物活性片段或毒素,包括部分加工的和成熟的毒素形式,例如,没有n-末端肽基片段和/或c-末端原毒素尾部。
[0174]“递送”杀昆虫蛋白意指该杀昆虫蛋白与昆虫相接触,产生毒性作用以及对昆虫的控制。这种杀昆虫蛋白可以按照许多公认的方式进行递送,例如,通过表达该杀昆虫蛋白的转基因植物、一种或多种配制的蛋白组合物、一种或多种可喷洒的蛋白组合物、诱饵基质、或领域公认的任何其他的毒素递送系统。
[0175]
术语“结构域”是指沿着进化相关蛋白的序列的比对在特定位置处保守的一组氨基酸。虽然其他位置上的氨基酸可在同系物之间有所不同,但是在特定位置处高度保守的氨基酸指示在蛋白质的结构、稳定性或功能中很可能是必需的氨基酸。通过其在蛋白质同系物家族的经比对序列中的高度保守性进行鉴别,其可用作鉴别物(identifier),用来确定所讨论的任何多肽是否属于先前鉴别的多肽组。
[0176]“控昆虫有效量”意指杀昆虫蛋白的浓度,其通过毒性作用抑制昆虫存活、生长、摄食和/或繁殖的能力,或限制昆虫相关的损害或作物植物损失。“控昆虫有效量”可以是或可以不是意指杀死昆虫,尽管其优选意指杀死昆虫。
[0177]
如本文所使用的“表达盒”意指能够在适当的宿主细胞中指导特定的核苷酸序列的表达的核酸序列,包含可操作地连接至目的核苷酸序列的启动子,该目的核苷酸序列可操作地连接至终止信号。它还典型地包含适当翻译该核苷酸序列所需要的序列。包含该目的核苷酸序列的表达盒可能具有其组分中的至少一种,该组分相对于它的其他组分中的至少一种而言是异源的。该表达盒还可以是天然存在的但已经是以对于异源表达有用的重组形式而获得的表达盒。然而,典型地,该表达盒相对于该宿主而言是异源的,即该表达盒的特定核酸序列不是天然存在于该宿主细胞中的,并且必须已经通过转化事件引入到该宿主细胞或该宿主细胞的祖先中。在表达盒中核苷酸序列的表达可以是在组成型启动子或诱导型启动子的控制之下,该启动子只有当该宿主细胞暴露于一些特定的外界刺激时才引发转录。在多细胞生物体(例如植物)的情况下,该启动子对于特定组织、或器官、或者发育阶段也可以是特异的。
[0178]
包含目的核苷酸序列的表达盒可以是嵌合的,这意味着它的组分中的至少一种相对于它的其他组分中的至少一种而言是异源的。表达盒也可以是包含驱动其天然基因的天然启动子的表达盒;然而,它已经以可用于异源表达的重组形式获得。表达盒的这种使用意味着表达盒不是天然存在于其被引入的细胞中。
[0179]
表达盒还可以任选地包括在植物中发挥作用的转录和/或翻译终止区(即,终止区)。多种转录终止子是可供用于在表达盒中使用的并且负责在超出目的异源核苷酸序列时的转录终止以及正确的mrna聚腺苷酸化。终止区对于转录起始区可以是天然的,对于可操作地连接的目的核苷酸序列可以是天然的,对于植物宿主可以是天然的,或者可以是衍生自另一种来源(即,对于启动子、目的核苷酸序列、植物宿主、或其任何组合而言是外来的
或异源的)。适当的转录终止子包括但不限于camv 35s终止子、tml终止子、胭脂碱合酶终止子和/或豌豆rbcs e9终止子。这些终止子可以在单子叶植物和双子叶植物两者中使用。此外,可以使用编码序列的天然转录终止子。任何已知在植物中发挥作用的可供使用的终止子均可以在本发明的上下文中使用。
[0180]
当参考多核苷酸(如植物的基因、orf或其部分、或转基因)使用时,术语“表达”是指通过基因的“转录”(即经由rna聚合酶的酶促作用)将基因中编码的遗传信息转化为rna(例如,mrna、rrna、trna或snrna),并在适用的情况下(例如,如果基因编码蛋白)通过mrna的“翻译”转化为蛋白的过程。基因表达可以在该过程的许多阶段进行调节。例如,在反义构建体或dsrna构建体的情况下,各自地表达可仅指该反义rna或仅指dsrna的转录。在实施例中,“表达”是指正义(mrna)或功能性rna的转录和稳定累积。“表达”还可指蛋白的产生。
[0181]“基因”是位于基因组内并且包含编码核酸序列的限定区域,并且典型地还包含其他负责控制该编码部分表达(也就是转录和翻译)的主要调节性核酸。基因还可以包含其他5'和3'未翻译序列和终止序列。另外的可能存在的元件是例如内含子。如在自然界中所发现,基因的调节核酸序列在正常情况下可能不与该相关联的核酸序列进行可操作地连接,并因此不会是嵌合基因。
[0182]“目的基因”是指当转移至植物时,在该植物上赋予所希望的性状(如抗生素抗性、病毒抗性、昆虫抗性、疾病抗性、或对其他有害生物的抗性、除草剂耐受性、非生物胁迫耐受性、雄性不育、改性脂肪酸代谢、改性碳水化合物代谢、改善的营养价值、工业过程中改善的性能或改变的繁殖能力)的任何核酸分子。“目的基因”还可以是被转移至植物用于在该植物中产生商业上有价值的酶或代谢物的基因。
[0183]“肠蛋白酶”是在昆虫的消化道中天然发现的蛋白酶。这种蛋白酶通常参与被摄取的蛋白质的消化。肠蛋白质酶的实例包括胰蛋白酶,其典型地切割赖氨酸(k)或精氨酸(r)残基的c-末端侧上的肽;以及胰凝乳蛋白酶,其典型地切割苯丙氨酸(f)、色氨酸(w)或酪氨酸(y)的c-末端侧上的肽。
[0184]“异源”核酸序列或核酸分子是天然地不与将该核酸序列引入其中的宿主细胞相关联的核酸序列或核酸分子,包括天然存在的核酸序列的非天然存在的多个拷贝。异源核酸序列或核酸分子可以包含嵌合序列,如嵌合表达盒,在该表达盒中,启动子和编码区源自多源生物。启动子序列可以是组成型启动子序列、组织特异性启动子序列、化学诱导型启动子序列、伤口诱导型启动子序列、胁迫诱导型启动子序列、或发育阶段特异性启动子序列。
[0185]“同源”核酸序列是与其被引入的宿主细胞天然相关联的核酸序列。
[0186]“同源重组”是在同源核酸分子之间的核酸片段的相互交换。
[0187]“同一性”或“同一性百分比”是指两个核苷酸或氨基酸序列之间的相似程度。对于序列比较,典型地,一个序列充当与测试序列进行比较的参考序列。当使用序列比较算法时,将测试序列和参考序列输入到计算机中(如有必要,指定子序列坐标),并且指定序列算法程序的参数。然后,该序列比较算法基于所指定的程序参数来计算一种或多种测试序列相对于该参考序列的序列同一性百分比。在两个核苷酸或两个氨基酸序列的上下文中,短语“基本上同一的”是指当针对最大对应性进行比较和比对时具有至少约50核苷酸或氨基酸残基同一性(如使用以下序列比较算法之一或通过目测检查所测量的)的两个或更多个序列或子序列。在某些实施例中,基本上相同的序列具有至少约60、或至少约70、或至少约
80、或至少约85、或甚至至少约90或95核苷酸或氨基酸残基同一性。在某些实施例中,基本上的同一性存在于这些序列的整个长度为至少约50个残基的区域中,或在整个至少约100个残基的区域中,或这些序列在至少约150个残基中是基本上相同的。在另外的实施例中,当这些序列在编码区的整个长度上时,这些序列是基本上同一的。
[0188]
用于比较的序列的最佳比对可以按照以下方式进行,例如通过smith和waterman,adv.appl.math.[应用数学进展]2:482(1981)的局部同源性算法、通过needleman和wunsch,j.mol.biol.[分子生物学杂志]48:443(1970)的同源比对算法、通过pearson和lipman,proc.nat'l.acad sci.usa[美国国家科学院院刊]85:2444(1988)的相似性方法的搜索,通过这些算法的计算机化实施(威斯康星州遗传学分析软件包中的gap、bestfit、fasta和tfasta,遗传学计算机组(genetics computer group),科学街575号(575science dr.),麦迪逊,威斯康星州)、或通过目测检查(总体上参见ausubel等人,下文)。
[0189]
适合于确定序列同一性百分比以及序列相似性的算法的一个实例是blast算法,其描述于以下文献中:altschul等人,j.mol.biol.[分子生物学杂志]215:403-410(1990)。执行blast分析的软件可通过美国国家生物技术信息中心(national center for biotechnology information)公开地获得(http://www.ncbi.nlm.nih.gov/)。这种算法涉及首先通过鉴定查询序列中具有长度w的短字码而鉴定得分高的序列对(hsp),这些得分高的序列对当与数据库序列中具有相同长度的字码(word)进行比对时匹配或满足一些正值阈值的得分t。t被称为邻近字码得分阈值(altschul等人,1990)。这些初始的邻近字码命中充当种子用于起始搜索以发现含有它们的较长的hsp。然后,将这些字码命中在两个方向上沿着每个序列延伸直到累积的比对得分可以增加。对于核苷酸序列,使用参数m(对于一对匹配残基的奖赏得分;总是》0)和n(对于错配残基的罚分;总是《0)来计算累积得分。对于氨基酸序列,使用得分矩阵来计算累积得分。当累积的比对得分从它的最大达到值降低了数量x;由于累积一个或多个负得分的残基比对使累积得分趋于0或0以下;或者到达任一序列的末端时,停止这些字码命中在每个方向上的延伸。blast算法的参数w、t、以及x决定了比对的灵敏度与速度。blastn程序(对核苷酸序列来说)使用字长(w)为11、期望值(e)为10、截止值(cutoff)为100、m=5、n=-4、以及两条链的比较作为默认值。对于氨基酸序列,blastp程序使用字长(w)为3、期望值(e)为10、以及blosum62评分矩阵作为默认值(参见henikoff&henikoff,proc.natl.acad.sci.usa[美国国家科学院院刊]89:10915(1989))。
[0190]
除了计算序列同一性百分比之外,blast算法还进行两个序列之间相似性的统计分析(参见,例如karlin和altschul,proc.nat'l.acad.sci.usa[美国国家科学院院刊]90:5873-5787(1993))。由blast算法提供的相似性的一种量度是最小概率总和(p(n)),它提供了在两个核苷酸或氨基酸序列之间会偶然发生匹配的概率的指示。例如,若在测试核酸序列与参考核酸序列的比较中最小概率总和小于约0.1、更优选地小于约0.01、并且最优选地小于约0.001,则该测试核酸序列被认为是与该参考序列相似的。
[0191]
用于进行序列比对的另一种被广泛使用和接受的计算机程序是clustalw v1.6(thompson,等人nuc.acids res.[核酸研究],22:4673-4680,1994)。将匹配的碱基或氨基酸的数目除以碱基或氨基酸的总数目并乘以100,以获得百分比同一性。例如,如果两个580个碱基对序列具有145个匹配的碱基,则它们会是25%同一的。如果两个进行比较的序列具有不同的长度,则将匹配的数目除以这两个长度中较短的一个。例如,如果在200个氨基酸
的蛋白质与400个氨基酸的蛋白质之间存在100个匹配的氨基酸,则相对于较短的序列而言这两个蛋白质是50%同一的。如果所述较短的序列在长度上小于150个碱基或50个氨基酸,则将匹配的数目除以150(对于核酸碱基而言)或50(对于氨基酸而言)并乘以100,以获得百分比同一性。
[0192]
两个核酸基本上同一的另一个指标是这两个分子在严格条件下彼此进行杂交。短语“特异性杂交”是指分子在严格条件下仅与特定的核苷酸序列结合、双链化或杂交,这是在该序列存在于dna或rna复合混合物(例如,总细胞的)中时进行的。“基本上结合”是指在探针核酸与靶核酸之间的互补杂交,并且涵盖少量错配,这些错配可以通过降低杂交介质的严格来容纳,以实现靶核酸序列的所希望的检测。
[0193]
在核酸杂交实验(如dna杂交和rna杂交)的上下文中,“严格杂交条件”和“严格杂交洗涤条件”是序列依赖性的,并且在不同的环境参数下是不同的。较长的序列在较高的温度下特异性杂交。对核酸杂交的广泛指导见于以下文献中:tijssen(1993)laboratory techniques in biochemistry and molecular biology-hybridization with nucleic acid probes[生物化学和分子生物学实验室技术-使用核酸探针的杂交]第2章第i部分“overview of principles of hybridization and the strategy of nucleic acid probe assays[杂交原理和核酸探针检验策略综述]”elsevier[爱思唯尔集团],纽约。通常,高严格杂交和洗涤条件在限定的离子强度和ph下被选定为比特定序列的热熔点(tm)低约5℃。典型地,在“严格条件”下,探针将会与其靶子序列进行杂交,但不会与其他序列杂交。
[0194]
tm是50的靶序列与完全匹配的探针进行杂交时的温度(在限定的离子强度和ph下)。非常严格条件被选定为等于特定探针的tm。针对互补核酸(它们在dna或rna印迹中在滤纸上具有超过100个互补的残基)的杂交的严格杂交条件的一个实例是具有1mg肝素的50甲酰胺、在42℃、将该杂交过夜进行。高严格洗涤条件的一个实例是0.15m nacl,在72℃持续约15分钟。严格洗涤条件的实例是在65℃以0.2x ssc洗涤持续15分钟(参见sambrook,下文,针对ssc缓冲液的描述)。通常,高严格洗涤之前会先进行低严格洗涤,以去除背景探针信号。对于例如多于100个核苷酸的双链体的中严格洗涤的实例是在45℃以1x ssc持续15分钟。对于例如多于100个核苷酸的双链体的低严格洗涤的实例是在40℃以4-6x ssc持续15分钟。对于短探针(例如,约10至50个核苷酸),严格条件典型地涉及小于约1.0m的na离子的盐浓度,典型地在ph 7.0至8.3下约0.01至1.0m的na离子浓度(或其他盐),并且温度典型地是至少约30℃。还可以通过添加去稳定剂(如甲酰胺)来达到严格条件。一般而言,在特定的杂交测定中相比于不相关的探针观察到的高出2x(或更高)的信噪比表明检测到特异性杂交。如果在严格条件下彼此不杂交的核酸所编码的蛋白是基本上相同的,则它们仍然是基本上相同的。例如,当使用遗传密码允许的最大程度的密码子简并而创建核酸的拷贝时,则发生这种情况。
[0195]
以下是可以用来克隆同源核苷酸序列(这些序列与本发明的参考核苷酸序列基本上相同)的杂交/洗涤条件的设置的实例:参考核苷酸序列在以下条件下优选地与该参考核苷酸序列杂交:在7十二烷基硫酸钠(sds)、0.5m napo4、1mm edta中在50℃,并且在2x ssc、0.1sds中在50℃洗涤;更令人希望的是在7十二烷基硫酸钠(sds)、0.5m napo4、1mm edta中在50℃,并且在1x ssc、0.1sds中在50℃洗涤;仍更令人希望的是在7十二烷基硫酸钠
(sds)、0.5mnapo4、1mm edta中在50℃,并且在0.5x ssc、0.1sds中在50℃洗涤;优选地在7十二烷基硫酸钠(sds)、0.5m napo4、1mm edta中在50℃,并且在0.1x ssc、0.1sds中在50℃洗涤;更优选地在7十二烷基硫酸钠(sds)、0.5m napo4、1mm edta中在50℃,并且在0.1x ssc、0.1sds中在65℃洗涤。
[0196]
两个核酸或蛋白质是基本上同一的另一个指标是由第一核酸所编码的蛋白质与由第二核酸所编码的蛋白质具有免疫交叉反应性或与其特异性结合。因此,蛋白质典型地是与第二蛋白质基本上一致的,例如其中这两种蛋白质仅区别于保守性取代。
[0197]
当核苷酸序列编码了多肽(该多肽与由参考核苷酸序列所编码的多肽具有相同的氨基酸序列)时,这种核苷酸序列与这种参考核苷酸序列“同类编码”。
[0198]“分离的”核酸分子、或分离的多核苷酸或分离的txp40蛋白是借助于人的手脱离其天然环境存在的核酸分子、或多核苷酸或蛋白,并且因此不是天然产物。分离的核酸分子、或多核苷酸或蛋白可能以纯化的形式存在或可能存在于非天然的环境中,如例如没有限制,存在于重组的微生物细胞、植物细胞、植物组织或植物中。
[0199]
术语“基序”或“共有序列”或“特征(signature)”是指进化相关蛋白的序列中的短保守区。基序常常是结构域的高度保守部分,但是也可只包括结构域的一部分,或位于保守结构域外部(如果基序的所有氨基酸位于所限定结构域的外部)。
[0200]“天然”或“野生型”核酸、多核苷酸、核苷酸序列、多肽或氨基酸序列是指天然存在的或内源性的核酸、多核苷酸、核苷酸序列、多肽或氨基酸序列。
[0201]“核酸分子”或“核苷酸序列”是可以从任何来源中分离的单链或双链dna或rna的区段。在本发明的上下文中,核酸分子典型地是dna的区段。在一些实施例中,本发明的核酸分子是分离的核酸分子。
[0202]
术语“核酸分子”和“多核苷酸”在本文中可以互换使用。
[0203]“可操作地连接”是指在单一核酸片段上多核苷酸的关联,这样使得一者的功能影响另一者的功能。例如,当启动子能够影响编码多核苷酸或功能rna的表达时(即,该编码多核苷酸或功能rna处于该启动子的转录控制之下),则该启动子与该编码多核苷酸或功能rna是可操作地连接的。正义方向或者反义方向的编码多核苷酸能够与调节多核苷酸可操作地连接。
[0204]
