一种比酶活提高的肝素酶I的定向改造酶及分子改造方法和表达工程菌与流程
本发明属于基因工程和蛋白质表达技术领域,涉及phhepia259d和phhepis169d/a259d酶基因的构建和表达,以及该酶的性质和制备研究,尤其是一种比酶活提高的肝素酶i的定向改造酶及分子改造方法和表达工程菌。
背景技术:
肝素(heparin)在自然界中广泛存在,是动物体内一种天然抗凝血物质。最开始它是在肝脏中被发现,所以被命名为肝素。它广泛分布在哺乳动物组织中,比如肝、肺、肠粘膜、心、胎盘和肌肉等,以共价键形式和蛋白结合。目前商业化的肝素主要从牛肺和猪肠粘膜中提取得到。低分子肝素(lowmolecularweightheparin,lmwh)是由肝素经过一定的化学物理方法裂解后产生的小肝素片段,低分子量肝素与蛋白或细胞的结合作用力下降,但是在抗凝血活性方面会增强。同普通肝素相比,lmwh能降低抗因子ⅱa的活性。lmwh具有发生出血率低,体内生物利用度高等优点。它是近些年来临床应用发展极快的抗凝血和抗血栓类药物。
低分子肝素的制备可以分为物理方法,化学方法,生物法以及合成法。化学解聚法技术比较成熟且应用广泛,但是存在一些缺陷,如生产过程溶液对环境造成污染,且反应过程中需要的强氧化剂和激烈的反应条件可能会导致lmwh的药理活性降低。生物解聚法与化学解聚法相比具有反应条件温和、选择性强、对环境污染小等特点,所以生物解聚法越来越受到人们关注。
目前肝素类药物的市场需求日益增多,2016年肝素原料药的需求达到44.30万亿单位,全球肝素类药物市场已达到125.8亿美元,市场巨大,并且持续增加。肝素类药物的发展方向趋向于低分子肝素和超低分子肝素,目前对于低分子肝素的制备,大都采用化学裂解法,但是此方法存在组分单一性差,分离成本高;三废污染,纯度差等缺点,越来越不适合绿色环保的理念。而酶解法条件温和,对肝素本身的结构特征不造成影响,且没有副反应发生,得率较高;反应一步完成,比较容易实现生产;反应没有产生不纯物,比较利于下游的分离纯化,其中以肝素酶i的酶解应用最为广泛。目前市场上肝素酶i的价格极其昂贵,并且其本身的酶活性较差,很难适应工业化生产的要求。
肝素酶i(heparanase)是一类能够裂解肝素类结构物质的多糖裂解酶,主要用于制备低分子肝素以及肝素类物质分子的机构测定。肝素酶i分子量大小约为42kd,它既存在于原核生物,也存在于真核生物中。原核生物来源的肝素酶i主要来自于肝素黄杆菌(flavobacteriumheparinum),还包括一些拟杆菌和芽孢杆菌等,它在制备低分子量肝素、清除体外血液循环中肝素和确定肝素精细结构方面有着重要作用。目前对于肝素酶i的研究大都来自于肝素黄杆菌,但基于理性设计的肝素酶i的分子改造,尤其是关于肝素酶i比酶活的分子改造研究未见有报道。
通过检索,尚未发现与本发明专利申请相关的专利公开文献。
技术实现要素:
本发明的目的在于克服现有技术的不足之处,克服当前肝素酶i产量低、活性低的缺点,提供一种比酶活提高的肝素酶i的定向改造酶及分子改造方法和表达工程菌,与野生型相比,本发明定向改造酶具有更高的比酶活,有较大应用潜力,也为hepi的研究奠定了理论基础。
本发明解决其技术问题是采取以下技术方案实现的:
一种比酶活提高的肝素酶i的定向改造酶phhepia259d,其核苷酸序列为seqidno:1。
如上所述的比酶活提高的肝素酶i的定向改造酶phhepia259d编码的高活性肝素酶i,其完整的氨基酸序列为seqidno:2。
一种比酶活提高的肝素酶i的定向改造酶phhepis169d/a259d,其核苷酸序列为seqidno:3。
如上所述的比酶活提高的肝素酶i的定向改造酶phhepis169d/a259d编码的高活性肝素酶i,其完整的氨基酸序列为seqidno:4。
一种高活性肝素酶i的分子改造方法,步骤如下:
通过以多形拟杆菌来源的肝素酶ibthepi为模板,利用同源建模和序列同源性比对肝素黄杆菌来源的肝素酶iphhepi进行分析,对其进行定向改造,得到比酶活显著提高的高活性肝素酶iphhepia259d和/或phhepis169d/a259d。
而且,具体步骤如下:
⑴phhepi的空间结构的模拟:登录蛋白质结构数据库,经过blast比对,找到与phhepi具有高度同源性的序列:选择已经具有晶体结构的bthepi为模板,两者的序列同源性为62.5%,运用swiss-model服务器在线对phhepi进行模拟空间结构;运用在线网站对同源建模的结构模型进行评估和优化,获得准确性更高的模拟结构;
⑵比酶活提高的phhepi突变体的理性设计:已知bthepi序列中155位-asp、253位-asp的氨基酸为重要氨基酸,分别处于钙离子结合区域以及底物结合区域;通过将phhepi和bthepi的结构和序列进行比对分析以及预测,在phhepi结构序列中对应的氨基酸残基位点是169位-ser和259位-ala,综合以上因素选择将phhepi序列中的169位丝氨酸突变为天冬氨酸、259位丙氨酸突变为天冬氨酸以及两个位点的复合突变。
而且,所述蛋白结构数据库网址为:htpp://rcsb.org;所述具有晶体结构的bthepi,其proteindatabank登录号为3ikw;所述在线网站为:http://services.mbi.ucla.edu/saves/。
一种含突变基因phhepia259d和/或phhepis169d/a259d表达工程菌,其构建方法,步骤如下:
基于突变位点,设计并合成定点突变的引物如下:
s169d-f:seqidno:5:
5′-cagtggcacggtgctccggatcgtactcttgttgctact-3′
s169d-r:seqidno:6:
5′-agtagcaacaagagtacgatccggagcaccgtgccactg-3′
a259d-f:seqidno:7:5′-gtggcttactgataaagatgatcgtaacaacgcta-3′
a259d-r:seqidno:8:5′-tagcgttgttacgatcatctttatcagtaagccca-3′
