分化的肠内分泌细胞和胰岛素产生细胞的制备的制作方法
本申请要求2016年1月8日提交的美国临时申请no.62/276,814的权益。上述申请的全部教导通过引用结合在本文中。政府支持本发明在国立卫生研究院(nationalinstitutesofhealth)授予的批准号r01de013023的政府支持下完成。政府拥有本发明的某些权利。
背景技术:
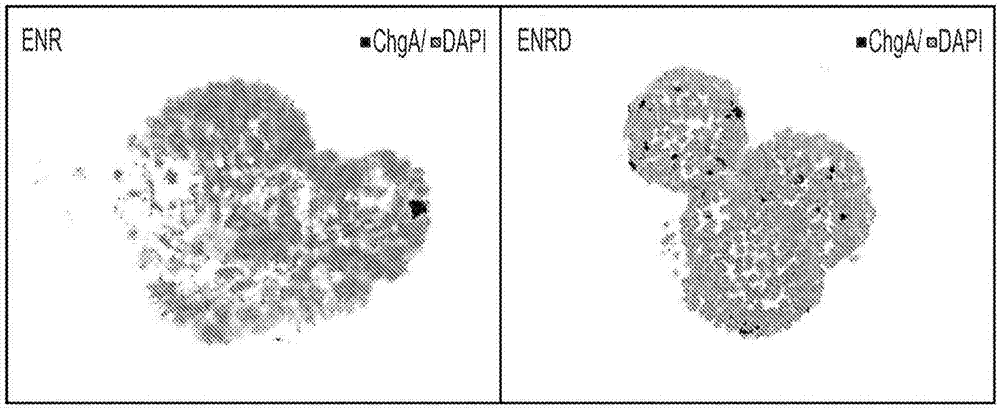
肠内分泌系统通过使用多种激素来协调身体如何对营养物质反应,以调整体内广泛的生理反应,从而在消化道和代谢性疾病中发挥重要作用,例如胃肠道(gi)疾病、糖尿病和肥胖症。肠内分泌细胞(eec)形成体内最大的内分泌系统(grible和reimann,2015)。eec沿着隐窝-绒毛轴独立分散在整个肠上皮中,但体内仅存在一小部分(~1%)(gunawardene等,2011)。eec的关键功能是感觉腔内容物,特别是营养物质,并且通过分泌多种激素(例如glp-1)而响应,所述多种激素调节食物摄入、能量稳态和葡萄糖耐受(furness,2013)。还提出,eec通过分泌激素(例如glp-1、pyy和glp-2)在胃旁路手术中发挥关键作用(mumphrey,2013)。因此,eec是糖尿病和肥胖症的治疗靶点。此外,确凿的证据证明了eec在先天免疫中的免疫调节功能(moran,2008)。eec表达功能性toll样受体(tlr)并直接响应于由共生细菌产生的代谢产物(bogunovic,2007)。最近的证据也表明eec可通过改变炎症期间的数量和激素分泌来直接协调免疫细胞功能(worthington,2015)。eec也被认为在代谢性疾病(例如糖尿病和肥胖症)和胃肠道病理,例如肠易激综合症、感染性肠炎和炎性肠病中起关键作用(moran,2008,和manocha和kahn,2012)。因此,eec对疾病干预的探索和发展有很大兴趣。然而,由于体外培养eec的能力相对缺乏,以及eec的分散分布和eec在肠上皮中的相对稀缺性(1%),肠内分泌细胞的研究受到阻碍。特别是,控制eec分化和功能的信号的知识在很大程度上是未知的。此外,eec的直接体外研究还不可能,因为它们是不分裂的末端分化的细胞。因此,由于eec在肠道上皮中的分散分布和稀缺性(1%),难以原位研究eec的功能和调节(sternini,anselmi和rosengut,2008,以及gunawerdene,corfe和staton,2011)。因此,仍然需要体外eec培养物,以允许研究代谢和消化疾病。还需要调节eec的功能并获得特定的eec亚型的能力,以例如用于疾病状态的发现和实施治疗。技术实现要素:本公开内容的方面包括:通过用上调chga并促进所述细胞分化形成所述肠内分泌细胞的多种小分子处理哺乳动物的出生后细胞群(例如出生后干细胞群),从而从哺乳动物的出生后细胞群(例如出生后干细胞群)获得肠内分泌细胞群(eec)。chga的上调使得所获得的细胞群中通过chga免疫染色检测所测定的表达cga的细胞分数为至少约1.5%。能够用于使出生后干细胞分化为肠内分泌细胞的小分子可包括以下中的至少一种:wnt激活剂、notch抑制剂、wnt抑制剂、mek/erk抑制剂、生长因子、hdac抑制剂、组蛋白甲基化抑制剂、tgf-β抑制剂和neurod1激活剂。本公开内容的方面还包括:通过用诱导细胞以增加胰岛素表达的多种小分子处理哺乳动物细胞群,来增加该群的胰岛素表达的方法。可以在细胞群中增加胰岛素表达水平,从而使得在使用胰岛素-gfp报告物的胰岛素活性检测中,胰岛素gfp报告物被激活的细胞的分数为至少约1%。可以进行处理以增加胰岛素表达的哺乳动物细胞可以包括出生后细胞或肠内分泌细胞,例如出生后干细胞、出生后多能子代细胞和肠内分泌细胞。可用于处理以增加细胞中胰岛素表达的小分子可包括dna甲基化抑制剂、tgf-β抑制剂和neurod1激活剂。根据本公开内容的一个方面,还包括了以本文所述的eec和/或胰岛素产生细胞治疗疾病状态的方法和/或组合物。此外,根据本公开内容的一个方面,还包括对应于所获得的eec和/或胰岛素产生细胞的细胞群,以及包含用于制备eec和/或胰岛素产生细胞的小分子的试剂盒。附图说明从以下对本发明的示例实施方式的更具体的描述,前述内容将变得显而易见,如附图中所示,其中相似的附图标记在不同图中指代相同的部分。附图不一定是按比例的,而是强调用于说明本发明的实施方式。图1显示利用notch抑制剂增加了从isc向肠内分泌细胞(eec)的分化。图2显示在所培养的肠干细胞中chga的mrna表达。图3显示了增加eec分化的小分子的96孔筛选平台。图4显示了小分子筛选结果中的阳性命中。图5显示利用小分子和在end条件下分化的肠干细胞对阳性命中的验证。图6显示mapk/erk或egfr抑制剂特异性增加eec分化。图7显示小分子吉非替尼(ge)、as703026(as)或pd0325901(pd)在eec分化过程中降低ngn3表达,并且r-spondin1促进ngn3表达。图8显示优化的分化方案。图9显示在eec分化过程中第24h时的ngn3表达。图10显示分化过程中第3天eec的关键标志物的表达。图11显示在eec分化过程中关键基因的时间进程研究。图12显示5天分化后eec标志物chga表达增加的2步方案。图13显示通过5天分化后对eec标志物chga进行染色而显示eec分化增加的2步方案。比例尺:50μm。图14显示另外的小分子对分化方案的改善。图15显示当在分化方案的第1步中加入时杜巴他丁(tubastatin)a(tu)增加eec分化。图16显示当在分化的两个步骤中加入时反苯环丙胺增加eec分化。图17显示去除egf进一步增加eec的分化。图18显示改善的分化方案。图19显示从isc向eec的高效分化。比例尺:20μm。图20显示从isc产生的功能性l细胞(glp-1)。图21显示另外的因子,包括wnt-c59和isx-9及其组合,增加eec分化。图22显示在多种条件下功能性eec标志物的表达。图23显示从isc分化为eec的分化方案。图24显示5-aza(5)、616452(6)和wnt-c59(c)的组合,在第5天诱导分化的肠干细胞中胰岛素-gfp表达。图25显示在第5天5-aza在诱导胰岛素-gfp表达中的剂量响应。图26显示在第5天616452在诱导胰岛素-gfp表达中的剂量响应。图27显示培养物中5天后isx-9(i)进一步增加胰岛素-gfp表达。图28显示在第5天isx-9在诱导胰岛素-gfp表达中的剂量响应。图29显示培养物中5天后在多种条件下细胞的胰岛素-gfp表达的facs分析。图30显示在第7天不以或以药物处理的细胞的gfp和明视野。图31显示基因表达数据。图32显示用低浓度(2mm)和高浓度(20mm)葡萄糖处理细胞诱导不同水平的胰岛素释放。图33显示分化方案的流程图。图34显示通过wnt和notch途径控制的体内eec分化的模型。图35显示如通过针对chga的免疫染色所确定的,notch和egfr/mek/erk抑制以及wnt失活(通过r-spondin1撤回)的组合诱导isc向eec方向特化。图36显示从所述组合中去除egf和/或noggin诱导了更高水平的chga表达,而添加wnt途径抑制剂iwp2(或i)或gsk3β抑制剂chir99021(或c)没有进一步增加向eec的分化。图37显示在多种分化条件下细胞集落的形态。图38显示在多种分化条件下细胞集落的形态。注意在notch、mek抑制和wnt失活(d.pd条件)以及notch/mek/wnt抑制(d.pd.c59条件)的条件下具有死细胞的集落的突出的腔。而2步分化条件(enrd-end.pd)诱导了较少的细胞死亡。图39显示另外的小分子增加eec分化。图39a.在第1步中添加或不添加杜巴他丁a(tu)的多种条件下分化的细胞的gip、gcg、cck的mrna表达。图39b.在第1步、第2步或两步中添加或不添加反苯环丙胺(tc)的多种条件下分化的细胞的gip、gcg、cck的mrna表达。图39c.反苯环丙胺的eec标志物(chga,gcg,gip,tph1)的剂量依赖性诱导。图39d.在多种条件下分化的细胞的多种eec标志物(gip,gcg,cck)的mrna表达。图39e.在多种条件下分化的细胞的lyz1的mrna表达。图40a-40d显示方案优化导致从isc向eec的高效分化。图40a.在多个时间点添加或不添加wnt途径抑制剂wnt-c59(c59)和tgf-β途径抑制剂repsox(rep)的杯状细胞(muc2)、潘氏细胞(lyz1)和eec(chga、gcg、gip、cck、tph1、sst、sct和pyy)的标志物的mrna表达。s1、s2表示2步分化方案的第1步和第2步。0h或12h提示加入c59的时间点。enr用作自发分化的对照。图40b.2步分化方法中多基因的时间进程研究。图40c.与它们相应的峰值水平相比的多种基因的表达水平(第2天的sox9、math1、ngn3和neurod1,第5天的chga,见图40d)。图40e.分化的eec细胞的免疫染色。图41a-41c显示方案优化导致从isc到eec的高效分化。图41a.具有或不具有repsox的条件下多种基因的mrna表达。所用的基础条件是enr.d.tu.tc-d.pd.tc.c59。repsox在第2步添加。图41b.所示条件下多种基因的mrna表达。图41c.分化的细胞集落的形态。箭头指示从集落中逐出的死细胞。图42显示repsox和fgf10在第1阶段添加时增加胰岛素mrna表达。图43显示更长期的5aza处理增加胰岛素mrna表达。图44显示第2阶段中wnt-c59的延迟添加增加胰岛素mrna表达。图45显示bayk8644在第1阶段添加时增加胰岛素mrna表达。图46显示bayk8644在促进胰岛素表达中的剂量依赖活性。图47显示bayk8644在第1-3阶段添加时增加胰岛素mrna表达。图48显示dapt在促进胰岛素表达中的剂量依赖活性。图49显示iox1增加胰岛素和nkx6.1表达。图50.显示repsox在促进胰岛素和nkx6.1表达中的剂量依赖活性。图51显示isx9在促进胰岛素和nkx6.1表达中的剂量依赖活性。图52显示wnt-c59在促进胰岛素和nkx6.1表达中的剂量依赖活性。图53显示dmh-1在第2-3阶段添加时增加胰岛素和nkx6.1表达水平。图54显示地塞米松在第2-3阶段添加时增加胰岛素和nkx6.1表达水平。图55显示t3在第3阶段添加时增加胰岛素mrna表达。图56显示t3和n-乙酰半胱氨酸(n-alc)在第3阶段添加时增加胰岛素mrna表达。图57显示chir99021(chir)在第3阶段添加时增加胰岛素mrna表达。图58显示艾塞那肽-4和极光激酶抑制剂ii以及佛司可林增加胰岛素mrna表达。图59.显示在促进胰岛素mrna表达时gc1能够替代t3。图60显示利用因子组合从胃肠干细胞获得的胰岛素-gfp表达和胰岛样结构的形态。图61显示细胞培养过程的图。定义在本申请中,“或”的使用是指“和/或”,除非另有说明。如在本申请中所使用的,术语“包括”以及该术语的变体,例如“包含”和“含有”不用于在本申请中排除其他添加剂、成分、整数或步骤。在本申请中,术语“约”和“近似”用作等同物。本申请中使用的具有或不具有约/近似的任何数字意指覆盖相关领域普通技术人员所理解的任何正常波动。在某些实施方式中,术语“近似”或“约”是指值的范围落入所述参考值的任一方向(大于或小于)上的25%、20%、19%、18%、17%、16%、15%、14%、13%、12%、11%、10%、9%、8%、7%、6%、5%、4%、3%、2%、1%以下范围内,除非另有说明或从上下文中明显看出(除非此类数字超过可能值的100%)。“施用”是指将物质引入受试者中。在一些实施方式中,施用是口服或通过注射进行。在某些实施方式中,“使得被施用”是指在已经施用第一组分后施用第二组分(例如,在不同的时间和/或由不同的操作者)。“抗体”是指具有免疫原结合能力的免疫球蛋白多肽或其片段。如本文所用,“激动剂”是分别引起靶基因、蛋白质或途径的表达或活性增加的试剂。因此,激动剂可以以某种方式结合并激活其同源受体,这直接或间接地对靶基因或蛋白质产生这种生理作用。激动剂还可以通过调节途径组分的活性来增加途径的活性,例如,通过抑制途径的负调节剂的活性。因此,“wnt激动剂”可以定义为增加wnt途径活性的药剂,其可以通过在细胞中增加tcf/lef介导的转录来测量。因此,“wnt激动剂”可以是真正的wnt激动剂,其结合并激活卷曲蛋白受体家族成员,包括任何和所有wnt家族蛋白,细胞内β-连环蛋白降解的抑制剂和tcf/lef的激活剂。“notch激动剂”可以定义为增加notch途径活性的药剂,其可以通过测量notch的转录活性来确定。“拮抗剂”是指与受体结合的试剂,这进而减少或消除被其他分子的结合。如本文与特定细胞类型相关使用的“细胞密度”是在代表性显微镜样品中每个区域的该细胞类型的平均数。细胞类型可以包括但不限于lgr5+细胞、肠内分泌细胞或胰岛素产生细胞。可以在给定样品、器官或组织中用给定细胞类型评估细胞密度。“细胞分化”是指细胞变得特化以执行特定功能的过程,例如出生后干细胞转化为具有更特化功能的细胞。在一个实施方式中,lgr5+肠干细胞分化为肠内分泌细胞。本文使用的“chga免疫染色检测”是用于通过免疫染色方法确定表达chga的细胞群中细胞的分数的检测。在chga免疫染色检测的实例中,已经处理了细胞群的细胞培养基被去除,并以pbs洗涤样品。在matrigel中培养的类器官或细胞集落通过加入4%pfa并在室温下温育20分钟而直接固定。然后机械破坏matrigel,细胞转移至bsa包被的eppendorf管。