术语“蛋白质”、“肽”和“多肽”在本文中可互换使用。
[0205]“植物”是在发育的任何阶段的任何植物,特别是种子植物。
[0206]“植物细胞”是植物的结构和生理单位,包含原生质体和细胞壁。植物细胞可以处于分离的单个细胞或培养细胞的形式,或者是作为较高级的组织单位(如例如,植物组织、植物器官、或全株植物)的一部分。
[0207]“植物细胞培养物”意指植物单元(例如像,原生质体、细胞培养物细胞、植物组织中的细胞、花粉、花粉管、胚珠、胚囊、接合子以及处于不同发育阶段的胚)的培养物。
[0208]“植物材料”是指叶、茎、根、花或花的部分、果实、花粉、卵细胞、接合子、种子、切条、细胞或组织培养物、或植物的任何其他部分或产物。
[0209]“植物器官”是植物的独特而明显的已结构化并且分化的部分,如根、茎、叶、花蕾或胚。
[0210]
如本文所使用的“植物组织”意指组织化成结构和功能单元的一组植物细胞。包括
植物中或培养物中的任何植物组织。这个术语包括但不限于全株植物、植物器官、植物种子、组织培养物以及被组织化成结构和/或功能单元的任何植物细胞群组。这个术语与如以上列出的或由该定义以其他方式涵盖的任何特定类型的植物组织的联合应用或单独应用并不旨在排除任何其他类型的植物组织。
[0211]
本发明多肽的“部分”或“片段”将理解为意指相对于本发明多肽的参考氨基酸序列而言长度减少的氨基酸序列。根据本发明,在适宜的情况下,这种部分或片段可以包括于它作为组分的较大多肽内(例如,标记的或融合蛋白)。在实施例中,所述“部分”或“片段”基本上保留了杀昆虫活性(例如,全长蛋白活性的至少40、50、60、70、80、85、90、95或甚至100,或者具有比全长蛋白更高的杀昆虫活性)。
[0212]“启动子”是编码区的不被翻译的dna序列上游,其含有rna聚合酶结合位点并且起始dna的转录。启动子区还可以包括充当基因表达的调节物的其他元件。
[0213]
如本文所使用的,术语“重组”是指核酸分子(例如,dna或rna)或蛋白质或生物的如下形式,该形式通常不会在自然界中发现并且正因为如此通过人类干预来产生。如本文所使用的,“重组核酸分子”是包括多核苷酸组合的核酸分子,这些多核苷酸不会天然地一起存在并且是人类干预的结果,例如,由至少两种彼此异源的多核苷酸的组合组成的核酸分子,或人工合成的(例如,使用组装的核苷酸序列合成的多核苷酸)并且包含偏离通常存在于自然界中的多核苷酸的多核苷酸的核酸分子,或包含人工掺入至宿主细胞的基因组dna中和该宿主细胞基因组相关侧翼dna中的转基因的核酸分子。重组核酸分子的另一个实例是由将转基因插入至植物的基因组dna中产生的dna分子,其可以最终导致该生物中的重组rna/或蛋白分子的表达。如本文所使用的,“重组植物”是通常不会在自然界中存在的植物,是人类干预的结果,并且含有掺入至其基因组中的转基因和/或异源核酸分子。由于此类基因组改变,重组植物明显不同于相关的野生型植物。
[0214]
如在此使用的,术语“减少(reduce、reduced、reducing、reduction)”、“减小(diminish)”、和“抑制(suppress)”(及其语法变体)和类似术语是指植物有害生物的存活、生长和/或繁殖的减少,例如通过使植物与本发明的多肽接触(如例如,通过转基因表达或通过局部应用方法)。这种存活、生长和/或繁殖的减少可以参考在本发明多肽不存在的情况下观察到的水平(例如,不是转基因表达多肽或不是用多肽局部处理的植物)。因此,在实施例中,术语“减少(reduce、reduced、reducing、reduction)”、“减小(diminish)”、和“抑制(suppress)”(及其语法变体)和类似术语意指如与不与本发明多肽接触的植物(例如,不是转基因表达多肽或不是用多肽局部处理的植物)相比降低至少约5、10、15、20、25、30、35、40、45、50、55、60、65、70、75、80、85、90、95或更多。在代表性实施例中,所述减少导致无或基本上无(即不显著量,例如少于约10、少于约5或甚至少于约1)可检测的植物有害生物的存活、生长和/或繁殖。
[0215]“调节元件”是指参与控制核苷酸序列的表达的序列。调节元件包含可操作地连接至目的核苷酸序列的启动子以及终止信号。它们还典型地涵盖适当翻译该核苷酸序列所需的序列。
[0216]“转化”是用于将异源核酸引入到宿主细胞或生物的方法。在特定实施例中,“转化”意指dna分子稳定地整合到目的生物的基因组(核或质体)中。
[0217]“转化的/转基因的/重组的”是指异源核酸分子已经引入其中的宿主生物例如细
菌或植物。核酸分子可以被稳定地整合到宿主的基因组中,或者核酸分子还可以作为染色体外分子存在。这种染色体外分子能够自主复制。转化的细胞、组织或植物应当理解为不仅涵盖转化过程的终产物,而且涵盖其转基因子代。“非转化的”、“非转基因的”、或“非重组的”宿主是指不含该异源的核酸分子的野生型生物,例如细菌或植物。
[0218]
如本文所用,“txp40蛋白”是由两种细菌属致病杆菌属和发光杆菌属产生的天然毒素,它们分别与线虫属斯氏线虫属和异小杆线虫属共生相关。线虫-细菌对能够侵入和杀死某些昆虫。txp40蛋白首先从发光杆菌中分离出来,并显示出对多种昆虫有害生物具有注射毒性。
[0219]
根据本发明的“变体txp40蛋白”是指与seq id no:1具有至少95%同一性的非天然存在的工程化蛋白,其对灰翅夜蛾属昆虫有害生物,特别是对秋粘虫有害生物(草地贪夜蛾)具有口服毒性。
[0220]
本发明涉及txp40杀昆虫蛋白,其在本文中显示对草地贪夜蛾(秋粘虫,faw)具有令人惊讶的口服毒性,草地贪夜蛾是对许多类别的杀昆虫蛋白,包括源自苏云金芽孢杆菌(bt)的cry蛋白来说难以控制的昆虫有害生物物种。尽管txp40蛋白是本领域中是已知的,但由于它们的天然生物学,它们已显示主要通过直接注射到昆虫的血腔中而对某些昆虫具有杀昆虫性(参见例如,美国专利6,630,619;park等人2012.j.agric.food chem.[农业与食品化学杂志]60:4053-4059以及brown等人2006.app.environ.microbiol.[应用与环境微生物学]72:1653-1662)。这种直接注射模拟了txp40蛋白通过病原性线虫(例如,异小杆线虫科和斯氏线虫科中的昆虫病原线虫)递送至昆虫的天然途径,所述病原性线虫首先侵入靶昆虫血腔并释放产生txp40蛋白的病原性细菌,例如发光杆菌属和致病杆菌属细菌。一个组报道了txp40蛋白仅对大蜡螟(galleria mellonella)具有口服毒性(mathur等人2018.toxicon 154:59-73),大蜡螟是一种非作物有害生物和对许多种类的杀昆虫蛋白高度敏感的昆虫,包括对许多cry蛋白高度敏感。本发明首次公开了txp40蛋白对faw昆虫有害生物的口服毒性。因此,本发明提供了使用txp40杀昆虫蛋白和txp40蛋白变体控制faw有害生物群体的新方法。本发明的新方法特别可用于控制对其他昆虫控制剂如cry蛋白和/或vip3蛋白具有抗性或可能变得具有抗性的faw群体。
[0221]
因此,在一些实施例中,本发明提供了一种控制灰翅夜蛾属昆虫有害生物的方法,其包括使灰翅夜蛾属昆虫有害生物与txp40蛋白接触,所述txp40蛋白包含与seq id no:1具有至少95%同一性的氨基酸序列,基本上由其组成或由其组成。在其他实施例中,txp40蛋白包含在其全长上与seq id no:1具有至少95%、或至少96%、或至少97%、或至少98%、或至少99%同一性的氨基酸序列,基本上由其组成或由其组成。在其他实施例中,txp40蛋白包含seq id no:1、seq id no:2或seq id no:74-136中的任一个,基本上由其组成或由其组成。在其他实施例中,方法包括向灰翅夜蛾属昆虫有害生物或其环境递送,例如口服递送有效量的本发明的txp40蛋白。通常,为了有效,txp40蛋白被灰翅夜蛾属昆虫有害生物口服摄入。将本发明的txp40蛋白质经口递送至灰翅夜蛾属昆虫有害生物的方式的实例包括但不限于(1)在转基因植物中提供蛋白,其中昆虫摄食(摄取)转基因植物的一个或多个部分,从而摄取在转基因植物中表达的txp40多肽;(2)在可施用于或掺入例如昆虫生长培养基的一个或多个配制的蛋白质组合物中提供蛋白;(3)在一种或多种蛋白组合物中提供蛋白,该蛋白组合物可以施用到表面,例如喷雾到植物部分的表面上,然后当昆虫食用一个或
多个喷雾的植物部分时被昆虫摄取;(4)诱饵基质;或(5)任何其他本领域公认的蛋白质递送系统。因此,口服递送到灰翅夜蛾属昆虫有害生物的任何方法可用于递送本发明的毒性蛋白。在一些特定的实施例中,将本发明的txp40杀昆虫蛋白经口递送至灰翅夜蛾属昆虫有害生物,例如,其中昆虫摄取本发明的转基因植物的一个或多个部分。在进一步的实施例中,灰翅夜蛾属昆虫有害生物是草地贪夜蛾(秋粘虫;faw)。
[0222]
在其他实施例中,将本发明的txp40杀昆虫蛋白经口递送到灰翅夜蛾属昆虫有害生物,例如秋粘虫昆虫(草地贪夜蛾),其中昆虫摄取用包含本发明的txp40杀昆虫蛋白的组合物喷雾的植物的一个或多个部分。可以使用本领域技术人员已知的用于将化合物、组合物、配制品等应用于植物表面的任何方法将本发明的组合物递送至植物表面。递送至或接触植物或其部分的一些非限制性实例包括喷雾、撒粉、喷洒、分散、下雾、雾化、撒播、浸泡、土壤注入、土壤掺入、浸透(例如,根、土壤处理)、浸渍、灌注、涂覆、叶或茎渗透、侧施或种子处理等及其组合。用于使植物或其部分与一种或多种化合物、一种或多种组合物或一种或多种配制品接触的这些和其他程序是本领域技术人员熟知的。
[0223]
在进一步的实施例中,本发明提供了一种控制对cry蛋白和/或营养期杀昆虫蛋白例如vip3具有抗性的faw昆虫有害生物或有害生物群体的方法,其包括向faw有害生物或有害生物群体或其环境递送有效量的本发明的txp40蛋白或组合物。在一些实施例中,txp40蛋白包含与seq id no:1具有至少95%同一性的氨基酸序列,基本上由其组成或由其组成。在其他实施例中,txp40蛋白包含在其全长上与seq id no:1具有至少95%、或至少96%、或至少97%、或至少98%、或至少99%同一性的氨基酸序列,基本上由其组成或由其组成。在其他实施例中,txp40蛋白的氨基酸序列包含seq id no:1、seq id no:2或seq id no:74-136中的任一个,基本上由其组成或由其组成。在其他实施例中,vip3是vip3a蛋白。在其他实施例中,vip3a蛋白是在包含转基因事件mir162的玉米植物中表达的vip3aa20蛋白。在又其他实施例中,cry蛋白是cry1a、cry1b、cry1c、cry1d、cry1f、cry1j或cry2a蛋白。在进一步的实施例中,cry1f蛋白是在包含转基因事件tc1507的玉米植物中表达的cry1fa蛋白。
[0224]
在其他实施例中,本发明提供了一种防止灰翅夜蛾属昆虫有害生物(例如秋粘虫)群体对转基因植物中表达的cry蛋白和/或vip3蛋白发展抗性的方法,该方法包括向靶灰翅夜蛾群体递送包含多核苷酸的转基因植物,该多核苷酸包含编码vip3蛋白的核苷酸序列和/或编码cry蛋白的核苷酸序列;和表达本发明的txp40杀昆虫蛋白的本发明的多核苷酸表达盒或载体。在其他实施例中,vip3蛋白是vip3a蛋白。在其他实施例中,vip3a蛋白是vip3aa20蛋白。在又其他实施例中,cry蛋白是cry1a、cry1b、cry1c、cry1d、cry1f、cry1j或cry2a蛋白。在进一步的实施例中,cry1f蛋白是cry1fa蛋白。在又其他实施例中,转基因植物是包含事件mir162和/或事件tc1507的玉米植物。在一些实施例中,txp40蛋白包含在其全长上与seq id no:1具有至少95%同一性的氨基酸序列,基本上由其组成或由其组成。在另一实施例中,txp40蛋白包含seq id no:1、seq id no:2或seq id no:74-136中的任一个,基本上由其组成或由其组成。根据前述实施例,转基因植物可包含编码本发明的txp40杀昆虫蛋白的两个或更多个基因的育种叠加、编码本发明的txp40杀昆虫蛋白的两个或更多个基因的分子叠加、或两者的组合。
[0225]
本发明还提供了变体txp40杀昆虫蛋白,其包含与seq id no:1具有至少95%、96%、97%、98%、99%、99.1%、99.2%、99.3%、99.4%、99.5%、99.6%、99.7%、99.8%或
99.9%同一性的氨基酸序列,基本上由其组成或由其组成,其中所述变体txp40蛋白对灰翅夜蛾属昆虫有害生物,特别是对草地贪夜蛾(秋粘虫,faw)昆虫有害生物具有口服活性。在一些实施例中,本发明的变体txp40蛋白包含seq id no:2或seq id no:74-136中的任一个的氨基酸序列,基本上由其组成或由其组成。
[0226]
在一些实施例中,本发明提供了与seq id no:1具有至少95%、96%、97%、98%、99%、99.1%、99.2%、99.3%、99.4%、99.5%、99.6%、99.7%、99.8%或99.9%同一性且与seq id no:1相比具有氨基酸取代的变体txp40杀昆虫蛋白。在其他实施例中,本发明提供了变体txp40杀昆虫蛋白,其与seq id no:1具有至少95%、96%、97%、98%、99%、99.1%、99.2%、99.3%、99.4%、99.5%、99.6%、99.7%、99.8%或99.9%同一性,并且与seq id no:1相比,在对应于seq id no:1的氨基酸位置10、11、26、31、38、46、48、49、73、75、86、96、101、103、111、116、119、133、134、143、155、167、169、170、182、186、187、191、200、208、209、210、213、217、221、226、238、240、247、250、252、256、257、263、265、266、271、275、284、293、296、305、308、309、311、313、315、320、326、328或333或其任何组合的位置具有氨基酸取代。
[0227]
在一些实施例中,本发明提供了变体txp40杀昆虫蛋白,其与seq id no:1具有至少95%、96%、97%、98%、99%、99.1%、99.2%、99.3%、99.4%、99.5%、99.6%、99.7%、99.8%或99.9%同一性,并且与seq id no:1相比,在对应于seq id no:1的氨基酸位置119和/或213的位置具有氨基酸取代。在其他实施例中,变体txp40蛋白在对应于seq id no:1的氨基酸位置119的位置具有a和/或在对应于氨基酸位置213的位置具有a。在其他实施例中,变体txp40蛋白在seq id no:1中具有k119a和/或k213a取代。在其他实施例中,变体txp40蛋白包含seq id no:18、seq id no:26或seq id no:127的氨基酸序列,基本上由其组成或由其组成。
[0228]
在一些实施例中,本发明提供了变体txp40杀昆虫蛋白,其与seq id no:1具有至少95%、96%、97%、98%、99%、99.1%、99.2%、99.3%、99.4%、99.5%、99.6%、99.7%、99.8%或99.9%同一性,并且与seq id no:1相比,在对应于seq id no:1的氨基酸位置11、119和/或213的位置具有氨基酸取代。在其他实施例中,变体txp40蛋白在对应于seq id no:1的氨基酸位置11的位置具有y、在对应于seq id no:1的氨基酸位置119的氨基酸位置具有a、和/或在对应于seq id no:1的氨基酸位置213的氨基酸位置具有a。在其他实施例中,变体txp40蛋白是seq id no:1,其中位置11的氨基酸s被y取代,位置119的氨基酸k被a取代和/或位置213的氨基酸k被a取代。在其他实施例中,变体txp40蛋白包含seq id no:131的氨基酸序列,基本上由其组成或由其组成。
[0229]
认识到编码本发明的txp40蛋白的核苷酸序列可通过各种方法改变,并且这些改变可导致编码其氨基酸序列不同于天然存在的txp40蛋白所编码的序列的变体txp40蛋白的核苷酸序列。txp40蛋白可以按照不同的方式进行改变,包括seq id no:1的一个或多个氨基酸的氨基酸取代、缺失、截短、以及插入,包括多达约2个、约3个、约4个、约5个、约6个、约7个、约8个、约9个、约10个、约15个、约17个、或约20个氨基酸取代、缺失或插入。用于此类操作的方法在本领域中通常是已知的。例如,通过在编码txp40蛋白的多核苷酸中的突变可以制备天然txp40蛋白的氨基酸序列变体。这还可以通过若干种诱变形式之一或在定向进化中来完成。在一些实施例中,在该氨基酸序列中所编码的改变将基本上不影响该蛋白质
的功能。此类变体将具有所希望的杀昆虫活性。在其他实施例中,所得变体txp40蛋白与野生型txp40蛋白相比具有增加的杀昆虫活性,特别是针对灰翅夜蛾属昆虫有害生物。在其他实施例中,所得变体蛋白质由合成变体多核苷酸编码。在又其他实施例中,本发明的txp40蛋白由包含seq id no:11-73中的任一个的核苷酸序列,基本上由其组成或由其组成的核酸分子编码。