以含有phhepi基因的质粒pe-sumo-phhepi为模板,使用以上引物进行pcr,反应条件为:95℃2min;95℃30s,60℃30s,72℃7min,30个循环;72℃5min;对其pcr产物进行dnpi酶消化,进行核酸电泳以及切胶回收,将其转化入dh5α感受态,通过抗生素筛选提取阳性克隆子的质粒并送去测序;将测序正确的表达质粒pe-sumo-phhepia169d、pe-sumo-phhepia259d、pe-sumo-phhepis169d/a259d转化入表达菌株rosettade3中,即可成功构建phhepi突变体的表达工程菌。
一种如上所述的含突变基因phhepia259d和/或phhepis169d/a259d表达工程菌的高效表达方法,步骤如下:
将含突变基因phhepia259d和/或phhepis169d/a259d表达工程菌分别接种于含卡那霉素和氯霉素抗性的lb培养基,37℃,220r/min培养过夜;
其中,所述lb培养基的配方为:nacl10g/l,酵母提取物5g/l,蛋白胨10g/l,含50ug/l卡那霉素和34ug/l氯霉素;
将过夜培养菌液按1%接种量接入lb培养基中,37℃,220r/min培养至od600为0.6-0.8时,加入终浓度为0.4mm的iptg,在25℃,200r/min条件下诱导12h;4℃8000r/min离心10min,收集菌体,用缓冲液洗涤两遍,再将其悬于40ml缓冲液中,超声破碎细胞;12000r/min、4℃离心20min,收集上清,即可;
其中,所述缓冲液的配方为:20mmol/ltris-hcl,200mmol/lnacl,ph7.4。
一种如上所述的含突变基因phhepia259d和/或phhepis169d/a259d表达工程菌的高效纯化方法,步骤如下:
使用co-nta亲和层析进行纯化,co柱使用10ml的平衡缓冲液平衡后上样,并使用10ml结合缓冲液冲洗除掉非特异性结合蛋白,使用3ml洗脱缓冲液洗脱重组目的蛋白,收集洗脱液即为纯化后的重组酶;
使用pd-10预装脱盐柱对纯化后的酶进行脱盐处理;使用25ml的平衡缓冲液平衡后上样2.5ml,3.5ml缓冲液洗脱,从上样开始收集2.5-6ml的洗脱液为脱盐后的酶液;
其中,所述co柱使用的平衡缓冲液的配方为:20mmol/ltris-hcl,300mmol/lnacl,ph7.4;所述结合缓冲液的配方为:20mmol/ltris-hcl,300mmol/lnacl,5mmol/l咪唑;所述洗脱缓冲液的配方为:20mmol/ltris-hcl,300mmol/lnacl,150mmol/l咪唑;所述pd-10预装脱盐柱使用的平衡缓冲液的配方为:20mmol/ltris-hcl,200mmol/lnacl,ph7.4。
本发明取得的优点和积极效果是:
1、本发明提供了两种phhepia259d和phhepis169d/a259d突变酶的设计与构建方法,新型突变酶工程菌的构建以及突变酶的高效表达的方法。与野生型相比,本发明突变酶具有更高的比酶活,有较大应用潜力,也为hepi的研究奠定了理论基础。
2、本发明利用同源建模和序列同源性分析对肝素酶i的比酶活进行理性改造,构建出能高效表达高活性的肝素酶i突变体工程菌并实现重组肝素酶i突变体的高效表达。由定向改造得到的突变肝素酶i命名为phhepia259d和phhepis169d/a259d,其相应的基因为phhepia259d、phhepis169d/a259d,与野生想相比突变体的比酶活得到了明显的提高,分别提高40.43%和122.05%。具有较大的工业应用潜力以及经济价值
3、本发明利用同源建模和序列同源性分析方法对肝素酶i进行定向改造,运用基因工程技术构建出高效表达可溶性的突变肝素酶i工程菌。将来源于肝素黄杆菌(pedobacterheparinus)的肝素酶i(genbank:l12534.1)进行提高比酶活的定向改造,获得了两种肝素酶i突变基因命名为phhepia259d和phhepis169d/a259d。本发明还公开了突变酶工程菌的构建以及高效表达和纯化方法,制备的突变酶具有更高比酶活的特性,并且突变前后的酶学性质未发生改变,具有较大的工业化生产潜力。
附图说明
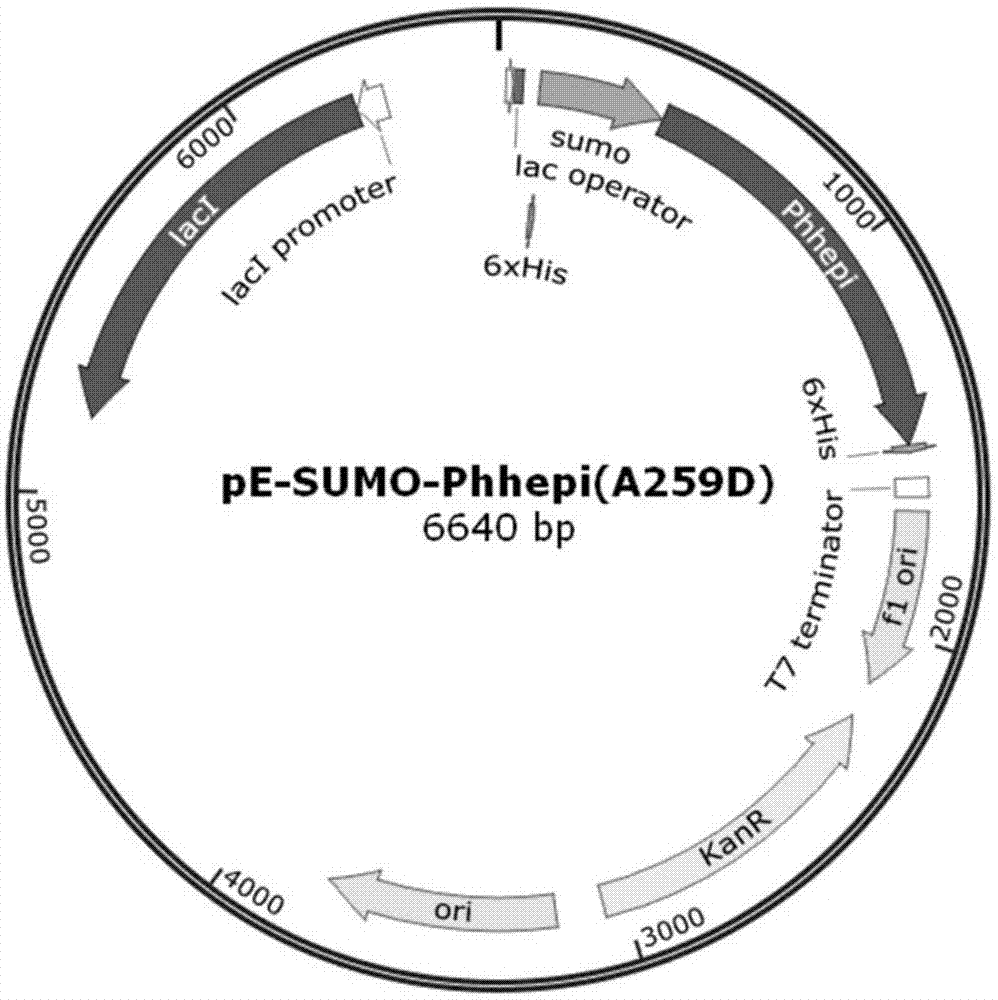
图1为本发明中重组质粒pe-sumo-phhepia259d构建示意图;
图2为本发明中重组质粒pe-sumo-phhepis169d/a259d的构建示意图;
图3为本发明中重组突变酶phhepia259d和phhepis169d/a259d的表达和纯化的sds-page图;
其中:m、蛋白质分子量标准,条带自上至下大小为170kd,130kd,100kd,70kd,55kd,40kd,35kd,25kd,15kd;泳道1、pe-sumo-phhepia259d重组菌的超声破壁上清,上样20ul,泳道2、pe-sumo-phhepia259d表达产物的纯化,上样20ul,泳道3、pe-sumo-phhepis169d/a259d重组菌的超声破壁上清,上样量20ul,泳道4、pe-sumo-phhepis169d/a259d表达产物的纯化;
图4为本发明中温度对肝素酶i突变前后活性的影响图;
图5为本发明中ph对肝素酶i突变前后活性的影响图;
图6为本发明中钙离子对肝素酶i突变前后活性的影响图;
图7为本发明中sumo-phhepia259d结构解析示意图;
图8为本发明中sumo-phhepis169d/a259d结构解析示意图。
具体实施方式
下面结合通过具体实施例对本发明作进一步详述,以下实施例只是描述性的,不是限定性的,不能以此限定本发明的保护范围。
本发明中所使用的原料,如无特殊说明,均为常规的市售产品;本发明中所使用的方法,如无特殊说明,均为本领域的常规方法。
一种比酶活提高的肝素酶i的定向改造酶phhepia259d,其核苷酸序列为seqidno:1。
