人α‑防御素5改造体及其应用的制作方法
本发明涉及生物技术领域,具体涉及人α-防御素5改造体及其应用。
背景技术:
抗生素耐药是威胁人类健康的重大科学问题。耐甲氧西林金黄色葡萄球菌(MRSA)、泛耐药鲍曼不动杆菌、耐碳青霉烯类铜绿假单胞菌等广谱甚至超广谱耐药菌的出现使得一些感染的治疗相当棘手,成为这些感染患者死亡的重要危险因素。目前耐药菌突变的速度快于新型抗菌药物的研发速度,人类对于一些耐药菌感染无能为力,面临“后抗生素时代”的严峻挑战。因此,探寻新型的抗菌药物,尤其是对广谱和超广谱耐药菌具有杀菌能力的新药,已经迫在眉睫。
阳离子抗菌肽是一类具有广谱抗菌活性的小分子多肽,是生物体对抗外界微生物侵袭的一道重要的免疫防线。阳离子抗菌肽的抗菌作用具有杀菌快速、作用广谱和难以形成抗性的特点,这使得其成为理想的候选新型抗生素。阳离子抗菌肽作用机理是其带正电荷,可与带负电荷的细菌表面结合,嵌入细菌的细胞膜内,形成孔洞,破坏其细胞膜的连续性,使其细胞质外流,最终导致细菌的死亡,区别于传统抗生素的杀菌靶点如蛋白体、RNA酶等,杀菌效果直接快速强大,且不易诱导传统的依赖基因突变、水解酶产生等的耐药性。一些抗菌肽作为抗耐药细菌的抗菌素,已进入临床试验(M.Zasloff,Nature,2002,415,389-395)。
人α-防御素5(human defensin-5,HD-5)是一种主要表达于肠道粘膜的天然阳离子抗菌肽,对大肠杆菌、金黄色葡萄球菌、蜡样芽胞杆菌、产气肠杆菌等多种细菌及真菌均具有较强的杀菌作用。此外,HD-5还可以通过调节肠道局部炎症因子的释放发挥免疫调节作用。因此,HD-5在肠道免疫屏障中发挥极其重要作用。
尽管目前HD-5的提取及人工合成已较成熟,但是由于完全仿生天然的HD-5抗菌活性容易受到诸多机体内环境因素的影响,如生理浓度的氯化钠的存在即可极大地降低其抗菌活性,限制其成为适宜产业化开发生产、临床应用的新型抗菌药物。
因此,本发明根据HD-5的理化性质对HD-5进行改造,克服HD-5的功能缺陷,使其有望成为应用于临床的新型抗菌药物。
技术实现要素:
本发明提供了一种人α-防御素5改造体,在人α-防御素5的C端连接不同长度的脂肪酸链,使其成为阳离子两亲性自组装纳米抗菌肽,获取了作用稳定、高效、适宜产业化生产且具备多种功能的人工抗菌肽。
人α-防御素5改造体,由人α-防御素5在C末端添加氨基酸残基并修饰连接脂肪酸改造获得,所添加的氨基酸残基中至少有一个氨基酸残基具有游离氨基,所述游离氨基上修饰连接碳原子数为8-24的脂肪酸。
本发明的改造以HD-5为模板,HD-5由32个氨基酸残基组成,其中有6个半胱氨酸,按N端到C端的顺序排列,第1个和第6个、第2个和第4个、第3个和第5个半胱氨酸之间分别两两形成二硫键,一共3个二硫键。本发明在其C端进行了延伸,最终得到了本发明的改造体。
研究表明,对HD-5的C端进行脂肪酸链修饰,能够显著提高人α-防御素5的杀菌活性,而且C端修饰得到的改造体较N端修饰具备更强的杀菌能力。
本发明改造体具备杀菌力强而溶血率低的特点。作为优选,所述脂肪酸的碳原子数为12-14个。更为优选的,所述脂肪酸为肉豆蔻酸或月桂酸。