[0230]
根据一些实施例,本发明提供了包含编码本发明txp40蛋白的核苷酸序列,基本上由其组成或由其组成的合成多核苷酸,所述txp40蛋白对灰翅夜蛾属有害生物,特别是对秋粘虫具有口服毒性,其中所述核苷酸序列已进行密码子优化以在转基因生物体中表达。在其他实施例中,转基因生物是转基因细菌或转基因植物。在另外的实施例中,该转基因植物是转基因玉米植物。尽管在许多情况下,来自微生物的基因能够在植物中高水平表达而无需修饰,但在转基因植物中的低表达可能是由于微生物核苷酸序列的缘故,这些序列具有在植物中并不优选的密码子。在本领域中已知,活生物具有特定的密码子使用偏好,而且在本发明中所描述的这些核苷酸序列的密码子可以被改变以符合植物偏好,同时维持由其编码的氨基酸。此外,在本领域中已知,在植物(例如玉米植物)中高表达可以是由如下编码序列实现的,所述编码序列具有至少约35、或至少约45、或至少约50、或至少约60的gc含量。具有低gc含量的微生物核苷酸序列在植物中也许表达欠佳。尽管某些核苷酸序列均可以在单子叶植物和双子叶植物物种两者中充分表达,但是可以对序列进行修饰以便迎合单子叶植物或双子叶植物特定的密码子偏好以及gc含量偏好,因为这些偏好已经被证明是不同的(murray等人,nucl.acids res.[核酸研究]17:477-498(1989))。此外,在实施例中,修饰所述核苷酸序列以去除可能导致消息平截(message truncation)的不正常剪接位点。使用描述于例如美国专利号5,625,136、5,500,365和6,013,523中的方法,使用熟知的定点诱变、pcr以及合成基因构建技术,可以对这些核苷酸序列做出此类修饰。在本发明的一些实施例中,本发明的密码子优化的核苷酸序列包含具有至少50、55、60或至少65gc含量的核苷酸序列,基本上由其组成或由其组成。在又其他实施例中,密码子优化的核苷酸序列包含seq id no:4、seq id no:5或seq id no:11-73中任一个的核苷酸序列,基本上由其组成或由其组成。本领域技术人员将认识到,这种gc含量可以以多种不同的方式实现。例如,根据美国专利号5,625,136中公开的程序制备这样的合成编码序列或多核苷酸。在这个程序中,使用了玉蜀黍优选的密码子,即最频繁地编码玉蜀黍中的氨基酸的单一密码子。针对特定的氨基酸的玉蜀黍偏好的密码子可衍生自例如来自玉蜀黍的已知基因序列。例如,针对来自玉蜀黍植物的28个基因的玉蜀黍密码子使用发现于以下文献中:murray等人,nucleic acids research[核酸研究]17:477-498(1989)。应当认识到,密码子进行优化用于在一种植物物种中的表达也将作用于其他植物物种,但密码子进行优化用于表达的植物物种可能不在相同水平。以这种方式,这些核苷酸序列可以进行优化用于在任何植物中表达。应当认识到,核苷酸序列的全部或任何部分可以是优化的或合成的。也就是说,多核苷酸可以包含作为部分天然序列和部分密码子优化序列的核苷酸序列。
[0231]
在本发明的一些实施例中,提供了嵌合基因,其包含与异源核酸分子可操作地连接的启动子,所述异源核酸分子包含编码对灰翅夜蛾属昆虫有害生物具有杀昆虫活性的txp40蛋白的核苷酸序列,其中所述核苷酸序列:(a)是seq id no:3-5或11-73中的任一个;(b)与seq id no:3-5或11-73中的任一个具有至少95%同一性;(c)编码包含seq id no:1、
2或74-136中的任一个的氨基酸序列的蛋白质;(d)编码包含与seq id no:1具有至少95%同一性的氨基酸序列的蛋白质;或(e)与(a)至(d)中的任一个的核苷酸序列互补。
[0232]
在一些实施例中,核酸分子包含玉蜀黍密码子优化的核苷酸序列。在其他实施例中,启动子是植物可表达的启动子,特别是在玉蜀黍植物中表达的启动子。例如但不限于,该植物可表达型启动子选自下组启动子,该组由以下组成:泛素、夜香树属黄病毒、玉米trpa、osmads6、玉蜀黍h3组蛋白、噬菌体t3基因9 5'utr、玉米蔗糖合成酶1、玉米醇脱氢酶1、玉米捕光复合物、玉米热休克蛋白、玉蜀黍mtl、豌豆小亚基rubp羧化酶、水稻肌动蛋白、水稻亲环蛋白、ti质粒甘露碱合酶、ti质粒胭脂碱合酶、矮牵牛查尔酮异构酶、豆类富甘氨酸蛋白1、马铃薯糖蛋白(potato patatin)、凝集素、camv 35s以及s-e9小亚基rubp羧化酶启动子。
[0233]
尽管已经显示来自双子叶植物的很多启动子在单子叶植物中是可操作的并且反之亦然,但在实施例中,选择双子叶植物启动子用于在双子叶植物中表达,并且选择单子叶植物启动子用于在单子叶植物中表达。不过,对于所选的启动子的起源没有限制;只要它们可有效驱动核苷酸序列在所希望细胞中表达就足够了。
[0234]
启动子的选择可以依赖于表达的时间和空间需要而变化,并且还依赖于有待转化的宿主细胞而变化。因此,例如,本发明的核苷酸序列的表达可以处于任何植物和/或植物部分中(例如,处于叶子中、处于茎杆或茎中、处于穗中、处于花序中(例如,穗状花序、圆锥花序、穗轴等)、处于根、种子和/或幼苗等中)。例如,在希望在特异性组织或器官中表达的情况下,可以使用组织特异性或组织优选的启动子(例如,根特异性或优选的启动子)。相比之下,在希望响应刺激而表达的情况下,可以使用可由刺激或化学物诱导的启动子。在希望在植物的所有细胞中以相对恒定水平连续表达的情况下,可以选择组成型启动子。
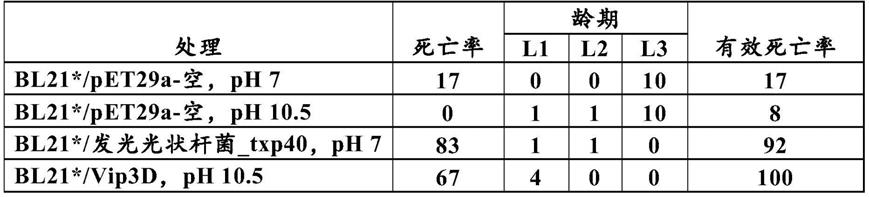
[0235]
适用于本发明的启动子包括但不限于组成性地驱动核苷酸序列的表达的那些启动子、在诱导时驱动表达的那些启动子、以及以组织或发育特异性方式驱动表达的那些启动子。这些不同类型的启动子在本领域是已知的。
[0236]
合适的组成型启动子包括例如camv 35s启动子(odell等人,nature[自然]313:810-812,1985);拟南芥at6669启动子(参见pct公开号w004081173a2);玉蜀黍ubi 1(christensen等人,plant mol.biol.[植物分子生物学]18:675-689,1992);水稻肌动蛋白(mcelroy等人,plant cell[植物细胞]2:163-171,1990);pemu(last等人,theor.appl.genet.[理论与应用遗传学]81:581-588,1991);camv19s(nilsson等人,physiol.plant[植物生理学]100:456-462,1997);gos2(de pater等人,plant j[植物杂志]11月;2(6):837-44,1992);泛素(christensen等人,plant mol.biol.[植物分子生物学]18:675-689,1992);水稻亲环蛋白(bucholz等人,plant mol biol.[植物分子生物学]25(5):837-43,1994);玉蜀黍h3组蛋白(lepetit等人,mol.gen.genet.[分子遗传学和普通遗传学]231:276-285,1992);肌动蛋白2(an等人,plant j.[植物杂志]10(1);107-121,1996)、组成型根尖ct2启动子(seq id no:1535;还参见pct申请号il/2005/000627)和合成超级mas(ni等人,the plant journal[植物杂志]7:661-76,1995)。其他组成型启动子包括美国专利号5,659,026、5,608,149;5,608,144;5,604,121;5,569,597;5,466,785;5,399,680;5,268,463;和5,608,142中的那些。
[0237]
对于在植物(任选地是玉蜀黍)中表达本发明的多肽是有用的组织特异性的或组
织优选的启动子包括那些直接在根、髓、叶或花粉中表达的启动子。合适的组织特异性启动子包括但不限于叶特异性启动子(例如yamamoto等人,plant j.[植物杂志]12:255-265,1997;kwon等人,plant physiol.[植物生理学]105:357-67,1994;yamamoto等人,plant cell physiol.[植物与细胞生理学]35:773-778,1994;gotor等人,plant j.[植物杂志]3:509-18,1993;orozco等人,plant mol.biol.[植物分子生物学]23:1129-1138,1993;和matsuoka等人,proc.natl.acad.sci.usa[美国国家科学院院刊]90:9586-9590,1993所述)、种子优选的启动子(例如来自种子特异性基因;simon等人,plant mol.biol.[植物分子生物学]5.191,1985;scofield等人,j.biol.chem.[生物化学杂志]262:12202,1987;baszczynski等人,plant mol.biol.[植物分子生物学]14:633,1990)、巴西坚果白蛋白(pearson等人,plant mol.biol.[植物分子生物学]18:235-245,1992)、豆球蛋白(ellis等人plant mol.biol.[植物分子生物学]10:203-214,1988)、谷蛋白(takaiwa等人,mol.gen.genet.[分子遗传学和普通遗传学]208:15-22,1986;takaiwa等人,febs letts.221:43-47,1987)、玉米醇溶蛋白(matzke等人,plant mol biol,[植物分子生物学]143).323-32 1990)、napa(stalberg等人,planta[植物学]199:515-519,1996)、小麦spa(albanietal,plant cell[植物细胞],9:171-184,1997)、向日葵油质蛋白(cummins等人,plant mol.biol.[植物分子生物学]19:873-876,1992)、胚乳特异性启动子(例如,小麦lmw和hmw、麦谷蛋白-1(mol gen genet[分子遗传学和普通遗传学]216:81-90,1989;nar17:461-2)、小麦a、b和g麦醇溶蛋白(emb03:1409-15,1984)、大麦ltrl启动子、大麦b1、c、d大麦醇溶蛋白(theor appl gen[理论与应用遗传学]98:1253-62,1999;plant j[植物杂志]4:343-55,1993;mol gen genet[分子遗传学和普通遗传学]250:750-60,1996)、大麦dof(mena等人,the plant journal[植物杂志],116(1):53-62,1998)、biz2(ep99106056.7)、合成启动子(vicente-carbajosa等人,plant j.[植物杂志]13:629-640,1998)、水稻醇溶蛋白nrp33、水稻球蛋白glb-1(wu等人,plant cell physiology[植物细胞生理学]39(8)885-889,1998)、水稻α-球蛋白reb/ohp-1(nakase等人.plant mol.biol.[植物分子生物学]33:513-s22,1997)、水稻adp-葡萄糖pp(trans res[翻译研究]6:157-68,1997)、玉蜀黍esr基因家族(plant j[植物杂志]12:235-46,1997)、高粱γ-高粱醇溶蛋白(plant mol.biol[植物分子生物学]32:1029-35,1996)]、胚胎特异性启动子(例如水稻osh1;sato等人,proc.nati.acad.sci.usa[美国国家科学院院刊],93:8117-8122)、knox(postma-haarsma等人,plant mol.biol.[植物分子生物学]39:257-71,1999)、水稻油质蛋白(wu等人,j.biochem.[生物化学杂志],123:386,199)]花特异性启动子,例如atprp4、查尔酮合成酶(chsa)(van der meer等人,plant mol.biol.[植物分子生物学]15,95-109,1990)、lat52(twell等人,mol.gen genet.[分子遗传学和普通遗传学]217:240-245;1989)、无瓣-3和植物生殖组织特异性启动子(例如,osmads启动子;美国专利公开2007/0006344)。
[0238]
适用于在绿色组织中优选表达的启动子的实例包括调节参与光合作用的基因的许多启动子,并且这些中的许多已经从单子叶植物和双子叶植物两者中得以克隆。一种此类启动子是来自磷酸烯醇羧化酶基因的玉蜀黍pepc启动子(hudspeth和grula,plant molec.biol.[植物分子生物学]12:579-589(1989))。另一种用于根特异性表达的启动子是由de framond(febs 290:103-106(1991)或美国专利号5,466,785)描述的启动子。另一种在本发明中有用的启动子是描述于美国专利号5,625,136中的茎特异性启动子,它天然地
驱动玉蜀黍trpa基因的表达。
[0239]
此外,可以使用在质体中发挥作用的启动子。此类启动子的非限制性实例包括噬菌体t3基因9 5'utr以及其他的披露于美国专利号7,579,516中的启动子。适用于本发明的其他启动子包括但不限于s-e9小亚基rubp羧化酶启动子和kunitz胰蛋白酶抑制剂基因启动子(kti3)。
[0240]
在本发明的一些实施例中,可以使用诱导型启动子。因此,例如,可以使用化学调节的启动子以通过应用外源化学调节物来调节植物中的基因表达。本发明的核苷酸序列的表达经由化学调节过的启动子进行的调节使得本发明的多肽仅当用诱导的化学物处理作物植物时能被合成。取决于目的,在应用化学物诱导基因表达时启动子可以是化学诱导型启动子,或者在应用化学物阻抑基因表达时启动子可以是化学阻抑型启动子。用于基因表达的化学诱导的此类技术的实例详述于公开申请ep 0 332 104和美国专利号5,614,395中。
[0241]
化学诱导型启动子在本领域是已知的,并且包括但不限于玉蜀黍in2-2启动子(它是由苯磺酰胺除草剂安全剂激活)、玉蜀黍gst启动子(它是由用作发芽前除草剂的疏水亲电子化合物激活)、以及烟草pr-1a启动子(它是由水杨酸激活)(例如,pr1a系统)、类固醇反应性启动子(参见例如,schena等人(1991)proc.natl.acad.sci.usa[美国国家科学院院刊]88,10421-10425和mcnellis等人(1998)plant j.[植物杂志]14,247-257)中的糖皮质激素诱导型启动子)以及四环素诱导型启动子和四环素-阻抑型启动子(参见例如,gatz等人(1991)mol.gen.genet.[分子遗传学与普通遗传学]227,229-237和美国专利号5,814,618和5,789,156)、lac阻遏物系统启动子、铜诱导型系统启动子、水杨酸诱导型系统启动子(例如,pr1a系统)、糖皮质激素诱导型启动子(aoyama等人(1997)plant j.[植物杂志]11:605-612)以及蜕皮激素诱导型系统启动子。
[0242]
诱导型启动子的其他非限制性实例包括aba诱导型和细胞膨胀诱导型启动子、植物生长素结合蛋白基因启动子(schwob等人(1993)plant j.[植物杂志]4:423-432)、udp葡萄糖类黄酮糖基转移酶启动子(ralston等人(1988)genetics[遗传学]119:185-197)、mpi蛋白酶抑制剂启动子(cordero等人(1994)plant j.[植物杂志]6:141-150)以及甘油醛-3-磷酸脱氢酶启动子(kohler等人(1995)plant mol.biol.[植物分子生物学]29:1293-1298;martinez等人(1989)j.mol.biol.[分子生物学杂志]208:551-565;和quigley等人(1989)j.mol.evol.[分子进化杂志]29:412-421)。还包括苯磺酰胺诱导型(美国专利号5,364,780)和乙醇诱导型(国际专利申请公开号wo 97/06269和wo 97/06268)系统和谷胱甘肽s-转移酶启动子。同样地,可以使用描述于以下文献中的诱导型启动子中的任一种:gatz(1996)current opinion biotechnol.[生物技术新见]7:168-172和gatz(1997)annu.rev.plant physiol.plant mol.biol.[植物生理学与植物分子生物学年度综述]48:89-108。适用于指导本发明的核苷酸序列在植物中的表达的其他化学诱导型启动子披露于美国专利5,614,395中。基因表达的化学诱导还详述于ep 0 332 104(授予汽巴-嘉基公司(ciba-geigy))和美国专利5,614,395中。
[0243]
