用于适配子对选择的方法与流程
本申请要求2016年6月17日提交的美国临时申请序列号62/351,890的优先权的权益,其内容通过引用并入本文。
本发明涉及用于从随机化寡核苷酸文库开始获得对于目标靶点有选择性的成对寡核苷酸适配子的方法、试剂和试剂盒。
背景技术:
适配子是小的人工配体,包括能够以高亲和力结合特定目标靶点部分的单链dna、rna或多肽分子。约二十五年来,适配子一直被认为是抗体的潜在替代物,用于作为诊断和/或治疗目的使用。适配子通常由通过基于亲和力的针对目标靶点的分配筛选候选寡核苷酸的随机文库而获得。适配子在宽的ph和温度范围内具有高的结构稳定性,使其成为用于广谱的体外、离体和体内应用的理想试剂。
在1990年,tuerk等,1990(science249,505-510)和ellington1990(nature346,818-822,doi:10.1038/346818a0)开发了模拟进化过程的体外方法。该过程被称为指数富集的配体系统进化(selex)。selex过程利用进化的基本概念,运用变异、选择和复制以从核酸分子(即寡核苷酸)的起始库获得高靶点亲和力和特异性。通常,为了选择核酸(寡核苷酸dna或rna)适配子,通过合成在从约20至约100个核苷酸的大小范围内的短寡核苷酸文库(约1014个不同序列)来实现变异。每个寡核苷酸包括侧翼为引物区域的内部随机区域,用于随后通过合适的扩增反应扩增,例如用于dna的聚合酶链式反应(pcr),或用于rna的逆转录聚合酶链式反应(rt-pcr)。由于文库中大数量的独特序列,至少一些适配子分子以特异性和亲和力结合靶点的可能性很高。接下来,通过将核酸库与固定在珠上的靶点分子一起温育,然后洗去非结合序列来实现选择。结合的适配子分子被洗脱并且扩增,形成用于下一轮selex的输入。通过使用pcr或一些其他扩增方法扩增结合的寡核苷酸来实现复制。然后将从一轮selex获得的寡核苷酸库(即选择后扩增)用作用于下一轮selex的输入,直到一组寡核苷酸(即核酸适配子)被富集,其紧密并且特异性地结合至靶点分子。最近在随机文库的制备以及在基于亲和力的分配中的技术进步使得适配子具有与抗体等同的亲和力。
该技术已应用于针对无机组分、小有机分子(ellington,等,1990,nature346,818-822,doi:10.1038/346818a0)、核苷酸(sassanfar1993,nature5;364(6437):550-3)、辅因子(lorsch1994,biochemistry,33(4):973-82)、核酸(boiziau,等,1999,jbiolchem.30;274(18):12730-7)、氨基酸(majerfeld等,2005jmolevol.61(2):226-35)、碳水化合物(jeong等,2001,biochembiophysrescommun.16;281(1):237-43)、抗生素(wang1996,biochemistry,35(38):12338-46.)、肽(ylera等,2002,.biochembiophysrescommun290(5):1583-8)、蛋白质(tuerk,1990同上)以及甚至结构复杂,诸如细胞(shangguan等,2006procnatlacadsciusa.103(32):11838-43)来选择dna或rna适配子。
适配子优于抗体的优点有容易体外合成、修饰灵活、靶点范围广泛、可重复使用性和热/化学稳定性高。非免疫原性和解毒剂的可用性增加了适配子作为治疗药物的价值。