本发明通过在C端添加氨基酸延长肽链,利于脂肪酸的修饰。作为优选,添加的氨基酸残基的数量为2-5个。
脂肪酸的修饰是通过脂肪酸的羧基与C端氨基酸残基上的游离氨基发生脱水缩合反应实现。因此,HD-5改造体C端添加氨基酸至少有一个为至少带有两个氨基的氨基酸,如Lys,其中α氨基用于合成肽链的肽键,侧链上氨基用于脂肪酸链的修饰。
作为优选,所述改造体的肽链由34个氨基酸残基组成,其氨基酸序列为:
Ala1-Thr2-Cys3-Tyr4-Cys5-Arg6-Thr7-Gly8-Arg9-Cys10-Ala11-Thr12-Arg13-Glu14-Ser15-Leu16-Ser17-Gly18-Val19-Cys20-Glu21-Ile22-Ser23-Gly24-Arg25-Leu26-Tyr27-Arg28-Leu29-Cys30-Cys31-Arg32-Gly33-Lys34。
作为优选,所述改造体的C端羧基酰胺化。
本发明将上述序列第34位的Lys的C-末端酰胺化,且ε氨基上连接肉豆蔻酰基或月桂酰基,分别命名为HD-5Myr和HD-5Lau,研究表明,改造体HD-5Myr和HD-5Lau对革兰氏阳性菌和革兰氏阴性菌均有杀灭作用。HD-5Myr和HD-5Lau可以形成胶束结构,胶束结构的形成使这些肽的单体聚集成纳米级别的颗粒,形成多聚体形式,使正电荷更加集中,其与细菌的结合能力更强,从而发挥更强大的杀菌作用。
本发明还提供了所述的人α-防御素5改造体在制备抗菌药物中的应用。
针对金黄色葡萄球菌ATCC 25923,HD-5Myr的LD90值约为HD-5的1/4,HD-5Lau的LD90值约为HD-5的1/3;针对大肠杆菌ATCC 25922,HD-5Myr的LD90值小于HD-5的1/8,HD-5Lau的LD90值小于HD-5的1/6,表明HD-5Myr和HD-5Lau的杀菌作用强于HD-5。针对临床分离的耐甲氧西林的金黄色葡萄球菌菌株MRSA-1,HD-5Myr的LD90值约为HD-5的1/15,HD-5Lau的LD90值约为HD-5的1/4。
HD-5、HD-5Myr和HD-5Lau针对耐甲氧西林金黄色葡萄球菌的LD90值远低于金黄色葡萄球菌株ATCC 25923的LD90值,表明其抗菌机制区别于传统的抗生素。因此,可将本发明的改造体HD-5Myr和HD-5Lau应用于制备抗耐药菌药物。
在耐盐能力方面,HD-5Myr显著强于HD-5Lau和HD-5,即使在生理浓度下(氯化钠浓度135mM-145mM)仍能发挥较强的杀菌作用,因此,HD-5Myr可作为抗菌的有效成分应用于制备静脉注射药物中。
本发明还提供了所述的人α-防御素5改造体在制备免疫调节药物中的应用。
研究表明,改造体HD-5Myr和HD-5Lau能够促进巨噬细胞释放炎症因子IL-1β,IL-1β在免疫调节过程中起重要作用。因此,可将其应用到制备免疫调节剂中。
本发明还提供了一种药物组合物,包含有效量的人α-防御素5改造体和药学上可接受的载体。
所述HD-5改造体的血药浓度达到25μg/ml时,即可发挥高效的杀菌作用,且溶血率较低,是一种理想的代替传统抗生素的抗菌药物。
本发明具备的有益效果:
(1)本发明在人α-防御素5的C末端进行延伸和修饰,有效增强其抗菌活性,尤其是针对耐甲氧西林金黄色葡萄球菌,有望成为代替传统抗生素的抗菌药物。
(2)本发明得到的改造体HD-5Myr和HD-5Lau可以形成胶束结构,胶束结构的形成使正电荷更加集中,其与细菌的结合能力更强,从而发挥更强大的杀菌作用。
(3)本发明在人α-防御素5的C末端增加Gly和Lys,并进行肉豆蔻酸修饰,显著提高改造体的耐盐性。
(4)本发明的改造体能够显著促进巨噬细胞释放炎症因子,可将其应用到免疫调节药物的制备中。
附图说明
图1为HD-5及本发明改造体HD-5Myr和HD-5Lau的透射电镜图,其中A为HD-5,B为HD-5Myr,C为HD-5Lau。
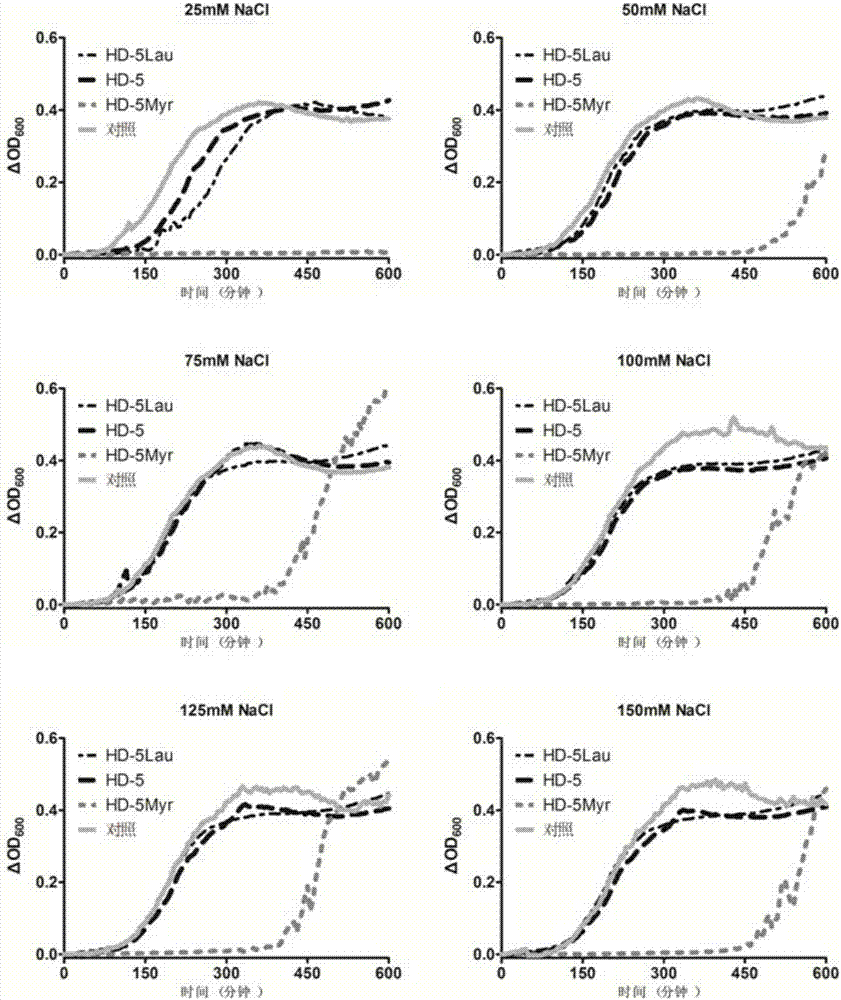
图2为实施例5中HD-5及改造体在不同氯化钠浓度条件下的抗菌活性结果图。
图3为实施例6中HD-5及改造体对人红细胞的溶血作用结果图。
图4为实施例7中HD-5及改造体调节巨噬细胞IL-1β成熟体的释放结果图。
具体实施方式
为了更好地理解本发明,下面结合实施例和附图对本发明做进一步的详细说明,但下述实施例不是对本发明保护范围的限制,任何在本发明基础上做出的改变和变化,都在本发明的保护范围之内。
实施例1
HD-5的氨基酸序列如SEQ ID NO.1所示,具体为:
Ala1-Thr2-Cys3-Tyr4-Cys5-Arg6-Thr7-Gly8-Arg9-Cys10-Ala11-Thr12-Arg13-Glu14-Ser15-Leu16-Ser17-Gly18-Val19-Cys20-Glu21-Ile22-Ser23-Gly24-Arg25-Leu26-Tyr27-Arg28-Leu29-Cys30-Cys31-Arg32,其中,Cys3和Cys31、Cys5和Cys20、Cys10和Cys30之间分别形成分子内二硫键。
本发明的改造体HD-5Myr和HD-5Lau是在HD-5分子基础上,在其C-端进行改造而获得的。HD-5Myr和HD-5Lau的氨基酸序列如SEQ ID NO.2所示。
HD-5Myr由34个氨基酸残基组成,分子量为3976.8Da。
HD-5Myr的氨基酸序列为:
Ala1-Thr2-Cys3-Tyr4-Cys5-Arg6-Thr7-Gly8-Arg9-Cys10-Ala11-Thr12-Arg13-Glu14-Ser15-Leu16-Ser17-Gly18-Val19-Cys20-Glu21-Ile22-Ser23-Gly24-Arg25-Leu26-Tyr27-Arg28-Leu29-Cys30-Cys31-Arg32-Gly33-Lys34-NH2,其中,Cys3和Cys31、Cys5和Cys20、Cys10和Cys30之间分别形成分子内二硫键。第34位为一个Lys,其ε氨基上连接一个肉豆蔻酰基。
HD-5Lau由34个氨基酸组成,分子量为3948.7Da。
HD-5Lau的氨基酸残序列为:
Ala1-Thr2-Cys3-Tyr4-Cys5-Arg6-Thr7-Gly8-Arg9-Cys10-Ala11-Thr12-Arg13-Glu14-Ser15-Leu16-Ser17-Gly18-Val19-Cys20-Glu21-Ile22-Ser23-Gly24-Arg25-Leu26-Tyr27-Arg28-Leu29-Cys30-Cys31-Arg32-Gly33-Lys34-NH2,其中,Cys3和Cys31、Cys5和Cys20、Cys10和Cys30之间分别形成分子内二硫键。第34位为一个Lys,其ε氨基上连接一个月桂酰基。
此外,在HD-5基础上,在其N-端进行改造,获得的改造体,命名为MyrHD-5。MyrHD-5由33个氨基酸残基组成,分子量为3849.6Da。
MyrHD-5的氨基酸序列如SEQ ID NO.3所示,具体为:
Gly1-Ala2-Thr3-Cys4-Tyr5-Cys6-Arg7-Thr8-Gly9-Arg10-Cys11-Ala12-Thr13-Arg14-Glu15-Ser16-Leu17-Ser18-Gly19-Val20-Cys21-Glu22-Ile23-Ser24-Gly25-Arg26-Leu27-Tyr28-Arg29-Leu30-Cys31-Cys32-Arg33,其中,Cys4和Cys32、Cys6和Cys21、Cys11和Cys31之间分别形成分子内二硫键。第1位为一个Gly,其氨基上连接一个肉豆蔻酰基。
实施例2
上述实施例1的HD-5和HD-5Myr由“上海吉尔多肽有限公司”合成。上述实施例1的HD-5Lau和MyrHD-5由“杭州中肽生化有限公司”合成。
实施例3
抗菌肽的抗菌活性测定,比较实施例2合成的HD-5的改造体HD-5Myr、HD-5Lau及MyrHD-5与天然HD-5的抗菌作用的强弱。
依据Ericksen等2005年报道的检测抗菌肽抗菌活性的方法(Ericksen B,et al.,Antibacterial activity and specificity of the six human{alpha}-defensins.Antimicrob Agents Chemother,2005)进行实验。
将细菌接种在LB琼脂平板上,置于37℃恒温培养至长出菌落。挑取单菌落,接种到5mlLB培养基中,37℃恒温振荡培养4-6小时。用无菌水洗涤2次。在多功能酶标仪上测定600nm处的吸光值,1OD600=4×108CFU/ml,将受试细菌用无菌水稀释至2×106CFU/ml。
在无菌96孔板中每孔加入50μl无菌水,然后在A1孔中加入稀释到一定浓度的抗菌肽HD-5样品。混匀后取50μl加入A2孔,依次倍比稀释,最后从A6孔吸出50μl弃去。同样的方法稀释MyrHD-5、HD-5Myr和HD-5Lau样品。对照孔为50μl无菌水。然后各个孔分别加入50μl的2×106CFU/ml细菌。37℃恒温振荡培养2小时。然后分别加入100μl的2倍浓度的LB培养基。在酶标仪中恒温37℃,并每间隔5分钟测量一次600nm处吸光值。
将所得吸光值减去0时刻的吸光值,并取对数。于坐标系中以时间为横坐标、以OD值的变化值的对数为纵坐标,所得的点在对数生长期中呈线性。根据此线性规律,计算出不同组的杀菌率。并依据各个不同浓度的杀菌率计算出杀灭90%的细菌时所需要的抗菌肽的浓度,称之为LD90。HD-5及MyrHD-5、HD-5Myr和HD-5Lau对不同菌种的的LD90见下表1。