另一类在本发明中有用的启动子是创伤诱导型启动子。已经描述了数量众多的在创伤部位并且还在植物病原菌感染的部位表达的启动子。理想的是,这样的启动子在昆虫入侵的部位应该仅有局部活性,并且以此方式这些杀昆虫蛋白仅在需要合成这些杀昆虫蛋
白的细胞中积聚以杀死入侵的昆虫有害生物。这种启动子的实例包括stanford等人mol.gen.genet.[分子遗传学与普通遗传学]215:200-208(1989)、xu等人plant molec.biol.[植物分子生物学]22:573-588(1993)、logemann等人plant cell[植物细胞]1:151-158(1989)、rohrmeier&lehle,plant molec.biol.[植物分子生物学]22:783-792(1993)、firek等人plant molec.biol.[植物分子生物学]22:129-142(1993)、以及warner等人plant j.[植物杂志]3:191-201(1993)所述的那些。
[0244]
除与本发明的核苷酸序列可操作地相关联的启动子之外,本发明的表达盒还可以包括其他调节元件。调节元件包括但不限于增强子、内含子、翻译前导序列、终止信号、以及多腺苷酸化信号序列。适合的转录终止子信号的实例是可获得的并且在本领域中是已知的(例如来自camv的tml,来自rbcs的e9)。任何已知在植物中发挥作用的可供使用的终止子均可以在本发明的上下文中使用。
[0245]
可以将数量众多的其他序列掺入本发明所描述的嵌合基因中。这些序列包括已经显示出增强表达的序列,如内含子序列(例如,来自adhl和bronzel)以及病毒的前导序列(例如,来自tmv、mcmv、和amv)。
[0246]
为了更加有效的翻译起始,可修饰与起始甲硫氨酸相邻的序列。例如,它们可以通过包含已知在植物中有效的序列而被修饰。joshi已经提出了针对植物的适当的共有序列(nar 15:6643-6653(1987)),并且clonetech提出了另一种共有翻译起始子(1993/1994目录,第210页)。这些共有序列适于与本发明的核苷酸序列一起使用。将这些序列掺入至包含核苷酸序列的构建体中,达到atg并且包括atg(同时保持不修饰第二氨基酸),或者可替代地达到atg后的gtc并且包括atg后的gtc(具有修饰该转基因的第二氨基酸的可能性)。
[0247]
在一些实施例中,希望的是将本发明的多肽的表达靶向至植物细胞中的特定细胞定位。在一些情况下,在胞质溶胶中的定位可能是令人希望的,而在其他情况下,在某个亚细胞器中的定位可能是优选的。用于靶向例如植物中的基因产物的任何机构都可以用于实践本发明,并且已知此类机构存在于植物中并且已经相当详细地表征了控制这些机构的功能的序列。已经表征了导致将基因产物靶向其他细胞区室的序列。例如,氨基末端序列可以负责将目的蛋白质靶向细胞区室,如植物细胞的液泡、线粒体、过氧化物酶体、蛋白体、内质网、叶绿体、淀粉颗粒、淀粉体、质外体或细胞壁(例如unger等人,plant molec.biol.[植物分子生物学]13:411-418(1989);rogers等人,(1985)proc.natl.acad.sci.usa[美国国家科学院院刊]82:6512-651;美国专利号7,102,057;wo 2005/096704)。任选地,信号序列可以是来自waxy的n-末端信号序列、来自γ-玉米醇溶蛋白的n-末端信号序列、淀粉结合结构域、c-末端淀粉结合结构域、将成熟蛋白质导入叶绿体的叶绿体靶向序列(comai等人(1988)j.biol.chem.[生物化学杂志]263:15104-15109;van den broeck等人(1985)nature[自然]313:358-363;美国专利号5,639,949)或来自糊粉细胞的分泌信号序列(koehler&ho,plant cell[植物细胞]2:769-783(1990))。另外,与羧基末端序列结合的氨基末端序列负责基因产物的液泡靶向,并且可以随本发明一起使用(shinshi等人,(1990)plant molec.biol.[植物分子生物学]14:357-368)。在一个实施例中,所选择的信号序列包括已知的切割位点,并且构建的融合体考虑了在一个或多个切割位点之后的需要切割的任何氨基酸。在一些情况下,这个要求可以通过在切割位点与转基因atg之间添加小数目的氨基酸,或可替代地置换转基因序列内的一些氨基酸来满足。这些构建技术在本领域是熟
知的并且同样适用于任何细胞区室。
[0248]
应认识到,用于细胞靶向的上述机制不仅可以与其同源启动子结合使用,还可以与异源启动子结合使用,从而在启动子的转录调节下实现特定的细胞靶向目标,该启动子具有不同于自其衍生靶向信号的启动子的表达谱。
[0249]
本发明的嵌合基因还可以包括用于选择性标记的核苷酸序列,所述选择性标记可以用于选择转化的植物、植物部分和/或植物细胞。适合的选择性标记的许多例子是本领域已知的并且可以用于在此描述的表达盒中。
[0250]
选择性标记的实例包括但不限于编码neo或nptii的核苷酸序列,其赋予对卡那霉素、g418等的抗性(potrykus等人(1985)mol.gen.genet.[分子遗传学和普通遗传学]199:183-188);编码bar的核苷酸序列,其赋予对膦丝菌素的抗性;编码改变的5-烯醇丙酮酰莽草酸-3-磷酸(epsp)合酶的核酸序列,其赋予对草甘膦的抗性(hinchee等人(1988)biotech.[生物科技]6:915-922);编码腈水解酶如来自臭鼻杆菌的bxn的核苷酸序列,其赋予对溴苯腈的抗性(stalker等人(1988)science[科学]242:419-423);编码改变的乙酰乳酸合酶(als)的核苷酸序列,其赋予对咪唑啉酮、磺酰脲或其他als抑制性化学品的抗性(ep专利申请号154204);编码甲氨蝶呤抗性二氢叶酸还原酶(dhfr)的核苷酸序列(thillet等人(1988)j.biol.chem.[生物化学杂志]263:12500-12508);编码茅草枯脱卤素酶的核苷酸序列,其赋予对茅草枯的抗性;编码甘露糖-6-磷酸异构酶(也称为磷酸甘露糖异构酶(pmi))的核苷酸序列,其赋予代谢甘露糖的能力(美国专利号5,767,378和5,994,629);编码改变的邻氨基苯甲酸合酶的核苷酸序列,其赋予对5-甲基色氨酸的抗性;或编码hph的核苷酸序列,其赋予对潮霉素的抗性。本领域技术人员能够选择用于在本发明的表达盒中使用的适合的选择性标记。
[0251]
另外的选择性标记包括但不限于编码β-葡糖醛酸糖苷酶的核苷酸序列或编码多种显色底物已知的酶的uida(gus);编码调节植物组织中花色苷色素(红色)产生的产物的r-基因座核苷酸序列(dellaporta等人,“molecular cloning of the maize r-nj allele by transposon-tagging with ac[通过ac转座子标签技术对玉蜀黍r-nj等位基因的分子克隆]”263-282:chromosome structure and function:impact of new concepts[染色体结构和功能:新概念的影响],18th stadler genetics symposium[第十八届斯塔德勒遗传学研讨会](gustafson&appels编著,plenum出版社1988));编码β-内酰胺酶的核苷酸序列,β-内酰胺酶是多种显色底物已知的酶(例如padac,一种显色头孢菌素)(sutcliffe(1978)proc.natl.acad.sci.usa[美国国家科学院院刊]75:3737-3741);编码xyle的核苷酸序列,所述xyle编码儿茶酚双加氧酶(zukowsky等人(1983)proc.natl.acad.sci.usa[美国国家科学院院刊]80:1101-1105);编码酪氨酸酶的核苷酸序列,所述酪氨酸酶是能够将酪氨酸氧化成dopa和多巴醌的酶,dopa和多巴醌又缩合形成黑色素(katz等人(1983)j.gen.microbiol.[普通微生物学杂志]129:2703-2714);编码β-半乳糖苷酶的核苷酸序列,β-半乳糖苷酶是存在显色底物的酶;编码允许生物发光检测的荧光素酶(lux)的核苷酸序列(ow等人(1986)science[科学]234:856-859);编码可用于钙敏感生物发光检测的水母发光蛋白的核苷酸序列(prasher等人(1985)biochem.biophys.res.comm.[生物化学和生物物理学研究通讯]126:1259-1268);或编码绿色荧光蛋白的核苷酸序列(niedz等人(1995)plant cell reports[植物细胞报告]14:403-406)。本领域技术人员能够选择用于
在本发明的表达盒中使用的适合的选择性标记。
[0252]
在一些实施例中,本发明的嵌合基因还可以包括编码除本发明的嵌合杀昆虫蛋白之外的其他所希望的性状的多核苷酸。此类其他多核苷酸的实例包括编码多肽或dsrna以获得感兴趣的其他所希望的性状的那些。此类包含“叠加”性状的表达盒可以用来例如产生具有所希望的具有叠加性状(即,分子叠加)的表型的植物、植物部分或植物细胞。植物中的此类叠加的组合还可以通过其他方法来产生,包括但不限于通过任何常规的方法学来杂交育种植物(即,育种叠加)。如果是通过遗传转化这些植物来进行叠加的,感兴趣的核苷酸序列可以在任何时间并且以任何次序进行组合。例如,包含一种或多种所希望的性状的转基因植物可以用作通过后续转化而引入另外的性状的靶标。另外的核苷酸序列可以在共转化方案中与由表达盒的任何组合提供的本发明的核苷酸序列、核酸分子、核酸构建体、或组合物同时引入。例如,如果将引入两个核苷酸序列,则它们可以合并在分开的盒(反式)中或可以合并在相同的盒(顺式)上。多核苷酸的表达可以通过相同的启动子或通过不同的启动子驱动。进一步认识到多核苷酸可以使用位点特异性重组系统在所希望的基因组位置处叠加。参见,例如,国际专利申请公开号wo 99/25821;wo 99/25854、wo99/25840、wo 99/25855和wo 99/25853。
[0253]
在代表性实施例中,所述嵌合基因还可以包括用于农艺性状(例如,主要受益者是种子公司、栽培者或谷物加工者的农艺性状)的感兴趣的一种或多种多肽或双链rna分子(dsrna)的另外的编码序列。感兴趣的多肽可以是由感兴趣的核苷酸序列编码的任何多肽。适合用于在植物中生产的感兴趣的多肽的非限制性实例包括产生农艺学重要性状的那些多肽,这些性状如除草剂抗性(有时也称为“除草剂耐受性”)、病毒抗性、细菌病原体抗性、昆虫抗性、线虫抗性、或真菌抗性。参见,例如美国专利号5,569,823、5,304,730、5,495,071、6,329,504和6,337,431。在实施例中,感兴趣的多肽可以是提高植物活力或产量(包括允许植物在不同的温度、土壤条件以及日光和沉淀水平下生长的性状)的多肽、或是允许对展现感兴趣的性状(例如,选择性标记、种皮颜色等)的植物进行鉴定的多肽。不同的目的多肽,连同用于将这些多肽引入植物中的方法描述于例如美国专利号4,761,373、4,769,061、4,810,648、4,940,835、4,975,374、5,013,659、5,162,602、5,276,268、5,304,730、5,495,071、5,554,798、5,561,236、5,569,823、5,767,366、5,879,903、5,928,937、6,084,155、6,329,504和6,337,431,连同美国专利公开号2001/0016956中。还参见互联网上lifesci.sussex.ac.uk/home/neil_crickmore/bt/。
[0254]
赋予对抑制生长点或分生组织的除草剂(如咪唑啉酮或磺酰脲)的抗性/耐受性的多核苷酸也可以适用于本发明的一些实施例中。对于突变型als和ahas酶在这一分类号中的示例性多核苷酸如描述于例如,美国专利号5,767,366和5,928,937中。美国专利号4,761,373和5,013,659针对抵抗不同的咪唑啉酮或磺酰脲除草剂的植物。美国专利号4,975,374涉及含有如下核酸的植物细胞和植物,所述核酸编码突变型谷氨酰胺合成酶(gs),所述突变型谷氨酰胺合成酶抵抗已知抑制gs的除草剂(例如,草丁膦和甲硫氨酸磺基肟(methionine sulfoximine))的抑制作用。美国专利号5,162,602披露了抵抗环己二酮和芳氧苯氧丙酸除草剂的抑制作用的植物。该抗性由改变的乙酰辅酶a羧化酶(accase)赋予。
[0255]
赋予对草甘膦抗性的由核苷酸序列编码的多肽也适用于本发明。参见,例如,美国专利号4,940,835和美国专利号4,769,061。美国专利号5,554,798披露了抗草甘膦的转基
因玉蜀黍植物,该抗性由改变的5-烯醇丙酮莽草酸-3-磷酸(epsp)合成酶基因赋予。
[0256]
编码对磷酰基化合物(如草铵膦或草丁膦、以及吡啶氧丙酸或苯氧丙酸以及环己酮)的抗性的多核苷酸也是适合的。参见欧洲专利申请号0 242 246。还参见美国专利号5,879,903、5,276,268和5,561,236。
[0257]
其他适合的多核苷酸包括编码对抑制光合作用的除草剂(如三嗪和苯基氰(腈水解酶))的抗性的那些。参见美国专利号4,810,648。编码用于除草剂抗性的另外的适合的多核苷酸包括编码对2,2-二氯丙酸、烯禾啶、吡氟氯禾灵、咪唑啉酮除草剂、磺酰脲除草剂、三唑并嘧啶除草剂、均三嗪除草剂以及溴草腈的抗性的那些。同样适合的是赋予对原卟啉原氧化酶的抗性或者提供增强的对植物疾病的抗性、增强的对不利环境条件(非生物胁迫)的耐受性(这些条件包括但不限于干旱、极冷、极热、或极端的土壤盐度或极端的酸度或碱度)、以及在植物构造或发育中的改变(包括发育时间方面的变化)的多核苷酸。参见例如,美国专利公开号2001/0016956和美国专利号6,084,155。
[0258]
另外的适合的多核苷酸包括对杀有害生物(例如杀昆虫)多肽进行编码的那些。这些多肽可以按足以控制例如昆虫有害生物的量(即,昆虫控制量)进行生产。在实施例中,所述多肽是具有鳞翅目活性的、鞘翅目活性的、半翅目活性的和/或双翅目活性的多肽,或其任何组合。应当认识到在植物中对控制昆虫或其他有害生物必要的杀有害生物多肽的生产量可以变化,这取决于栽培品种、有害生物的类型、环境因素等。有用于另外的昆虫或有害生物抗性的多核苷酸包括例如编码芽孢杆菌属(bacillus)生物中鉴定到的毒素的那些。已经克隆了包含编码来自几个亚种的苏云金芽孢杆菌(bt)cry蛋白的核苷酸序列的多核苷酸,并且已经发现这些重组克隆对鳞翅目、双翅目和鞘翅目昆虫幼虫是有毒的。此类bt杀昆虫蛋白的实例包括如下cry蛋白,如cry1aa、cry1ab、cry1ac、cry1b、cry1c、cry1d、cry1ea、cry1fa、cry3a、cry9a、cry9b、cry9c等,连同营养期杀昆虫蛋白如vip1、vip2、vip3等,以及前述bt杀昆虫蛋白的任何组合。bt来源的蛋白的完整清单可以在万维网在苏塞克斯大学(university of sussex)维护的苏云金芽孢杆菌毒素命名法数据库中找到(还参见,crickmore等人,(1998),microbiol.mol.biol.rev.[微生物分子生物学综述]62:807-813)。
[0259]
在实施例中,另外的多肽是衍生自非bt来源的杀昆虫多肽,包括但不限于:α淀粉酶、过氧化物酶、胆固醇氧化酶、马铃薯糖蛋白、蛋白酶、蛋白酶抑制剂、脲酶、α-淀粉酶抑制剂、成孔蛋白、几丁质酶、凝集素、工程化抗体或抗体片段、蜡样芽孢杆菌杀昆虫蛋白、致病杆菌属物种(如嗜线虫致病杆菌(x.nematophila)或伯氏致病杆菌(x.bovienii))杀昆虫蛋白、发光杆菌属物种(如发光杆菌(p.luminescens)或p.asymobiotica)杀昆虫蛋白、短芽孢杆菌属物种(如侧孢短芽孢杆菌(b.laterosporous))杀昆虫蛋白、赖氨酸芽孢杆菌属物种(lysinibacillus spp.)(如球形赖氨酸芽孢杆菌(l.sphearicus))杀昆虫蛋白、色杆菌属物种(如c.subtsugae或c.piscinae)杀昆虫蛋白、耶尔森菌属物种(如耶尔森噬虫霉(y.entomophaga))杀昆虫蛋白、类芽孢杆菌属物种(如丙型类芽孢杆菌属(p.propylaea))杀昆虫蛋白、梭菌属物种(如双酶梭菌(c.bifermentans))杀昆虫蛋白、假单胞菌属物种(pseudomonas spp.)(如荧光假单胞菌(p.fluorescens)和木质素。
[0260]
适合在植物中生产的多肽进一步包括改善或通过其他方式促进收获的植物或植物部分转化成为商业上有用的产品(包括例如增加的或改变的碳水化合物含量或分布、改
善的发酵特性、增加的油含量、增加的蛋白质含量、改善的消化率、以及增加的营养成分含量(例如,增加的植物甾醇含量、增加的生育酚含量、增加的甾烷醇含量或增加的维生素含量))的那些。感兴趣的多肽还包括,例如在收获的作物中导致或促成不需要的成分(例如植酸、或降解糖的酶类)的含量降低的那些。“导致(resulting in)”或“促成(contributing to)”是指这种感兴趣的多肽可以直接或间接地促成感兴趣的性状的存在(例如,通过异源纤维素酶的使用来增加纤维素降解)。
