大麦HvAIR12基因及其用途的制作方法
本发明涉及一种大麦基因,具体是涉及一种青藏高原一年生野生大麦基因型xz16和xz61及栽培大麦品种dayton中hvair12基因及其用途,属于基因工程
技术领域:
。
背景技术:
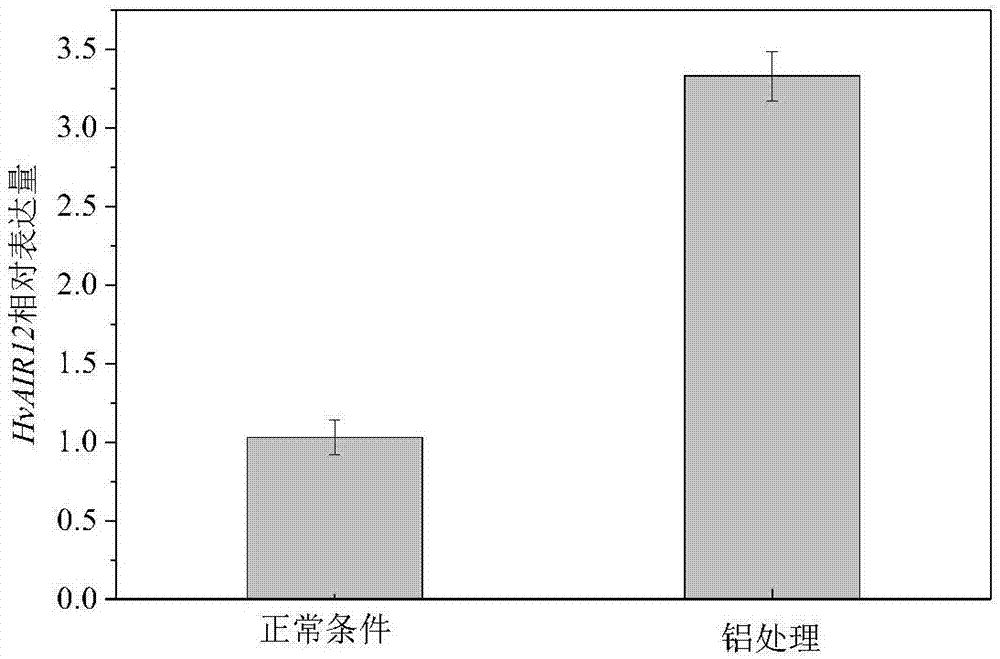
全世界大约有30%的可用耕地以及50%的潜在耕地土壤为酸性土壤,在酸性土壤条件下,铝毒害是非常重要的限制作物产量和植物生长的因子(kochian等,2004)。铝作为地壳中除了氧和硅之外含量最高的金属元素,一般以难溶性的硅酸盐或者氧化铝的形式存在于土壤中,此时铝的存在形态对植物无毒害作用,但当土壤ph小于5.5时,铝会从矿物态解离为离子态(其中以al3+离子毒性最强)释放进土壤,抑制植物根系的生长和功能,影响根系对水分和矿质元素的吸收,进而降低植物的产量(lou等,2016)。在早期,air12(auxin-inducedinrootcultures12)被认为是早期生长素响应基因aux/iaas家族成员(woodward等,2005),但意外的是,近年来的研究表明,air12被鉴定为是还原型抗坏血酸细胞色素b家族成员,这个结论在大豆、菜豆和拟南芥中都得到验证(preger等,2009)。air12具有糖基磷脂酰肌醇(glycosylphosphatidylinositol,gpi)锚定区域(borner等,2003),并且和其他氧化还原蛋白一起与脂筏连接(lefebvre等,2007),共同构成细胞膜内外的氧化还原链,调控下游ros信号转导,诱导根的形态建成。alex等的研究表明,过表达的air12在叶片中积累ros(过氧化物和过氧化氢)和脂质过氧化物,说明在特定条件下,air12可能改变了质外体中的氧化还原状态,将air12敲除,植物降低了对灰霉菌感染的敏感性(alex等,2015)。biniek(2017)等研究表明,nqr通过醌类物质和air12互相作用,使细胞溶质的nad(p)h电子传递到质外体的单脱氢抗坏血酸中,从而调控活性氧的水平和质外体的氧化还原状态。vigs(virus-inducedgenesilencing),病毒诱导的基因沉默,最初是一种由rna介导的植物抗病毒机制,现在它成为了一个重要的以反向遗传学研究基因功能的方法(holzberg等,2002)。bsmv-vigs除了转化效率高,性价比高之外,它和基于稳定遗传转化的rnai没有本质上的区别。bsmv-vigs的最主要的优点就是快,并且与rnai相比,它还具有很好的容纳较大目的基因的能力。目前为止,已经在一些重要的单子叶粮食作物如大麦、小麦(triticumaestivuml.),水稻和玉米(zeamaysl.)上得到了成功应用,弥补了其它一些vigs载体不能应用于单子叶粮食作物的不足,是近年来vigs研究的一个热点(lee等,2012),并且本课题组已经在野生大麦上建立了相关体系和应用。中国青藏高原是栽培大麦起源地之一(dai等,2012),青藏高原一年生野生大麦(hordeumvulgarel.ssp.spontaneum)为人们提供了丰富的基因库,并且由于其所处环境的原因,青藏高原一年生野生大麦具有自己独特的抗性基因调控网络,因此从野生大麦中发掘优异的抗性基因,进而进行应用,对作物品种改良及大麦育种具有重要的意义。技术实现要素:本发明提供一种大麦hvair12基因,该基因的cds区核苷酸序列如seqidno:1所示。大麦hvair12cdna来源于青藏高原一年生野生大麦(hordeumvulgarel.ssp.spontaneum,六棱大麦)xz16(耐酸(铝)基因型)、xz61(酸(铝)敏感基因型)和耐酸(铝)栽培大麦品种dayton。本发明还提供了上述hvair12基因编码的蛋白质,为seqidno:2所示的氨基酸序列。hvair12氨基酸序列来源于xz16、xz61和dayton。本发明还同时提供了一种bsmv:hvair12重组载体,将seqidno:3所示的241bp的hvair12基因片段连入rnaγ载体的nhei位点间,从而构建而得。seqidno:3为用于构建bsmv:hvair12重组载体的hvair12基因片段,来源于xz16。本发明还同时提供了所述大麦hvair12基因在增强大麦对酸铝耐受性方面的用途。该基因受到酸铝的诱导,沉默后使大麦对酸铝的耐受性显著降低。