如上所述的比酶活提高的肝素酶i的定向改造酶phhepia259d编码的高活性肝素酶i,其完整的氨基酸序列为seqidno:2。
一种比酶活提高的肝素酶i的定向改造酶phhepis169d/a259d,其核苷酸序列为seqidno:3。
如上所述的比酶活提高的肝素酶i的定向改造酶phhepis169d/a259d编码的高活性肝素酶i,其完整的氨基酸序列为seqidno:4。
一种高活性肝素酶i的分子改造方法,步骤如下:
通过以多形拟杆菌来源的肝素酶ibthepi为模板,利用同源建模和序列同源性比对肝素黄杆菌来源的肝素酶iphhepi进行分析,对其进行定向改造,得到比酶活显著提高的高活性肝素酶iphhepia259d和/或phhepis169d/a259d。
较优地,具体步骤如下:
⑴phhepi的空间结构的模拟:登录蛋白质结构数据库,经过blast比对,找到与phhepi具有高度同源性的序列:选择已经具有晶体结构的bthepi为模板,两者的序列同源性为62.5%,运用swiss-model服务器在线对phhepi进行模拟空间结构;运用在线网站对同源建模的结构模型进行评估和优化,获得准确性更高的模拟结构;
⑵比酶活提高的phhepi突变体的理性设计:已知bthepi序列中155位-asp、253位-asp的氨基酸为重要氨基酸,分别处于钙离子结合区域以及底物结合区域;通过将phhepi和bthepi的结构和序列进行比对分析以及预测,在phhepi结构序列中对应的氨基酸残基位点是169位-ser和259位-ala,综合以上因素选择将phhepi序列中的169位丝氨酸突变为天冬氨酸、259位丙氨酸突变为天冬氨酸以及两个位点的复合突变。
而且,所述蛋白结构数据库网址为:htpp://rcsb.org;所述具有晶体结构的bthepi,其proteindatabank登录号为3ikw;所述在线网站为:http://services.mbi.ucla.edu/saves/。
一种含突变基因phhepia259d和/或phhepis169d/a259d表达工程菌,其构建方法,步骤如下:
基于突变位点,设计并合成定点突变的引物如下:
s169d-f:seqidno:5:
5′-cagtggcacggtgctccggatcgtactcttgttgctact-3′
s169d-r:seqidno:6:
5′-agtagcaacaagagtacgatccggagcaccgtgccactg-3′
a259d-f:seqidno:7:5′-gtggcttactgataaagatgatcgtaacaacgcta-3′
a259d-r:seqidno:8:5′-tagcgttgttacgatcatctttatcagtaagccca-3′
以含有phhepi基因的质粒pe-sumo-phhepi为模板,使用以上引物进行pcr,反应条件为:95℃2min;95℃30s,60℃30s,72℃7min,30个循环;72℃5min;对其pcr产物进行dnpi酶消化,进行核酸电泳以及切胶回收,将其转化入dh5α感受态,通过抗生素筛选提取阳性克隆子的质粒并送去测序;将测序正确的表达质粒pe-sumo-phhepia169d(已申请专利,专利申请公布号:cn106497897a)、pe-sumo-phhepia259d、pe-sumo-phhepis169d/a259d转化入表达菌株rosettade3中,即可成功构建phhepi突变体的表达工程菌。
一种如上所述的含突变基因phhepia259d和/或phhepis169d/a259d表达工程菌的高效表达方法,步骤如下:
将含突变基因phhepia259d和/或phhepis169d/a259d表达工程菌分别接种于含卡那霉素和氯霉素抗性的lb培养基,37℃,220r/min培养过夜;
其中,所述lb培养基的配方为:nacl10g/l,酵母提取物5g/l,蛋白胨10g/l,含50ug/l卡那霉素和34ug/l氯霉素;
将过夜培养菌液按1%接种量接入lb培养基中,37℃,220r/min培养至od600为0.6-0.8时,加入终浓度为0.4mm的iptg,在25℃,200r/min条件下诱导12h;4℃8000r/min离心10min,收集菌体,用缓冲液洗涤两遍,再将其悬于40ml缓冲液中,超声破碎细胞;12000r/min、4℃离心20min,收集上清,即可;
其中,所述缓冲液的配方为:20mmol/ltris-hcl,200mmol/lnacl,ph7.4。
一种如上所述的含突变基因phhepia259d和/或phhepis169d/a259d表达工程菌的高效纯化方法,步骤如下:
使用co-nta亲和层析进行纯化,co柱使用10ml的平衡缓冲液平衡后上样,并使用10ml结合缓冲液冲洗除掉非特异性结合蛋白,使用3ml洗脱缓冲液洗脱重组目的蛋白,收集洗脱液即为纯化后的重组酶;
使用pd-10预装脱盐柱对纯化后的酶进行脱盐处理;使用25ml的平衡缓冲液平衡后上样2.5ml,3.5ml缓冲液洗脱,从上样开始收集2.5-6ml的洗脱液为脱盐后的酶液;
其中,所述co柱使用的平衡缓冲液的配方为:20mmol/ltris-hcl,300mmol/lnacl,ph7.4;所述结合缓冲液的配方为:20mmol/ltris-hcl,300mmol/lnacl,5mmol/l咪唑;所述洗脱缓冲液的配方为:20mmol/ltris-hcl,300mmol/lnacl,150mmol/l咪唑;所述pd-10预装脱盐柱使用的平衡缓冲液的配方为:20mmol/ltris-hcl,200mmol/lnacl,ph7.4。
具体地,相关操作如下:
本发明通过以多形拟杆菌来源的肝素酶i(bthepi)为模板,利用同源建模和序列同源性比对肝素黄杆菌来源的肝素酶i(phhepi)进行分析,对其进行定向改造,得到比酶活显著提高的肝素酶i(phhepia259d和phhepis169d/a259d),其核苷酸序列分别对应于序列表中的seqidno:1和seqidno:3;氨基酸序列对应序列表中的seqidno:2和seqidno:4。