[0261]
在一些实施例中,多肽促成了食品或饲料的改进的可消化度。木聚糖酶是半纤维素分解酶,这些酶改进了植物细胞壁的分解,这导致动物更好地利用这些植物营养素。这导致了改进的生长率和饲料转化。同样,可以减小含有木聚糖的饲料的粘度。在植物细胞内异源产生木聚糖酶也可以促进木质纤维素转化成工业加工中的可发酵糖。
[0262]
已经鉴定和表征了来自真菌和细菌微生物的许多木聚糖酶(参见,例如,美国专利号5,437,992;coughlin等人(1993)“proceedings of the second tricel symposium on trichoderma reesei cellulases and other hydrolases[关于里氏木霉纤维素酶和其他水解酶的第二届tricel研讨会的议定书]”espoo;souminen和reinikainen,编著(1993)foundation for biotechnical and industrial fermentation research[生物技术和工业发酵研究的基础]8:125-135;美国专利公开号2005/0208178;以及pct公开号wo 03/16654)。特别地,已经在里氏木霉中鉴定了三种特定的木聚糖酶(xyl-i、xyl-ii和xyl-iii)(tenkanen等人(1992)enzyme microb.technol.[酶与微生物技术]14:566;torronen等人(1992)bio/technology[生物/技术]10:1461;以及xu等人(1998)appl.microbiol.biotechnol.[应用微生物学和生物技术]49:718)。
[0263]
在其他实施例中,对于本发明有用的多肽可以是多糖降解酶。产生这样一种酶的本发明的植物对于产生例如用于生物加工的发酵原料会是有用的。在一些实施例中、可用于发酵过程的酶包括α淀粉酶、蛋白酶、支链淀粉酶、异淀粉酶、纤维素酶、半纤维素酶、木聚糖酶、环糊精糖基转移酶、脂肪酶、植酸酶、漆酶、氧化酶、酯酶、角质酶、颗粒淀粉水解酶以及其他葡糖淀粉酶。
[0264]
多糖降解酶包括:淀粉降解酶如α-淀粉酶(ec 3.2.1.1)、葡萄糖醛酸酶(e.c.3.2.1.131);外切-1,4-α-d葡聚糖酶,如淀粉葡糖苷酶和葡糖淀粉酶(ec 3.2.1.3)、β-淀粉酶(ec 3.2.1.2)、α-葡糖苷酶(ec3.2.1.20)以及其他外切淀粉酶;淀粉脱支酶,例如a)异淀粉酶(ec3.2.1.68)、支链淀粉酶(ec 3.2.1.41)等;b)纤维素酶如外切-1,4-3-纤维二糖水解酶(ec 3.2.1.91)、外切-1,3-β-d-葡聚糖酶(ec 3.2.1.39)、β-葡糖苷酶(ec 3.2.1.21);c)l-阿拉伯糖酶,例如内切-1,5-α-l-阿拉伯糖酶(ec 3.2.1.99)、α-阿拉伯糖苷酶(ec 3.2.1.55)等;d)半乳聚糖酶,例如内切-1,4-β-d-半乳聚糖酶(ec 3.2.1.89)、内切-1,3-β-d-半乳聚糖酶(ec 3.2.1.90)、α-半乳糖苷酶(ec 3.2.1.22)、β-半乳糖苷酶(ec 3.2.1.23)等;e)甘露聚糖酶,例如内切-1,4-β-d-甘露聚糖酶(ec 3.2.1.78)、β-甘露糖苷酶(ec 3.2.1.25)、α-甘露糖苷酶(ec3.2.1.24)等;f)木聚糖酶,例如内切-1,4-β-木聚糖酶(ec 3.2.1.8)、β-d-木糖苷酶(ec 3.2.1.37)、1,3-β-d-木聚糖酶等;和g)其他酶,例如α-l-岩藻糖苷酶(ec 3.2.1.51)、α-l-鼠李糖苷酶(ec 3.2.1.40)、果聚糖酶(ec 3.2.1.65)、菊粉酶(ec 3.2.1.7)等。在一个实施例中,所述α-淀粉酶是描述于美国专利号8,093,453中的合成α-淀粉酶amy797e。
[0265]
可以与本发明一起使用的另外的酶包括蛋白酶,如真菌和细菌蛋白酶。真菌蛋白酶包括但不限于从曲霉属(aspergillus)、木霉属(trichoderma)、毛霉属(mucor)和根霉属(rhizopus),如黑曲霉(a.niger)、泡盛曲霉(a.awamori)、米曲霉(a.oryzae)和米黑毛霉(m.miehei)获得的那些。在一些实施例中,本发明的多肽可以是纤维二糖水解酶(cbh)(ec 3.2.1.91)。在一个实施例中,该纤维二糖水解酶可以是cbh1或cbh2。
[0266]
可用于本发明的其他酶包括但不限于半纤维素酶,例如甘露糖酶和阿拉伯呋喃糖苷酶(ec 3.2.1.55);木质素酶;脂肪酶(例如,e.c.3.1.1.3)、葡萄糖氧化酶、果胶酶、木聚糖酶、转葡糖苷酶、α1,6葡糖苷酶(例如,e.c.3.2.1.20);酯酶,例如阿魏酸酯酶(ec 3.1.1.73)和乙酰木聚糖酯酶(ec 3.1.1.72);和角质酶(例如,e.c.3.1.1.74)。
[0267]
与本发明一起使用的双链rna(dsrna)分子包括但不限于抑制靶标有害生物(例如昆虫)基因的那些。在实施例中,所述dsrna靶向鳞翅目、鞘翅目、半翅目或双翅目昆虫有害生物中的基因,或前述的任何组合。如在此使用的词语“基因抑制”当一起考虑时旨在是指用于减少作为基因转录为mrna和该mrna的后续翻译的结果而产生的蛋白质的水平的任何熟知的方法。基因抑制还旨在意指减少从基因或编码序列表达蛋白质,包括转录后基因抑制和转录抑制。转录后基因抑制由从靶向抑制的基因或编码序列转录的mrna的全部或其一部分与用于抑制的相应双链rna之间的同源性介导,并且是指在细胞中可供被核糖体结合使用的可获得的mrna的量的实质且可测量的减少。经转录的rna可以处于正义方向而发挥作用,称为共抑制,处于反义方向而发挥作用,称为反义抑制,或在两个方向上产生dsrna而发挥作用,称为rna干扰(rnai)。转录抑制由细胞中存在作为基因抑制剂的与启动子dna序列或其补体展示出实质序列同一性而发挥作用的dsrna介导,称为启动子反式抑制。针对与性状相关联的天然植物基因,基因抑制可以是有效的,例如,以提供具有减少水平的由所述天然基因编码的蛋白质或具有增强或减少水平的受影响代谢物的植物。针对植物有害生物中的靶基因,基因抑制也可以是有效的,这些有害生物可以摄取或接触含有基因抑制剂的植物材料,这些基因抑制剂专门设计用于阻抑或抑制一种或多种同源或互补序列在该有害生物的细胞中的表达。靶向抑制的此类基因可以编码必需蛋白质,其预测功能选自下组,该组由以下组成:肌肉形成、保幼激素形成、保幼激素调节、离子调节和转运、消化酶合成、细胞膜电势的维持、氨基酸生物合成、氨基酸降解、精子形成、外激素(pheromone)合成、外激素感测、触角形成、翼形成、腿形成、发育和分化、卵形成、幼虫成熟、消化酶形成、血淋巴合成、血淋巴维持、神经传递、细胞分裂、能量代谢、呼吸以及凋亡。
[0268]
在一些实施例中,本发明的多核苷酸可进一步包含运载体,基本上由其组成或由其组成。在其他实施例中,本发明的多核苷酸和表达盒包含在运载体中。用于在植物和其他生物的转化中使用的运载体在本领域是熟知的。一般类别的运载体的非限制性实例包括质粒、噬菌体运载体、噬菌粒运载体、粘粒运载体、福斯质粒、噬菌体、人工染色体或病毒运载体。在实施例中,所述运载体是植物运载体,例如用于植物转化。在实施例中,所述运载体是细菌运载体,例如用于细菌转化。适用于植物、细菌和其他生物的运载体是本领域已知的。
[0269]
本发明还涵盖了如下转基因非人类宿主细胞,所述转基因非人类宿主细胞包含本发明的多核苷酸、核酸分子、表达盒、运载体或多肽。转基因非人类宿主细胞可以包括但不限于植物细胞(包括单子叶植物细胞和/或双子叶植物细胞)、酵母细胞、细菌细胞或昆虫细胞。因此,在一些实施例中,本发明提供了选自以下属的细菌细胞:芽孢杆菌属(bacillus)、
短芽孢杆菌属(brevibacillus)、梭菌属(clostridium)、致病杆菌属(xenorhabdus)、发光杆菌属(photorhabdus)、巴斯德氏芽菌属(pasteuria)、埃希氏菌属(escherichia)、假单胞菌属(pseudomonas)、欧文氏菌属(erwinia)、沙雷氏菌属(serratia)、克雷伯菌属(klebsiella)、沙门氏菌属(salmonella)、巴氏杆菌属(pasteurella)、黄单胞菌属(xanthomonas)、链霉菌属(streptomyces)、根瘤菌属(rhizobium)、红假单胞菌属(rhodopseudomonas)、嗜甲基菌属(methylophilius)、农杆菌属(agrobacterium)、醋杆菌属(acetobacter)、乳杆菌属(lactobacillus)、节杆菌属(arthrobacter)、固氮菌属(azotobacter)、明串珠菌属(leuconostoc)或产碱杆菌属(alcaligenes)。因此,例如,作为生物昆虫控制剂,本发明的txp40杀昆虫蛋白可以通过在细菌细胞中表达编码本发明的杀昆虫蛋白的多核苷酸而产生。例如,在一些实施例中,提供了包含编码本发明的txp40杀昆虫蛋白的多核苷酸的苏云金芽孢杆菌细胞。
[0270]
在一些实施例中,所述转基因植物细胞是双子叶植物细胞或单子叶植物细胞。在另外的实施例中,所述双子叶植物细胞是大豆细胞、向日葵细胞、番茄细胞、芸苔属作物细胞、棉花细胞、糖用甜菜细胞或烟草细胞。在另外的实施例中,所述单子叶植物细胞是大麦细胞、玉蜀黍细胞、燕麦细胞、水稻细胞、高粱细胞、甘蔗细胞或小麦细胞。在实施例中,本发明提供了多个双子叶植物细胞或单子叶植物细胞,这些细胞包含表达本发明的嵌合杀昆虫蛋白的多核苷酸。在其他实施例中,将该多个细胞并列以形成质外体并且使其在自然光照中生长。在其他实施例中,所述转基因植物细胞不能再生全株植物。
[0271]
在本发明的一些实施例中,在高等生物(例如,植物)中表达本发明的txp40杀昆虫蛋白。在一些实施例中,植物可被灰翅夜蛾属有害生物侵染,特别是可被秋粘虫侵染。在其他实施例中,可被秋粘虫侵染的植物是玉米植物。在这种情况下,表达有效量的txp40杀昆虫蛋白的转基因植物保护其自身至少免受灰翅夜蛾有害生物,特别是秋粘虫的伤害。当昆虫开始摄食这样一种转基因植物时,它摄取了所表达的txp40杀昆虫蛋白。这可以妨碍昆虫进一步咬食植物组织或者甚至可以伤害或杀死昆虫。在一些实施例中,本发明的多核苷酸被插入表达盒中,然后所述表达盒被稳定地整合到植物的基因组中。在其他实施例中,该多核苷酸被包括在非致病性自我复制病毒中。
[0272]
在本发明的一些实施例中,包含本发明的核酸分子或多肽的转基因植物细胞是植物部分、植物器官或植物培养物(各自如在此描述的)的细胞,包括但不限于根、叶、种子、花、果实、花粉细胞、器官或植物培养物等,或愈伤组织细胞或培养物。
[0273]
根据本发明的转基因植物或植物细胞可以是单子叶植物或双子叶植物或植物细胞,并且包括但不限于玉米(玉蜀黍)、大豆、水稻、小麦、大麦、黑麦、燕麦、高粱、粟、向日葵、红花、糖用甜菜、棉花、甘蔗、油菜、苜蓿、烟草、花生、蔬菜(包括甘薯、豆类、豌豆、菊苣、莴苣、甘蓝、花椰菜、西兰花、芜菁、胡萝卜、茄子、黄瓜、萝卜、菠菜、马铃薯、番茄、芦笋、洋葱、大蒜、瓜类、胡椒、芹菜、南瓜、西葫芦、绿皮西葫芦等)、水果(包括苹果、梨、榅桲、李子、樱桃、桃、蜜桃、杏、草莓、葡萄、覆盆子、黑莓、菠萝、鳄梨、番木瓜、芒果、香蕉等)和特种植物或植物细胞(如拟南芥),或者木本植物或植物细胞(如针叶树和/或落叶树)。在实施例中,本发明的植物或植物细胞是作物植物或植物细胞,如玉蜀黍、高粱、小麦、向日葵、番茄、十字花科植物、胡椒、马铃薯、棉花、水稻、大豆、糖用甜菜、甘蔗、烟草、大麦、油菜植物或植物细胞等。
[0274]
本发明进一步提供了本发明的转基因植物的一部分。任选地,所述植物部分包含本发明的嵌合杀昆虫蛋白和/或编码其的核酸。
[0275]
本发明进一步提供了本发明的转基因植物的种子或产生本发明的转基因植物的种子。任选地,所述种子包含本发明的嵌合杀昆虫蛋白和/或编码其的核酸。
[0276]
本发明的另外的实施例包括从本发明的转基因植物、植物部分或种子产生的收获产物,连同从所述收获产物产生的加工产物。收获产物可以是如在此描述的全株或任何植物部分。因此,在一些实施例中,收获产物的非限制性实例包括种子、果实、花或其部分(例如,花药、柱头等)、叶、茎等。在其他实施例中,加工产物包括但不限于从收获的本发明的种子或其他植物部分产生的细粉、粗粉、油、淀粉、谷物等。任选地,收获的产物或加工的产物包含本发明的嵌合杀昆虫蛋白和/或编码其的核酸。
[0277]
在其他实施例中,本发明提供了来自本发明的转基因植物、植物部分的提取物,任选地其中所述提取物包含本发明的嵌合杀昆虫蛋白和/或编码其的核酸。来自植物或植物部分的提取物可以根据本领域公知的程序制备(参见,de la torre等人,food,agric.environ.[食品、农业和环境杂志]2(1):84-89(2004);guidet,nucleic acids res.[核酸研究]22(9):1772-1773(1994);lipton等人,food agric.immun.[食品、农业和免疫杂志]12:153-164(2000))。
[0278]
所述嵌合杀昆虫蛋白可以作为昆虫控制剂在植物部分、植物细胞、植物器官、种子、收获产物、加工产物或提取物等中起作用。换言之,所述嵌合杀昆虫蛋白可以继续执行它在转基因植物中具有的杀昆虫功能。所述核酸可以起到表达所述嵌合杀昆虫蛋白的作用。作为编码本发明的杀昆虫蛋白的替代方案,所述核酸可用于鉴定本发明的转基因植物部分、植物细胞、植物器官、种子、收获产物、加工产物或提取物。
[0279]
在实施例中,本发明的转基因植物、植物部分、植物细胞、植物器官或种子对于本发明的多核苷酸或表达盒是半合子的。在实施例中,本发明的转基因植物、植物部分、植物细胞、植物器官或种子对于本发明的多核苷酸或表达盒是纯合的。
[0280]
在实施例中,如与不包含编码本发明杀昆虫蛋白的核酸的适合的对照相比,转基因植物、植物部分、植物细胞、植物器官、种子、收获产物、加工产物或提取物对一种或多种昆虫有害生物(例如鳞翅目有害生物,如秋黏虫)具有增加的抗性。
[0281]
用于转化植物的程序在本领域中是熟知且常规的并且普遍描述于文献中。用于植物转化的方法的非限制性实例包括通过以下的转化:细菌介导的核酸递送(例如,经由农杆菌)、病毒介导的核酸递送、碳化硅或核酸须晶介导的核酸递送、脂质体介导的核酸递送、微注射、微粒轰击、磷酸钙介导的转化、环糊精介导的转化、电穿孔、纳米粒子介导的转化、超声处理、渗入、peg介导的核酸吸收、连同使得核酸引入到植物细胞中的任何其他电学的、化学的、物理的(机械的)或生物的机制,包括其任何组合。对于本领域已知的不同植物转化方法的一般指导包括以下文献:miki等人(“procedures for introducing foreign dna into plants”in methods in plant molecular biology and biotechnology[植物分子生物学与生物技术方法中的“用于将外来dna引入植物中的程序”],glick,b.r.和thompson,j.e.编辑(crc press,inc.[crc出版公司],boca raton[波卡拉顿],1993),第67-88页)和rakowoczy-trojanowska(cell.mol.biol.lett.[细胞与分子生物学快报]7:849-858(2002))。
[0282]
对于农杆菌介导的转化,二元运载体或携带至少一个t-dna边界序列的运载体通常是适合的,而对于直接基因转移(例如,微粒轰击等),任何运载体都是适合的,并且可以使用仅含有感兴趣的构建体的线性dna。在直接基因转移的情况下,可以使用以单个dna物种的转化或共转化(schocher等人,biotechnology[生物技术]4:1093-1096(1986))。对于直接基因转移以及农杆菌介导的转移二者,转化通常(但不是必需的)用如下选择性标记进行,所述选择性标记可以是正向选择(例如,磷甘露糖异构酶),提供对抗生素(例如,卡那霉素、潮霉素或甲氨蝶呤)或除草剂(例如,草甘膦或草丁膦)的抗性。然而,选择性标记的选择对于本发明并不是至关重要的。
[0283]
农杆菌介导的转化是用于转化植物的常用方法,因为它的高转化效率以及因为它与许多不同物种的广泛实用性。农杆菌介导的转化典型地涉及将携带感兴趣的外来dna的二元运载体转移至适当的农杆菌菌株,这可能取决于由宿主农杆菌菌株在共同存在的ti质粒上或在染色体上携带的vir基因的互补物(uknes等人(1993)plant cell[植物细胞]5:159-169)。将该重组二元运载体转移至农杆菌可以使用携带该重组二元运载体的大肠杆菌、辅助大肠杆菌菌株(该辅助菌株携带能够将该重组二元运载体移动到靶标农杆菌菌株中的质粒)通过三亲本交配程序实现。可替代地,可以通过核酸转化将所述重组二元运载体转移至农杆菌中(和willmitzer,(1988)nucleic acids res.[核酸研究]16:9877)。