通常,生物测定需要一对配体以在检测中实现高灵敏度和特异性。此外,连接对将是增强其亲和力和特异性的新配体。在这方面,迄今为止,抗体胜过适配子。由于它们在生产之前预先设计的抗原结合位点,因此相当容易获得一对抗体。与之相比,由于传统selex方案的基础性限制,难以获得具有不同的靶点结合位点的一组适配子。适配子已经从随机化文库中富集,而没有结合位点的现有知识,因此不可能使用目前可用的方法将结合位点分配到库中的个体适配子。
在常规适配子选择方案中,来自适配子富集库的配对选择被认为是在鉴定个体适配子后的后选择步骤。到目前为止,只有三个成功的用于适配子对选择的平台。所有这些都基于“暴力”策略,以(1)选择单个适配子,阻断其结合位点,然后重新开始并且选择另一个单个适配子(ochsner,等,2014,biotechniques56,125-133,doi:10.2144/000114134和csordas,等,2016,anal.chem88,10842-10847,doi:10.1021/acs.analchem.6b03450)或(2)鉴定库或文库中的个体适配子,然后用构建的适配子矩阵进行同时结合的成对测试(cho,m.等,2015,analyticalchemistry87,821-828,doi:10.1021/ac504076k)。前者的途径耗时,而后者“鸟枪法”途径,为了其成功,必须依赖待测试的适配子矩阵的大小,而这与费用相关。
这种解耦途径导致了在寻找适配子对方面极其高的经济成本和低效率,因此,对选择适配子对的速度和效率的改进仍有长期的需求。因此,本发明提供了用于rna适配子对的靶点驱动的选择的高效率新方法,其中rna适配子同时被选择作为能够选择性结合相同目标靶点的对。
技术实现要素:
本发明提供了用于富集成对寡核苷酸适配子的适配子对选择方法、试剂和试剂盒。根据本发明的适配子对选择方法通过仅允许结合到目标靶点的寡核苷酸对在选择过程期间存活,而直接从随机库中选择针对自由溶液靶点的适配子对。通过本发明,可以同时选择成对的适配子,减轻了许多当前如高费用、低效率和工作流程繁琐的问题。本发明的同质性质对每轮选择提供可复用性、可延长性、稳健性和监控的容易性。
概括地说,本发明提供了分离用于选择性结合目标靶点的寡核苷酸适配子对的方法,该方法包括,
(a)制备两个序列随机化的寡核苷酸的文库,
(b)通过针对目标靶点的基于亲和力的分配独立地筛选(a)中的每个文库,以获得富集有结合目标靶点的寡核苷酸的寡核苷酸的各自的a和b库,
(c)将库a和库b寡核苷酸与目标靶点以及连接头寡核苷酸一起温育,以便形成两个寡核苷酸、连接头和目标靶点的四部分复合物,
(d)向(c)的产物中加入连接酶以连接复合物的寡核苷酸以形成连接的寡核苷酸,
(e)通过聚合酶链式反应(pcr)或通过逆转录聚合酶链式反应(rt-pcr)扩增(d)中的连接的寡核苷酸,以产生编码两个寡核苷酸适配子的dna寡核苷酸。
该方法还包括通过重复步骤(c)至(e)使寡核苷酸对经受一个或多个额外的富集循环,直至所获得的寡核苷酸适配子对的特异性已被优化,其中(a)、(b)、(c)和(d)中的寡核苷酸是dna、rna、表1中列出的具有修饰核苷酸的dna和/或表1中列出的具有修饰核苷酸的rna。
此外,两个文库中的随机化寡核苷酸的大小范围从约60至约200个核苷酸,并且包括内部随机区域,每个随机区域侧翼为引物区域,在相应的a和b寡核苷酸的5'和3'末端上包括独立选择的寡核苷酸标签。