样品用pbs洗涤,用0.25%tritonx-100透化30分钟,并用抗嗜铬粒蛋白a的一抗(例如抗嗜铬粒蛋白a,sc-13090,santacruz)和适当的二抗(例如alexafluor缀合的二抗,例如alexafluor594缀合的驴抗兔抗体,a-21207;lifetechnologies)染色。通过共聚焦显微镜获得图像。“chir99021”是化学式为c22h18cl2n8并且有以下别名的化学成分:ct99021;6-[[2-[[4-(2,4-二氯苯基)-5-(5-甲基-1h-咪唑-2-基)-2-嘧啶基]氨基]乙基]氨基]-3-吡啶甲腈。其化学结构如下:“互补核酸序列”是指能够与由互补核苷酸碱基对组成的另一核酸序列杂交的核酸序列。如本文所用,与特定细胞类型有关的“横截面细胞密度”是在代表性显微镜样品中通过组织的每个横截面积的该细胞类型的平均数。“减少”和“降低”是指与例如参考水平相比减少至少5%,例如,5%、6%、7%、8%、9%、10%、15%、20%、25%、30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、99%或100%,并且包括与例如参考水平相比减少至少1倍,例如,1、2、3、4、5、6、7、8、9、10、15、20、30、40、50、60、70、80、90、100、200、500、1000倍或更多。“egfr抑制剂”是抑制表皮生长因子受体的物质。egfr抑制剂的实例包括厄洛替尼hcl盐(osi-744)、吉非替尼(zd1839)、拉帕替尼(gw-572016)二甲苯磺酸盐、阿法替尼(bibw2992)、来那替尼(hki-272)、卡奈替尼(ci-1033)、拉帕替尼、ag-490(tyrphostinb42)、cp-724714、达克替尼(pf299804、pf299)、wz4002、azd8931(sapitinib)、cudc-101、ag-1478(tyrphostinag-1478)、pd153035hcl、培利替尼(ekb-569)、ac480(bms-599626)、aee788(nvp-aee788)、osi-420、wz3146wz8040、ast-1306、诺司替尼(co-1686、avl-301)、金雀异黄素(genistein)、瓦力替尼(varlitinib)、埃克替尼(icotinib)、tak-285、whi-p154、pd168393、cnx-2006、酪氨酸磷酸化抑制剂(tyrphostin)9、ag-18、波齐替尼(hm781-36b)、azd3759、奥希替尼(azd9291)、阿法替尼(bibw2992)二马来酸盐、厄洛替尼、奥莫替尼(hm61713、bi1482694)、cl-387785(eki-785)、nsc228155、az5104、ag490、ag494、ag555、ag556、ag825、ag879、ag99、ap24534、av412、bibu1361、bibx1382、bms599626、卡奈替尼、cgp52411、gw583340、hds029、hki357、易瑞沙(iressa)、jnj28871063、薰草菌素a、甲基2,5-二羟基肉桂酸盐、pd158780、pf6274484、pki166、r1530、raf265、和xl184等等。“消除”意指至少降低至不可检测的水平。“肠内分泌细胞”是指细胞,其是胃肠道和胰腺的特化的内分泌细胞,并且可见于肠道、胃和胰腺。肠内分泌细胞形成体内最大的内分泌系统,并且可以感知管腔内容物,特别是营养物质,并通过调节食物摄入、能量稳态和葡萄糖耐受的多种激素(例如glp-1)的分泌来响应。肠内分泌细胞的特定类型通常根据特定肠内分泌细胞亚群内的激素的表达来分类,所述亚群例如表达glp-1、5ht、sst、胃泌素、cck、sct、nts、pyy、胃泌素和胃饥饿素(ghrelin)等的细胞。肠内分泌的不同亚群有时也被称为k细胞、i细胞、l细胞,g细胞,肠嗜铬细胞,n细胞和s细胞,但越来越多地使用细胞的激素表达来鉴定如上所述的细胞压型。可以通过chga标志物的表达来鉴定肠内分泌细胞,chga标志物可以通过检测例如本文所述的mrnachga表达检测和chga免疫染色检测来检测。“植入”或“移植”是指干细胞或祖细胞通过与组织的现有细胞接触而体内引入感兴趣组织的过程。“上皮祖细胞”是指多能细胞,其具有被限制为产生上皮细胞的细胞谱系的潜力。“上皮干细胞”是指多能细胞,其具有成为多种细胞谱系的潜力,包括产生上皮细胞的细胞谱系。“片段”是指例如多肽或核酸分子的一部分。该部分含有例如参考核酸分子或多肽的全长的至少10%、20%、30%、40%、50%、60%、70%、80%,或90%。片段可含有10、20、30、40、50、60、70、80、90或100、200、300、400、500、600、700、800、900或1000个核苷酸或氨基酸。“生长因子”是指能够刺激细胞生长、增殖或分化的物质。如本文中可互换使用的“gsk3beta”、“gsk3β”和“gsk3b”是糖原合成酶激酶3β的首字母缩略词。“gsk3β抑制剂”是抑制gsk3β活性的物质。如本文所用的“hdac”是组蛋白脱乙酰酶的首字母缩略词。“hdac抑制剂”是抑制hdac活性的物质。“组蛋白甲基化抑制剂”是抑制组蛋白甲基化活性的物质。“杂交”是指在合适的严格条件下互补核苷酸碱基之间形成双链分子的配对(例如,在dna中腺嘌呤(a)与胸腺嘧啶(t)形成碱基对,如鸟嘌呤(g)与胞嘧啶(c)形成碱基对一样)。(例如见wahl,g.m.和s.l.berger(1987)methodsenzymol.152:399;kimmel,a.r.(1987)methodsenzymol.152:507)。“抑制剂”是指引起例如靶基因、蛋白质或途径的表达或活性降低的试剂。例如,“wnt抑制剂”是指导致wnt信号传导的活性减少的试剂,其可以是例如wnt受体抑制剂、wnt受体拮抗剂、抑制wnt分泌的porcupine抑制剂,或端锚聚合酶抑制剂,或干扰β-连环蛋白相互作用的药物。“拮抗剂”可以是抑制剂,但更具体地是与受体结合的试剂,这进一步减少或消除其他分子的结合。“增加”和“提高”是指例如与参照的水平相比增加至少5%,例如,5、6、7、8、9、10、15、20、25、30、35、40、45、50、55、60、65、70、75、80、85、90、95、99、100%或更多,并且包括与参照标准品的水平相比增加至少1倍,例如,1、2、3、4、5、6、7、8、9、10、15、20、30、40、50、60、70、80、90、100、200、500、1000倍或更多。本文所用的“胰岛素活性检测”是用于确定在细胞群中胰岛素基因转录、翻译或胰岛素释放被激活的程度的检测。在示例性胰岛素活性检测中,从胰岛素-gfp小鼠例如b6.cg-tg(ins1-egfp)1hara/j小鼠(也称为mit-gfp小鼠,jackson实验室库存号:006864)肠中分离出初始细胞。从肠的近端半部分离肠隐窝。将大约200个隐窝包埋在40μlmatrigel中,并在含有500μl隐窝培养基(高级dmem/f12,具有培养基补充剂(1xn2,1xb27,2mmglutamax,10mmhepes,1mmn-乙酰半胱氨酸、和100u/ml青霉素/100μg/ml链霉素))的24孔板中培养,并补充生长因子(50ng/mlegf,100ng/mlnoggin和500ng/mlr-spondin1)和小分子(5μmchir99021和1.25mmvpa)以获得富集的肠干细胞群。然后将细胞传代1-2次以产生用于检测的起始细胞群。为了测试试剂激活胰岛素基因转录、翻译或胰岛素释放的能力,将细胞与适当的培养基(例如上述隐窝培养基)、生长因子和/或正在测试的其他试剂一起温育。将适当的培养基(包括隐窝培养基以及待评估的试剂)加入每个孔中,并与细胞一起温育2-10天,每2天更换培养基。胰岛素-gfp阳性细胞(即,其中胰岛素-gfp报告物被激活的细胞)的分数可以使用流式细胞仪来量化,以测量总细胞群中存在的gfp+细胞群的分数。此外,细胞群的平均胰岛素活性可以通过以下测量:测量使用合适的参考或管家基因(例如,使用hprt的mrna表达作为基线)标准化的群的胰岛素的平均mrna表达水平。“肠干细胞”是指肠谱系的多能细胞,其具有成为多种细胞谱系的潜力,包括产生例如肠内分泌细胞、肠细胞细胞、杯状细胞和潘氏细胞等肠细胞的细胞谱系。“分离的”是指这样的材料:在不同程度上不含有与在其天然状态下发现的通常伴随其的组分。“分离”表示与原始来源或周围环境分开的程度。“lgr5”是含有富亮氨酸重复的g蛋白偶联受体5的缩写,也称为g蛋白偶联受体49(gpr49)或g蛋白偶联受体67(gpr67)。它是人体内由lgr5基因编码的蛋白质。本文所用的“lgr5+细胞”或“lgr5阳性细胞”是表达lgr5的细胞。本文所用的“lgr5-细胞”是并非lgr5+的细胞。“哺乳动物”是指任何哺乳动物,包括但不限于人、小鼠、大鼠、绵羊、猴、山羊、兔、仓鼠、马、牛或猪。“多能子代细胞”是指已经比干细胞更具特异性的细胞,意味着它具有分化成特定类型细胞的倾向,但仍保留分化成多种不同但有限的细胞类型的能力。多能子代细胞可以是多能细胞,其已经从干细胞中分化出来,但尚未分化成“目标”细胞类型。“mek/erk抑制剂”是抑制mek/erk信号传导途径的物质。mek抑制剂的实例包括牛蒡子苷元、bix02189、10z-海门地塞(hymenialdisine)、pd0325901、pd184352、pd198306、pd334581、pd98059、sl327、u0126、司美替尼(azd6244)、曲美替尼(trametinib)(gsk1120212)、pd184352(ci-1040)、pd98059、pimasertib(as-703026)、bix02188、tak-733、azd8330、比尼替尼(binimetinib)(mek162、arry-162、arry-438162)、pd318088、来法替尼(refametinib)(rdea119、bay86-9766)、bi-847325、卡比替尼(cobimetinib)(gdc-0973、rg7420)、gdc-0623和aps-2-79。erk抑制剂的实例包括sch772984、del-22379、vx-11e、erk5-in-1、xmd8-92、sc1(pluripotin)、乌里替尼(ulixertinib)(bvd-523、vrt75227、fr180204、gdc-0994、bix02189、tcserk11e、tmcb和二十碳五烯酸。“mrnachga表达检测”是指用于确定细胞群中相对chgamrna表达水平的检测。例如,检测可以确定与未测试的出生后干细胞群相比,用待测试的试剂处理后的分化的细胞群的chgamrna表达水平。在mrnachga表达检测的实例中,收集细胞并使用rna提取试剂盒(例如rneasyminikit,qiagen)从细胞中提取rna。chga表达水平然后使用一步qpcr试剂盒(例如quantitechprobepcr试剂盒,qiagen)和chga引物和探针(例如商业上可获得的用于小鼠chga的taqman探针,lifetechnologies)通过定量实时pcr评估。“mrna胰岛素表达检测”是指用于确定细胞群的相对胰岛素mrna表达水平的检测。例如,检测可以确定与未处理的细胞群相比,在经处理以增加细胞中胰岛素表达的细胞群中表达的胰岛素mrna水平。在示例性的mrna胰岛素表达检测中,收集细胞,并使用rna提取试剂盒(例如rneasyminikit,qiagen)从细胞中提取rna。然后对胰岛素表达水平使用一步qpcr试剂盒(例如quantitechprobepcr试剂盒,qiagen)以及ins1或ins2引物和探针(例如市售用于小鼠ins1和ins2的taqman探针,lifetechnologies)通过定量实时pcr评估。相对胰岛素mrna表达可以相对于基线或其他标记物确定,例如hprtmrna表达水平或其他基线标记物。“neurod1激活剂”是激活neurod1的物质。如本文所用,“非人哺乳动物”是指任何非人的哺乳动物。“notch抑制剂”是指notch信号传导途径的抑制剂。“ngn3”指的是神经生成素-3,一种在内分泌祖细胞中表达并与肠内分泌分化相关的蛋白质。如本文所用,“ngn3检测”是用于确定细胞(例如经受了分化方案的细胞)中ngn3的表达程度的检测。在示例性ngn3检测中,从已经根据分化方案培养的细胞中分离rna,使用市售的引物和探针(例如taqman探针)进行定量实时pcr,以确定细胞中ngn3表达的程度。如本文相关上下文中所使用的,术语细胞的“数量”可以是0、1或更多细胞。“类器官”或“上皮类器官”是指细胞簇或聚集体,其类似于器官或器官的一部分,并且具有与该特定器官相关的细胞类型。细胞“群”是指大于1的任何数量的细胞,甚至至少1×103个细胞、至少1×104个细胞、至少1×105个细胞、至少1×106个细胞、至少1×107个细胞、至少1×108个细胞、至少1×109个细胞或至少1×1010个细胞。“出生后细胞”是指非胚胎细胞。出生后细胞可包括以下中的至少一种:出生后干细胞,出生后祖细胞,和出生后多能子代细胞,以及一种或多种由这些细胞分化的细胞,例如肠内分泌细胞(eecs)。“出生后干细胞”是指具有自我更新和分化成多种细胞谱系的能力的非胚胎干细胞。出生后干细胞也可称为成体干细胞或体细胞干细胞。出生后干细胞可包括肠干细胞、上皮干细胞、造血干细胞、乳腺干细胞、间充质干细胞、内皮干细胞和神经干细胞。