表1、抗菌肽HD-5及其改造体的抗菌活性
所述的大肠杆菌ATCC 25922、金黄色葡萄球菌ATCC 25923购自美国模式培养物集存库(ATCC),MRSA-1为临床分离得到的耐甲氧西林的金黄色葡萄球菌菌株。
由表1可见,HD-5Myr和HD-5Lau对革兰氏阳性菌金黄色葡萄球菌、革兰氏阴性菌大肠杆菌及临床分离菌株MRSA-1均具有杀灭作用,且改造体HD-5Myr和HD-5Lau的作用均强于野生肽HD-5。
上述实验结果表明,本实验的HD-5Myr和HD-5Lau的抗菌活性,无论是对革兰氏阳性金黄色葡萄球菌,抑或革兰氏阴性大肠杆菌,还是临床分离得到的耐甲氧西林的金黄色葡萄球菌,均明显高于天然HD-5,而且HD-5Myr和HD-5Lau对MRSA的抗菌能力强于对甲氧西林敏感的金黄色葡萄球菌菌株ATCC 25923,表明其抗菌机制可能区别于传统抗生素。
MyrHD-5和HD-5Myr均为HD-5分子肉豆蔻酰化修饰后的改造体,MyrHD-5是在N端修饰,HD-5Myr是在C端修饰,而HD-5Myr的杀菌能力显著强于MyrHD-5。且MyrHD-5杀灭大肠杆菌ATCC 25922菌株的能力甚至弱于HD-5。说明了对HD-5分子C端进行脂肪酸修饰更有利于其发挥杀菌作用。
实施例4
抗菌肽HD-5的改造体HD-5Myr和HD-5Lau可以形成胶束结构,更有利于其发挥杀菌作用。
分别取少许HD-5、HD-5Myr和HD-5Lau样品滴在电镜用铜网上,用滤纸吸掉多余样品,室温干燥后,使用醋酸双氧铀染色。后置于透射电子显微镜中观察。
如附图1A所示,在天然HD-5中未观测到胶束结构。如附图1B中箭头所示,HD-5Myr可以形成胶束结构。如附图1C中箭头所示,大小均一的球状颗粒物为胶束结构,说明HD-5Lau可以形成胶束结构。胶束结构的形成使这些肽的单体聚集成纳米级别的颗粒,形成多聚体形式,使正电荷更加集中,其与带负电荷的细菌的结合能力更强,使其更有利于穿透细菌的细胞壁和细胞膜,从而发挥更强的杀菌作用。
实施例5
抗菌肽HD-5及其改造体HD-5Myr和HD-5Lau的抗菌活性对盐的耐受能力的强弱。
依据Ericksen等2005年报道的检测抗菌肽抗菌活性的方法(Ericksen B,et al.,Antibacterial activity and specificity of the six human{alpha}-defensins.Antimicrob Agents Chemother,2005)进行实验。
将大肠杆菌接种在LB琼脂平板上,置于37℃恒温培养至长出菌落。挑取单菌落,接种到5ml LB培养基中,37℃恒温振荡培养4-6小时。用无菌水洗涤2次。在多功能酶标仪上测定600nm处的吸光值,1OD600=4×108CFU/ml,将受试细菌用无菌水稀释至2×106CFU/ml。