在一个方面,基于亲和力的分配是指数富集的配体系统进化(selex),或selex的任何变体。通常,目标靶点选自由肽、蛋白质、核酸、细胞、活体组织的组分、有机分子和无机分子组成的组。
在更具体的实施方式中,本发明提供了一种分离用于选择性结合目标靶点的寡核苷酸适配子对的方法,从rna寡核苷酸开始,包括:
(a)制备两个大小范围约60至约200个核苷酸的随机化rna寡核苷酸文库,
(b)通过针对目标靶点的基于亲和力的分配独立地筛选(a)中的每个文库,以获得富集有结合目标靶点的rna寡核苷酸的寡核苷酸的各自的a和b库,
(c)将库a和库b的rna寡核苷酸与目标靶点以及连接头寡核苷酸一起温育,以便形成两个寡核苷酸、连接头和目标靶点的四部分复合物,其中保持与两个寡核苷酸接近的连接头寡核苷酸的大小范围为约40至约120个核苷酸,
(d)将连接酶加入到(c)中的温育的复合物中,以形成与靶点结合的来自库a的rna寡核苷酸和来自库b的rna寡核苷酸之间的共价连接,
(e)通过逆转录聚合酶链式反应(rt-pcr)扩增(d)中的连接的rna寡核苷酸,以产生编码两个rna适配子的dna寡核苷酸,
(f)用选择的引物扩增(e)中的dna寡核苷酸,以分离编码库a的rna适配子(适配子a)和来自库b的rna适配子(适配子b)的dna寡核苷酸,
(g)使(f)中的dna寡核苷酸经体外转录以产生rna寡核苷酸适配子对;并且
其中每个相应文库的寡核苷酸包括内部随机区域,每个随机区域的侧翼为引物区域,在相应的a和b寡核苷酸的5'和3'末端上包括独立选择的寡核苷酸标签。
在另一方面,引物的长度为至少15个核苷酸,并且优选长度为约20个核苷酸。
该方法还包括通过逆转录聚合酶链式反应(rt-pcr)扩增连接的rna寡核苷酸,以产生编码两个rna适配子的dna寡核苷酸,
另一方面,体外转录以产生rna寡核苷酸适配子的步骤任选在将合适的启动子5'引入到编码rna适配子的两个双链dna寡核苷酸后进行。该启动子是例如t7启动子。
另一方面,本方法还包括在步骤(c)之后加入衔接子或引物双链体,以便在库中延长固定的寡核苷酸,其中衔接子或引物双链体是包括引物的两个杂交的寡核苷酸。
另一方面,本方法还包括在步骤(f)之后加入,在碱性条件下水解库b的部分。
在更具体的实施方式中,本发明提供了一种分离用于选择性结合目标靶点的寡核苷酸适配子对的方法,该方法包括,
(a)制备两个大小范围为约60至约200个核苷酸的随机化rna寡核苷酸文库,
(b)通过针对目标靶点的基于亲和力的分配独立地筛选(a)中的每个文库,以获得富集有结合目标靶点的rna寡核苷酸的rna寡核苷酸的各自的a和b库,
(c)将库a和库b的rna寡核苷酸与目标靶点以及连接头寡核苷酸一起温育,以便形成两个寡核苷酸、连接头和目标靶点的四部分复合物,其中保持两个寡核苷酸接近的连接头寡核苷酸的大小范围为约40至约120个核苷酸,
(d)加入衔接子或引物双链体,以便在库中延长固定的寡核苷酸,其中衔接子或引物双链体是包含引物的两个杂交的寡核苷酸,
(e)将连接酶加入到(d)中的温育的复合物中,以形成与靶点结合的来自库a的寡核苷酸和来自库b的寡核苷酸之间的共价连接,以及衔接子和来自每个库的寡核苷酸之间的共价连接,
(f)通过逆转录聚合酶链式反应(rt-pcr)扩增(e)中的连接的寡核苷酸,以产生编码两个rna适配子的dna寡核苷酸,
(g)用选择的引物扩增(f)中的dna寡核苷酸以产生编码rna适配子的两个双链dna寡核苷酸,以分离编码库a的rna适配子(适配子a)和来自库b的rna适配子(适配子b)的dna寡核苷酸,以及