所述的高活性肝素酶i突变体的设计和表达方法:
(1)phhepi的空间结构模拟:登录蛋白结构数据库(htpp://rcsb.org),经blast比对,找到与phhepi同源性为62.5%的多形拟杆菌肝素酶i(bthepi)(pdbid3ikw),以bthepi为模板,运用swiss-model服务器在线进行phhepi的空间结构模拟。运用在线网站(http://services.mbi.ucla.edu/saves/)对同源建模的结构模型进行评估和优化,获得准确性更高的模拟结构。
(2)高活性的肝素酶i突变体的理性设计:已知bthepi的晶体结构和氨基酸序列,通过结构和序列的比对,确定bthepi的155位和253位的氨基酸在空间结构上对应phhepi序列中的169位和259位,phhepi的两个位点的氨基酸残基与bthepi中的不同,并且这两个氨基酸是关键氨基酸,因此确定将phhepi的第169位丝氨酸突变为天冬氨酸、259位丙氨酸突变为天冬氨酸以及两个位点的复合突变。
(3)含突变基因phhepia259d、phhepis169d/a259d表达工程菌的构建:根据上述分析结果以及phhepi的基因序列设计2对突变引物:
s169d-f:5′-cagtggcacggtgctccggatcgtactcttgttgctact-3′
s169d-r:5′-agtagcaacaagagtacgatccggagcaccgtgccactg-3′
a259d-f:5′-gtggcttactgataaagatgatcgtaacaacgcta-3′
a259d-r:5′-tagcgttgttacgatcatctttatcagtaagccca-3′
采用重叠pcr技术构造含有突变酶基因phhepia259d、phhepis169d/a259d的表达质粒,分别以s169d-f、s169d-r和a259d-f、a259d-r为引物、以pe-sumo-phhepi为模板进行pcr(95℃2min;95℃30s,60℃30s,72℃7min,30个循环;72℃5min),pcr产物使用dnpi酶消化2h后,使用1%琼脂糖凝胶电泳进行分析并切胶回收。将回收产物进行重组连接,进而转化入dh5α感受态细胞,提取质粒送去测序。将测序正确的突变表达质粒转化入rosetta(de3)感受态细胞,通过抗性验证以及测序,成功得到表达突变酶的工程菌。
phhepia259d和phhepis169d/a259d的表达、纯化以及活性测定:
1、将工程菌分别接种于含卡那霉素和氯霉素抗性的lb培养基(nacl10g/l,酵母提取物5g/l,蛋白胨10g/l,含50ug/l卡那霉素和34ug/l氯霉素),37℃,220r/min培养过夜。将过夜培养菌液按1%接种量接入50mllb培养基中(250ml摇瓶),37℃,220r/min培养至od600约为0.6-0.8时,加入终浓度为0.4mm的iptg,在25℃,200r/min条件下诱导12h。4℃8000r/min离心10min,收集菌体,用缓冲液(20mmol/ltris-hcl,200mmol/lnacl,ph7.4)洗涤两遍,再将其悬于40ml缓冲液中,超声破碎细胞。12000r/min、4℃离心20min,收集上清,进行sds-page分析。
2、使用co-nta亲和层析进行纯化,co柱使用10ml的平衡缓冲液(20mmol/ltris-hcl,300mmol/lnacl,ph7.4)平衡后上样,并使用10ml结合缓冲液(20mmol/ltris-hcl,300mmol/lnacl,5mmol/l咪唑)冲洗除掉非特异性结合蛋白,使用3ml洗脱缓冲液(20mmol/ltris-hcl,300mmol/lnacl,150mmol/l咪唑)洗脱重组目的蛋白,收集洗脱液即为纯化后的重组酶。使用pd-10预装脱盐柱对纯化后的酶进行脱盐处理。使用25ml的平衡缓冲液(20mmol/ltris-hcl,200mmol/lnacl,ph7.4)平衡后上样2.5ml,3.5ml缓冲液洗脱,从上样开始收集2.5-6ml的洗脱液为脱盐后的酶液。纯化后的突变体酶经sds-page分析,可以达到胶纯的效果。结果如图3所示。m为标准蛋白分子量maker条带大小分别为(170kda、130kda、100kda、70kda、55kda、40kda、35kda、25kda、10kda),泳道1、pe-sumo-phhepia259d重组菌的超声破壁上清;泳道2、pe-sumo-phhepia259d表达产物的纯化;泳道3、pe-sumo-phhepis169d/a259d重组菌的超声破壁上清;泳道4、pe-sumo-phhepis169d/a259d表达产物的纯化。
3、纯化后的突变酶使用酶活测定方法进行测定,测定结果为:与野生型酶相比突变体酶phhepia259d和phhepis169d/a259d的比酶活分别提高了40.43%和122.05%。具有较大的工业应用潜力以及经济价值。
其中,突变酶的活性测定方法为:
酶活测定采用232nm的光吸收法,以肝素钠为底物测定肝素酶i的活性,一个酶活力单位(1iu)指在30℃,1min内产生1μmol的△4,5不饱和糖醛酸反应效力。反应体系为:1.5ml的ep管中加入100μl的底物缓冲液(50mmol/l醋酸钠,5mmol/l醋酸钙,5mmol/l肝素钠),金属浴中40℃恒温孵育10min,加入使用缓冲液(20mmol/ltris-hcl,200mmol/lnacl)稀释150倍的纯化脱盐后的酶液10μl,反应10min,立即加入0.06mol/l盐酸1ml终止反应。在12000r/min条件下离心5min,取上清液测定其在232nm的吸光值。
本发明的相关检测:
一、ph-hepi、phhepia259d和phhepis169d/a259d的酶学性质测定
突变型肝素酶i的活性同样也会受到温度、金属离子和ph等各种条件的影响,为了研究各因素对肝素酶i的活性影响,探究最佳酶活条件,进行了一系列实验。分别将未突变ph-hepi、突变型phhepia259d和phhepis169d/a259d在相同条件下的酶学性质进行分析比较。