分别在A1至A6孔中加入25μl一定浓度的NaCl溶液。然后各个孔分别加入25μl的100μg/ml的HD-5溶液。同样的方法用于HD-5Myr组和HD-5Lau组。对照孔为25μl不同浓度的盐溶液与25μl无菌水的混合液。然后向各个孔中加入50μl的2×106CFU/ml大肠杆菌(ATCC25922)。37℃恒温振荡培养2小时。然后分别加入100μl的2倍浓度的LB培养基。在酶标仪中恒温37℃,并每间隔5分钟测量一次600nm处吸光值。
将所得吸光值减去0时刻的吸光值。以时间为横坐标,以OD值的变化量为纵坐标作图。根据曲线到达某一个OD值的时间的长短,来推断其0时刻的细菌存活量的多少,进而推断其杀菌作用受到盐的影响的大小。
如附图2所示,HD-5Myr的耐盐能力显著强于HD-5和HD-5Lau。即使在生理浓度氯化钠溶液(135mM-145mM)情况下,HD-5Myr仍可以发挥较强的杀菌作用。
实施例6
抗菌肽HD-5及改造体HD-5Myr和HD-5Lau对人红细胞的溶血作用测定
1)取新鲜人血,肝素化后离心取红细胞层。
2)用PBS洗涤3次。
3)在EP管中分别加入终浓度分别为50μg/ml、25μg/ml的HD-5及改造体。阳性对照组加入等体积10%Triton X-100。阴性对照组加入等体积PBS。
4)分别加入4%的人红细胞,37℃孵育1小时。
5)离心,取等量上清至96孔板中,测量540nm处的OD值,以阳性对照组为100%溶血,以阴性对照组为0%溶血,计算各孔的溶血率。
结果如附图3所示,在远高于LD90的浓度25μg/ml时,HD-5Myr的溶血率为6.83%,HD-5Lau的溶血率为2.24%。这说明改造体HD-5Myr和HD-5Lau均有较为广阔的安全范围,非常有利于在医药领域进一步的开发应用。
实施例7
抗菌肽的改造肽发挥免疫调节的作用,促进炎症因子IL-1β的释放
1)用加入10%的胎牛血清的RPMI-1640培养基养THP-1细胞,在其对数期时,接种在24孔板上,每孔106个细胞。加入终浓度为100nM的PMA刺激12小时,使细胞贴壁。
2)用加入10%的胎牛血清的RPMI-1640培养基换液,静置24小时。
3)组别设置有:空白组、LPS刺激组、LPS+5mM ATP组(阳性对照组)、LPS+HD-5组、LPS+HD-5Myr组、LPS+HD-5Lau组。用或者不用终浓度为100ng/ml的脂多糖(LPS)刺激3h,然后使用1640培养基轻柔洗涤2次,再加入或者不加终浓度为10μg/ml及20μg/ml的HD-5Myr或HD-5Lau或终浓度为5mM的ATP,或者终浓度为100μg/ml及200μg/ml的HD-5,刺激3h后,收集各组的上清。
4)将各组等量的上清中的蛋白浓缩,用蛋白裂解液裂解后,用免疫印迹法比较不同组别间IL-1β成熟体的释放情况。
如附图4所示,LPS+5mM ATP组(阳性对照组)、LPS+HD-5组、LPS+HD-5Myr组、LPS+HD-5Lau组均有IL-1β成熟体的释放,证明HD-5、HD-5Myr和HD-5Lau能够促进IL-1β成熟体的释放,而HD-5Myr和HD-5Lau的浓度远低于HD-5,表明改造体HD-5Myr和HD-5Lau的免疫调节活性远高于HD-5。
- 还没有人留言评论。精彩留言会获得点赞!