(h)在碱性条件下水解库b的部分,
(i)使(g)和(h)中的产物经历体外转录以产生富集rna寡核苷酸适配子的库;并且
其中每个相应文库的寡核苷酸包括内部随机区域,每个随机区域侧翼为至少一个引物区域,包括在相应的a和b寡核苷酸的5'和3'末端上的寡核苷酸标签,以及4-6个固定的核苷酸的寡核苷酸标签。
附图说明
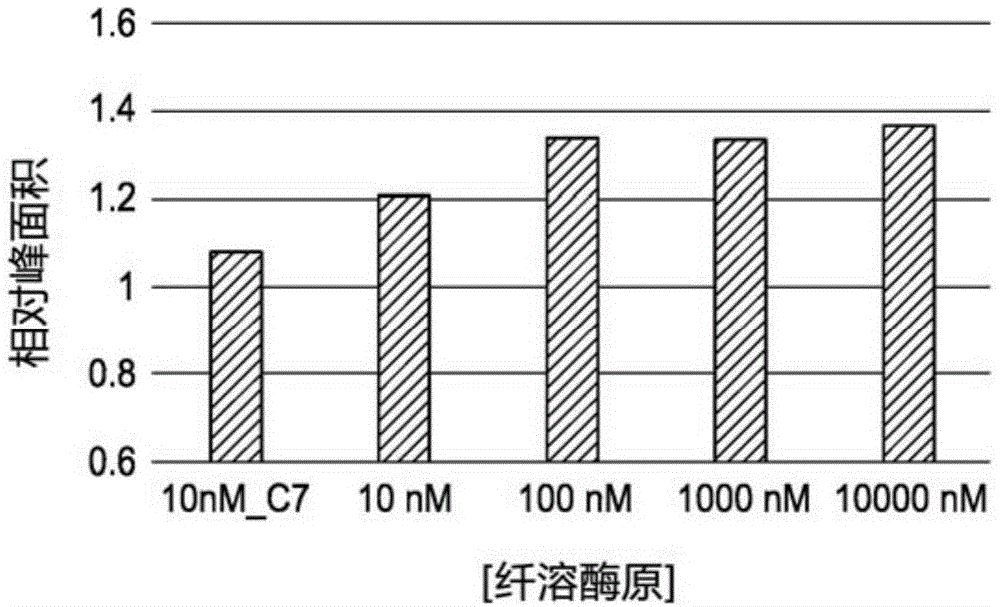
图1说明了用图1中描述的方法的第1轮适配子对的选择的靶点依赖性rna适配子对库富集。图1a说明了血纤维蛋白溶酶原的结果。图1b说明了人补体7的结果。
图2说明了靶点依赖性rna适配子对库的富集。1μl样品中的人血清蛋白质,nm
图3说明了混合的适配子库(库a和b,1:1摩尔比)的解离常数(kd),比较第零和第三轮适配子对库。
图4说明了用适配子对库作为配体的邻近连接测定(pla)的灵敏度,比较第零、第二和第三轮适配子对库,1μl样品中的人血清蛋白质,nm。
具体实施方式
将描述本发明的优选实施方式,但是本发明不限于该实施方式。本领域技术人员也可以进行变化和修改。
概括地说,本发明提供了rna适配子对的靶点驱动的选择,该rna适配子对同时被作为能够选择性结合相同目标靶点的适配子对。靶点可以是任何生物材料、生物分子和/或易于选择性结合适配子的其他组分或材料,包括但不限于肽、蛋白质、dna或rna分子、细胞、活体组织的组分,有机分子和/或无机分子、毒素、病毒、细菌。
通过产生两个随机化的单链rna寡核苷酸文库来起始该过程。
寡核苷酸的大小范围可以为约60至约200个核苷酸,其包括大小范围为20至100个核苷酸的随机化rna。通过基于亲和力的分配预筛选侧翼为在相应的5'和3'末端上的不同的rna序列的两个随机rna寡核苷酸文库。这可以通过应用selex方法来实现,或者可以通过任何其他本领域已知的基于亲和力的方法针对目标靶点预筛选文库。用于筛选适配子的方法描述于例如wo2000056930a1中。
对于本发明,selex仅通过几轮(例如,从1至6轮)进行,从示例的两个随机寡核苷酸文库开始,以便产生富集有能够选择性地结合目标靶点的分子(库a和库b)的寡核苷酸分子库。
从侧翼为两个引物的随机文库选择rna适配子对的示意图(方案1)示出了如何在靶点的存在下募集成对的rna寡核苷酸适配子候选物,然后将其制备成用于下一轮选择的库。在复合物中与靶点和核苷酸连接头协同结合的寡核苷酸将被优先“标记”用于扩增(例如,rt-pcr)。靶点分子募集起源于库a的适配子以及起始于库b的其成对适配子。
将从文库a和b富集的来自库的寡核苷酸与目标靶点一起温育,并且在连接头寡核苷酸的存在下温育。该连接头寡核苷酸与库a寡核苷酸的3’端以及与库b寡核苷酸的5’端互补,使得当与目标靶点混合时来自库a和库b的rna寡核苷酸可保持接近多至120个碱基对。该连接体寡核苷酸是寡核苷酸,其大小范围约10至约50个核苷酸,或更特别地是长度为约18至约22个核苷酸。
从侧翼为两个引物的随机文库选择rna适配子对的示意图(方案1)
子图1说明了用于rna适配子对选择的输入rna的制备;
子图2说明了如果候选rna寡核苷酸是适配子,则通过rt-pcr进行靶点依赖性连结和随后的扩增;以及
子图3说明了通过选择性扩增、体外转录、去磷酸化和适配子折叠的诱导,由扩增的连接的编码两个rna适配子库a和库b的进化。
方案1.从侧翼为两个引物的随机文库选择rna适配子对的示意图