如本文所用,“祖细胞”是指与干细胞一样具有分化成特定类型细胞的倾向的细胞,但是已经比干细胞更具特异性并被推动分化为其“目标”细胞。“参考”是指标准或对照条件(例如,未用测试试剂或测试试剂的组合处理)。术语“样品”是指获得、提供和/或进行分析的体积或质量。在一些实施方式中,样品是或包括组织样品、细胞样品、流体样品等。在一些实施方式中,样品取自(或是)受试对象(例如人或动物受试对象)。在一些实施方式中,组织样品是或包括脑、毛发(包括根)、口腔拭子、血液、唾液、精液、肌肉或来自任何内脏器官,或与以上任一种相关的癌症、癌前病变或肿瘤细胞。液体可以是但不限于尿液、血液、腹水、胸膜液和脊髓液等。身体组织可以包括但不限于脑、皮肤、肌肉、子宫内膜、子宫和子宫颈组织,或者与这些中的任何一种相关的癌症、癌前病变或肿瘤细胞。“自我更新”指的是干细胞分裂产生一个(不对称分裂)或两个(对称分裂)子细胞的过程,子细胞的发育潜力与母细胞的细胞无法区分。自我更新涉及增殖和维持不分化的状态。本文提及的“小分子”是指可以参与调节生物途径的有机化合物,并且是非核酸,通常是非肽和非寡聚物,并且可以具有小于1500道尔顿的分子量。“干细胞”是指具有自我更新和分化成多种细胞谱系能力的多能细胞。本文所用的“干细胞标志物”可以定义为在干细胞中特异性表达的基因产物(例如蛋白质,rna等)。一种类型的干细胞标志物是直接和特异性地支持干细胞特性维持的基因产物。实例包括lgr5和sox2。另外的干细胞标志物可以使用文献中描述的检测来鉴定。为了确定是否需要该基因来维持干细胞特性,可以使用功能获得和功能丧失研究。在功能获得研究中,特定基因产物(干细胞标志物)的过表达将有助于维持干细胞特性。而在功能丧失研究中,去除干细胞标志物会导致干细胞特征的丧失或诱导干细胞分化。另一种类型的干细胞标志物是仅在干细胞中表达的基因,但不一定具有维持干细胞特性的特定功能。这种类型的标志物可通过如微量阵列和qpcr等检测比较分选的干细胞和非干细胞的所述基因表达特征来鉴定。这种类型的干细胞标志物可以在文献中找到(例如liuq.等,intjbiochem,;60:99-111.http://www.ncbi.nlm.nih.gov/pubmed/25582750)。潜在干细胞标志物包括ccdc121、gdf10、opcm1、phex等。在给定的细胞或细胞群中,干细胞标志物例如lgr5或sox2的表达可以用例如qpcr、免疫组织化学、蛋白质印迹和rna杂交等检测来测量。干细胞标志物的表达也可以使用表达报告物的转基因细胞来测量,报告物可以指示给定干细胞标记物的表达,例如lgr5-gfp或sox2-gfp。然后可以使用流式细胞术分析来测量报告物表达的活性。荧光显微镜也可用于将报告物的表达直接可视化。干细胞标志物的表达可以通过微阵列分析确定,以用于全局基因表达谱分析。可以将给定细胞群或纯化细胞群的基因表达谱与干细胞的基因表达谱进行比较,以确定2个细胞群之间的相似性。干细胞功能可以通过集落形成检测或球形成检测、自我更新检测和分化检测测定。在集落(或球体)形成检测中,当在适当的培养基中培养时,干细胞应该能够在细胞培养物表面(例如细胞培养皿)中形成集落或包埋在细胞培养物基质(例如matrigel),或当在悬浮液中培养时能够形成球体。在集落/球形成检测中,在适当的培养基中以低细胞密度接种单个干细胞,并允许增殖一段时间(7-10天)。然后对形成的集落计数并对干细胞标志物表达进行评分,作为原始细胞的干细胞性的指示。任选地,然后挑选和传代形成的集落,以测试它们的自我更新和分化潜力。在自我更新检测中,当在适当的培养基中培养时,细胞应保持干细胞标志物(例如,lgr5)表达超过至少1次(例如,1次、2次、3次、4次、5次、10次、20次等)细胞分裂。“受试对象”包括人和哺乳动物(例如,小鼠、大鼠、猪、猫、狗和马)。在许多实施方式中,受试对象是哺乳动物,特别是灵长类动物,尤其是人。在一些实施方式中,受试对象是牲畜例如牛、绵羊、山羊、牛、猪等;家禽例如鸡、鸭、鹅、火鸡等;和驯养的动物,特别是宠物,例如狗和猫。在一些实施方式中(例如,特别是研究背景下)受试对象哺乳动物是例如啮齿动物(例如小鼠、大鼠、仓鼠)、兔子、灵长类动物或猪,如近交系猪等。“协同”或“协同效果”是大于单独取得的每种效果的总和的效果;大于加成效果。“tgfβ抑制剂”(tgf-β抑制剂)如本文所用,是降低tgfβ途径活性的物质。tgfβ抑制剂的实例可以是tgfβ受体抑制剂,其可以包括但不限于alk4、alk7和alk5/tgfβ-ri。“组织”是来自相同来源的类似细胞的集合,其共同执行特定功能。如本文所用与细胞群相关的“处理”意指递送物质至该群以得到结果。在体外群体的情况下,该物质可以直接(或甚至间接)递送至群。在体内群的情况下,该物质可以通过施用至主体受试对象来递送。“丙戊酸”(vpa)具有化学式c8h16o2和以下替代名称:2-丙基戊酸。丙戊酸的钠盐也可用于代替vpa,并且术语“vpa”在本文中可互换使用指代vpa或其药学上可接受的盐,例如钠盐。其化学结构如下:如本文所用,与物质或组合物有关的“wnt激活”是wnt信号传导途径的激活。如本文所用,“wnt激活剂”是指激活wnt信号传导途径的物质。如本文所用,“wnt抑制剂”是指抑制wnt信号传导途径的物质,其可以是例如wnt受体抑制剂,wnt受体拮抗剂,抑制wnt分泌的porcupine抑制剂,或端锚聚合酶抑制剂,或干扰β-连环蛋白相互作用的药物。wnt抑制剂的实例包括wnt-c59、iwp-2、iwr-1-endo、az6102、fh535、wiki4、icg-001、xav-939、pri-724、lgk-974、ya1797k、ky02111、心肌原(cardionogen)1和iwp12等。本文使用的术语“药学上可接受的”是指那些在合理的医学判断范围内适合用于与人类和动物的组织接触使用而不具有过度的毒性、刺激、过敏反应或其他问题或并发症(与合理的利益/风险比相称)的那些化合物、材料、组合物和/或剂型。如本文所用,“药学上可接受的载体、稀释剂或赋形剂”包括但不限于任何佐剂、载体、赋形剂、助流剂、甜味剂、稀释剂、防腐剂、染料/着色剂、增味剂、表面活性剂、润湿剂、分散剂、悬浮剂、稳定剂、等渗剂、溶剂、表面活性剂或乳化剂,其已经美国食品药品监督管理局批准可用于人类或家畜。药学上可接受的载体的实例包括但不限于糖,例如乳糖、葡萄糖和蔗糖;淀粉,例如玉米淀粉和马铃薯淀粉;纤维素及其衍生物,例如羧基甲基纤维素钠、乙基纤维素和纤维素醋酸盐;黄蓍胶;麦芽;明胶;滑石粉;可可脂、蜡、动物和植物脂肪、石蜡、硅酮、膨润土、硅酸、氧化锌;油,例如花生油、棉籽油、红花油、芝麻油、橄榄油、玉米油和大豆油;二醇,例如丙二醇;多元醇,例如甘油、山梨糖醇、甘露醇和聚乙二醇;酯,例如油酸乙酯和月桂酸乙酯;琼脂;缓冲剂,例如氢氧化镁和氢氧化铝;海藻酸;无热原水;等渗盐水;林格氏溶液;乙醇;磷酸盐缓冲溶液;和药物制剂中使用的任何其他相容物质。“药学上可接受的盐”包括酸和碱加成盐。“药学上可接受的酸加成盐”是指那些保留游离碱的生物有效性和性质的盐,其在生物学上或其它方面不是不期望的,并且由以下形成:无机酸,例如但不限于盐酸、氢溴酸、硫酸、硝酸、磷酸等,和有机酸,例如但不限于乙酸、2,2-二氯乙酸、己二酸、海藻酸、抗坏血酸、天冬氨酸、苯磺酸、苯甲酸、4-乙酰氨基苯甲酸、樟脑酸、樟脑-10-磺酸、癸酸、己酸、辛酸、碳酸、肉桂酸、柠檬酸、环己烷氨基磺酸、十二烷基硫酸、乙烷-1,2-二磺酸酸、乙磺酸、2-羟基乙磺酸、甲酸、富马酸、半乳糖二酸、龙胆酸、葡庚糖酸、葡萄糖酸、葡萄糖醛酸、谷氨酸、戊二酸、2-氧代戊二酸、甘油磷酸、乙醇酸、马尿酸、异丁酸、乳酸、乳糖酸、月桂酸、马来酸、苹果酸、丙二酸、扁桃酸、甲磺酸、粘酸、萘-1,5-二磺酸、萘-2-磺酸、1-羟基-2-萘甲酸、烟酸、油酸、乳清酸、草酸、棕榈酸、双羟萘酸、丙酸、焦谷氨酸、丙酮酸、水杨酸、4-氨基水杨酸、癸二酸、硬脂酸、琥珀酸、酒石酸、硫氰酸、甲苯磺酸、三氟乙酸、十一碳烯酸等。“药学上可接受的碱加成盐”是指保留游离酸的生物有效性和性质的那些盐,其在生物学上或其它方面不是不期望的。这些盐是通过在游离酸中加入无机碱或有机碱制备的。衍生自无机碱的盐包括但不限于钠、钾、锂、铵、钙、镁、铁、锌、铜、锰、铝盐等。例如,无机盐包括但不限于铵盐、钠盐、钾盐、钙盐和镁盐。衍生自有机碱的盐包括但不限于以下物质的盐:伯胺、仲胺和叔胺、取代的胺,包括天然存在的取代的胺,环胺和碱性离子交换树脂,例如氨、异丙胺、三甲基胺、二乙基胺、三乙基胺、三丙基胺、二乙醇胺、乙醇胺、癸醇、2-二甲基氨基乙醇、2-二乙基氨基乙醇、二环己胺、赖氨酸、精氨酸、组氨酸、咖啡因、普鲁卡因、哈胺(hydrabamine)、胆碱、甜菜碱、苯乙苄胺(benethamine)、苄星(苄星青霉素)、乙基二胺、葡糖胺、甲基葡糖胺、可可碱、三乙醇胺、氨丁三醇、嘌呤、哌嗪、哌啶、n-乙基哌啶、多胺树脂等。在某些实施方式中使用的有机碱的实例包括异丙胺、二乙基胺、乙醇胺、三甲基胺、二环己胺、胆碱和咖啡因。在组合物(例如,药物组合物)中也可存在润湿剂、乳化剂和润滑剂,例如十二烷基硫酸钠和硬脂酸镁,以及着色剂、脱模剂、包衣剂、甜味剂、调味剂和芳香剂、防腐剂和抗氧化剂。药学上可接受的抗氧化剂的实例包括:(1)水溶性抗氧化剂,例如抗坏血酸、盐酸半胱氨酸、硫酸氢钠、偏亚硫酸氢钠、亚硫酸钠等;(2)油溶性抗氧化剂,例如抗坏血酸棕榈酸酯、丁基化羟基苯甲醚(bha)、丁基化羟基甲苯(bht)、卵磷脂、没食子酸丙酯、α-生育酚等;和(3)金属螯合剂,如柠檬酸、乙二胺四乙酸(edta)、山梨糖醇、酒石酸、磷酸等。具体描述以下是对本公开的示例性实施方式进行的描述。本发明涉及通过用能够使导致出生后细胞群分化为eec的途径和机制激活和/或失活的多种小分子处理哺乳动物的出生后细胞群(例如出生后干细胞群)从而由出生后细胞群(例如出生后干细胞群)形成肠内分泌细胞(eecs)的方法。小分子作用的途径和机制可包括但不限于wnt信号传导途径、notch信号传导途径、egfr途径、mek/erf信号传导途径、通过生长因子作用的机制、组蛋白甲基化途径,tgf-β信号传导途径和neurod1途径。特别地,本公开的方面提供了可以促进出生后细胞例如出生后干细胞的分化为eec的小分子组合,以及将分化成特定的eec细胞类型的信号。本公开还涉及通过用多种小分子处理群来增加哺乳动物细胞群中胰岛素表达的方法,所述多种小分子能够使导致细胞上的胰岛素表达增加的途径和机制激活和/或失活。由小分子处理的细胞可以是出生后细胞(例如出生后干细胞或多能子代细胞)或肠内分泌细胞,例如通过如上所述的方法已经从出生后细胞例如出生后干细胞分化为的eec。由小分子起作用增加胰岛素生产的途径和机制可以包括但不限于上面针对eec分化所讨论的任何信号传导途径,例如wnt和notch信号传导途径,特别是包括发挥作用以抑制和/或激活dna甲基化途径、tgf-β信号传导途径和neurod1途径的小分子。特别地,本公开的方面提供促进细胞中胰岛素表达,并在所处理群中诱导细胞成为胰岛素产生细胞的小分子组合。在出生后细胞例如出生后干细胞分化为eec时,提供了多种小分子,从而使得分化标志物chga的表达由于细胞的分化而被上调。chga的表达被上调,从而使得用小分子处理的细胞群中表达chga的细胞分数在chga免疫染色检测中测量为至少约1.5%。通过chga免疫染色检测测量的表达chga的细胞分数甚至可以是至少约10%,和甚至至少约50%,例如细胞的分数在约60%至约100%的范围内。在一个实施方式中,表达chga的细胞分数可以在约1.5%至约100%的范围内。此外,如通过mrnachga表达检测所测定的,在分化的细胞群中的chgamrna表达可以是初始的出生后细胞群中chgamrna表达的至少约10倍。chgamrna表达甚至可以是初始的出生后细胞群中chgamrna表达的至少100倍,并且甚至是初始的出生后细胞群中chgamrna表达的约1000倍至约1,000,000倍的范围内。在一个实施方式中,chgamrna表达可以是初始的出生后细胞群中chgamrna表达的约10倍至约1,000,000倍的范围内。使用小分子使细胞分化形成eec可以提供高纯度的肠内分泌细胞群,其中,相当数量的出生后细胞,例如出生后干细胞转化为肠内分泌细胞。这种将出生后干细胞大量转化为eec的能力很重要,因为肠eec受限于粘膜,主要在其较深的一半中,并且通常仅占整个上皮细胞群的一小部分,例如小于1%。相比之下,用如本文所述的小分子处理将出生后细胞转化为肠内分泌细胞,使得表达针对eec的chga标志物的细胞百分比为总细胞群的至少2%,甚至高达细胞群的90%至甚至100%,例如总细胞群的约60%至约90%,甚至总细胞群的约70%至约80%。如本文所公开的,已经实现具有约80%eec的最终细胞群(参见例如图40)。在一个实施方式中,在所得的细胞群中,表达chga的细胞百分比与初始出生后细胞群是增加的,chga表达细胞的分数增加至少0.1%至至少1%,甚至增加至少约5%,例如增加至少约10%,和甚至增加至少约50%,例如至少约100%。由小分子处理的出生后细胞包括出生后干细胞,其是非胚胎干细胞例如成体干细胞,其具有自我更新和分化成多个细胞谱系的能力,并且还可以包括多能子代细胞和/或祖细胞。出生后干细胞可以包括肠干细胞、上皮干细胞、造血干细胞、乳腺干细胞间、间充质干细胞、内皮干细胞和神经干细胞,其通常具有保守信号传导和发育途径。在一个实施方式中,出生后干细胞是肠干细胞,并且可以通过标志物lgr5鉴定。lgr5是包含富亮氨酸重复的g蛋白偶联受体5,也称为g蛋白偶联受体49(gpr49)或g蛋白偶联受体67(gpr67),和在人类中由lgr5基因编码。