本发明利用重组质粒bsmv:hvair12在xz16上研究hvair12基因的用途。本发明以课题组前期筛选到的青藏高原一年生野生大麦xz16(耐酸(铝)基因型)为主要材料(戴华鑫,青藏高原一年生野生大麦耐铝毒机制的研究,2013年博士论文),克隆酸铝胁迫下大麦耐铝相关基因,对阐明大麦响应酸铝胁迫的分子机理和育种及生产具有重要意义。本发明分离克隆了大麦hvair12基因的cds区序列如seqidno:1所示,编码的蛋白质的氨基酸序列如seqidno:2所示。hvair12基因克隆与分析:基于根部转录组测序结果挑选出备选基因auxin-inducedproteinair12,从xz16、xz61和dayton中克隆了该基因的全长cds区序列,命名为hvair12。hvair12基因cds区全长783bp,编码一个260aa的蛋白序列,该蛋白分子量为25.35kda,等电点pi=10.13。将hvair12蛋白序列通过smart(http://smart.embl-heidelberg.de/)网站和interpro(http://www.ebi.ac.uk/interpro/)网站进行功能域预测分析,结果显示该蛋白含有1个功能域:domon功能域。将hvair12蛋白序列进行进化树分析,结果显示该蛋白与水稻osair12位于同一进化枝上。对hvair12基因进行铝的诱导表达,结果显示该基因的表达受到铝的诱导。bsmv:hvair12重组载体构建:将一个241bp的hvair12基因片段连入rnaγ载体的nhei位点间。反向插入的阳性克隆一方面送公司测序,另一方面将测序正确的单克隆提质粒进行酶切验证,确保了重组载体的准确性。利用bsmv-vigs方法在野生大麦xz16上验证hvair12基因功能:用mlui限制性内切酶将rnaα和rnaγ:hvair12质粒充分酶切,用spei限制性内切酶将rnaβ质粒充分酶切,使其线性化,以保证后续体外转录生产bsmv病毒。使用体外转录试剂盒将纯化后的线性化产物进行体外转录,将体外转录的bsmv:hvair12接种两叶期野生大麦xz16的第二片叶。为了研究hvair12基因的功能并确定bsmv:hvair12沉默效果,共设置四个处理,分别是:叶片接种bsmv:hvair12(处理1),叶片接种bsmv:γ(处理2),叶片接种bsmv:hvair12并进行200μm铝处理(处理3),叶片接种bsmv:γ并进行200μm铝处理(处理4)。接种14d后,观察主根根尖的生长情况和植株地下部铝含量。铝处理条件下,与模拟接种bsmv:γ的植株相比,接种bsmv:hvair12的植株地下部的铝含量显著提高(处理3vs.4)。对于接种bsmv:hvair12的植株来说,无论是在正常还是铝处理条件下,与其对照植株相比,根系中hvair12基因的表达量分别被抑制了57%和76%(处理3vs.4,处理1vs.2),并且接种bsmv:hvair12植株(处理3)相对于对照植株(处理4),植株根系含有更多的铝离子并且主根根尖部分受到的损伤更加严重,这些结果说明该基因沉默后使大麦对酸铝的耐受性显著降低。本发明通过对大麦hvair12基因的克隆和分析,并结合bsmv-vigs技术在xz16上对该基因进行功能验证发现,hvair12基因的表达会在铝诱导下显著提高,沉默hvair12基因后的xz16植株体内包含更多的铝离子,对酸铝的耐受性显著降低。本发明为大麦耐酸铝育种与生产提供了理论依据和相关基因。附图说明图1是铝处理和正常条件下zx16中的hvair12基因表达量。图2是三个大麦基因型(zx16,zx61和dayton)的核苷酸序列对比。图3是hvair12功能域预测图;数字代表氨基酸序列位点。图4是hvair12与水稻osair12、玉米zmair12和拟南芥atair12氨基酸序列对比,还包括四者在内的其他物种所构建的进化树。图5是bsmv:hvair12载体构建图、线性化图和体外转录图。(a)bsmv:hvair12载体示意图。(b)rnaγ:hvair12酶切产物琼脂糖凝胶电泳图。(c)rnaγ:hvair12线性化琼脂糖凝胶电泳图。(d)rnaγ:hvair12体外转录琼脂糖凝胶电泳图。1是rnaγ:hvair12,2是rnaγ:hvair12经mlui酶切,3是rnaγ:hvair12经nhei酶切,箭头所指为从载体上酶切下来的hvair12基因片段,4是rnaα经mlui酶切,5是rnaβ经spei酶切,6是rnaγ:hvair12经mlui酶切,7是体外转录的rnaα,8是体外转录的rnaβ,9是体外转录的rnaγ:hvair12。