一旦寡核苷酸已显示其会特异性结合目标靶点,术语适配子就应用于本文。源自库a和库b的寡核苷酸形成三分子的相互作用(来自库a的适配子,来自库b的适配子和靶点)以包括靶点-适配子对复合物,其大大增强募集的适配子通过其预先设计的低聚物尾部对与短寡核苷酸连接头的杂交能量。特别地,库a的rna寡核苷酸上的3’标签和库b的rna寡核苷酸上的5’标签是将与连接头寡核苷酸杂交的区域。库a寡核苷酸的5’标签和库b寡核苷酸的3’标签是在通过连接分别连结来自库a和库b的所选寡核苷酸后用于选择性扩增的引物结合的区域。
获得配体对的优点是通过应用靶向靶点部分的不同结合位点(例如表位)的两种不同的配体,可以在测定或临床应用中实现更高水平的灵敏度和选择性。这是协同稳定化或“邻近效应”的结果。由于靶点结合,邻近效应导致连接头附近的适配子对的浓度升高。这种邻近增强了接近短寡核苷酸连接头的两个适配子对的杂交能量。
当与连接头寡核苷酸复合时,产生四分子复合物(与寡核苷酸连接头杂交的靶点-适配子对复合物)。然后使四分子复合物经历连接反应,例如,通过加入连接酶酶,诸如rna或dna连接酶,以在成对的适配子之间形成共价连接,从而“标记”它们,用于扩增。编码一对募集的适配子的连接产物优先被例如使用特异性识别库a和库b寡核苷酸的一对引物通过rt-pcr扩增。
该选择过程导致靶点-适配子对复合物定量转化为成比例的量的连接适配子。
所有以上步骤都在自由溶液中进行,在扩增之前不需要对库进行物理分配(即洗涤)。通过使用随后的酶促反应解固来自连接产物库的两个适配子库,重复多轮(即反复)选择过程。这些酶反应不是改变dna群体的主要的选择压力。
在体外rna或dna选择中结合修饰的核苷酸,提供了许多潜在的优点,诸如所选核酸对核酸酶降解的稳定性增加,亲和力改善,化学功能扩展和文库多样性增加。引入具有新碱基配对的修饰可潜在地提供额外的化学和功能特性,不受未修饰的核苷酸碱基配对的限制。修饰的核苷酸库还可潜在地增加所选适配子的整体的结合亲和力。适配子-靶点结合通常由极性、氢键结合和电荷-电荷相互作用介导。相比之下,有助于蛋白质-蛋白质相互作用的疏水性接触是有限的。因此,加入模拟氨基酸侧链的官能团可以扩展化学多样性并且增强适配子的结合亲和力。
核糖2'-oh的修饰是一种增加rna稳定性的任选途径。小的电负性2'取代基,诸如2'-氟(2'-f),dna(2'-h),2'-o-甲基(2'-ome)是最广泛使用的,因为它们是耐受良好的,通常增强rna核酸酶抗性,而不显著影响rna热稳定性和构象。氟取代(2'-f)稍微稳定dsrna双链体(每个修饰,tm增加~1℃),是最耐受的修饰类型之一。除2'-f外,还已知2'-ome修饰在rna结构中是耐受良好的,并且增加核酸酶抗性。核糖核酸。常用的2'取代基包括核糖核酸、硫代磷酸酯、二硫代磷酸酯;ea、2'-氨乙基、脱氧核糖核酸、2'-氟、2'-o-甲基、2'-o-甲氧乙基、2'-脱氧-2'-氟-β-d-阿拉伯糖核酸、4'-c-羟甲基-dna、锁核酸、2',4'-碳环-lna-锁核酸、氧杂环丁烷-lna、解锁核酸、4'-硫代核糖核酸、2'-脱氧-2'-氟-4'-硫代核糖核酸、2'-o-me-4'-硫代核糖核酸、2'-氟-4'-硫代阿拉伯糖核酸、阿卓糖醇核酸、己糖醇核酸。