1、温度对肝素酶i突变前后活性的影响
温度既能改变酶发生催化反应速度,也能导致酶蛋白活性降低或失活。本实验将同时测定未突变型ph-hepi和突变型phhepia259d和phhepis169d/a259d的最佳反应温度。
准备若干组100ul反应液(50mmol/l醋酸钠,5mmol/l醋酸钙,5mmol/l肝素钠,ph7.4),分别置于25、29、33、37、41、45℃下反应,添加酶量均为10ul,定时测定反应液a232变化情况,将25℃下测得的未突变的酶的酶活值作为100%,计算其他温度下的相对酶活。结果如图4所示,突变后的肝素酶i的最适反应温度与未突变型肝素酶i的一致均为37℃,说明突变前后并未改变其最适的反应温度。
2、ph对肝素酶i突变前后活性的影响
酶反应都有其最佳的ph值范围,ph值过高或者过低对酶发生催化反应的活性都会产生影响,本实验将同时测定未突变型ph-hepi和突变型phhepia259d和phhepis169d/a259d的最佳反应ph。
准备若干组100ul反应液(50mmol/l醋酸钠,5mmol/l醋酸钙,5mmol/l肝素钠),分别调节ph值为4.0、5.0、6.0、7.0、8.0、9.0、10.0,添加酶量均为10ul,置于37℃反应10min,测定反应液a232变化情况,将ph值为4条件下测得的未突变的酶活值作为100%,计算其他ph值下的相对酶活。结果如图5所示,未突变型ph-hepi和突变型phhepia259d和phhepis169d/a259d的最佳反应ph都是7.0。说明突变前后并未改变肝素酶i的最适ph值。
3、ca2+对肝素酶i突变前后活性的影响
由于肝素酶i结构中存在ca2+结合位点,ca2+对底物的结合和催化有着重要作用,因此ca2+浓度会极大地影响肝素酶i的活性,本实验将同时测定未突变型ph-hepi和突变型phhepia259d和phhepis169d/a259d的最佳反应ca2+浓度值。
准备若干组100ul反应液(50mmol/l醋酸钠,5mmol/l醋酸钙,5mmol/l肝素钠,ph7.4),分别控制ca2+浓度为0、5、10、50、100、200(单位:mm),添加酶量均为10ul,置于37℃条件下反应10min,测定反应液a232变化情况,将不添加ca2+浓度条件下测得的未突变酶的酶活值作为100%,计算其他浓度下的相对酶活。结果如图6所示,未突变型ph-hepi和突变型phhepia259d的最佳ca2+浓度是10mm,而突变型phhepis169d/a259d的最佳ca2+浓度是50mm。因为肝素酶i中存在ca2+的结合位点,这个ca2+的结合的结合区域包括169位的氨基酸残基,169位氨基酸由s突变为d之后,结合ca2+的能力增强,进而影响底物的结合和催化,提高比酶活。
二、ph-hepi、phhepia259d和phhepis169d/a259d的空间结构分析
通过同源建模,多序列比对以及定点突变,构建了两株比酶活显著提高的肝素酶i的突变体phhepia259d和phhepis169d/a259d。通过使用pymol软件对突变前后的肝素酶i的空间结构进行结构解析,分析出比酶活提高的可能存在的原因。突变体phhepia259d,将259位的a突变为d后比酶活得到显著的提高,因为259位残基涉及到肝素酶i和底物的结合,它位于酶活性口袋的上方,类似于一个盖子将底物更好地结合在活性区域,进而促进酶对底物的催化作用,提高比酶活,结构图如图7所示;突变体phhepis169d/a259d是一个双突变,将169位和259位进行复合突变,突变后酶活的增加得到了叠加效果,比单一突变效果更好。肝素酶i中存在钙离子结合位点,而169位残基正好处于这个结合区域,将169位的s突变为d后,在空间结构上显示钙离子结合区域增加一条氢键,氢键可以稳定蛋白的结构稳定,因此突变后使钙离子的结合更加稳定,进而影响了底物的结合以及催化,提高了比酶活,结构图如图8所示。
序列表
天津科技大学
一种提高肝素酶i活性的分子改造方法
seqidno:1
phhepia259d核苷酸序列
1
cagcagaaaaaatccggtaacatcccgtaccgtgttaacgttcaggctgattctgctaaacagaaagctatcatcgataacaaatgggttgctgttggtatcaacaaaccgtacgctttacagtatgatgataaactgcgcttcaacggtaaaccgtcctaccgcttcgaacttaaagctgaagataactcccttgaaggttacgctgctggtgaaactaaaggtcgtactgaattgtcctactcctacgctactactaacgatttcaaaaaattcccgccgtccgtttaccagaacgctcagaaacttaaaactgtttaccattacggtaaaggtatctgtgaacagggttcctcccgctcctacactttctccgtttacatcccgtcctccttcccggataacgctactactatcttcgctcagtggcacggtgctccgtcccgtactcttgttgctactccggaaggtgaaatcaaaactctgtccatcgaagaattcttggctttatacgatcgcatgatcttcaaaaaaaacatcgctcacgataaagttgaaaaaaaagataaagatggtaaaatcacttacgttgctggtaaaccgaacggttggaaagttgaacagggtggttacccgactctggctttcggtttctctaaaggttacttctacatcaaagctaactccgatcgtcagtggcttactgataaagatgatcgtaacaacgctaacccggaaaactccgaagttatgaaaccgtactcctccgaatacaaaacttccactatcgcttacaaaatgccgttcgctcagttcccgaaagattgctggatcactttcgatgttgctatcgattggactaaatacggtaaagaagctaacactatcttgaaaccgggtaaactggatgttatgatgacttacactaaaaacaaaaaaccgcagaaagctcacatcgttaaccagcaggaaatcctgatcggtcgtaacgatgatgatggttactacttcaaattcggtatctaccgtgttggtaactccactgttccggttacttacaacctgtctggctattccgaaactgctcgt