[0284]
可以使用农杆菌转化双子叶植物以及单子叶植物。用于农杆菌介导的水稻转化的方法包括用于水稻转化的熟知的方法,例如在以下的任一个中描述的那些:欧洲专利申请ep 1198985 a1、aldemita和hodges(planta[植物学]199:612-617,1996);chan等人(plant mol biol[植物分子生物学]22(3):491-506,1993)、hiei等人(plant j[植物杂志]6(2):271-282,1994),将这些披露通过引用结合在此,其引用程度如同完全阐明一样。在玉米转化的情况下,优选方法是如ishida等人(nat.biotechnol[自然生物技术]14(6):745-50,1996)或frame等人(plant physiol[植物生理学]129(1):13-22,2002)中所描述的,将这些披露通过引用结合在此,其引用程度如同完全阐明一样。所述方法通过以下中的实例来进一步描述:b.jenes等人,techniques for gene transfer[基因转移技术],于:transgenic plants[转基因植物],第1卷,engineering and utilization[工程与利用],编著s.d.kung和r.wu,学院出版社(1993)128-143以及potrykus annu.rev.plant physiol.plant molec.biol.[植物生理学与植物分子生物学年度综述]42(1991)205-225)。有待表达的核酸或构建体优选地克隆至适合于转化根癌农杆菌(agrobacterium tumefaciens)的运载体例如pbin19中(bevan等人,nucl.acids res.[核酸研究]12(1984)8711)。然后,能够以已知的方式使用由这种运载体转化的农杆菌来转化植物,如用作模型的植物(如拟南芥)或作物植物(如烟草植物),方法是例如通过将捣碎的叶或切碎的叶浸没于农杆菌溶液中,并且然后将其在适合的培养基中培养。通过根癌农杆菌转化植物例如hagen和willmitzer在nucl.acid res.[核酸研究](1988)16,9877中描述或尤其从f.f.white,vectors for gene transfer in higher plants[在高等植物中基因转移的载体];transgenic plants[转基因植物],第1卷,engineering and utilization[工程与利用],编著s.d.kung和r.wu,学院出版社,1993,第15-38页中已知。
[0285]
通过重组农杆菌进行的植物转化通常涉及该农杆菌与来自该植物的外植体的共
培养,并且遵循本领域熟知的方法。在携带位于这些二元质粒t-dna边界之间的抗生素或除草剂抗性标记的选择培养基上对转化的组织进行再生。
[0286]
如先前所讨论的,另一种用于转化植物、植物部分和植物细胞的方法涉及在植物组织和细胞上推进惰性或生物活性的粒子。参见例如,美国专利号4,945,050、5,036,006和5,100,792。通常,这种方法涉及在有效于穿透该细胞的外表面并提供掺入在其内部中的条件下在植物细胞处推进惰性或生物活性的粒子。当使用惰性粒子时,可以通过用含有目的核酸的载体包被这些粒子而将该载体引入该细胞中。可替代地,一个或多个细胞可以被该载体围绕以使得该载体通过该粒子的激发而被带入该细胞中。也可以将生物活性的粒子(例如,干酵母细胞、干细菌或噬菌体,各自含有一种或多种被试图引入的核酸)推进到植物组织中。
[0287]
在其他实施例中,本发明的多核苷酸可以被直接转化进质体基因组中。质体转化的主要优点在于质体通常能够表达细菌基因而无需实质性的修饰,而且质体能够在单个启动子的控制下表达多个开放阅读框。在美国专利号5,451,513、5,545,817和5,545,818中,在pct申请号wo 95/16783中,以及在mcbride等人(1994)proc.nati.acad.sci.usa[美国国家科学院院刊]91,7301-7305中广泛描述了质体转化技术。基本的叶绿体转化技术涉及将位于选择性标记侧翼的经克隆的质体dna区连同感兴趣的基因一起引入适合的靶组织中,这是例如使用生物射弹(biolistic)或原生质体转化(例如,氯化钙或peg介导的转化)来进行的。这些1至1.5kb的侧翼区(被命名为靶向序列)促进了与质体基因组的同源重组,并且因而允许置换或修饰原质体(plastome)的特定区域。最初,可以将叶绿体16s rrna和rps12基因(赋予针对大观霉素或链霉素的抗性)的点突变用作供转化用的选择性标记(svab,z.、hajdukiewicz,p.和maliga,p.,(1990),proc.natl.acad.sci.usa[美国国家科学院院刊]87,8526-8530);staub,j.m.和maliga,p.,(1992),plant cell[植物细胞]4,39-45)。在这些标记之间克隆位点的存在允许建立质体靶向运载体用于外来基因的引入(staub,j.m.和maliga,p.(1993)embo j.[欧洲分子生物学杂志]12,601-606)。转化频率的实质性增加可以是通过用显性选择性标记(细菌aada基因,其编码了大观霉素-解毒酶氨基糖苷-3'-腺苷酰转移酶)来置换这些隐性rrna或r-蛋白抗生素抗性基因(svab,z.和maliga,p.(1993)proc.natl.acad.sci.usa[美国国家科学院院刊]90,913-917)。先前,这种标记已经被成功地用于莱茵衣藻(chlamydomonas reinhardtii)这种绿藻的质体基因组的高频率转化(goldschmidt-clermont,m.(1991)nucl.acids res.[核酸研究]19:4083-4089)。有用于质体转化的其他选择性标记在本领域是已知的,并且被包括在本发明的范围之内。典型地,转化之后需要大约15-20个细胞分裂循环以便达到同质状态。质体表达(其中基因通过同源重组被插入到在每个植物细胞中存在的所有数千个环状质体基因组的拷贝中)利用了超过核表达的基因的庞大的拷贝数目的优点,以便允许能够很容易超过总的可溶性植物蛋白的10的表达水平。在一个实施例中,可以将本发明的多核苷酸插入质体靶向运载体中并转化进所希望的植物宿主的质体基因组中。因此,可以获得与含有本发明的核苷酸序列的质体基因组同型的植物,这些植物能够高表达该多核苷酸。
[0288]
选择转化的转基因植物、植物细胞或植物组织培养物的方法在本领域中是常规的,并且可以用于在此提供的本发明的方法中。例如,本发明的重组运载体还可以包括包含用于选择性标记的核苷酸序列的表达盒,该选择性标记可以用于选择转化的植物、植物部
分或植物细胞。
[0289]
此外,如本领域中所熟知的,完整的转基因植物可以使用多种已知技术中的任何技术从转化的植物细胞、植物组织培养物或培养的原生质体再生而来。从植物细胞、植物组织培养物或培养的原生质体再生植物描述于例如evans等人(handbook of plant cell cultures[植物细胞培养手册],第1卷,macmilan publishing co.[麦克米兰出版公司]纽约(1983));和vasil i.r.(编著)(cell culture and somatic cell genetics of plants[植物的细胞培养和体细胞遗传学],学院出版社,奥兰多,第i卷(1984)和第ii卷(1986))中。
[0290]
另外,工程化入以上所述的本发明的转基因种子和植物、植物部分或植物细胞中的遗传特性可以通过有性生殖或营养生长来传递,并且因此可以在子代植物中维持并传代。通常,维持和传代利用了被开发以适合特定目的(如收获、播种或耕作)的已知农业方法。
[0291]
因此,可以按本领域熟知的任何数目的方法(如上所述的)将多核苷酸引入该植物、植物部分或植物细胞中。因此,没有依赖用于将一种或多种多核苷酸引入植物中的特定方法,相反可以使用允许将该一种或多种多核苷酸稳定地整合到该植物的基因组中的任何方法。在有待引入一种以上多核苷酸的情况下,这些对应的多核苷酸可以作为单一核酸分子的一部分、或者作为分开的核酸分子而进行组装,并且可以位于相同的或不同的核酸分子上。因此,可以在单个转化事件中、在分开的转化事件中、或者例如作为育种方案的一部分在植物中,将这些多核苷酸引入感兴趣的细胞中。
[0292]
一旦所希望的多核苷酸已经被转化进特定的植物物种中,便可以使用传统的育种技术将其在该物种中繁殖或将其转移到相同物种的其他品种中,特别是包括商业品种。
[0293]
在一些实施例中,本发明提供了杀昆虫组合物,所述杀昆虫组合物包含农业上可接受的载体中的本发明的嵌合杀昆虫蛋白。如在此使用的“农业上可接受的载体”可以包括与活性蛋白组合以促进它应用至植物或其部分或者在植物或其部分中应用的天然或合成的有机或无机材料。农业上可接受的载体的实例包括但不限于粉剂、尘剂、丸剂、颗粒剂、喷雾剂、乳剂、胶体以及溶液。农业上可接受的载体进一步包括但不限于可用于农业配制品中的惰性组分、分散剂、表面活性剂、佐剂、增粘剂、粘着剂、粘合剂或其组合。此类组合物可以按使杀昆虫蛋白或其他有害生物控制剂与这些有害生物接触的任何方式施用。因此,可以将这些组合物施用于植物或植物部分的表面,包括种子、叶、花、茎、块茎、根等。在其他实施例中,在植物体内产生本发明的杀昆虫蛋白的植物是被表达的杀昆虫蛋白的农业上可接受的载体。在实施例中,本发明的组合物和农业上可接受的载体不包括转基因植物。
[0294]
在另外的实施例中,所述杀昆虫组合物包含细菌细胞或本发明的转基因细菌细胞,其中所述细菌细胞或转基因细菌细胞产生本发明的杀昆虫蛋白。这样一种杀昆虫组合物可以通过脱水、冷冻干燥、均化、提取、过滤、离心、沉降或浓缩苏云金芽孢杆菌(bt)的培养物(包括转基因bt培养物)而制备。在实施例中,本发明的组合物可包含按重量计至少约1、至少约5、至少约10、至少约20、至少约25、至少约30、至少约35、至少约40、至少约50、至少约60、至少约70、至少约80、至少约90、至少约95、至少约97或至少99的本发明的多肽。在另外的实施例中,所述组合物包含按重量计从约1至约99的本发明的杀昆虫蛋白。
[0295]
本发明的杀昆虫蛋白可以与其他有害生物控制剂组合使用,以增加有害生物靶标
谱和/或用于预防或管理昆虫抗性。此外,本发明的杀昆虫蛋白与杀昆虫剂(其具有不同的作用方式或靶向昆虫肠中不同的受体)相组合进行使用对于预防和/或管理昆虫抗性而言具有特定的功用。
[0296]
因此,在一些实施例中,本发明提供了控制一种或多种植物有害生物(例如昆虫有害生物,如鳞翅目昆虫有害生物、鞘翅目昆虫有害生物、半翅目昆虫有害生物和/或双翅目昆虫有害生物)的组合物,其中所述组合物包含第一有害生物控制剂(其是本发明的嵌合杀昆虫蛋白)和至少第二有害生物控制剂(其不同于所述第一有害生物控制剂)。在其他实施例中,该组合物是用于局部施用至植物的配制品。在仍其他实施例中,该组合物是转基因植物。在另外的实施例中,该组合物是局部施用至转基因植物的配制品的组合。在一些实施例中,当所述转基因植物包含所述第二有害生物控制剂时,所述配制品包含所述第一有害生物控制剂(其是本发明的嵌合杀昆虫蛋白)。在其他实施例中,当所述转基因植物包含所述第一有害生物控制剂(其是本发明的嵌合杀昆虫蛋白)时,所述配制品包含所述第二有害生物控制剂。
[0297]
在一些实施例中,所述第二有害生物控制剂可以是以下中的一种或多种:化学杀有害生物剂(如杀昆虫剂)、苏云金芽孢杆菌(bt)杀昆虫蛋白、和/或非bt杀有害生物剂,包括但不限于致病杆菌属杀昆虫蛋白、发光杆菌属杀昆虫蛋白、侧孢短芽孢杆菌(brevibacillus laterosporus)杀昆虫蛋白、球形芽孢杆菌(bacillus sphaericus)杀昆虫蛋白、蛋白酶抑制剂(丝氨酸和半胱氨酸类型两者)、凝集素、α-淀粉酶、过氧化物酶、胆固醇氧化酶或双链rna(dsrna)分子。
[0298]
在其他实施例中,所述第二有害生物控制剂是一种或多种化学杀有害生物剂,其任选地是种子包衣。化学杀有害生物剂的非限制性实例包括拟除虫菊酯、氨基甲酸酯、新烟碱、神经元钠通道阻断剂、杀昆虫大环内酯、γ-氨基丁酸(gaba)拮抗剂、杀昆虫脲以及保幼激素模拟物。在其他实施例中,所述化学杀有害生物剂是以下中的一种或多种:阿巴美丁、乙酰甲胺磷、啶虫脒、磺胺螨酯(amidoflumet)(s-1955)、除虫菌素(avermectin)、印楝素、甲基谷硫磷、联苯菊酯、联苯肼酯(binfenazate)、噻嗪酮、克百威、溴虫腈、定虫隆、毒死蜱、甲基毒死蜱、环虫酰肼、噻虫胺、氟氯氰菊酯、β-氟氯氰菊酯、三氯氟氰菊酯、λ-三氯氟氰菊酯、氯氰菊酯、灭蝇胺、溴氰菊酯、杀螨隆、二嗪磷、除虫脲、乐果、苯虫醚、甲氨基阿维菌素、硫丹、高氰戊菊酯、乙虫腈、苯硫威(fenothicarb)、苯氧威、甲氰菊酯、唑螨酯、氰戊菊酯、氟虫腈、氟啶虫酰胺、氟氰戊菊酯、τ-氟胺氰菊酯、嘧虫胺(ur-50701)、氟虫脲、地虫硫磷、氯虫酰肼、氟铃脲、吡虫啉、茚虫威、异柳磷、氯芬奴隆、马拉硫磷、聚乙醛、甲胺磷、杀扑磷、灭多威、烯虫酯、甲氧氯、久效磷、甲氧虫酰肼、噻虫醛(nithiazin)、双苯氟脲、多氟脲(xde-007)、杀线威、对硫磷、甲基对硫磷、氯菊酯、甲拌磷、伏杀磷、亚胺硫磷、磷胺、抗蚜威、丙溴磷、吡蚜酮、啶虫丙醚、蚊蝇醚、鱼藤酮、多杀菌素、螺甲螨酯(spiromesifin)(bsn 2060)、硫丙磷、虫酰肼、伏虫隆、七氟菊酯、特丁硫磷、杀虫畏、噻虫啉、噻虫嗪、硫双威、杀虫双(thiosultap-sodium)、四溴菊酯、敌百虫和杀铃脲、涕灭威、杀线威、苯线磷、双甲脒、灭螨猛、乙酯杀螨醇、三环锡、三氯杀螨醇、除螨灵、依杀螨、喹螨醚、苯丁锡、甲氰菊酯、唑螨酯、噻螨酮、克螨特、哒螨灵以及吡螨胺。在仍其他实施例中,所述化学杀有害生物剂选自以下中的一种或多种:氯氰菊酯、三氯氟氰菊酯、氟氯氰菊酯和β-氟氯氰菊酯、高氰戊菊酯、氰戊菊酯、四溴菊酯、苯硫威、灭多威、杀线威、硫双威、噻虫胺、吡虫啉、噻虫啉、茚虫威、多杀菌
素、阿巴美丁、除虫菌素(avermectin)、甲氨基阿维菌素、硫丹、乙虫腈、氟虫腈、氟虫脲、杀铃脲、苯虫醚、蚊蝇醚、吡蚜酮以及双甲脒。
[0299]
在另外的实施例中,所述第二有害生物控制剂可以是任何数目的苏云金芽孢杆菌杀昆虫蛋白中的一种或多种,包括但不限于cry蛋白、营养期杀昆虫蛋白(vip)以及任何前述杀昆虫蛋白的杀昆虫嵌合体。