预期的寡核苷酸可任选地包括硫代磷酸酯核苷酸间连接修饰、糖修饰、核酸碱基修饰和/或磷酸盐骨架修饰。寡核苷酸可包含天然磷酸二酯骨架或硫代磷酸酯骨架或任何其他修饰的骨架类似物,包括任选的lna(锁核酸)、pna(具有肽骨架的核酸)、cpg寡聚体等,诸如2002年的tides、2002年5月6-8日在内华达州(nv)拉斯维加斯(lasvegas)的寡核苷酸和肽技术会议(oligonucleotideandpeptidetechnologyconferences)、2003年11月18日&19日在德国(germany)汉堡(hamburg)的寡核苷酸&肽技术(oligonucleotide&peptidetechnologies)中公开的那些,其内容通过引用并入本文。
根据本发明的寡核苷酸还可任选地包括任何合适的本领域已知的核苷酸类似物和衍生物,包括由下表1列出的。
对本发明所预期的寡核苷酸的修饰包括,例如,用允许寡核苷酸与所需聚合物共价连接的官能团或部分添加到或取代所选核苷酸,和/或将附加电荷、极化性、氢键结合、静电相互作用和官能性并入到寡核苷酸的官能部分的添加或取代。这些修饰包括但不限于2'-位糖修饰、5-位嘧啶修饰、8-位嘌呤修饰、环外胺的修饰、4-硫代尿苷的取代、5-溴或5-碘尿嘧啶的取代、骨架修饰、甲基化、碱基配对组合,诸如异碱基(isobases)异胞苷和异胍,以及类似的组合。预期在本发明范围内的寡核苷酸还可包括3'和/或51帽结构。参见freier&altmann,1997,nυcl.acidres.;25,4429-4443和uhlmann,2000,curr.opinionindragdevelopment,3(2),293-213中描述的核苷类似物的实例。
不受任何理论束缚,本发明的若干优点包括:
·本发明的rna适配子对选择不需要特征化的适配子库来寻找适配子对。只要具有两个不同的靶点结合位点的适配子可在两个库中获得,本rna适配子对选择则产生一个库中的适配子和另一个库中的成对适配子。这允许适配子对的同质选择,其中适配子的表征被继承。
·通过在存在/不存在靶点时产生的扩增的连接产物的量,即连续的定量评估,在每一轮容易地监测选择过程(参见图1、图2和图4)。
·未纯化的、未测序的适配子库可直接在最终rna适配子对选择轮之后立即用于夹心测定,诸如邻近连接测定(参见图4)。这提供了比较便宜的替代,类似于多克隆抗体对(尽管批次与批次间的差异较大)。
·本rna适配子对选择使得许多后选择步骤不再必要(例如,用二维适配子矩阵的成对组合筛选)。与筛选适配子对的成对匹配仅通过对连接产物测序来完成。选择应在仅几轮rna适配子对选择后完成。该技术基于邻近效应,即在靶点的存在下来自两个库的适配子的熵稳定的协同进化。zhang等人在2013年(angewandtechemie,doi:10.1002/anie.201210022)指出,在靶点的存在下的邻近测定中的dna组装导致两个探针的局部浓度增加~4×105倍。从这个数字开始,如果假设在1012个随机序列中有数百个适配子,那么在仅两轮rna适配子对选择后,适配子对与随机序列的摩尔比率将达到1:1。在另一研究中,liu等人在2014年(journaloftheamericanchemicalsociety,doi:10.1021/ja412934t)中估计,在67,858种可能的组合中,存在五种适配子,其结合亲和力范围从kd=0.2nm至3.2μm。从这个对于丰度的假设开始,找到适配子对所需的rna适配子对选择轮次数量为至多三轮。