当用增加胰岛素表达的小分子处理细胞时,被处理的细胞可以是这些出生后干细胞和/或已经进一步特化的细胞,例如多能子代细胞和/或祖细胞和/或肠内分泌细胞。也可以进行用多种小分子处理细胞群以提供细胞的胰岛素表达的显著的增加。在一个实施方式中,通过用小分子处理使胰岛素表达增加,使得如胰岛素活性检测所测量的,在处理的细胞中胰岛素gfp报告物激活的细胞的分数为至少约1%,甚至至少约20%。在一个实施方式中,胰岛素gfp报告物激活的细胞分数为约20%至约100%。根据不同类型的测量,如通过mrna胰岛素表达检测所确定的,用多种小分子处理可能能够相对于初始细胞群增加胰岛素表达,增加至显著大于基线水平的水平。例如与hprtmrna(作为qpcr中表达水平的参考基因)相比,mrna胰岛素表达检测可以提供mrna胰岛素表达水平,其与使用hprtmrna水平(其中胰岛细胞的胰岛素水平通常是hprt水平的约100倍)的标准相比为1倍和甚至至少约2倍,从而证明细胞成功地转化为表达胰岛素的细胞。如本文图31中所示,当在qpcr中对干细胞样品设定任意对照ct值时,与干细胞中的表达相比,可以实现胰岛素表达2x107倍增加(在胰岛素mrna在干细胞样品中在设定ct值处或之上检测不到的情况下)。此外,通过mrna胰岛素表达检测测量的胰岛素mrna表达水平与胰岛细胞中的胰岛素mrna水平相比,被认为提供了与胰岛相比至少约0.01%,例如至少约0.1%,设置至少约1%,高达至少约10%,甚至100%或200%的胰岛素mrna活性。根据一个实施方式,使细胞群接受多种小分子处理可以提供这样的所得细胞群:约1%的所获得的细胞是胰岛素产生细胞,并且所获得的细胞群中的胰岛素产生细胞的数量甚至可为总细胞群的约1%至约20%。也可以进行用多种小分子对eec群的处理以产生子群,所述子群包括例如l-细胞、k-细胞、i-细胞、g-细胞、ec-细胞、n-细胞、和s-细胞。由小分子处理的哺乳动物细胞群可以是体外群,例如分散在细胞培养基中的细胞群。体外细胞群也可以是类器官的形式,它是类似器官或器官的一部分的细胞簇或聚集体,并且具有与该特定器官相关的细胞类型(sato,2009)。作为选择,由小分子处理的哺乳动物细胞群可以是体内群,例如治疗疾病状态的人或者其他哺乳动物的体内治疗。在另一个替代方案中,由小分子处理的哺乳动物细胞群可以是离体群,例如从人或其他哺乳动物(例如,需要治疗的人或其他哺乳动物)获得(例如,分离,衍生)的细胞群。图34中提供了用于从出生后细胞例如出生后干细胞分化出eec的一个实施方式的提出机制。不希望受到任何特定理论的限制,但目前认为通过控制wnt和notch途径,和可选的其他途径,可以从干细胞中特化出eec。特别地,在所示的机制中提供激活wnt途径同时抑制notch途径,然后同时抑制notch和wnt途径以形成eec。如下所述,也可以激活和/或抑制其他机制和/或途径,从而进一步指定沿着途径向eec分化(与杯状细胞、肠细胞、潘氏细胞等相反),并且其他机制和/或途径可以也可以用于进一步特化特定类型的eec,和/或eec功能。例如,虽然图34显示wnt抑制,但在一些实施方式中,可以无需添加wnt抑制剂就提供向eec的分化。而且,虽然图34示出了用于获得eec的两阶段方法,但是某些实施方式还可以提供仅单阶段处理,或甚至3个或更多个阶段处理以实现eec。可以特化的特定类型的eec可以包括表达glp-1、5ht、sst、胃泌素、cck、sct、nts、pyy和胃饥饿素等的细胞。在如本文所述的增加哺乳动物细胞的胰岛素产生的方法的实施方式中,哺乳动物细胞的处理可导致沿着与图34中所示类似的途径分化,例如从出生后干细胞至eec,得到的细胞是那些提供增加胰岛素生产的细胞,和/或可以对eec本身进行处理,从而将这些细胞转化为胰岛素生产增加的细胞。也就是说,增加细胞胰岛素生产的处理可以在出生后干细胞分化为eec的同时进行、之前或之后进行,或者可以对在干细胞和eec之间的分化阶段的细胞进行,以及对出生后干细胞或eec本身进行。因此,增加胰岛素生产的处理可以对包括出生后干细胞(例如lgr5+细胞)、祖细胞和分化的肠内分泌细胞中的至少一种的哺乳动物细胞群进行。图34还显示了不同类型的肠细胞,例如eec、肠细胞、杯状细胞、潘氏细胞和干细胞,以及在肠的隐窝中发现的所述细胞的一般构造。该图显示,如上所述,肠内分泌(eec)细胞通常位于这样的隐窝中,并且仅以总细胞的一小部分(<1%)存在于其中,因此在没有获得细胞的eec群的体外方法的情况下,难以测试疾病治疗的机制。如上所述,在一个实施方式中,为了对出生后细胞群(例如出生后干细胞群)进行处理以实现向eec的分化,和/或对细胞群进行处理以增加其胰岛素表达而提供的所述多种小分子包括激活和/或抑制wnt途径的小分子。wnt信号传导途径是通过wnt蛋白配体与卷曲蛋白家族受体结合而激活的信号转导途径。可以作为处理的一部分而提供的wnt激活剂包括例如以下中的至少一种:r-spondin1、wnt3a、gsk抑制剂,例如至少一种chir99021、ly2090314、np031112(tideglusib)、锂、a1070722、skl2001,和其他能够激活wnt的药剂。在一个实施方式中,作为多种小分子的一部分提供的wnt激活剂可以是r-spondin1,其是属于r-spondin家族的蛋白质。可以作为治疗的一部分提供的wnt抑制剂可以包括例如以下中的至少一种:wnt-c59、dkk家族蛋白质(例如dkk-1、dkk2、dkk3和dkk4中的至少一种)、sfrp(例如frp-1和sfrp-2中的至少一种)、抗wnt受体的抗体例如omp-18r5、其他小分子wnt抑制剂例如lgk974、cwp232291、pri-724、iqr-1、iwp2、iwp-l6、icg-001、wiki4、ky02111、fh535、xav939、nsc668036、fj9、3289-8625,等等(kahn,2014)。在一个实施方式中,作为用于处理的多种小分子的一部分提供的wnt抑制剂可以是wnt-c59,其具有以下化学结构:还可提供wnt激活剂和/或wnt抑制剂的衍生物和/或药学上可接受的盐。可以适合作为wnt激活剂的wnt激动剂的其他实例可以在下表中找到。表-wnt激动剂在一个实施方式中,为了对出生后细胞群(例如出生后干细胞群)进行处理以实现向eec的分化,和/或对细胞群进行处理以增加其胰岛素表达而提供的所述多种小分子包括起作用以抑制notch途径的小分子。可作为治疗的一部分提供的notch抑制剂可包括例如以下中的至少一种:dapt;ly411575;mdl-28170;ro4929097;l-685458((5s)-(叔丁氧基羰基氨基)-6-苯基-(4r)羟基-(2r)苄基己酰基)-l-亮氨基-l-苯并氨酸-酰胺);bms-708163(avagacestat);bms-299897(2-[(1r)-1-[[(4-氯苯基)磺酰基](2,5-二氟苯基)氨基]乙基-5-氟苯丁酸);m-0752;yo-01027;mdl28170(sigma);ly411575(n-2((2s)-2-(3,5-二氟苯基)-2-羟基乙酰基)-n1-((7s)-5-甲基-6-氧代-6,7-二氢-5h-二苯并[b,d]氮杂卓-7-基)-1-丙氨酰胺);eln-46719(ly411575的2-羟基-戊酸酰胺类似物;pf-03084014((s)-2-((s)-5,7-二氟-1,2,3,4-四氢萘-3-基氨基)-n-(1-(2-甲基-1-(新戊基氨基)丙-2-基)-1h-咪唑-4-基)戊酰胺);化合物e((2s)-2-{[(3,5-二氟苯基)乙酰基]氨基}-n-[(3s)-l-甲基-2-氧代-5-苯基-2,3-二氢-1h-1,4-苯并二氮杂-3-基]丙酰胺;和semagacestat(ly450139;(2s)-2-羟基-3-甲基-n--((1s)-1-甲基-2-{[(1s)-3-甲基-2-氧代-2,3,4,5-四氢-1h-3-苯并氮杂-1-基]-氨基}-2-氧代乙基)丁酰胺)。在一个实施方式中,作为多种小分子的一部分提供的notch抑制剂可以是dapt,也称为n-[n-(3,5-二氟苯乙酰基)-l-丙氨酰基]-s-苯基甘氨酸叔丁酯。还可提供notch抑制剂的衍生物和/或药学上可接受的盐。为处理而提供的多种小分子还可以包括作用以抑制mek/erk途径的小分子。这可以包括核心ras-raf-mek-erk信号传导级联的抑制剂,以及该核心信号传导级联的上游或下游的蛋白质的抑制剂,例如egf受体(egfr)抑制剂。为处理而提供的mek/erk抑制剂可包括例如以下中的至少一种:pd0325901、azd8330(arry-424704)、来法替尼(bay86-9766、rdea119)、卡比替尼(gdc-0973、xl-518、rg7421)、e6201、mek162(arry-438162)、pimasertib(as703026、msc1936369b)、ro4987655(ch4987655)、ro5126766(ch5126766)、司美替尼(azd6244、arry-142,886)、tak-733、曲美替尼(gsk1120212)、和gdc-0623、wx-554(zhao和adjei、2014),并且也可以和/或替代地包括egfr抑制剂,例如厄洛替尼、吉非替尼、拉帕替尼、阿法替尼、来那替尼、az5104、阿法替尼、pd153035、cl-387785、ast-1306、pd168393、卡奈替尼、和其他egfr抑制剂,以及ras和raf抑制剂。在一个实施方式中,作为多种小分子的一部分提供的mek/erk抑制剂可以是pd0325901,也称为n-[(2r)-2,3-二羟基丙氧基]-3,4-二氟-2-[(2-氟-4-碘苯基)氨基]-苯甲酰胺。还可提供mek/erk抑制剂的衍生物和/或药学上可接受的盐。为处理而提供的多种小分子还可以包括对应于各种生长因子(例如表皮生长因子(egf)和noggin中至少一种)的小分子。在一个实施方式中,作为为处理而提供的多种小分子的一部分包括egf和/或noggin。还可提供生长因子的衍生物和/或药学上可接受的盐。在另一个实施方式中,多种小分子可以包括hdac抑制剂(组蛋白脱乙酰抑制剂)。可以作为为处理而提供的多种小分子的一部分的hdac抑制剂可包括杜巴他丁a、acy1215、丙戊酸、saha、曲古抑菌素a、shba、cbha、laq-824、pdx-101、lbh-589、itf2357、pci-24781、化合物7(chemietek)、jnk-24681585(quisinostat)sb939(pracinostat)、4sc-201(resminostat)、tefinostat(chr-2845)、chr-3996、cg200745、缩酚酸肽(罗米地辛)、丁酸酯、ms-275、mgcd0103和ci994等。在一个实施方式中,为进行处理而提供的hdac抑制剂可以是杜巴他丁a,也称为n-羟基-4-(2-甲基-1,2,3,4-四氢-吡啶并[4,3-b]吲哚-5-基甲基)苯甲酰胺盐酸盐。还可以提供hdac抑制剂的衍生物和/或药学上可接受的盐。hdac抑制剂的其他实例可见于下表中。表-hdac抑制剂可以作用用于处理的多种小分子的一部分提供的组蛋白甲基化抑制剂(例如,组蛋白去甲基酶(hdm)抑制剂)可包括以下中的至少一种:反苯环丙胺、gsk-2879552、gsk-lsd1、sp-2509、gskj4、2,4-吡啶二羧酸、ml324、iox1、og-l002、cbb1007、gskj1、gskj2、和gskj5等等。在一个实施方式中,为了处理而提供的组蛋白甲基化抑制剂可以是反苯环丙胺。在实施方式中,为了处理而提供的组蛋白甲基化抑制剂可以是赖氨酸特异性组蛋白去甲基酶(lsd1)。还可以提供组蛋白甲基化抑制剂的衍生物和/或药学上可接受的盐。组蛋白甲基化抑制剂的实例包括:jmjc-结构域去甲基酶:jmjd2、jmjd2c、jmjd3;赖氨酸特异性去甲基酶:lsd1抑制剂,例如反苯环丙胺(lsd1)、rn1(lsd1)、gsk2879552(lsd1)、cbb1003(lsd1)、og-l002(lsd1)、cbb1007(lsd1)、2,4-吡啶二羧酸(lsd)、sp2509(lsd1)、ory-1001(rg-6016)、gsklsd1(lsd1);jmjd:丁酰肼(jmjd)、gskj1(jmjd3/utx)、gskj4(jmjd3)、iox1(jmjd)、jib04(jmjd)、nsc636819(kdm4a/kdm4b)、tc-e5002(kdm2/7)、巴吉林(pargyline)、ml324(jmjd2);可以作为用于处理的多种小分子的一部分提供的tgf-β抑制剂可包括以下中的至少一种:616452(repsox)、ly-364947、sb-505124、a-83-01、sb-431542、tgf-βri激酶抑制剂vii、sb-525334、tgf-βri激酶抑制剂ix、gw788388、ly2109761、galunisertib(ly2157299)、ew-7197、吡非尼酮、k02288、d4476、r268712、a77-01、和sm16以及针对tgf-β受体的抗体。在一个实施方式中,为处理而提供的tgf-β抑制剂可以是616452,也称为repsox,化学式为2-[3-(6-甲基-2-吡啶基)-1h-吡唑-4-基]-1,5萘啶。还可以提供组蛋白甲基化抑制剂的衍生物和/或药学上可接受的盐。tgf-β抑制剂的其他实例可见于下表。tgf-β抑制剂可以作为提供用于处理的多种小分子的一部分提供的neurod1激活剂可以包括isx-9和其他异噁唑分子(schneider,2008)。在一个实施方式中,提供用于处理的neurod1激活剂可以是isx-9,化学式为n-环丙基-5-(2-噻吩基)-3-异噁唑甲酰胺。