m1是15000bpdnamarker,m2是500bpdnamarker,m3是5000bpdnamarker。图6是利用bsmv-vigs方法在野生大麦xz16上验证hvair12基因的功能。(a)rt-pcr分析根中hvair12基因的表达情况。(b)icp法测定植株体内离子含量。(c)体视显微镜观察根尖表型。(d)桑色素染色植株根尖并计算荧光量。处理(1)是接种bsmv:hvair12并在基本培养液(bns)中生长14d,处理(2)是接种bsmv:γ并在bns中生长14d,处理(3)是接种bsmv:hvair12并在bns中生长7d并进行200μm铝处理7d,处理(4)是接种bsmv:γ并在bns中生长7d并进行200μm铝处理7d。具体实施方式下面通过具体实施例,对本发明的技术方案作进一步的具体说明。应当理解,本发明的实施并不局限于下面的实施例,对本发明所做的任何形式上的变通和/或改变都将落入本发明保护范围。在本发明中,若非特指,所有的份、百分比均为重量单位,所采用的设备和原料等均可从市场购得或是本领域常用的。下述实施例中的方法,如无特别说明,均为本领域的常规方法。实施例1、hvair12基因cds区和启动子序列的克隆和分析1、大麦生长条件青藏高原一年生野生大麦xz16、xz61和栽培大麦品种dayton种子用2%的h2o2消毒30min,随后用蒸馏水冲洗5-6次,将消毒过的种子置于铺有双层湿润滤纸的发芽盒上,发芽盒由收纳盒改造,盖子上打孔,发芽盒内加入蒸馏水并通气,于生长室中进行暗培养(22℃/18℃),萌发之后进行补光(22℃/18℃,白天/夜间)。第6d时,选取长势一致的幼苗转移至1l盛有大麦基本培养液的黑色塑料桶中,用5孔塑料盖覆盖,每孔两株幼苗,使用海绵进行固定,在大麦培养室中进行培养,大麦基本培养液使用1/5hogland配方。将预培养3d后的zx16植株进行铝胁迫(200μmal溶于基本营养液,ph4.3)试验,以基本营养液(ph4.3)为对照,处理时间为24h,进行hvair12基因的铝诱导表达。通过qrt-pcr的结果显示,zx16中的hvair12基因受到铝的诱导后,表达量提高了2.33倍(图1)。2、hvair12基因cds区序列的克隆根据rna提取试剂盒(takara,日本)的说明书进行xz16、xz61和dayton根部总rna的提取,并用dnasei去除总rna中基因组dna污染,将提取的总rna反转录成cdna。根据blast序列进行特异性引物设计,具体引物序列为:hvair12-cds-f:atggcctccacgatgccgchvair12-cds-r:tcatactaccgcgaggaggc将扩增出来跑胶验证之后的产物,进行胶回收连接pmd18-t载体,转化大肠杆菌dh5α,将阳性克隆送至公司测序,测序正确的分别进行提质粒和甘油保存。xz16、xz61和dayton三个大麦基因型的cds区核苷酸序列完全相同,遂统称为hvair12(图2),所得质粒命名为pmd18-t-hvair12质粒。pcr引物由生工生物工程(上海)股份有限公司完成,基因测序工作由上海铂尚生物技术有限公司完成。大麦hvair12基因的cds区核苷酸序列如seqidno:1所示。3、hvair12基因序列分析将hvair12蛋白序列通过smart(http://smart.embl-heidelberg.de/)网站进行功能域的预测和分析,结果显示该蛋白只含有一个功能域:duf568功能域(图3),但是dufdomain在此网站上没有详细介绍,功能也不明确。随后将hvair12蛋白序列通过interpro(http://www.ebi.ac.uk/interpro/)网站进行功能域预测分析,结果显示该蛋白含有一个功能域:domondomain,其与纤维乙糖脱氢酶和细胞色素domain存在着从属关系。通过clustalw和mega6软件将包含了hvair12在内的不同物种中的air12序列和拟南芥中不同air家族蛋白序列进行分析,构建系统发育树。结果显示hvair12在进化上与水稻osair12位于同一进化枝(图4)。