在从侧翼为一个引物的随机文库的rna适配子对选择的示意图中(方案2)示出的适配子对选择方案允许在选择期间最小化固定序列的参与,使得所选适配子长度短并且具有更大的修饰灵活性。通过标准selex过程鉴定的适配子通常包括60-120个核苷酸:30-70个核苷酸长的随机化区域加上每侧~15-25个核苷酸的固定引物位点。然而,截短selex在30-40个随机化核苷酸序列的每一侧需要约4-6个固定核苷酸,使得后选择步骤被简化。在wo2000056930a1中描述了在我们的适配子对选择平台中适用的该截短selex技术。我们的rna文库由随机化区域组成,其在文库a的5'端(1b)和文库b的3'端(4b)上侧接六个核苷酸长的固定序列段。实际上,用作连续扩增的引物是不够长的。选择后,它们用作预退火的双链衔接子(1a-1a'-1b'和4a-4a'-4b')中的桥接寡核苷酸的杂交位点。连接后,库a(1a+1b)中的连接物不仅是正向引物位点,而且在其3'端还包含t7启动子。库b(4a+4b)中的连接物是用于pcr的反向引物位点。在库b经历体外转录以产生其相应的rna库之前,4a'中的尿苷(u)允许在碱性条件下引物除去。
从侧翼为一个引物的随机文库的rna适配子对选择的示意图(方案2):
·子图1说明了用于rna适配子选择的输入rna的制备。
·子图2说明了靶点依赖性适配子连结,如果rna是适配子。
·子图3说明了通过库b的选择性扩增和碱性水解,由编码两个rna适配子的连接物进化库a和库b。
·子图4说明了从编码rna适配子的dna库制备rna库。
方案2:从侧翼为一个引物的随机文库的rna适配子对选择的示意图
实施例
将参考以下实验和对比实施例进一步详细描述本发明的所选实施方式。这些实施例仅用于说明目的,并且不旨在限制本发明的范围。
实施例1.制备用于selex的随机rna文库
包含在5’和3’端两者上侧接不同标签的随机化序列的两个单链dna文库(即文库a和文库b)被转化为双链dna文库,并且通过三轮聚合酶链式反应进行扩增。将扩增的双链dna文库在四种不同的脱氧核糖核苷三磷酸(即datp、dgtp、2'-氟代-2'-脱氧胞苷-5'-三磷酸、2'-氟-2'-脱氧尿苷-5'-三磷酸)的混合物中经历体外转录以产生相应的rna文库。然后使用5’-rna多磷酸酶将每个文库中的rna在其5’端去磷酸化。通过在94℃加热5min,然后冷却至22℃,从这些去磷酸化的rna库中制备用于下一轮选择的输入库。
实施例2.用三种人血清蛋白质验证rna适配子对选择
对于每个靶点,使5’和3’端两者上侧接不同标签的两个随机文库经历selex以富集适配子库(图1)。在用人血纤维蛋白溶酶原或补体7的第5轮selex后,将一对富集的适配子库(例如库a和库b)与在第1轮rna适配子选择中的不同量的靶点一起温育。随后的连接和扩增允许一对rna适配子被物理相连的连接产物可以通过凝胶电泳鉴定。
在图1中,电泳图中的峰面积(来自电泳分离的结果图)表示来自库a和b的两个rna寡核苷酸彼此共价相连并且随后逆转录并且扩增的dna寡核苷酸的量。因此,以峰面积形式的靶点依赖性增加表明靶点蛋白质在第一rna适配子对选择期间在附近募集更多rna寡核苷酸。考虑到邻近连接测定中靶点蛋白质的探针的邻近效应,我们合理地认为,如果rna寡核苷酸是一对适配子,则这种募集相当容易完成。示出了通过(子图a)人血纤维蛋白溶酶原以及通过(子图b)人补体7的rna适配子对富集。
使用从随机文库的rna适配子对选择的示意图中所示的截短的selex进行使用另一血清蛋白质的适配子富集。在两轮适配子富集后,使库a和b经历适配子对选择。图4示出了一对库如何通过表示编码两个rna寡核苷酸(rna连结的结果)的dna寡核苷酸的量来响应靶点蛋白质。在25nm蛋白质的情况下,人血清蛋白质增强了rna连结,因此将扩增的连接的双链dna的量增加至高达1.8倍。在50nm蛋白质的情况下,扩增的连接物减少量表明过量的靶点蛋白质对靶点驱动的适配子连结具有负面影响。