在其他实施例中,第二有害生物控制剂是选自以下的cry蛋白:cry1aa、cry1ab、cry1ac、cry1ad、cry1ae、cry1af、cry1ag、cry1ah、cry1ai、cry1aj、cry1ba、cry1bb、cry1bc、cry1bd、cry1be、cry1bf、cry1bg、cry1bh、cry1bi、cry1ca、cry1cb、cry1da、cry1db、cry1dc、cry1dd、cry1ea、cry1eb、cry1fa、cry1fb、cry1ga、cry1gb、cry1gc、cry1ha、cry1hb、cry1hc、cry1ia、cry1ib、cry1ic、cry1id、cry1ie、cry1if、cry1ig、cry1ja、cry1jb、cry1jc、cry1jd、cry1ka、cry1la、cry1ma、cry1na、cry1nb、cry2aa、cry2ab、cry2ac、cry2ad、cry2ae、cry2af、cry2ag、cry2ah、cry2ai、cry2aj、cry2ak、cry2al、cry2ba、cry3aa、cry3ba、cry3bb、cry3ca、cry4aa、cry4ba、cry4ca、cry4cb、cry4cc、cry5aa、cry5ab、cry5ac、cry5ad、cry5ba、cry5ca、cry5da、cry5ea、cry6aa、cry6ba、cry7aa、cry7ab、cry7ac、cry7ba、cry7bb、cry7ca、cry7cb、cry7da、cry7ea、cry7fa、cry7fb、cry7ga、cry7gb、cry7gc、cry7gd、cry7ha、cry7ia、cry7ja、cry7ka、cry7kb、cry7la、cry8aa、cry8ab、cry8ac、cry8ad、cry8ba、cry8bb、cry8bc、cry8ca、cry8da、cry8db、cry8ea、cry8fa、cry8ga、cry8ha、cry8ia、cry8ib、cry8ja、cry8ka、cry8kb、cry8la、cry8ma、cry8na、cry8pa、cry8qa、cry8ra、cry8sa、cry8ta、cry9aa、cry9ba、cry9bb、cry9ca、cry9da、cry9db、cry9dc、cry9ea、cry9eb、cry9ec、cry9ed、cry9ee、cry9fa、cry9ga、cry10aa、cry11aa、cry11ba、cry11bb、cry12aa、cry13aa、cry14aa、cry14ab、cry15aa、cry16aa、cry17aa、cry18aa、cry18ba、cry18ca、cry19aa、cry19ba、cry19ca、cry20aa、cry20ba、cry21aa、cry21ba、cry21ca、cry21da、cry21ea、cry21fa、cry21ga、cry21ha、cry22aa、cry22ab、cry22ba、cry22bb、cry23aa、cry24aa、cry24ba、cry24ca、cry25aa、cry26aa、cry27aa、cry28aa、cry29aa、cry29ba、cry30aa、cry30ba、cry30ca、cry30da、cry30db、cry30ea、cry30fa、cry30ga、cry31aa、cry31ab、cry31ac、cry31ad、cry32aa、cry32ab、cry32ba、cry32ca、cry32cb、cry32da、cry32ea、cry32eb、cry32fa、cry32ga、cry32ha、cry32hb、cry32ia、cry32ja、cry32ka、cry32la、cry32ma、cry32mb、cry32na、cry32oa、cry32pa、cry32qa、cry32ra、cry32sa、cry32ta、cry32ua、cry33aa、cry34aa、cry34ab、cry34ac、cry34ba、cry35aa、cry35ab、cry35ac、cry35ba、cry36aa、cry37aa、cry38aa、cry39aa、cry40aa、cry40ba、cry40ca、cry40da、cry41aa、cry41ab、cry41ba、cry42aa、cry43aa、cry43ba、cry43ca、cry43cb、cry43cc、cry44aa、cry45aa、cry46aa、cry46ab、cry47aa、cry48aa、cry48ab、cry49aa、cry49ab、cry50aa、cry50ba、cry51aa、cry52aa、cry52ba、cry53aa、cry53ab、cry54aa、cry54ab、cry54ba、cry55aa、cry56aa、cry57aa、cry57ab、cry58aa、cry59aa、cry59ba、cry60aa、cry60ba、cry61aa、cry62aa、cry63aa、cry64aa、cry65aa、cry66aa、cry67aa、cry68aa、cry69aa、cry69ab、cry70aa、cry70ba、cry70bb、cry71aa、cry72aa、cry73aa、或以上任何组合。在实施例中,所述cry蛋白是cry1fa,例如,如由玉蜀黍事件tc1507所代表的。
[0300]
在另外的实施例中,所述第二有害生物控制剂是选自以下的一种或多种vip3营养期杀昆虫蛋白:vip3aa1、vip3aa2、vip3aa3、vip3aa4、vip3aa5、vip3aa6、vip3aa7、vip3aa8、vip3aa9、vip3aa10、vip3aa11、vip3aa12、vip3aa13、vip3aa14、vip3aa15、vip3aa16、
vip3aa17、vip3aa18、vip3aa19、vip3aa20、vip3aa21、vip3aa22、vip3aa2、vip3aa24、vip3aa25、vip3aa26、vip3aa27、vip3aa28、vip3aa29、vip3aa30、vip3aa31、vip3aa32、vip3aa33、vip3aa34、vip3aa35、vip3aa36、vip3aa37、vip3aa38、vip3aa39、vip3aa40、vip3aa41、vip3aa42、vip3aa43、vip3aa44、vip3ab1、vip3ab2、vip3ac1、vip3ad1、vip3ad2、vip3ae1、vip3af1、vip3af2、vip3af3、vip3ag1、vip3ag2、vip3ag3 hm117633、vip3ag4、vip3ag5、vip3ah1、vip3ba1、vip3ba2、vip3bb1、vip3bb2、vip3bb3,或前述的任何组合。在在实施例中,所述vip3蛋白是vip3aa(美国专利号6,137,033),例如,如由玉米事件mir162(美国专利号8,232,456;美国专利号8,455,720;和美国专利号8,618,272)所代表的。
[0301]
在实施例中,所述第二有害生物控制剂可以衍生自除苏云金芽孢杆菌外的来源。例如,所述第二有害生物控制剂可以是α淀粉酶、过氧化物酶、胆固醇氧化酶、马铃薯糖蛋白、蛋白酶、蛋白酶抑制剂、脲酶、α-淀粉酶抑制剂、成孔蛋白、几丁质酶、凝集素、工程化抗体或抗体片段、蜡样芽孢杆菌杀昆虫蛋白、致病杆菌属物种(如嗜线虫致病杆菌(x.nematophila)或伯氏致病杆菌(x.bovienii))杀昆虫蛋白、发光杆菌属物种(如发光杆菌(p.luminescens)或p.asymobiotica)杀昆虫蛋白、短芽胞杆菌属物种(如侧孢短芽孢杆菌(b.laterosporous))杀昆虫蛋白、赖氨酸芽胞杆菌属物种(lysinibacillus spp.)(如球形赖氨酸芽孢杆菌(l.sphearicus))杀昆虫蛋白、色杆菌属物种(如c.subtsugae或c.piscinae)杀昆虫蛋白、耶尔森菌属物种(如耶尔森噬虫霉(y.entomophaga))杀昆虫蛋白、类芽孢杆菌属物种(如丙型类芽孢杆菌属(p.propylaea))杀昆虫蛋白、梭菌属物种(如双酶梭菌(c.bifermentans))杀昆虫蛋白、假单胞菌属物种(pseudomonas spp.)(如荧光假单胞菌(p.fluorescens)杀昆虫蛋白和木质素。在其他实施例中,所述第二试剂可以是至少一种衍生自杀昆虫毒素复合物(tc)(所述复合物来自发光杆菌属、异短杆菌属(xenorhabus)、沙雷氏菌属或耶尔森菌属)的杀昆虫蛋白。在其他实施例中,杀昆虫蛋白可以是来源于杀昆虫细菌如发光杆菌属物种的adp-核糖基转移酶。在其他实施例中,杀昆虫蛋白可以是vip蛋白,如来自蜡样芽孢杆菌的vip1和/或vip2。在仍其他实施例中,所述杀昆虫蛋白可以是衍生自杀昆虫细菌(如来自侧孢短芽孢杆菌的isp1a和isp2a或来自球形芽孢杆菌的bina和binb)的二元毒素。在仍其他实施例中,所述杀昆虫蛋白可以被工程化或可以是前述杀昆虫蛋白中的任一个的杂交体或嵌合体。
[0302]
在一些实施例中,所述第二杀有害生物剂可以是非蛋白质的(例如干扰rna分子,如dsrna),其可以被转基因表达或作为组合物的一部分应用(例如,使用局部方法)。干扰rna分子典型地包含针对靶基因的至少一种rna片段、间隔序列、和与第一rna片段互补的第二rna片段,从而可以形成双链rna结构。当生物识别双链rna(dsrna)分子并水解它们时,rna干扰(rnai)发生。所得的水解产物是约19-24个核苷酸长度的小rna片段,这些小rna片段被称为小干扰rna(sirna)。然后这些sirna扩散或被携带至整个生物,包括越过细胞膜,在那里它们与mrna(或其他的rna)杂交并且导致rna的水解。干扰rna由rna干扰沉默复合体(risc)识别,rna的效应链(或“引导链”)位于所述复合体中。此引导链充当双链体序列的识别和破坏模板。每次sirna与其互补rna靶标杂交,此过程都被重复,有效防止那些mrna被翻译,并且因此“沉默”mrna自其中转录的特异性基因的表达。本领域已知干扰rna可用于昆虫控制(参见例如,公开物wo 2013/192256,其通过引用并入本文)。设计用于昆虫控制的干扰rna产生非天然存在的双链rna,其利用昆虫中的天然rnai途径来触发靶基因的下调,这可
能导致停止摄食和/或生长并可能导致昆虫有害生物死亡。干扰rna分子可赋予针对与本发明的蛋白质相同的靶标有害生物的昆虫抗性或可靶向不同的有害生物。靶标昆虫植物有害生物可以通过咀嚼、吸吮或刺穿来摄食。本领域已知干扰rna可用于昆虫控制。在实施例中,可用于昆虫控制的dsrna描述于2016年8月5日提交的美国临时申请号62/371,259、62/371,261或62/371,262中。在实施例中,可用于昆虫控制的dsrna描述于美国专利号9,238,8223、9,340,797或8,946,510中。在实施例中,可用于昆虫控制的dsrna描述于美国专利申请号12/868,994、13/831,230、14/207,313或14/207318中。在其他实施例中,干扰rna可赋予针对非昆虫植物有害生物(如线虫有害生物或病毒有害生物)的抗性。
[0303]
在仍另外的实施例中,在转基因植物中共表达所述第一昆虫控制剂(其是本发明的嵌合杀昆虫蛋白)和所述第二有害生物控制剂。可以通过将植物遗传工程化以含有并表达编码昆虫控制剂的核酸序列来实现一种以上杀有害生物成分在相同转基因植物中的共表达。例如,一种以上杀有害生物剂在相同转基因植物中的共表达可以通过制备单一的重组运载体(其在分子叠加(molecular stack)中包含一种以上杀有害生物剂的编码序列)并对植物进行遗传工程化以在所述转基因植物中含有并表达所有的杀有害生物剂而实现。此类分子叠加还可以通过使用微型染色体进行制备,如在例如美国专利7,235,716中所述的。可替代地,可以将植物(亲本1)遗传工程化,用于本发明的嵌合杀昆虫蛋白的表达。可以将第二植物(亲本2)遗传工程化,用于第二有害生物控制剂的表达。通过将亲本1与亲本2杂交,获得了表达来自亲本1和亲本2二者的昆虫控制剂的子代植物。
[0304]
在其他实施例中,本发明提供了对植物有害生物侵染有抗性的叠加性转基因植物,所述植物包含编码用于在靶标有害生物中抑制必需基因的dsrna的核酸(例如dna)序列和编码针对所述靶标有害生物展示出杀昆虫活性的本发明的嵌合杀昆虫蛋白的核酸例如(dna)序列。据报道,dsrna对某些鳞翅目有害生物无效(rajagopol等人2002.j.biol.chem.[生物化学杂志]277:468-494),可能是由于中肠的高ph使dsrna不稳定。因此,在一些靶标有害生物是鳞翅目有害生物的实施例中,本发明的嵌合杀昆虫蛋白起作用以瞬时降低中肠ph,这用于稳定共摄取的dsrna,从而使得所述dsrna有效沉默靶基因。
[0305]
包含和/或表达本发明的杀昆虫蛋白的转基因植物或种子还可以用杀昆虫剂或杀昆虫种子包衣进行处理,如美国专利号5,849,320和5,876,739中所述的。在实施例中,在本发明的杀昆虫剂或杀昆虫种子包衣以及转基因植物或种子针对相同靶标昆虫例如鳞翅目有害生物(例如秋黏虫)具有活性的情况下,所述组合(i)在用于进一步增强本发明的组合物针对所述靶标昆虫的活性的方法中和/或(ii)在用于通过提供针对所述靶标昆虫的又另一种作用机制而预防对本发明的组合物产生抗性的方法中是有用的。因此,在实施例中,本发明提供了增强鳞翅目昆虫群体的控制的方法,所述方法包括提供本发明的转基因植物或种子并且向所述植物或种子施用本发明的杀昆虫剂或杀昆虫种子包衣。
[0306]
即使在杀昆虫剂或杀昆虫种子包衣针对不同昆虫具有活性的情况下,所述杀昆虫剂或杀昆虫种子包衣对于扩展昆虫控制的范围是有用的,例如通过将针对鞘翅目昆虫具有活性的杀昆虫剂或杀昆虫种子包衣添加到本发明的转基因种子(在一些实施例中针对鳞翅目昆虫具有活性)中,所产生的包被的转基因种子控制鳞翅目和鞘翅目昆虫有害生物两者。
[0307]
本发明还涵盖产生抗昆虫(例如,抗鳞翅目昆虫)的转基因植物的方法。在代表性实施例中,所述方法包括:向植物中引入包含编码本发明的嵌合杀昆虫蛋白的核苷酸序列
(包括与在此明确披露的多肽基本上同一的毒素片段和修饰形式)的本发明的多核苷酸、表达盒或运载体,其中所述核苷酸序列被表达于所述植物中以产生本发明的嵌合杀昆虫蛋白,从而赋予所述植物对昆虫有害生物的抗性,并且产生抗昆虫的转基因植物(例如,如与适合的对照植物相比,如不包含本发明的多核苷酸、表达盒或运载体和/或不表达本发明多肽的植物)。
[0308]
在实施例中,向所述植物中引入本发明的多核苷酸、表达盒或运载体的方法包含首先用多核苷酸、表达盒或运载体转化植物细胞并由此再生转基因植物,其中所述转基因植物包含多核苷酸、表达盒或运载体,并表达本发明的嵌合杀昆虫蛋白。
[0309]
可替代地或另外地,所述引入步骤可以包含使包含多核苷酸、表达盒或运载体的第一植物与第二植物(例如,与第一植物不同的植物,例如不包含多核苷酸、表达盒或运载体的植物)杂交,并且任选地,产生包含多核苷酸、表达盒或运载体并表达本发明的嵌合杀昆虫蛋白的子代植物,从而导致对至少一种昆虫有害生物的增加的抗性。因此,本发明的转基因植物涵盖是转化事件的直接结果的植物及其(任一代)子代,所述植物及其子代包含多核苷酸、表达盒或运载体并任选地表达嵌合杀昆虫蛋白,导致对至少一种昆虫有害生物的增加的抗性。
[0310]
本发明进一步提供了鉴定本发明的转基因植物的方法,所述方法包括检测植物(或由其衍生的植物细胞、植物部分等)中的本发明的多核苷酸、表达盒、运载体或嵌合杀昆虫蛋白的存在,从而基于本发明的多核苷酸、表达盒、运载体或嵌合杀昆虫蛋白的存在将所述植物鉴定为本发明的转基因植物。
[0311]
本发明进一步提供了产生对至少一种昆虫有害生物(例如,至少一种鳞翅目有害生物)具有增加的抗性的转基因植物的方法,所述方法包括:种植包含本发明的多核苷酸、表达盒或运载体的种子,并且使来自所述种子的转基因植物生长,其中所述转基因植物包含多核苷酸、表达盒或运载体并产生嵌合杀昆虫蛋白。
[0312]
在实施例中,通过本发明的方法产生的转基因植物包含本发明的多核苷酸、表达盒或运载体。在实施例中,通过本发明的方法产生的转基因植物包含本发明的嵌合杀昆虫蛋白,并且任选地对至少一种昆虫有害生物具有增加的抗性。
[0313]
产生在此描述的转基因植物的方法任选地包括从所述转基因植物中收获种子的另外的步骤,其中所述种子包含多核苷酸、表达盒或运载体并产生嵌合杀昆虫蛋白。任选地,所述种子产生另外的转基因植物,所述转基因植物包含多核苷酸、表达盒或运载体并产生嵌合杀昆虫蛋白,从而对至少一种昆虫有害生物具有增加的抗性。
[0314]
本发明进一步提供了通过本发明的方法生产的转基因植物的植物部分、植物细胞、植物器官、植物培养物、种子、植物提取物、收获产物和加工产物。
[0315]
作为另外的方面,本发明还提供了产生种子的方法,所述方法包括:提供包含本发明的多核苷酸、表达盒或运载体的转基因植物,并且收获来自所述转基因植物的种子,其中所述种子包含多核苷酸、表达盒、运载体并产生嵌合杀昆虫蛋白。任选地,所述种子产生另外的转基因植物,所述转基因植物包含多核苷酸、表达盒或运载体并产生嵌合杀昆虫蛋白,从而对至少一种昆虫有害生物具有增加的抗性。在代表性实施例中,提供所述转基因植物的步骤包括种植产生所述转基因植物的种子。
[0316]
本发明进一步提供了产生杂交植物种子的方法,所述方法包括:使第一近交植物
(其是包含本发明的多核苷酸、表达盒或运载体并任选地表达本发明的嵌合杀昆虫蛋白的转基因植物)与不同近交植物(例如,不包含本发明的多核苷酸、表达盒或运载体的近交植物)杂交,并且允许形成杂交种子。任选地,所述方法进一步包括收获杂交种子。在实施例中,所述杂交种子包含本发明的多核苷酸、表达盒或运载体,并且在实施例中可进一步包含本发明的嵌合杀昆虫蛋白并任选地对昆虫有害生物具有增加的抗性。在实施例中,所述杂交种子产生转基因植物,所述转基因植物包含本发明的多核苷酸、表达盒或运载体,表达本发明的嵌合杀昆虫蛋白,以及对至少一种昆虫有害生物具有增加的抗性。
[0317]
在一些实施例中,本发明的转基因植物对至少一种鳞翅目昆虫有害生物(如在此描述的)具有抗性。在实施例中,所述转基因植物控制秋黏虫昆虫有害生物或集落,所述秋黏虫昆虫有害生物或集落对vip3a(例如,vip3aa蛋白,例如,如在玉蜀黍事件mir162中表达的)和/或cry1f蛋白(例如,cry1fa蛋白,例如,如在玉蜀黍事件tc1507中表达的)具有抗性。
[0318]
在另外的实施例中,控制至少一种昆虫有害生物(例如,至少一种鳞翅目昆虫有害生物,如秋黏虫)的方法包括提供本发明的嵌合杀昆虫蛋白。在实施例中,所述方法包括向昆虫有害生物或其环境递送(例如,经口递送)有效量的本发明的嵌合杀昆虫蛋白。通常,为了有效,所述多肽被昆虫经口摄取。然而,所述嵌合杀昆虫蛋白可以按许多公认的方式被递送至昆虫。将蛋白质经口递送至昆虫的方式包括但不限于(1)在转基因植物中提供蛋白,其中昆虫摄食(摄取)转基因植物的一个或多个部分,从而摄取在转基因植物中表达的多肽;(2)在可施用于或掺入例如昆虫生长培养基的一个或多个配制的蛋白质组合物中提供蛋白;(3)在一种或多种蛋白组合物中提供蛋白,该蛋白组合物可以施用到表面,例如喷雾到植物部分的表面上,然后当昆虫食用一个或多个喷雾的植物部分时被昆虫摄取;(4)诱饵基质;或(5)任何其他本领域公认的蛋白质递送系统。因此,可以使用经口递送至昆虫的任何方法来递送本发明的毒性蛋白。在一些特定实施例中,将本发明的嵌合杀昆虫蛋白经口递送至昆虫,其中所述昆虫摄取本发明的转基因植物的一个或多个部分。