实施例2、bsmv-vigs方法在野生大麦zx16中验证hvair12基因功能1、bsmv:hvair12载体构建利用rna提取试剂盒(takara,日本)的说明书进行正常生长条件下的xz16根部总rna的提取,并用dnasei去除总rna中基因组dna污染,根据primescripttmii1ststrandcdnasynthesiskit(takara,日本)说明书步骤将叶片rna反转录成cdna。以之前保存的pmd18-t-hvair12质粒为模板设计引物,扩增241bp的hvair12基因片段。kod酶pcr反应体系μl2×pcrbufferforkodfx252mmdntps1010pmol/μlhvair12-γ-f1.510pmol/μlhvair12-γ-r1.5templatedna1kodfx1ddh2o10总体积50扩增程序为:94℃2min,(98℃10s,59℃30s,68℃15s)35个循环,68℃5min。引物序列为(下划线为酶切位点):hvair12-γ-f:gtacgctagcacaacatcaccggctacgtghvair12-γ-r:gtacgctagcaggacgagcttggacttgga将hvair12基因片段连接到pmd18-t载体上,转化大肠杆菌dh5α,挑取阳性克隆送往公司测序,测序正确的单克隆进行摇菌提质粒和甘油保存。提取的pmd18-t-hvair12质粒使用nheⅰ限制性内切酶进行酶切,分别与经过同样限制酶内切酶酶切并且去磷酸化的病毒载体rnaγ:tapds载体(贺小彦,干旱胁迫下西藏野生大麦根毛生长调控基因hvexpb7的分离、克隆和功能鉴定,博士论文(2015))通过t4连接酶进行连接反应。连接之后的产物转化大肠杆菌dh5α,使用rnaγ载体上的引物γ-stain-f和hvair12基因的正向引物hvair12-γ-f验证反向插入,将反向插入的阳性克隆送到公司测序,测序正确的单克隆进行摇菌提质粒和甘油保存,提取的质粒进行酶切再次验证(图5b),将该质粒命名为rnaγ:hvair12。γ-stain-f:caactgccaatcgtgagtagg。pcr引物由生工生物工程(上海)股份有限公司完成,基因测序工作由上海铂尚生物技术有限公司完成。2、bsmv载体线性化和体外转录使用mluⅰ限制性内切酶单酶切rnaα、rnaγ和rnaγ:hvair12,同时使用speⅰ限制性内切酶单酶切rnaβ,分别使其线性化,酶切产物跑胶,胶回收纯化产物(图5c、d)。将纯化后的一定浓度的rnaα、rnaβ、rnaγ和rnaγ:hvair12按照ribomaxtmlargescalernaproductionsystem-t7kit和ribom7gcapanalogkit试剂盒实验说明书进行反转录(promega,美国),所有的操作需要保证无rnase的干扰。将体外转录后的rnaα、rnaβ和rnaγ/rnaγ:hvair12质粒按照1:1:1的体积比进行混合,加入三倍体积的rnase-free水稀释,随后在稀释产物中加入等体积的2×gkp缓冲液(1%bentonite、1%celite、50mm甘氨酸和30mm磷酸氢二钾ph9.2),混合均匀,用于随后的接种。所得产物命名为bsmv:hvair12。3、bsmv接种前大麦幼苗培养实验所需野生大麦zx16种子用2%的h2o2消毒30min,随后用蒸馏水冲洗5-6次,将消毒过的种子置于铺有双层湿润滤纸的发芽盒上,发芽盒由收纳盒改造,盖子上打孔,发芽盒内加入蒸馏水并通气,于生长室中进行暗培养(22℃/18℃),萌发之后进行补光(22℃/18℃)。第6d时,选取长势一致的幼苗转移至1l盛有大麦基本培养液的黑色塑料桶中,用5孔塑料盖覆盖,每孔两株幼苗,使用海绵进行固定,在大麦培养室中进行培养,大麦基本培养液使用1/5hogland配方。每3d更换一次培养液,培养液的ph用naoh或hcl调至5.9±0.1,整个培养过程用气泵持续通气。4、bsmv接种验证基因功能选取两叶期的大麦第二片叶子在rnase-free环境下进行bsmv摩擦接种,每个植株使用8μl的上述混合物,具体为:以rnaα+rnaβ+rnaγ+gkp混合物bsmv:γ为对照,以rnaα+rnaβ+rnaγ:hvair12+gkp混合物bsmv:hvair12为处理。接种后的植物随即喷施适量的depc水,并用透明的塑料罩保湿,保湿3d后,将玻璃罩取掉,在大麦生长培养室中继续培养(22℃/18℃,白天/夜间),定时观察植株的表型。