seqidno:2
phhepia259d氨基酸序列
2
qqkksgnipyrvnvqadsakqkaiidnkwvavginkpyalqyddklrfngkpsyrfelkaednslegyaagetkgrtelsysyattndfkkfppsvyqnaqklktvyhygkgiceqgssrsytfsvyipssfpdnattifaqwhgapsrtlvatpegeiktlsieeflalydrmifkkniahdkvekkdkdgkityvagkpngwkveqggyptlafgfskgyfyikansdrqwltdkddrnnanpensevmkpysseyktstiaykmpfaqfpkdcwitfdvaidwtkygkeantilkpgkldvmmtytknkkpqkahivnqqeiligrndddgyyfkfgiyrvgnstvpvtynlsgysetar
seqidno:3
phhepis169d/a259d核苷酸序列
3
cagcagaaaaaatccggtaacatcccgtaccgtgttaacgttcaggctgattctgctaaacagaaagctatcatcgataacaaatgggttgctgttggtatcaacaaaccgtacgctttacagtatgatgataaactgcgcttcaacggtaaaccgtcctaccgcttcgaacttaaagctgaagataactcccttgaaggttacgctgctggtgaaactaaaggtcgtactgaattgtcctactcctacgctactactaacgatttcaaaaaattcccgccgtccgtttaccagaacgctcagaaacttaaaactgtttaccattacggtaaaggtatctgtgaacagggttcctcccgctcctacactttctccgtttacatcccgtcctccttcccggataacgctactactatcttcgctcagtggcacggtgctccggatcgtactcttgttgctactccggaaggtgaaatcaaaactctgtccatcgaagaattcttggctttatacgatcgcatgatcttcaaaaaaaacatcgctcacgataaagttgaaaaaaaagataaagatggtaaaatcacttacgttgctggtaaaccgaacggttggaaagttgaacagggtggttacccgactctggctttcggtttctctaaaggttacttctacatcaaagctaactccgatcgtcagtggcttactgataaagatgatcgtaacaacgctaacccggaaaactccgaagttatgaaaccgtactcctccgaatacaaaacttccactatcgcttacaaaatgccgttcgctcagttcccgaaagattgctggatcactttcgatgttgctatcgattggactaaatacggtaaagaagctaacactatcttgaaaccgggtaaactggatgttatgatgacttacactaaaaacaaaaaaccgcagaaagctcacatcgttaaccagcaggaaatcctgatcggtcgtaacgatgatgatggttactacttcaaattcggtatctaccgtgttggtaactccactgttccggttacttacaacctgtctggctattccgaaactgctcgt
seqidno:4
phhepis169d/a259d氨基酸序列
4
qqkksgnipyrvnvqadsakqkaiidnkwvavginkpyalqyddklrfngkpsyrfelkaednslegyaagetkgrtelsysyattndfkkfppsvyqnaqklktvyhygkgiceqgssrsytfsvyipssfpdnattifaqwhgapdrtlvatpegeiktlsieeflalydrmifkkniahdkvekkdkdgkityvagkpngwkveqggyptlafgfskgyfyikansdrqwltdkddrnnanpensevmkpysseyktstiaykmpfaqfpkdcwitfdvaidwtkygkeantilkpgkldvmmtytknkkpqkahivnqqeiligrndddgyyfkfgiyrvgnstvpvtynlsgysetar
尽管为说明目的公开了本发明的实施例,但是本领域的技术人员可以理解:在不脱离本发明及所附权利要求的精神和范围内,各种替换、变化和修改都是可能的,因此,本发明的范围不局限于实施例所公开的内容。
序列表
<110>天津科技大学
<120>一种比酶活提高的肝素酶i的定向改造酶及分子改造方法和表达工程菌
<160>8
<170>siposequencelisting1.0
<210>1
<211>1089
<212>dna/rna
<213>定向改造酶phhepia259d核苷酸序列(unknown)
<400>1
cagcagaaaaaatccggtaacatcccgtaccgtgttaacgttcaggctgattctgctaaa60