[0319]
在其他实施例中,将本发明的杀昆虫蛋白经口递送至昆虫,其中所述昆虫摄取用包含本发明的杀昆虫蛋白的组合物喷雾的植物的一个或多个部分。可以使用本领域技术人员已知的用于将化合物、组合物、配制品等应用于植物表面的任何方法将本发明的组合物递送至植物表面。递送至或接触植物或其部分的一些非限制性实例包括喷雾、撒粉、喷洒、分散、下雾、雾化、撒播、浸泡、土壤注入、土壤掺入、浸透(例如,根、土壤处理)、浸渍、灌注、涂覆、叶或茎渗透、侧施或种子处理等及其组合。用于使植物或其部分与一种或多种化合物、一种或多种组合物或一种或多种配制品接触的这些和其他程序是本领域技术人员熟知的。
[0320]
在一些实施例中,本发明涵盖为农民提供控制灰翅夜蛾属有害生物昆虫的手段的方法,该方法包括向该农民供应或出售植物材料如种子,该植物材料包含能够表达本发明的txp40杀昆虫蛋白的txp40多核苷酸、表达盒或载体。在其他实施例中,灰翅夜蛾属昆虫有害生物是秋粘虫有害生物,并且植物材料包含本发明的txp40杀昆虫蛋白,并且任选地至少对秋粘虫具有增加的抗性。在其他实施例中,植物材料是玉米种子,并且从种子生长的玉米植物包含本发明的txp40多核苷酸、表达盒或运载体,其表达本发明的txp40杀昆虫蛋白,并且至少对秋粘虫具有增加的抗性。
[0321]
实例
[0322]
通过参考以下具体的实例可以进一步描述本发明。这些实例仅仅是出于说明性目的而提供的,并且并不旨在进行限制,除非另外指明。这里使用的标准重组dna和分子克隆技术是本领域熟知的,并由j.sambrook等人,molecular cloning:a laboratory manual[分子克隆:实验室手册],第3版,冷泉港,纽约:冷泉港实验室出版社(2001);t.j.silhavy,m.l.berman和l.w.enquist,experiments with gene fusions[基因融合实验],冷泉港实验室,冷泉港,纽约(1984)以及ausubel,f.m.等人,current protocols in molecular biology[分子生物学现代方法],纽约,john wiley and sons inc.[约翰
·
威利父子公司],(1988),reiter,等人,methods in arabidopsis research[拟南芥研究方法],world scientific press[世界科学出版社](1992),以及schultz等人,plant molecular biology manual[植物分子生物学手册],kluwer academic publishers[克吕韦尔学术出版集团](1998)所述。
[0323]
实例1.txp40蛋白对cry抗性秋粘虫的杀昆虫活性。
[0324]
使用人造饮食生物测定法测定从发光光状杆菌获得的txp40蛋白(seq id no:1)的杀昆虫活性,其中将包含txp40蛋白的大肠杆菌提取物覆盖在人造昆虫饮食的表面上。用于这些生物测定法的faw菌株来自对cry1f和cry1ab杀昆虫蛋白具有抗性的faw田间群体。将vip3蛋白用作阳性对照。将包含空载体(即没有txp40编码序列的载体)的大肠杆菌克隆用作阴性对照。生物测定法的结果记录为第7天的死亡率和有效死亡率百分比,其中有效死亡率考虑在测试期间faw幼虫达到的生长阶段,即具有生长抑制和垂死的处理过的幼虫计数为有效死亡。
[0325]
结果示出在表1中。当在昆虫生物测定法中测试包含txp40的大肠杆菌提取物时,txp40令人惊讶地显示出对cry1f抗性faw的显著杀昆虫活性。这些结果不仅证明了txp40蛋白对faw的令人惊讶的口服活性,而且表明txp40具有与来自苏云金芽孢杆菌的cry1f和cry1ab杀昆虫蛋白不同的作用模式。
[0326]
表1.txp40对cry抗性秋粘虫的杀昆虫活性
[0327][0328]
为了测试txp40蛋白的效力,如上所述制备txp40大肠杆菌提取物的稀释物并将其添加到人造昆虫饮食的表面。还对txp40处理和vip3阳性对照处理测试了未稀释的提取物。
[0329]
表2中所示的结果表明,txp40蛋白即使在1:32稀释度下也具有显著活性,这表明txp40在转基因玉米植物中的表达将是可行的faw控制选择,特别是对于cry抗性faw。
[0330]
表2.txp40-1对cry抗性faw的效力
[0331][0332]
实例2.txp40变体的faw活性。
[0333]
在txp40-1(seq id no:1)中引入突变,并测定来自各txp40变体的细菌裂解物的蛋白质稳定性和杀昆虫活性。突变包括在多个残基处的氨基酸改变以及氨基酸残基的插入和/或缺失。使用基本上如实例1中所述进行的饮食覆盖测定法,测试具有一个或多个突变的变体txp40蛋白对两种秋粘虫菌株(来自巴西的菌株(br-faw;草地贪夜蛾)和来自北美的菌株(na-faw;草地贪夜蛾))的杀昆虫活性。营养期杀昆虫蛋白vip3d、野生型txp40蛋白和/或苏云金芽孢杆菌菌株c0756用作阳性对照。pet29空载体用作阴性对照。表3-4显示了具有靶向突变的变体txp40蛋白的表达、溶解度和/或杀昆虫活性测定法的结果。
[0334]
表3.具有单突变的变体txp40蛋白的活性
[0335]
[0336][0337]
表4.具有多个突变的变体txp40蛋白的活性
[0338]
[0339][0340]
表3中的结果表明,可以产生与野生型txp40蛋白相比对秋粘虫(faw)具有相等或更好杀昆虫活性的变体txp40蛋白。例如,在k73a、k191a、k209a、k238a、k309a、r10a、r26a、r46a、r186a和r226a处具有单突变的变体蛋白对na-faw具有与野生型txp40大致相同的杀昆虫活性,而k49a、k200a、k221a、r26a、r167a、r208a和r217a对br-faw具有与野生型txp40大致相同的杀昆虫活性。
[0341]
与野生型txp40蛋白相比,txp40中的某些突变产生对na-faw具有增加或显著增加(≥20%)的杀昆虫活性的变体蛋白。例如,与野生型txp40相比,在k31a、k49a、k73a、k75a、k103a、k111a、k119a、k170a、k191a、k200a、k210a、k213a、k221a、k284a、k333a、r167a、r208a和r252a处具有单突变的变体蛋白对na-faw都具有更好的活性,并且在这些突变体中,与野生型txp40相比,k31a、k49a、k75a、k111a、k119a、k200a和k333a对na-faw的杀昆虫活性增加20%或更多。与野生型txp40蛋白相比,txp40中的某些突变产生对br-faw的杀昆虫活性增加或显著增加(≥14%)的变体蛋白。例如,与野生型txp40相比,在k31a、k49a、k73a、k75a、k103a、k111a、k119a、k200a、k213a和k333a处具有单突变的变体蛋白对br-faw都具有更好的活性,并且其中,与野生型txp40蛋白相比,k73a、k103a和k213a对br-faw具有显著增加的杀昆虫活性。
[0342]
结果进一步证明,与br-faw相比,txp40(seq id no:1)中的某些突变产生对na-faw具有惊人差异毒性的变体蛋白。一些突变体对na-faw的毒性大于br-faw,例如k31a、k48a、k75a、k111a、k119a、k133a、k170a、k209a、k210a、k238a、k333a、r10a、r208a和r226a。一些变体对br-faw的毒性大于na-faw,例如,k73a、k103a、k213a、k247a、k271a、k284a、r26a、r46a、r187a、r217a和r240a,表明这些位置对于差异faw活性是重要的。
[0343]
与野生型txp40相比,大多数包含多个突变的变体txp40蛋白对na-faw具有降低的杀昆虫活性,并且一些突变完全敲除活性,再次表明这些氨基酸位置对txp40的生物活性的重要性。一些具有多个突变的变体具有与野生型txp40相同的活性,与野生型txp40相比,在位置k119a/k213a处具有突变的一种变体(seq id no:127)显著增加了杀昆虫活性。
[0344]
将编码变体txp40-k119a/k213a蛋白(seq id no:127)的dna用作模板,通过(皮斯卡塔韦,新泽西州)对全长txp40-k119a/k213a dna使用简并寡核苷酸随机诱变技术产生进一步的突变。将包含随机突变的变体文库亚克隆到pet29a载体中。测试数百种包含变体txp40蛋白的细菌裂解物对na-faw、br-faw和大豆尺蠖(sbl;黄豆银纹夜蛾)的杀昆虫活性,如上所述。某些变体的结果示于表5中。
[0345]
表5.包含随机突变的变体txp40蛋白的活性
[0346][0347]
如上所述,野生型txp40(seq id no:1)和变体txp40蛋白对na-faw和br-faw具有差异毒性。随机化实验的目的是鉴定一种或多种txp40变体,其包含赋予对na-和br-faw菌株以及其他鳞翅目物种的高杀昆虫活性的突变。用作随机诱变的模板的包含k119a/k213a突变的txp40变体(seq id no:127)对na-faw具有非常高的活性,但对br-faw和sbl具有低得多的活性(参见表5)。
[0348]
数百种随机诱变的txp40突变体的生物测定结果表明,大多数保持对na-faw和br-faw的差异毒性。然而,三个变体,txp40-k119a/k213a/s11y/m86i(seq id no:130);txp40-k119a/k213a/s11y(seq id no:131)和txp40-k119a/k213a/f169v(seq id no:132)被鉴定为,与用作模板的野生型txp40(seq id no:1)和txp40-k119a/k213a变体(seq id no:127)的活性相比,保持了对na-faw的高活性,并且令人惊讶地对br-faw也具有非常高的活性。与变体模板txp40-k119a/k213a相比,这些变体中的一个txp40-k119a/k213a/s11y(seq id no:131)也令人惊讶地对大豆尺蠖(sbl)具有显著增加的活性。这些结果强烈表明,seq id no:1的氨基酸位置11、119和213在确定txp40蛋白的杀昆虫效力方面是非常重要的。
[0349]
在表6中显示了如上所述制备并针对na-faw测试的另外其他变体txp40蛋白。表6中的所有突变均是指seq id no:1中的氨基酸取代。
[0350]
表6.seq id no:1中包含突变的变体txp40蛋白的活性
[0351]
[0352][0353]
表6中所示的结果支持以下发现:seq id no:1中的至少氨基酸位置119和213对于确定faw活性是重要的。
[0354]
实例3.测试txp40对vip3抗性faw的活性
[0355]
为了确定txp40和/或变体蛋白的毒性是否通过与vip3a蛋白不同的moa,如实例2所述产生上述txp40野生型和txp40-k119a变体蛋白。将txp40蛋白分别在txp40缓冲液中混合,并使用bio-rad experion系统(伯乐公司(biorad),加利福尼亚州赫拉克勒斯(hercules,ca))监测溶解的蛋白制剂的纯度。
[0356]
测试纯化的蛋白针对对vip3a杀昆虫蛋白具有抗性的faw菌株的效力。基本上如实例1所述进行饮食覆盖测定。将vip3a蛋白溶解在pbs中。两种阴性对照处理为pbs和txp40缓冲液。cry1fa蛋白用作vip3a抗性faw菌株的阳性对照。测试多个浓度的各蛋白质。将杀昆虫活性评估为第7天的有效死亡率(将具有生长抑制和垂死的幼虫记为有效死亡)。
[0357]
结果表明vip3a抗性faw菌株不受vip3a控制,表明该菌株对该蛋白质具有抗性。相反,txp40野生型蛋白和txp40-k119a变体对vip3抗性昆虫引起60%的死亡率,表明txp40的作用模式不同于vip3a蛋白的作用模式,因此可用于与vip3叠加组合以减轻faw群体中抗性
的发展。
[0358]
实例4.对大肠杆菌裂解物制剂进行的模拟胃液测试
[0359]
本实例描述了确定txp40和/或txp40变体的sgf消化率的测定。每种txp40蛋白和/或变体在大肠杆菌菌株bl21*(de3)中产生。确定变体在细菌菌株中的表达水平和变体的溶解度。将缓冲液中的细菌裂解物稀释至3mg/ml(总蛋白浓度)用于消化率分析。在37℃下,通过将15μl裂解物添加至285μl模拟胃液[10个单位的胃蛋白酶/μg蛋白,或大约1579个单位的胃蛋白酶/ml,在g-con溶液(2mg/ml氯化钠,ph 1.2)]中来启动消化反应。在5分钟时,将100μl的裂解物-sgf反应液去除,并通过将其添加到100μl的预热(95℃)的终止溶液中来终止该反应,该终止溶液由65tricine加样缓冲液(bio-rad 2x tricine加样缓冲液w/10β-巯基乙醇)和35 500mm碳酸氢钠(ph 11.0)组成。通过将5μl测试裂解物添加到预热(95℃)的100μl终止溶液和95μl模拟胃液中来产生零时间点(t0)。将所有样品在95℃下加热5分钟,并然后储存在冰上直至sds-page分析。在标准蛋白质凝胶电泳之前,将30微升的每种反应液加样到10-20tris-tricine肽凝胶上。电泳后立即用40甲醇:10乙酸混合物固定tris-tricine凝胶20分钟。然后将凝胶用gelcode蓝蛋白染色剂在室温下染色1小时。1小时后,将聚丙烯酰胺凝胶用蒸馏水脱色至少12小时。结果包括“通过”“未通过”评估。t5测试的“未通过”表示在凝胶电泳后可通过gelcode蓝蛋白染色检测到完整的或部分消化的txp40蛋白质和/或变体,这表明该蛋白质在sgf测定中不能被完全消化。t5测试的“通过”表示不可检测到完整的txp40蛋白质和/或变体,这表明txp40蛋白质和/或变体在sgf测定中是可消化的。这些实验的结果表明,txp40和变体txp40蛋白在sgf测定中被完全消化。
[0360]
实例5.用txp40-1密码子优化基因转化玉蜀黍。
[0361]
产生适于农杆菌介导的玉蜀黍转化的二元载体构建体。二元载体包含编码txp40-k119a/k213a(seq id no:127)的玉蜀黍优化核酸和编码txp40-k119a/k213a/s11y(seq id no:131)的玉蜀黍优化核酸,在5’端可操作地连接至适合于驱动在植物中表达的启动子,并在3’端可操作地连接至终止子序列。例如,使用美国专利号6,320,100(通过引用并入本文)中所述的方法进行玉蜀黍密码子优化。使用本领域技术人员已知的标准分子生物学技术将该构建体转化到根癌农杆菌中。为了制备用于转化的农杆菌,将细胞在28℃和220rpm下在液体ypc培养基中培养过夜。基本上如在negrotto等人,2000(plant cell reports[植物细胞通讯]19:798-803)中所说明进行未成熟玉蜀黍胚的农杆菌转化。对于这个实例,所有的培养基组分基本上如negrotto等人(如上)所述。然而,本领域内已知的多种培养基组分可以被替代。
[0362]
在转化、选择和再生后,使用分析来测定植物中是否存在编码选择性标记的基因和txp40玉蜀黍密码子优化的编码序列。还测试了植物中是否存在载体骨架。将对载体骨架为阴性且包含该转基因的一个拷贝的植物转移至温室并测定其对faw损害的抗性。结果显示两种蛋白均在植物中表达并且对秋粘虫具有活性。
[0363]
实例6.与第二杀昆虫剂组合的txp40-1。
[0364]
txp40和/或txp40变体如实例1纯化为细菌裂解物或纯化为蛋白质。制备第二杀昆虫剂,例如cry蛋白和/或vip和/或dsrna。在非限制性实例中,cry蛋白可以是cry1ab、cry1b、cry1c、cry1d、cry1f、cry1j和/或cry2a蛋白。在其他非限制性实例中,vip3蛋白可以是vip3a和/或vip3b蛋白。dsrna可靶向编码液泡atp合酶、β-微管蛋白、26s蛋白体亚基p28
蛋白、ef1α48d、肌钙蛋白i、跨膜四蛋白、γ-外被体、β-外被体和/或保幼激素环氧化物水解酶的基因(wo公开号wo 2018/026770、wo 2018/026773、和wo2018/026774;美国专利号7,812,219;各自通过引用并入本文)。在饮食覆盖测定中测试纯化的txp40蛋白和第二杀昆虫剂对faw的杀昆虫功效,所述测定基本上如实例1中所述进行,除了添加了第二杀昆虫剂。
[0365]
应当理解的是,本文描述的实例和实施例仅是出于说明性的目的,并且根据其说明书的不同修改或变化将提示本领域的技术人员并且将会包含在本技术的精神和范围内以及所附权利要求的范围内。
[0366]
在本说明书中提到的所有公开物和专利申请均指示本发明所属领域中的技术人员的技术水平。所有公开物和专利申请均通过援引并入本文,其程度如同每个单独的公开物或专利申请被明确地并单独地指示通过援引而并入。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1