图3示出了具有1:1摩尔比的混合的库a和b的解离常数(kd)。所示出的两个混合库是(1)未经历任何适配子对选择的那些以及(2)通过第3轮适配子对选择而富集的那些。两个混合库之间难以区分的kd值表明成对的适配子在适配子对多轮选择期间保持在库a和b的每个中。
图4显示了用三个不同的适配子对库作为探针进行的pla的灵敏度,(1)库a和b未经历适配子对选择,(2)通过第2轮适配子对选择富集的库a和b,以及(3)通过第3轮适配子对选择富集的库a和b。未经历适配子对选择的适配子库完全不响应人血清蛋白。通过第3轮适配子对选择富集的适配子库对人血清蛋白的响应比通过第2轮适配子对选择的那些更灵敏。它表示随着多轮选择的进行,成对的适配子进一步被富集。
实施例3.使用本发明在给定靶点的存在下优化适配子对选择平台
在每轮选择中监测适配子对富集使得可以调节应用于下一轮选择的选择压力。例如,选择的压力可以在多轮选择中保持恒定,直到观察到的连接产物的量显着增加。然后,在接下来的选择中的一对库将暴露于非常严格的条件,诸如低量的靶点,以允许一个库中富集的适配子彼此竞争寻找在另一个库中它们的对。最终,这将导致缩短选择轮次的数量以获得最佳适配子对。
本发明还增强了对用于适配子对选择的初始库的理解。在给定量的靶点蛋白质的情况下,若干不同的初始库(例如第3、第5和第7轮适配子富集的库)平行进行,预期在寻找适配子对中的高成功率。例如,如果仅第3轮库在寻找适配子对中产生阳性结果,那么阴性结果(例如第5和第7轮适配子富集的库)被解释为单独的适配子在与适配子对竞争中胜出。如果仅第7轮库产生阳性结果,那么额外的适配子对选择方法轮次可以在第3和第5轮两轮适配子富集的库中给出阳性结果。这类数据并未对初始库中所需的适配子富集循环的数量达成任何共识,但该数据通过建议个体适配子富集的步骤中允许的最大量的pcr循环,有助于大大减少系统误差,诸如pcr伪影/伪像。
最初使用邻近连接测定(pla)评估适配子富集的库,其与本适配子对选择方案高度兼容。在我们的技术中公开用作适配子对选择的平台的pla的专利出版物如下。us7306904b2是对于pla的提交的第一个专利。us20080293051a1教导了用rna作为探针的pla。该测定除了rna用作探针并且使用rna连接酶代替dna连接酶之外,与us7306904b2所公开的测定相同。公开截短selex以最小化我们的技术中使用的固定序列的参与的出版物是wo2000056930a1。pla测定也是高度灵敏的,通常表现出在低阿摩尔范围内的检测下限(lod)。
然后,进行对于适配子富集库的pla动态范围的筛选。在评估pla响应(qpcr读数)的同时,蛋白质量在多个数量级上变化,从1amol到1nmol(10-18到10-9mol)。在确定测定的近似动态范围后,进一步细化测定范围,并且在该窄范围(至少10种不同的蛋白质浓度)下进行pla。最终,测量诸如lod、loq、动态范围和灵敏度的性能指标。
选择的对将在多轮选择期间协同进化,以便预期通过对连接的双链dna测序来完成对来自库的适配子对的鉴定。来自连接的双链dna的所得到的rna可以是一种新的配体,在其折叠机制不受适配子的共价连接影响的情况下,增强了其从两个rna适配子继承的亲和力和特异性。
通过引用并入
上文引用了许多出版物,所有出版物都通过引用整体并入本文。
- 还没有人留言评论。精彩留言会获得点赞!