cagaaagctatcatcgataacaaatgggttgctgttggtatcaacaaaccgtacgcttta120
cagtatgatgataaactgcgcttcaacggtaaaccgtcctaccgcttcgaacttaaagct180
gaagataactcccttgaaggttacgctgctggtgaaactaaaggtcgtactgaattgtcc240
tactcctacgctactactaacgatttcaaaaaattcccgccgtccgtttaccagaacgct300
cagaaacttaaaactgtttaccattacggtaaaggtatctgtgaacagggttcctcccgc360
tcctacactttctccgtttacatcccgtcctccttcccggataacgctactactatcttc420
gctcagtggcacggtgctccgtcccgtactcttgttgctactccggaaggtgaaatcaaa480
actctgtccatcgaagaattcttggctttatacgatcgcatgatcttcaaaaaaaacatc540
gctcacgataaagttgaaaaaaaagataaagatggtaaaatcacttacgttgctggtaaa600
ccgaacggttggaaagttgaacagggtggttacccgactctggctttcggtttctctaaa660
ggttacttctacatcaaagctaactccgatcgtcagtggcttactgataaagatgatcgt720
aacaacgctaacccggaaaactccgaagttatgaaaccgtactcctccgaatacaaaact780
tccactatcgcttacaaaatgccgttcgctcagttcccgaaagattgctggatcactttc840
gatgttgctatcgattggactaaatacggtaaagaagctaacactatcttgaaaccgggt900
aaactggatgttatgatgacttacactaaaaacaaaaaaccgcagaaagctcacatcgtt960
aaccagcaggaaatcctgatcggtcgtaacgatgatgatggttactacttcaaattcggt1020
atctaccgtgttggtaactccactgttccggttacttacaacctgtctggctattccgaa1080
actgctcgt1089
<210>2
<211>363
<212>prt
<213>定向改造酶phhepia259d编码的高活性肝素酶i完整的氨基酸序列(unknown)
<400>2
glnglnlyslysserglyasnileprotyrargvalasnvalglnala
151015
aspseralalysglnlysalaileileaspasnlystrpvalalaval
202530
glyileasnlysprotyralaleuglntyraspasplysleuargphe
354045
asnglylysprosertyrargphegluleulysalagluaspasnser
505560
leugluglytyralaalaglygluthrlysglyargthrgluleuser
65707580
tyrsertyralathrthrasnaspphelyslyspheproproserval
859095
tyrglnasnalaglnlysleulysthrvaltyrhistyrglylysgly
100105110
ilecysgluglnglyserserargsertyrthrpheservaltyrile
115120125
proserserpheproaspasnalathrthrilephealaglntrphis
130135140
glyalaproserargthrleuvalalathrprogluglygluilelys
145150155160
thrleuserilegluglupheleualaleutyraspargmetilephe
165170175
lyslysasnilealahisasplysvalglulyslysasplysaspgly
180185190
lysilethrtyrvalalaglylysproasnglytrplysvalglugln
195200205
glyglytyrprothrleualapheglypheserlysglytyrphetyr
210215220
ilelysalaasnseraspargglntrpleuthrasplysaspasparg
225230235240
asnasnalaasnprogluasnsergluvalmetlysprotyrserser
245250255
glutyrlysthrserthrilealatyrlysmetprophealaglnphe
260265270
prolysaspcystrpilethrpheaspvalalaileasptrpthrlys
275280285
tyrglylysglualaasnthrileleulysproglylysleuaspval
290295300
metmetthrtyrthrlysasnlyslysproglnlysalahisileval
305310315320
asnglnglngluileleuileglyargasnaspaspaspglytyrtyr
325330335
phelyspheglyiletyrargvalglyasnserthrvalprovalthr
340345350
tyrasnleuserglytyrsergluthralaarg
355360
<210>3
<211>1089
<212>dna/rna
<213>定向改造酶phhepis169d/a259d核苷酸序列(unknown)