还可以提供neurod1激活剂的衍生物和/或药学上可接受的盐。提供用于处理的的多种小分子还可包括一种或多种单胺氧化酶(mao)抑制剂。mao抑制剂的实例包括沙芬酰胺甲磺酸盐、雷萨吉兰甲磺酸盐、反苯环丙胺、吗氯贝胺(ro111163)、靛红、8-(3-氯苯乙烯基)咖啡因、二苯美仑盐酸盐、(r)-(-)-苄甲炔胺盐酸盐、哈尔满碱(harmane)、拉扎贝胺盐酸盐、吡吲哚甲磺酸盐、rn1二盐酸盐、和特吲哚甲磺酸盐(tetrindolemesylate)等等。为处理而提供的多种小分子可以还包含一种或多种bmp受体(包括alk2)抑制剂。bmp受体(包括alk2)抑制剂的实例包括dmh-1、dmh2、多索莫芬(dorsomorphin)、k02288、ldn193189和ml347。在处理细胞以增加胰岛素表达的过程中,已显示有效增加细胞的胰岛素表达的多种小分子是包含dna甲基化抑制剂、tgf-β抑制剂和neurod1激活剂中至少一种的多种小分子。在一个实施方式中,所有三种小分子一起作为治疗的一部分提供,以增加细胞群中胰岛素的产生。tgf-β抑制剂和neurod1激活剂可以是任何上面讨论的那些,和/或它们的衍生物和/或其药学上可接受的盐,特别是可以对应于616452和isx-9。616452(repsox)的结构如下:dna甲基化抑制剂可包含以下中的至少一种:5-氮杂胞苷、5-氮杂-2-脱氧胞苷、rg108、sgi1027、七尾霉素(nanaomycin)a、泽布拉林(zebularine)、洛美谷里(lomeguatrib)、sgi-110和七尾霉素c等。在美国专利no.8,207,142、加拿大专利no.2,454,147,和wo2012/087889中描述了dna甲基化抑制剂的实例,其中每一个通过引用明确地并入本文。在一个实施方式中,dna甲基化抑制剂包括5-氮杂胞苷。也可以提供dna甲基化抑制剂的衍生物和/或药学上可接受的盐。另外,dna甲基化抑制剂(例如,dna甲基转移酶(dnmt)抑制剂)的其他实例包括:dna类似物5-氮杂胞苷、泽布拉林、地西台宾(decitabine);dnmt抑制剂:咖啡酸普鲁姆(caffeicacidpurum)、绿原酸、(-)-表没食子儿茶素没食子酸酯(egcg)、肼苯哒嗪、普鲁卡因酰胺、普鲁卡因、匹萨玛林(psammaplin)a、rg108、非瑟酮、洛美谷里、sgi1027,5-碘杀结核菌素(iodotubercidin)、6-硫鸟嘌呤、mg98、dc-05、和dc-517。下面的表1列出了分子列表,它们在本文描述中另外使用的缩写,以及当用于本文所述的体外实验时每种试剂的最终浓度的实例。表1.研究中使用的生长因子和小分子概述试剂名称缩写终浓度egfe50ng/mlnogginn100ng/mlr-spondin1r500ng/mlchir99021chir,c5μm丙戊酸钠盐vpa,v1.25mmdaptd5μm杜巴他丁atu10μm反苯环丙胺tranyl,ty2μmisx-9isx,i10μmwnt-c59c59,c5μmpd0325901pd1μm616452/repsox65μm5-氮杂胞苷5-aza,51μm根据一个实施方式,使用多种小分子进行的处理可以分几个阶段进行,例如第一和第二阶段,甚至更多阶段。用于处理的阶段数量可根据要进行的分化和/或胰岛素表达增加方法、以及根据所用的小分子类型和所要激活和/或抑制的途径来选择。例如,在进行处理从而将出生后细胞例如出生后干细胞分化为eec的方法中,可以进行第一阶段,使出生后细胞群接触上调ngn3的一种或多种第一小分子,并且在第二阶段,出生后细胞群可以接触下调ngn3的一种或多种第二小分子,从而降低细胞中的ngn3表达。该实施方式的第一和第二阶段可促进出生后细胞例如出生后干细胞分化形成eec。在增加胰岛素的方法的实施方式中,可以进行第一至第三阶段从而处理细胞以上调胰岛素增加表达。根据一个方面,使出生后细胞例如出生后干细胞分化为eec的处理可包括第一和第二阶段,其中第一阶段包括使所述细胞接触包括notch抑制剂和wnt激活剂在内的第一小分子,并且第二阶段包括使用所述细胞接触包括egfr抑制剂和mek/erk抑制剂中至少一个以及notch抑制剂在内的第二小分子。例如,第一阶段可以提供r-spondin1和dapt,而第二阶段可以提供dapt和pd0325901。第二阶段也可以任选地包括wnt抑制剂,例如wnt-c59。第一阶段和第二阶段还可以包含一种或多种生长因子,例如egf和/或noggin。根据又一个其他方面,第一阶段还包括使所述细胞接触hdac抑制剂、组蛋白甲基化抑制剂和neurod1激活剂,例如至少一种杜巴他丁a、反苯环丙胺和isx-9中的至少一种。第二阶段可还包括使所述细胞接触tgf-β抑制剂例如616452,和/或使所述细胞接触组蛋白甲基化抑制剂,例如反苯环丙胺。在一个实施方式中,在细胞群中增加胰岛素表达的处理可以包括第一、第二和第三阶段,其中第一阶段包括使所述细胞接触包括notch抑制剂、wnt激活剂和dna甲基化抑制剂在内的第一小分子,第二阶段包括使所述细胞接触包括notch抑制剂、任选wnt抑制剂、tgf-β抑制剂和neurod1激活剂在内的第二小分子,并且第三阶段包括使所述细胞接触包括notch抑制剂、可选的wnt抑制剂、mek/erk抑制剂和tgf-β抑制剂在内的第三小分子。例如,第一阶段可以提供r-spondin1、dapt、以及5-氮杂胞苷和5-氮杂-2-脱氧胞苷中的至少一种,而第二阶段可以提供dapt、wnt-c59、616452和isx-9,并且第三阶段可以提供dapt、wnt-c59、6116452和pd0325901。第一、第二和/或第三阶段可以还包含一种或多种生长因子,例如egf和/或noggin。根据又一个其他方面,第一阶段还包括使所述细胞接触hdac抑制剂、组蛋白甲基化抑制剂和neurod1激活剂中的至少一种,例如杜巴他丁a、反苯环丙胺和isx-9中的至少一种。第二阶段可以还包括使所述细胞接触组蛋白甲基化抑制剂,例如反苯环丙胺。如上所述,小分子可以以不同的组合提供,并且在不同的治疗阶段提供,以使出生后细胞例如出生后干细胞分化为eec和/或增加胰岛素表达。用于处理的小分子组合的一些实施方式在下文中有所描述。根据一个方面,分化方法可以包括:用多种小分子处理哺乳动物出生后细胞,例如出生后干细胞,所述多种小分子包括notch抑制剂(例如,dapt)、mek/erk抑制剂(例如,pd0325901)和一种或多种生长因子(例如,egf和noggin)。处理可以以单阶段进行。根据另一个方面,所述分化方法可以包括:第一阶段,其采用第一组小分子,包括:notch抑制剂(例如,dapt)、wnt激活剂(例如,r-spondin1)和一种或多种生长因子(例如,egf和noggin);和第二阶段,其采用notch抑制剂(例如,dapt)、mek/erk抑制剂(例如,pd0325901)和一种或多种生长因子(例如,egf和noggin)。根据另一个方面,所述分化方法可以包括:第一阶段,其采用第一组小分子,包括:notch抑制剂(例如,dapt)、wnt激活剂(例如,r-spondin1)、和一种或多种生长因子(例如,egf和noggin)、hdac抑制剂(例如,杜巴他丁a)和组蛋白甲基化抑制剂(例如,反苯环丙胺);和第二阶段,其采用notch抑制剂(例如,dapt)、mek/erk抑制剂(例如,pd0325901)和组蛋白甲基化抑制剂(例如,反苯环丙胺)。根据另一个方面,所述分化方法可以包括:第一阶段,其采用第一组小分子,包括notch抑制剂(例如,dapt)、wnt激活剂(例如,r-spondin1)、和一种或多种生长因子(例如,egf和noggin)、hdac抑制剂(例如,杜巴他丁a)和组蛋白甲基化抑制剂(例如,反苯环丙胺);和第二阶段,其采用notch抑制剂(例如,dapt)、mek/erk抑制剂(例如,pd0325901)、组蛋白甲基化抑制剂(例如,反苯环丙胺)和wnt抑制剂(例如,wnt-c59)。根据另一个方面,所述分化方法可以包括:第一阶段,其采用第一组小分子,包括notch抑制剂(例如,dapt)、wnt激活剂(例如,r-spondin1)、和一种或多种生长因子(例如,egf和noggin)、hdac抑制剂(例如,杜巴他丁a)、组蛋白甲基化抑制剂(例如,反苯环丙胺)、和neurod1激活剂(例如,isx9);和第二阶段,其采用notch抑制剂(例如,dapt)、mek/erk抑制剂(例如,pd0325901)、组蛋白甲基化抑制剂(例如,反苯环丙胺)、wnt抑制剂(例如,wnt-c59)、和tgf-β抑制剂(例如,616452)。在一个实施方式中,增加细胞群中胰岛素的方法可包括:使所述细胞接触小分子,包括dna甲基化抑制剂(例如,5-氮杂胞苷和/或5-aza2-脱氧胞苷)、tgf-β抑制剂(例如,616452)和neurod1激活剂(例如,isx-9)。细胞群可以在单个阶段与分子接触,或者可以在多个阶段与一种或多种分子接触。在又一个方面中,增加细胞群中胰岛素的方法包括三阶段过程,包括:在第一阶段,使所述细胞接触notch抑制剂(例如,dapt)、wnt激活剂(例如,r-spondin1)和dna甲基化抑制剂(例如,5-氮杂胞苷和/或5-aza2-脱氧胞苷),在第二阶段,使所述细胞接触notch抑制剂(例如,dapt)、wnt抑制剂(例如,wnt-c59)、tgf-β抑制剂(例如,616452)和neurod1激活剂(例如,isx-9),并且在第三阶段,使所述细胞接触notch抑制剂(例如,dapt)、wnt抑制剂(例如,wnt-c59)、tgf-β抑制剂(例如,616452)和mek/erk抑制剂(例如,pd0325901)。生长因子例如egf和noggin也可以在第一阶段中提供,并且组蛋白甲基化抑制剂(例如,反苯环丙胺)可以在第一和第二阶段中提供。hdac抑制剂(例如,杜巴他丁a)和/或neurod1激活剂(例如,isx-9)也可以在第一阶段中提供。通过对出生后细胞例如出生后干细胞进行处理从而使细胞分化为肠内分泌细胞,可以获得eec含量相对高的细胞群。根据一个方面,可通过用多种小分子处理而提供具有源自出生后细胞例如出生后干细胞的肠内分泌细胞的细胞群,其中肠内分泌细胞的分数为总细胞群的至少约1%。在另一方面,细胞群中肠内分泌细胞的分数可以为总细胞群的至少约10%,例如20%至100%。细胞群的细胞也可以冷冻保存以用于体外或体内应用。细胞群是未分选的,意味着细胞群未经过过滤或其它分选以实现细胞密度。根据一个方面,分化的细胞可以能够表达5-ht和/或glp-1,如本文包括的实施例中所示。此外,本文所述的增加胰岛素产生的方法可以导致细胞群具有相对高分数的胰岛素产生细胞,例如胰岛素产生细胞的分数为总细胞群的至少约0.05%,甚至总细胞群的至少的至少约1%。本文所述的处理细胞的方法可以通过使用试剂盒进行,所述试剂盒提供:细胞培养基例如基质胶,以及靶向用于处理方法的细胞群,和多种小分子。该试剂盒可以还包括使用该试剂盒的说明书,和有助于进行处理方法的其它器具。在一个实施方式中,试剂盒包括细胞培养基、哺乳动物出生后细胞例如出生后干细胞,和上调chga和分化出生后细胞的多种小分子。在其他实施方式中,试剂盒包括细胞培养基、哺乳动物细胞和在细胞中增加胰岛素产生的多种小分子。本公开的方面还涉及使用本文所述的处理方法治疗哺乳动物(例如人)中的疾病状态。根据一个方面,以内分泌或肠内分泌细胞产物不足为特征的疾病状态得到治疗,例如肥胖症、糖尿病、肠易激综合征、感染性肠炎和炎性肠病中的至少一种。在其他方面,治疗以胰岛素生产不足为特征的疾病状态,例如是肥胖症和i型糖尿病中的至少一种。疾病状态的治疗通过施用多种小分子至需要这种治疗的哺乳动物来实现,所述多种小分子对应于上述任何小分子和/或小分子组合。在实施方式中,多种小分子中所有的小分子被同时(例如,在单个施用步骤中)施用(例如,递送)至需要这种治疗的哺乳动物(例如,人)。在其他实施方式中,多种小分子的小分子在多个施用(例如,作为多步过程的一部分)中被施用(例如,递送)至需要这种治疗的哺乳动物(例如,人)。在一些实施方式中,多个分子体内递送至受试对象的上皮细胞,从而原位诱导细胞分化。在特定实施方式中,所述多个分子被递送至多个组织(例如,胃,小肠,结肠,口腔粘膜)。根据又一个其他方面,通过提供多个小分子以及药物上可接受的赋形剂,可以制备包含多种小分子的药物组合物。在一个实施方式中,该小分子可以制备成药物组合物的一部分,所述药物组合物能够提供保护性制剂,使得小分子在施用(例如,口服)时免于分解直至它们到达目标区域。例如,制剂可以提供保护性涂层,使得多种小分子可以通过胃而达到小肠,并在此发生治疗。这样的制剂可以包括聚合物材料,例如泊洛沙姆,并且还可以包括ph敏感的聚合物,例如eudragit,以提供小分子在身体目标区域的释放。制剂也可以是这样的,即控制一种或多种小分子的释放,以例如控制小分子的释放的开始,或者控制小分子在目标区域释放的持续时间。此外,制剂可以包括可以涂覆胃或肠的其他化合物,例如硫糖铝。虽然药物组合物可以口服施用,例如通过丸剂、胶囊锭(gelcap)、液体等,其他施用方法可以包括通过装置(例如,支架)以及通过栓剂,灌肠剂和/或通过贴剂。在其它实施方式中,通过本文描述的方法获得的细胞群可以通过细胞疗法施用,例如通过输注、注射、在植入物上、在载体材料内或载体材料上,和可以与可以免疫分离的材料或装置组合(例如,同种异体移植)。在一个实施方式中,用于治疗以胰岛素产生不足为特征的疾病状态的药物组合物可包括:dna甲基化抑制剂、tgf-β抑制剂和neurod1激活剂,和/或其衍生物和/或药学上可接受的盐,并组合有药学上可接受的赋形剂。