本实验共有4个处理:接种bsmv:hvair12并在基本培养液(bns)中生长14d(处理1),接种bsmv:γ并在基本培养液(bns)中生长14d(处理2),接种bsmv:hvair12并在基本培养液(bns)中生长7d,再进行200μm铝处理7d(处理3),接种bsmv:γ并在基本培养液(bns)中生长7d,再进行200μm铝处理7d(处理4)。每个处理3个重复,每个重复8个植株。接种14d后,观察主根根尖的生长情况和植株的铝含量。铝处理条件下,与模拟接种bsmv:γ的植株相比,接种bsmv:hvair12的植株地下部的铝含量显著提高(处理3vs.4)(图6b),并且接种bsmv:hvair12植株(处理3)相对于对照植株(处理4),主根根尖部分受到的损伤更加严重并且含有更多的铝离子(图6c、d),这些结果表明,沉默hvair12基因后的xz16植株体内包含更多的铝离子,对酸铝的耐受性显著降低。5、rt-pcr验证hvair12基因的表达量利用rna提取试剂盒(takara,日本)的说明书分别提取xz16不同处理样本的rna,并用dnasei去除总rna中基因组dna污染,用primescripttmrtreagentkit(takara,日本)将各样本rna分别反转录成cdna。利用sybr绿色荧光酶复合物(takara,日本)和lightcycler480pcr仪(roche,瑞士)对相应样本中hvair12基因的表达进行荧光定量pcr分析(qrt-pcr),并用一个内参基因actin对表达值进行矫正处理。pcr体系为:takarasybr绿色荧光酶体系μlsybrpermixextaqⅱ(tlirnasehplus)(2×)10hvair12-rt-pcr-f/actin-f0.8hvair12-rt-pcr-r/actin-r0.8cdna2ddh2o6.4总体积20pcr的具体程序为:95℃30s,(95℃5s,60℃30s)40个循环。溶解曲线程序为:95℃5s,60℃1min,95℃,50℃30s降温。利用2-δδcq相对定量方法计算基因表达值变化。每组实验重复三次。rt-pcr引物序列为:hvair12-rt-pcr-f:5'-ggggaagacgtacgccaagt-3',hvair12-rt-pcr-r:5'-cacgaacgccagcgagag-3',actin-f:5'-aagcatgaagatacagggagtgtg-3',actin-r:5'-aaatttattctcggaagaggttgtaca-3'。所得结果为:对于接种bsmv:hvair12的植株来说,无论是在正常还是铝处理条件下,与其对照植株相比,根系中hvair12基因的表达量分别被抑制了57%和76%(处理3vs.4,处理1vs.2)(图6a)。综上所述,通过对大麦hvair12的克隆和分析,并结合bsmv-vigs技术在青藏高原一年生野生大麦xz16上对该基因进行功能验证发现,hvair12基因的表达会在铝诱导下显著提高,沉默hvair12基因后的xz16植株体内包含更多的铝离子,对酸铝的耐受性显著降低。最后,还需要注意的是,以上列举的仅是本发明的若干个具体实施例。显然,本发明不限于以上实施例,还可以有许多变形。本领域的普通技术人员能从本发明公开的内容直接导出或联想到的所有变形,均应认为是本发明的保护范围。序列表<110>浙江大学<120>大麦hvair12基因及其用途<160>3<170>siposequencelisting1.0<210>1<211>783<212>dna<213>青藏高原一年生野生大麦(hordeumvulgarel.ssp.spontaneum)<400>1atggcctccacgatgccgcgccacatcgcgctgctgttgctcctgctcctcgtggcgtcc60acgcgcgcggcgtccgcggccggcgccggcgcgtgcgcggccgagaagttcccggcgggg120aagacgtacgccaagtgcgaggacctcccgcagctgggggccgcgctgcactggacctac180gacgagtccaagtcgtccctctcgctggcgttcgtggccgcgccggcgggggccaacggg240tgggtggcctgggcgctcaaccccacgggcgagggcatggccggcgcgcaggcgctcgtg300gcgctcaagggctccggcgccgccgcgcccaccgtcaggacgtacaacatcaccggctac360gtgccgctcggcaaggcctccacgcccatcgcgttcccggccaccgacctcgccgccgac420tccggcagcgccggcaagatccggctctacggcaagctgcagctgcacagcggcatgaag480gccgtcaaccatatatggcaggtcggcacctccgtcaccgccggggcccccgacaagcac540gctttcgcccccggcaacctcgcctccaagtccaagctcgtcctctcgggcaaagcggct600tccgcgacctcgccttcgccgtcgcccgcccccatggccggtggcccgtccggaagcgac660gccggcgcggcggcgaccacggcgccttctgccggcaagtccccgtcggccgcggccgcg720gccggcgtgtcggcgccggcactccttgtgctcgcgttgatgggcctcctcgcggtagta780tga783<210>2<211>260<212>prt<213>青藏高原一年生野生大麦(hordeumvulgarel.ssp.spontaneum)<400>2metalaserthrmetproarghisilealaleuleuleuleuleuleu151015leuvalalaserthrargalaalaseralaalaglyalaglyalacys202530alaalaglulyspheproalaglylysthrtyralalyscysgluasp354045leuproglnleuglyalaalaleuhistrpthrtyraspgluserlys505560serserleuserleualaphevalalaalaproalaglyalaasngly65707580trpvalalatrpalaleuasnprothrglygluglymetalaglyala859095glnalaleuvalalaleulysglyserglyalaalaalaprothrval100105110argthrtyrasnilethrglytyrvalproleuglylysalaserthr115120125proilealapheproalathraspleualaalaaspserglyserala130135140glylysileargleutyrglylysleuglnleuhisserglymetlys145150155160alavalasnhisiletrpglnvalglythrservalthralaglyala165170175proasplyshisalaphealaproglyasnleualaserlysserlys180185190leuvalleuserglylysalaalaseralathrserproserproser195200205proalaprometalaglyglyproserglyseraspalaglyalaala210215220alathrthralaproseralaglylysserproseralaalaalaala225230235240alaglyvalseralaproalaleuleuvalleualaleumetglyleu245250255leualavalval260<210>3<211>241<212>dna<213>青藏高原一年生野生大麦(hordeumvulgarel.ssp.spontaneum)<400>3acaacatcaccggctacgtgccgctcggcaaggcctccacgcccatcgcgttcccggcca60ccgacctcgccgccgactccggcagcgccggcaagatccggctctacggcaagctgcagc120tgcacagcggcatgaaggccgtcaaccatatatggcaggtcggcacctccgtcaccgccg180gggcccccgacaagcacgctttcgcccccggcaacctcgcctccaagtccaagctcgtcc240t241当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1