例如,该组合物可包括5-氮杂胞苷和5-氮杂-2'脱氧胞苷中的至少一种、616452和isx-9,以及/或者其衍生物和/或药学上可接受的盐,并组合有药学上可接受的赋形剂。进一步设计组合物制剂,从而使其提供治疗有效量的小分子组合,所述小分子组合有效治疗要治疗的哺乳动物的疾病状态。在另一方面,本发明涉及一种制备用于移植到有需要的受试对象(例如人)中的细胞群的方法。在该方面的一个实施方式中,该方法包括:a)(例如,从受试对象的组织)分离包含lgr5+细胞的细胞群;b)用靶向选自以下的一种或多种过程的一种或多种分子处理细胞:wnt激活、notch抑制、tgfβ抑制和表观遗传调节(例如,lsd1抑制和/或hdac抑制),或其任何组合,和c)用靶向选自以下的一种或多种过程的一种或多种分子处理细胞:wnt抑制、egfr抑制、mek抑制、erk抑制、表观遗传调节(例如、lsd1抑制)、单胺氧化酶(mao)抑制、和notch抑制,或其任何组合。对于步骤b)和c)中的每一步,可以用所述分子对细胞处理各种时间段,例如,大于或等于至约4、8、24、48、96或192小时。通常,步骤b)和c)中使用的分子以大于或等于其ic50值的约10、25、50、100、500或1000%施用。当在步骤b)和c)中使用多个分子处理细胞时,所述分子可以同时施用(例如,在1步中)或分开施用(例如,连续施用)。在另一实施方式中,制备用于移植的细胞群的方法包括:根据上述段落的步骤b)和/或c)处理祖细胞群(例如,过渡扩增细胞(transitamplifyingcell))。根据该实施方式,该方法可包括步骤b)和c),或者其可包括步骤c)而无步骤b)。当分子施用给体内群时,该方法可包括步骤b)和c),包括或步骤b)没有步骤c),或包括步骤c)而没有步骤b)。在另外的实施方式中,制备用于移植的细胞群的方法还包括:d)递送经处理的细胞群至受试对象。在一些实施方式中,本文描述的细胞群用于筛选化合物,所述化合物用于多种目的(例如,用于增强eec及其子群的效率,用于体内控制eec及其子群的大小和功能,用于控制肠促胰岛素表达,例如glp-1和/或glp-2表达、胰岛素表达、和血清素表达,用于毒性试验)的化合物。实施例实施例1小鼠lgr5-egfp-ires-creert2、b6.129小鼠和胰岛素-gfp(ins-gfp,b6.cg-tg(ins1-egfp)1hara/j)小鼠获自jacksonlabs,并且6-至12周龄小鼠用于隐窝细胞分离。动物实验方案由麻省理工学院动物护理委员会(cac)批准。实施例2隐窝分离如前所述(yin等,nature方法,2014),分离隐窝。收集小肠的近端半部(proximalhalf),纵向打开并用冷pbs洗涤以除去腔内容物。然后用剪刀将组织切成2mm至4mm的块,通过10ml移液管上、下吸取还用冷pbs洗涤5-10倍。将组织片段与在pbs中的2mmedta在冰上温育30分钟。去除edta后,用pbs洗涤组织片段以释放隐窝。然后收集隐窝、洗涤并用于细胞培养。将释放的隐窝收集并通过70μm细胞滤网。将分离的隐窝包埋在matrigel中并在24孔板的孔中心铺板。实施例3细胞培养如前所述培养分离的隐窝或单细胞。将200-300个隐窝与40μlmatrigel混合,并在24孔板的孔中心铺板。在matrigel聚合后,提供含有生长因子(egf,noggin,r-spondin1)和小分子(chir99021和vpa)的500μl隐窝培养基(高级dmem/f12,具有n2、b27、glutamax、hepes和n-乙酰半胱氨酸)。对于细胞分化实验,细胞培养基按照如下所述的分化方案更换为含有表2所列生长因子和小分子的培养基。根据所使用的分化条件,每1-2天更换一次培养基。表2:研究中使用的小分子和生长因子实施例4免疫染色通过轻轻吸移到1.5ml蛋白质lobindeppendorf管中,而收集分化的细胞集落。除去细胞培养基,用pbs洗涤样品。通过直接加入4%pfa并在室温下温育10-30分钟来固定在基质胶中培养的类器官或细胞集落。然后将matrigel机械破碎,将细胞转移到bsa包被的eppendorf管中。用pbs洗涤样品,用0.25%tritonx-100透化30分钟,并用一抗和二抗染色。使用的抗体列于表3中。通过共聚焦显微镜(zeisslsm710)或通过倒置显微镜(evos;advancedmicroscopygroup)获得图像。表3:研究中使用的抗体实施例5rna提取和qpcr收获类器官或分化的细胞,并使用rneasymicrokit(qiagen)根据制造商的说明提取rna。使用quantitectprobepcr试剂盒(qiagen),使用市售引物和taqman探针(lifetechnologies)进行定量实时pcr。实施例6glp-1分泌/释放检测将细胞收集在1.5mleppendorf管中,并用基础检测培养基(hbss,补充有10mmhepes,0.1%不含脂肪酸的bsa,并且无葡萄糖,ph7.4)洗涤。将细胞在基础培养基中在300rpm的热混合器中温育2小时。然后洗涤细胞并在含有1mg/ml抑二肽素a(sigma-aldrich)的50μl基础培养基中温育1小时,并收集上清液。然后将细胞在50μl含有1mg/ml抑二肽素a和10mm葡萄糖的基础培养基中温育1小时,并收集上清液。将来自24孔板的类器官收集在1.5ml蛋白质lobindeppendorf管(每管1个孔)中并在基础培养基(hanks'平衡盐溶液(lifetechnologies),补充有10mmol/lhepes,0.1%无脂肪酸bsa,并且无葡萄糖,ph7.4)中,在热混合器中以300转/分钟温育2小时。然后洗涤类器官并在含有1mg/ml抑二肽素a(sigma-aldrich)的100μl基础溶液中温育1小时。收集上清液,并将类器官进一步在含有1mg/ml抑二肽素a和10mm葡萄糖的100μl基础溶液中温育1小时。然后将细胞和类器官在细胞裂解m缓冲液(sigma-aldrich)中裂解。通过elisa(多物种glp-1总elisa,millipore)测定上清液中的glp-1浓度。使用picogreenkit定量细胞裂解物中的dna含量并将其用于标准化glp-1含量。实施例7胰岛素释放检测将细胞收集在1.5mleppendorf管中,并用补充有0.25%bsa并且不含葡萄糖的krebs-ringer缓冲液(krb)洗涤。然后将细胞与具有2mm或20mm葡萄糖的krb在37℃下温育1小时。收集上清液并使用htrf胰岛素检测(cisbio)测量胰岛素。实施例8notch抑制剂增加从isc至肠内分泌细胞(eec)的分化(图1)肠干细胞可在3dmatrigel中培养至形成类器官,其中干细胞自发分化并产生所有的肠上皮细胞类型,包括eec。用小分子γ-分泌酶抑制剂(dapt)进行的notch抑制增加eec分化,约5%-10%的干细胞转变为eec,如图1所示,其中在类器官培养条件(enr)或在存在notch抑制剂dapt(enrd)条件下对eec标志物chga进行免疫染色。dapi用于细胞核染色。实施例9eec分化的筛选系统(图2)为了进一步提高分化效率,测试了干细胞扩增和分化系统中使用的生长因子和wnt/notch调节剂的多种组合,以鉴定具有最高分化效率的基础组合。egf、noggin和dapt(end)的组合,但没有r-spondin1或其他wnt激活剂/抑制剂,诱导高水平的chga表达,如图2所示,其中测量了在多种条件下培养的肠干细胞的chga的mrna表达,所述多种条件包括wnt激活剂chir99021(c)、vpa和wnt抑制剂iwp-2(i)或notch抑制剂dapt(d)的分化组合,以及生长因子egf(e)、noggin(n)、r-spoindin1(r)。总之,采用end条件作为小分子筛选的基础条件。实施例10增加eec分化的小分子的96孔筛选平台(图3)还使用96孔筛选系统,通过qpcr就chga表达作为eec分化的指示来筛选小分子文库,如图3所示,其中lgr5肠干细胞在96孔板中扩增并传代。然后用egf、noggin和dapt(end)以及小分子培养细胞。分化2-3天后,裂解细胞并通过qrt-pcr测量chga表达。实施例11阳性筛选命中(图4)记录示例性筛选结果,如图4所示。egfr抑制剂吉非替尼和mek/erk途径抑制剂as703026增加chga表达。实施例12阳性命中的验证(图5)小分子还使用24孔板培养和分化系统进行验证。egfr抑制剂(例如,吉非替尼)或mek/erk抑制剂(例如,as703026和pd0325901)在促进chga表达方面具有相似的活性,并且goblet细胞标志物(muc2)和潘氏细胞标志物(lyz1)的表达没有类似的增加,表明在eec方向上的特异性诱导,如图5所示,其中肠干细胞在end条件下通过加入小分子而分化。测定分泌谱系细胞(包括杯状细胞(muc2)、肠内分泌细胞(chga)和潘氏细胞(lyz1))的标志物的表达。通过针对chga的免疫染色也证实了eec的产生(图35)。在该条件中,使用notch和egfr/mek/erk抑制和wnt失活(通过r-spondin1撤出)的组合来诱导isc向eec方向的特化。从组合中进一步去除egf和/或noggin诱导了更高水平的chga表达(图36),但在这些条件中观察到更高的细胞凋亡水平(图37)。添加wnt途径抑制剂iwp2(或i)或gsk3β抑制剂chir99021(或c)没有增加朝向eec的分化(图36)。实施例13mapk/erk或egfr抑制剂特异性增加eec分化(图6)肠道干细胞在多种条件下培养,并且小分子as703026(as)、吉非替尼(ge)和pd0325901(pd)特异性增加了肠内分泌细胞标志物chga的表达。实施例14小分子(例如,as、ge和pd)在eec分化过程中降低ngn3表达,并且r-spondin1促进ngn3表达(图7)在分泌细胞分化期间,显示ngn3对于eec的特化是必需的。先前有人推测ngn3阳性分泌祖细胞分化为eec(genesdev.2002)。在肠干细胞体外分化期间检测ngn3表达水平令人惊讶地揭示:去除r-spondin1诱导了ngn3表达的强降低,而添加吉非替尼(ge)、as703026(as)或pd0325901(pd)降低ngn3表达,这在r-spondin1存在下不显著,如图7所示,其中肠干细胞在多种条件下培养2天,并测量ngn3的mrna水平。实施例15优化的分化方案(图8)虽然wnt信号传导(在r-spondin1的条件下)激励eec的初始特化从而促进ngn3表达,但ngn3表达随后在eec成熟期间被下调。这与在eec特化期间的ngn3的短暂表达(transitexpression)一致。因此,测试了2步分化方案,目的是增加ngn3表达和随后的eec分化。添加以r-spondin1和notch抑制剂dapt进行的初始(第一)始发步骤(primestep),以允许ngn3的上调,接着是没有wnt激活但使用小分子(例如,pd)的第二步,以允许eec的末端分化,如图8所示,其中在分化方案中加入存在r-spondin1的初始步骤,从而在第1步中增加ngn3表达。实施例16在eec分化过程中第24小时的ngn3表达(图9)在多种条件下24小时后测量ngn3表达。在存在r-spondin1和notch抑制剂dapt的情况下,极大地诱导ngn3表达,如图9所示,其中r-spondin1和notch抑制剂dapt组合增加ngn3表达。实施例17第3天分化过程中eec关键标志物的表达(图10)切换培养基至没有wnt激动剂或r-spondin1但有pd的条件下,进行另外的2天,显示:与具有r-spondin1或没有pd的条件相比,ngn3表达水平降低并且chga表达被上调,如图10所示,其中2步分化方案在第3天减少ngn3表达并增加chga表达。进一步如图10所示,后期eec标志物neurod1也增加。实施例18在eec分化过程中关键基因的时间进程研究(图11、12和13)eec标志物表达的时间进程研究表明,2步分化方案在第一步中有效增加ngn3水平,在第二步过程中增加chga水平。与ngn3和chga相比,neurod1显示出中等变化趋势,表明neurod1在ngn3之后但在chga之前表达。如图11所示,2步方案在处理5天后诱导出最高的chga表达,并且有效地诱导了从isc至eec的分化,如图12(5天分化后,2步方案增加eec标志物chga表达)和图13(在5天分化后,通过染色eec标志物chga发现,2步方案增加eec分化)所示,其比直接wnt、notch和mek/erk失活(d.pd或d.pd.c59条件)具有更少的细胞凋亡(图38)。实施例19以另外的小分子对分化方案的进一步改进(图14)为了在促进eec分化的第一步过程中增加ngn3水平,在enrd条件下通过另外的筛选鉴定了增加ngn3表达的因子。筛选出超过80个小分子和生长因子,发现杜巴他丁a和反苯环丙胺增加ngn3表达,如图14所示,其中在2天分化后,反苯环丙胺(tranyl)增加ngn3和chga水平,而杜巴他丁a增加ngn3水平,但减少chga水平。然而,虽然反苯环丙胺也增加chga表达,但注意到杜巴他丁a降低了chga表达(也在图14中显示)。实施例20杜巴他丁a(tu)在分化方案的第1步中加入时增加eec分化(图15)杜巴他丁a在第1步中应用,但反苯环丙胺在分化期间在两个步骤中应用。在这些条件下,两种化合物在2步分析方案的5天后增加eec标志物chga表达,并且k细胞(gip)、l细胞(gcg)、和i细胞(cck)的标志物的表达也增加,如图15所示,其中是包括杯状细胞(muc2)、肠内分泌细胞(chga)、潘氏细胞(lyz1)和k-细胞(gip)的细胞的标记物的mrna表达;并且如图16所示,其中是包括杯状细胞(muc2)、肠内分泌细胞(chga)、潘氏细胞(lyz1)、k-细胞(gip)、l-细胞(gcg)和i-细胞(cck)的多个细胞类型的标志物的mrna;并且如图39a-39c所示。值得注意的是,加入反苯环丙胺也显著降低了杯状细胞(muc2)和潘氏细胞(lyz1)标志物的表达,表明eec分化的特异性诱导,也如图16所示。实施例21去除egf还增加eec的分化(图17)此外,由于pd/as靶向mek/erk途径(其是egf途径的下游靶标之一),因此在分化时测试egf的作用。从培养基中去除egf还增加eec标志物(即,chga,gip、gcg和cck)表达,如图17和图39d所示,其中肠干细胞在多种条件下分化5天,并通过qpcr测量肠内分泌细胞的标志物如chga、gip、gcg和cck,但是当在两个步骤中去除egf时,在分化的细胞集落的腔中倾向于存在更多死细胞,这可能是由于egf在促进细胞存活中的作用(例如,pi3k)。因此,在分化过程的第一步中保持egf。noggin在第2步中的作用是微不足道的,因此将它从组合物中除去。实施例22改善的分化方案(图18)为了表征从肠干细胞产生的eec,使用图18中所鉴定和所示的条件进行细胞分化。实施例23从isc至eec的高效分化(图19)针对eec的标志物进行免疫荧光染色。在肠类器官培养(enr条件)中,只有极少数细胞分化为eec,这由类器官中的chga+或glp-1+细胞的数量表示,如图19的上图所示,其中isc在enrcv条件下扩增并在如所示条件下进一步分化。也进行针对肠内分泌细胞标志物(例如,chga)或eec的多种亚型的免疫染色。dapi用于细胞核染色。这些发现与先前的报道(diabetes,2014)一致。在受控的分化条件下,大部分细胞在5天的分化方案后转变为eec,并且eec亚型的产生也升高(参见图19)。有趣的是,大多数eec似乎是血清素分泌细胞,通过强烈的5-ht染色表明(参见图19)。还测试了来自分化的l细胞的glp-1释放。其他eec亚型也以更高的频率存在,如通过标志物例如glp-1、gip和sst的阳性染色所鉴定的。分化方案显著增加类器官的glp-1分泌,并且10mm葡萄糖刺激诱导glp-1分泌的5倍增加,如图20(由isc产生的功能性l细胞(glp-1))所示,其中,在所示条件下培养的细胞中测量glp-1的分泌,并且表明这些eec是成熟的和功能性的(图40c)。在eec分化中,在诱导的第一步中需要wnt信号传导活性,而在eec特化的第二步中不需要。在本文所述的2步分化方案中,虽然具有wnt激活的第1步增加ngn3表达,但它也诱导了潘氏细胞分化,其还可以在第2步中分泌wnt配体和激活wnt途径。这与和end.pd条件相比潘氏细胞标志物lyz1表达的更高水平一致,end.pd条件中最初的wnt失活步骤首先阻止了潘氏细胞的产生(图39e)。因此,测试了在第2步中用小分子抑制剂(wnt-c59或c59)抑制wnt信号传导。c59有效地降低了潘氏细胞标志物lyz1表达,表明它在第2步中阻止了潘氏细胞分化。并且它还增加了chga表达,表明eec分化的水平更高(图40a)。在第2步中延迟添加c5912-24小时还增加chga表达的可能性,这是由于第1步中ngn3诱导的水平更高(图40a)。类似地,添加tgf-β抑制剂(repsox,rep)极大地促进了chga表达并降低杯状细胞标志物muc2表达(图40a,图41a)。rep也大大增加了成熟eec标志物(gcg、gip、cck、tph1、sst、sct、pyy)的表达,相对于enr条件的表达水平为120x(tph1),300x(sct),430x(cck),800x(gip),高达15,000x(pyy)。据发现,在第1步加入repsox足以诱导与在两个步骤中加入它相当的chga水平,但成熟eec标志物的表达大大减少(图41b),表明tgf-β抑制诱导eec特化和进一步成熟。有趣的是,repsox的添加诱导集落从腔中排出凋亡细胞(图41c),并形成高纯度eec细胞集落。使用优化的分化(图40b),测试分化过程中关键分化基因的表达变化(图40c)。干细胞基因lgr5在第2天略微上调,这可能是由于潘氏细胞分化或notch下调的增加(tian等,2015)。sox9的表达牵涉在isc调节和eec分化中(formeister等人,2009),并在胰腺内分泌细胞分化中起重要作用,部分地由ngn3的表达的诱导进行。据发现sox9最初上调,然后从第2天开始下调,其显示与lgr5类似的表达模式。与lgr5相比,sox9表达维持在相对较高的水平,与其在eec中的表达一致(formeister等,2009)。作为激活的notch信号传导的下游基因,hes1的表达被下调。多个notch靶基因的表达类似地调节,包括math1/atoh1、ngn3、和neurod1,这包括初始的上调阶段和之后的下调阶段。并且chga的表达不断增加,与逐渐增加的eec分化一致。有趣的是,这些基因达到峰值水平秒(sox9、math1、ngn3、neurod1为第2天,chga为第5天)的上调速率不同,顺序为sox9、math1、ngn3、neurod1和chga(图40d)。这可能反映了这些基因的天然基因调控过程。进行免疫荧光染色以鉴定在我们的培养物中产生的不同eec亚型。发现多个eec亚型的高频率存在于集落中,通过gip、glp1、血清素、cck、sct和sst的阳性染色进行鉴定(图40e)。有趣的是,在分化的eec集落中发现chga阳性细胞2个群,其中chga高表达群与表达血清素的细胞共定位,而chga低表达群与glp-1、gip、cck、sst和sct表达细胞共定位(图40e)。实施例24增加eec分化的其他因子(图21)鉴定了能够进一步增加eec分化的其他分子,如图21所示,其中包括wnt-c59和isx-9以及它们的组合在内的小分子进一步增加分化细胞的chga表达。对照:以前的分化条件。实施例25多种条件下功能性eec标志物的表达(图22)发现tgf-β抑制剂616452大大增加功能性eec标志物例如gip、gcg、cck、pyy等的表达,而isx9降低其表达,如图22所示,其显示了相对于参照基因hprt的相对表达。因此,调整分化方案以包括616452并在后面阶段去除isx9,如图23所示,其显示了从isc分化至eec的分化方案。实施例26将chga阳性eec细胞转化为胰岛素产生细胞5-aza(5)、616452(6)和wnt-c59(c)的组合在第5天诱导分化肠干细胞中的胰岛素-gfp表达(图24)由于eec和产生胰岛素的β细胞有许多相似之处,最近的数据表明eec可以通过另外的小分子转化为胰岛素产生细胞。使用分离自胰岛素-gfp转基因小鼠的胰岛素-gfp细胞以识别能够诱导胰岛素产生的小分子。在从isc至eec分化期间,当在tgf-β抑制剂616452存在下培养细胞时观察到胰岛素-gfp阳性细胞,约1%的细胞变为gfp阳性。wnt抑制剂wnt-c59和dna甲基化抑制剂5-aza(5-氮杂胞苷)的进一步添加随后增加胰岛素-gfp+细胞的百分比,如图24所示,显示gfp+细胞百分比的facs分析和24孔板的一个孔中gfp+细胞数的定量。值得注意的是,56c(5-aza、616452、wnt-c59)的组合导致最高数量的gfp+细胞(参见图24)。实施例27第5天5-aza在诱导胰岛素-gfp表达中的剂量响应,和第5天616452在诱导胰岛素-gfp表达中的剂量响应(图25)小分子5-aza和616452也在培养物中诱导细胞死亡,因此,确定了5-aza(见图25)和616452(见图26)的最佳浓度。由此,在将来的实验中使用0.5至1μm的5-aza和5μm的616452。实施例28培养5天后isx-9(i)进一步增加胰岛素-gfp表达(图27)还发现在组合中添加isx9(i)进一步增加gfp+细胞的数量(参见图27),并且使用10μm的浓度,如图28中,其中显示第5天isx-9在诱导胰岛素-gfp表达中的剂量响应数据。实施例29培养5天后在多种条件下细胞的胰岛素-gfp表达的facs分析(图29)还在两个转换步骤中测试了wnt信号传导。通过添加chir99021在第1步增加wnt激活没有增加gfp表达,而r-spondin1和wnt-c59的组合导致系统中最高数量的gfp+细胞,如图29所示,其中n:无;chir:chir99201;并且c59:wnt-c59。实施例30在第7天不用药物或用药物处理的细胞的gfp和亮视野(图30)在实施例29的条件下,在分化7天后,高达20%的细胞表达胰岛素-gfp,如图30所示,其中对照:在eec分化条件中培养的细胞。(在第1天enr.d.tu.ty,并且从第2天开始为d.pd.ty.c59)。添加的药物包括5-aza、616452和isx9;在第4天而不是第2天加入pd;并且比例尺:200μm。实施例31过程中的基因表达(图31)细胞打开了对胰岛素表达重要的主要转录因子例如pdx1、ngn3、mafa、neurod1、nkx6.1和nkx2.2的表达,并且大大增加胰岛素mrna表达,如图31所示,其中对照条件为enr.d.tu.ty.5aza.616452.isx9,从第2天开始去除5aza;对于药物处理,首先用enr.d.tu.ty.5aza.616452.isx9培养细胞,并在第2天改变为d.ty.616452.isx9.c59条件,第4天加入pd0325901;并且在第0天,所有细胞均在enr.cv条件下培养。实施例32用低浓度(2mm)和高浓度(20mm)葡萄糖处理细胞诱导不同水平的胰岛素释放(图32)测试了胰岛素从分化的细胞中的释放。用葡萄糖处理细胞诱导剂量依赖性的胰岛素释放,表明这些细胞具有功能性并对葡萄糖响应,如图32所示。实施例33分化方案的流程图(图33)蓝色和绿色的因素代表重要因素。橙色表示支持因素。还列出了分化期间的关键事件,以及每个阶段的关键标志物,如图33所示。实施例34由wnt和notch途径控制的体内eec分化模型(图34)通过反式激活或顺式抑制来调节notch途径的活性。通过notch受体与相邻细胞(即,分泌细胞,例如潘氏细胞、eec和杯状细胞或分泌细胞祖细胞)上的notch配体(dll)的结合,激活notch途径。失去与这些细胞的接触导致notch失活,其进一步通过顺式抑制而增强。wnt途径通过源自固有层的细胞或位于隐窝底部的潘氏细胞的wnt配体梯度而激活。离开隐窝底部导致wnt失活。notch失活还导致wnt信号传导途径的去阻遏,而wnt途径的减弱挽救了与notch阻断相关的表型(tian等,细胞reports,2015)。对于eec分化,当wnt途径是(中度)激活时(即,当wnt在notch失活前失活时,atoh1将被诱导,但不诱导ngn3,导致杯状细胞分化)。nnt3阳性细胞在wnt失活时进一步分化为eec。并且在notch失活时连续(强)的wnt激活导致潘氏细胞的命运。小分子药物例如5-aza、tgf-b抑制剂、isx-9或其他药物可作用于多种细胞类型,包括干细胞、祖细胞、或分化的肠内分泌细胞并诱导这些细胞中的胰岛素表达,如图34所示。实施例35高效肠内分泌细胞分化的临时组合的化学控制在上文描述的研究中,鉴定了在eec的特化中起重要作用的若干信号传导途径。notch途径已经显示决定吸收或分泌分化,并且其失活对于包括杯状细胞、潘氏细胞、簇状细胞(tuftscell)和肠内分泌细胞在内的分泌细胞是必要的。继续沿着分化道路,wnt途径在杯状细胞和潘氏细胞的命运决定中起着重要的作用,其激活诱导潘氏细胞命运并且其失活诱导杯状细胞命运。wnt在eec分化中的作用尚不清楚。如上所述,对于eec特化,wnt激活对于ngn3表达是重要的,并且随后的wnt失活有助于eec的进一步分化和多个eec亚型的成熟(图34)。肠干细胞在体内的分化是动态过程,其中细胞在分化过程中向上移动(潘氏细胞除外)。沿着隐窝-绒毛轴的信号梯度影响分化/迁移过程中使干细胞和祖细胞分化的命运决定。虽然干/祖细胞的特化在隐窝底部以上开始,其中细胞膜结合的wnt配体(由潘氏细胞分泌)通过细胞分裂稀释,当干细胞离开与干细胞壁龛细胞-潘氏细胞的接触时,r-spondin/lgr信号传导模块延长wnt激活(farin等,2016,nature)。该机制允许在notch失活时诱导ngn3表达。因此,wnt/notch失活的顺序和水平似乎在分泌祖细胞的命运决定中发挥作用。并且分泌祖细胞的位置相应地帮助细胞进行选择。例如,在wnt激活的低水平存在的情况下,notch的早期失活可能有利于eec的命运,这归因于ngn3的诱导(例如,在+4位置),wnt信号传导的早期失活以及随后的notch信号传导失活可以促进杯状细胞分化,这是因为ngn3诱导的低水平(例如,高于+4位置),而wnt信号传导的强激活和notch失活有利于潘氏细胞命运(例如,在隐窝中)。因此,来自肠干细胞的eec和其他肠上皮细胞类型的分化是空间和时间上精确控制的过程。虽然wnt和notch途径在包括eec的干细胞分化中起着重要作用,但其他信号传导途径也参与分化过程。如上所述,egfr/mek/erk信号传导级联的抑制增加eec分化水平,并且tgf-β的抑制也极大地促进了eec的特化和成熟。表观遗传调控也参与分化过程,例如hdac和组蛋白去甲基化。这些途径的特定组合控制着isc和祖细胞的命运。实施例36基于将胃肠干细胞转化为胰岛素产生细胞的方案,开发了3阶段过程,具体而言第0-1天为第1阶段,第1-3天为第2阶段,第3-5天为第3阶段。在第3阶段后,细胞开启了胰岛素表达的显著水平,如ins-egfp表达和胰岛素mrna表达的qpcr所示。任选地,细胞可以继续培养1-2周,作为阶段4。对于每个阶段,添加一组小分子(和/或生长因子)的特定组合。目的是通过筛选和测试新的小分子(或生长因子)来优化每个阶段的重编程方案以增加胰岛素表达,具体而言胰岛素mrna表达。repsox(616452)之前在第2-3阶段添加。发现当从第1阶段开始加入(即在第1阶段-3中加入)repsox(616452)时,与第2-3阶段中加入repsox时相比,胰岛素mrna(ins2)增加。当与fgf10(在第1阶段中添加)组合时,胰岛素表达进一步增加,如图42所示。发现当将5aza(5-氮杂-2'-脱氧胞苷)的处理时间从仅为第1阶段延长至第1阶段和第2阶段时,胰岛素mrna表达略微增加(图43)。在第2阶段,当wnt-c59的添加延迟12小时时,胰岛素mrna表达增加。这种延迟的wnt抑制可能允许细胞中ngn3表达的水平更高(因为ngn3表达需要wnt活性),这还增加胰岛素mrna表达(图44)。另一种小分子bayk8644,作为l型ca2+通道激活剂,当在第1阶段中添加时增加胰岛素mrna表达,如图45所示。bayk8644当以2μm添加时显示最高活性(图46)。bayk8644处理当在第1-3阶段中加入时显示最高活性(图47)。dapt的最佳浓度是5μm(图48)。nkx6.1是胰岛素表达和胰岛功能的重要转录因子。测试了另外的因子增加nkx6.1表达以及胰岛素表达的能力。作为组蛋白去甲基酶jmjd抑制剂的iox1,当以5-10μm使用时增加胰岛素和nkx6.1表达(图49)。还测试了repsox和isx9的效果,以鉴定促进胰岛素和nkx6.1表达的最佳浓度。发现在促进胰岛素和nkx6.1表达方面,repsox的最佳剂量是10-15μm,isx9的最佳剂量是10-20μm(图50-51)。对于wnt-c59,最佳剂量是5μm(图52)。另外,发现当在第2-3阶段中添加时,骨形态发生蛋白(bmp)alk2受体抑制剂dmh-1增加nkx6.1以及胰岛素的表达(图53)。此外,地塞米松在第2-3阶段中加入时也增加nkx6.1和胰岛素表达(图54)。当在第2-3阶段中添加时,维生素c具有有益效果。还测试了另外的小分子(生长因子)在第3阶段促进胰岛素表达的能力。当小分子t3(三碘甲状腺氨酸)在第3阶段加入时,它大大增加胰岛素mrna表达。而维生素c(pvc)没有显示出这种效果,如图55所示。类似地,包括n-乙酰半胱氨酸(2mm)在内的另外的分子也增加胰岛素mrna表达,并且当与t3组合时还增加胰岛素表达,如图56所示。还发现通过在第3阶段中用chir99021(chir)转换wnt-c59,胰岛素mrna大大增加,如图57所示。当chir和t3组合时,胰岛素mrna进一步增加。鉴定了包括艾塞那肽-4(ex4)和极光激酶抑制剂ii(aki)的另外的小分子或因子增加胰岛素mrna表达,如图58所示。还发现佛司可林(50-100μm)在第3阶段加入时改善了细胞存活率。此外,使用甲状腺激素β受体激活剂gc-1来替代t3并显示出类似的促进胰岛素mrna表达的效果(图59)。通过使用这些鉴定出的因子的组合,在5天内从肠干细胞诱导胰岛素表达和胰岛素-gfp表达。继续培养细胞进一步增加胰岛素表达水平并产生胰岛样细胞集落,如图60所示。对于继续培养胰岛样的表达胰岛素的细胞集落,发现佛司可林的添加增加细胞存活。包括dapt、pd0325901、bayk8644、dmh-1的因子可以被移除而没有太大影响,如图61所示。eec分化的其他数据。基于如本文所述的观察,开发了eec分化的修改的流程图(图61),如下所示。主要差异:1.从第0天开始,而不是从第1天开始添加repsox(616452)。2.isx-9改为可选,并且未在eec-v6.docx中描述的方案中使用。3.repsox改为重要因素,因为它大大增加了eec分化,如eec-v6.docx所述。4.wnt-c59在从第0天起36h加入。从以上描述中显而易见的是,对于本文所述的公开内容可以进行变化和修改,以使其适用于各种使用和条件。本文描述了用于本公开使用的方法和材料;本领域已知的其他合适的方法和材料也可以使用。所述材料、方法和实施例仅是说明性的而不是限制性的。这些实施方式也在所附权利要求的范围内。在此处变量的任何定义中对元素列表的叙述包括作为所列元素中任何单个元素或组合(或子组合)的该变量的定义。本文的实施方式的叙述包括作为任何单个实施方式或与任何其他实施方式或其部分组合的该实施方式。本文引用的所有专利、公开申请和参考文献的教导通过引用整体并入本文。虽然本发明已特别示出并参考其示例性实施方式进行描述,本领域技术人员将理解,在不脱离所附权利要求所涵盖的本发明的范围的情况下,可以在其中进行形式和细节的各种改变。参考文献和等同物的方式加入本文引用的所有专利、公开申请和参考文献的教导通过引用整体并入本文。虽然本发明已特别示出并参考其示例性实施方式进行描述,本领域技术人员将理解,在不脱离所附权利要求所涵盖的本发明的范围的情况下,可以在其中进行形式和细节的各种改变。参考文献barker,n.,vanes,j.h.,kuipers,j.,kujala,p.,vandenborn,m.,cozijnsen,m.,haegebarth,a.,korving,j.,begthel,h.,peters,p.j.等(2007).identificationofstemcellsinsmallintestineandcolonbymarkergenelgr5.nature449,1003-1007.farin,h.f.,vanes,j.h.,和clevers,h.(2012).redundantsourcesofwntregulateintestinalstemcellsandpromoteformationofpanethcells.gastroenterology143,1518-1529e1517.formeister,e.j.,sionas,a.l.,lorance,d.k.,barkley,c.l.,lee,g.h.,和magness,s.t.(2009).distinctsox9levelsdifferentiallymarkstem/progenitorpopulationsandenteroendocrinecellsofthesmallintestineepithelium.amjphysiolgastrointestliverphysiol296,g1108-1118.furness,j.b.,rivera,l.r.,cho,h.j.,bravo,d.m.,和callaghan,b.(2013).thegutasasensoryorgan.natrevgastroenterolhepatol10,729-740.gribble,f.m.,和reimann,f.(2015).enteroendocrinecells:chemosensorsintheintestinalepithelium.annurevphysiol.gunawardene,a.r.,corfe,b.m.,和staton,c.a.(2011).classificationandfunctionsofenteroendocrinecellsofthelowergastrointestinaltract.intjexppathol92,219-231.lee,c.s.,perreault,n.,brestelli,j.e.,和kaestner,k.h.(2002).neurogenin3isessentialfortheproperspecificationofgastricenteroendocrinecellsandthemaintenanceofgastricepithelialcellidentity.genesdev16,1488-1497.sato,t.,和clevers,h.(2013).growingself-organizingmini-gutsfromasingleintestinalstemcell:mechanismandapplications.science340,1190-1194.sato,t.,vanes,j.h.,snippert,h.j.,stange,d.e.,vries,r.g.,vandenborn,m.,barker,n.,shroyer,n.f.,vandewetering,m.,和clevers,h.(2011).panethcellsconstitutethenicheforlgr5stemcellsinintestinalcrypts.nature469,415-418.sato,t.,vries,r.g.,snippert,h.j.,vandewetering,m.,barker,n.,stange,d.e.,vanes,j.h.,abo,a.,kujala,p.,peters,p.j.等(2009).singlelgr5stemcellsbuildcrypt-villusstructuresinvitrowithoutamesenchymalniche.nature459,262-265.tian,h.,biehs,b.,chiu,c.,siebel,c.w.,wu,y.,costa,m.,desauvage,f.j.,和klein,o.d.(2015).opposingactivitiesofnotchandwntsignalingregulateintestinalstemcellsandguthomeostasis.cellrep11,33-42.vandussen,k.l.,carulli,a.j.,keeley,t.m.,patel,s.r.,puthoff,b.j.,magness,s.t.,tran,i.t.,maillard,i.,siebel,c.,kolterud,a.等(2012).notchsignalingmodulatesproliferationanddifferentiationofintestinalcryptbasecolumnarstemcells.development139,488-497.yin,x.,farin,h.f.,vanes,j.h.,clevers,h.,langer,r.,和karp,j.m.(2014).niche-independenthigh-purityculturesoflgr5+intestinalstemcellsandtheirprogeny.natmethods11,106-112.mumphrey,m.b.等,roux-en-ygastricbypasssurgeryincreasesnumberbutnotdensityofcck-,glp-1-,5-ht-,andneurotensin-expressingenteroendocrinecellsinrats.neurogastroenterolmotil,2013.25(1):p.e70-9.moran,g.w.等,enteroendocrinecells:neglectedplayersingastrointestinaldisorders?therapadvgastroenterol,2008.1(1):p.51-60.bogunovic,m.等,enteroendocrinecellsexpressfunctionaltoll-likereceptors.amjphysiolgastrointestliverphysiol,2007.292(6):p.g1770-83.worthington,j.j.,theintestinalimmunoendocrineaxis:novelcross-talkbetweenenteroendocrinecellsandtheimmunesystemduringinfectionandinflammatorydisease.biochemsoctrans,2015.43(4):p.727-33.manocha,m.和w.i.khan,serotoninandgidisorders:anupdateonclinicalandexperimentalstudies.clintranslgastroenterol,2012.3:p.e13.sternini,c.,l.anselmi,和e.rozengurt,enteroendocrinecells:asiteof'taste'ingastrointestinalchemosensing.curropinendocrinoldiabetesobes,2008.15(1):p.73-8.vanderflier,l.g.和h.clevers,stemcells,self-renewal,anddifferentiationintheintestinalepithelium.annurevphysiol,2009.71:p.241-60.petersen,n.等,generationoflcellsinmouseandhumansmallintestineorganoids.diabetes,2014.63(2):p.410-20.beucher,a.等,thehomeodomain-containingtranscriptionfactorsarxandpax4controlenteroendocrinesubtypespecificationinmice.plosone,2012.7(5):p.e36449.当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1