<400>3
cagcagaaaaaatccggtaacatcccgtaccgtgttaacgttcaggctgattctgctaaa60
cagaaagctatcatcgataacaaatgggttgctgttggtatcaacaaaccgtacgcttta120
cagtatgatgataaactgcgcttcaacggtaaaccgtcctaccgcttcgaacttaaagct180
gaagataactcccttgaaggttacgctgctggtgaaactaaaggtcgtactgaattgtcc240
tactcctacgctactactaacgatttcaaaaaattcccgccgtccgtttaccagaacgct300
cagaaacttaaaactgtttaccattacggtaaaggtatctgtgaacagggttcctcccgc360
tcctacactttctccgtttacatcccgtcctccttcccggataacgctactactatcttc420
gctcagtggcacggtgctccggatcgtactcttgttgctactccggaaggtgaaatcaaa480
actctgtccatcgaagaattcttggctttatacgatcgcatgatcttcaaaaaaaacatc540
gctcacgataaagttgaaaaaaaagataaagatggtaaaatcacttacgttgctggtaaa600
ccgaacggttggaaagttgaacagggtggttacccgactctggctttcggtttctctaaa660
ggttacttctacatcaaagctaactccgatcgtcagtggcttactgataaagatgatcgt720
aacaacgctaacccggaaaactccgaagttatgaaaccgtactcctccgaatacaaaact780
tccactatcgcttacaaaatgccgttcgctcagttcccgaaagattgctggatcactttc840
gatgttgctatcgattggactaaatacggtaaagaagctaacactatcttgaaaccgggt900
aaactggatgttatgatgacttacactaaaaacaaaaaaccgcagaaagctcacatcgtt960
aaccagcaggaaatcctgatcggtcgtaacgatgatgatggttactacttcaaattcggt1020
atctaccgtgttggtaactccactgttccggttacttacaacctgtctggctattccgaa1080
actgctcgt1089
<210>4
<211>363
<212>prt
<213>phhepis169d/a259d编码的高活性肝素酶i完整的氨基酸序列(unknown)
<400>4
glnglnlyslysserglyasnileprotyrargvalasnvalglnala
151015
aspseralalysglnlysalaileileaspasnlystrpvalalaval
202530
glyileasnlysprotyralaleuglntyraspasplysleuargphe
354045
asnglylysprosertyrargphegluleulysalagluaspasnser
505560
leugluglytyralaalaglygluthrlysglyargthrgluleuser
65707580
tyrsertyralathrthrasnaspphelyslyspheproproserval
859095
tyrglnasnalaglnlysleulysthrvaltyrhistyrglylysgly
100105110
ilecysgluglnglyserserargsertyrthrpheservaltyrile
115120125
proserserpheproaspasnalathrthrilephealaglntrphis
130135140
glyalaproaspargthrleuvalalathrprogluglygluilelys
145150155160
thrleuserilegluglupheleualaleutyraspargmetilephe
165170175
lyslysasnilealahisasplysvalglulyslysasplysaspgly
180185190
lysilethrtyrvalalaglylysproasnglytrplysvalglugln
195200205
glyglytyrprothrleualapheglypheserlysglytyrphetyr
210215220
ilelysalaasnseraspargglntrpleuthrasplysaspasparg
225230235240
asnasnalaasnprogluasnsergluvalmetlysprotyrserser
245250255
glutyrlysthrserthrilealatyrlysmetprophealaglnphe
260265270
prolysaspcystrpilethrpheaspvalalaileasptrpthrlys
275280285
tyrglylysglualaasnthrileleulysproglylysleuaspval
290295300
metmetthrtyrthrlysasnlyslysproglnlysalahisileval
305310315320
asnglnglngluileleuileglyargasnaspaspaspglytyrtyr
325330335
phelyspheglyiletyrargvalglyasnserthrvalprovalthr
340345350
tyrasnleuserglytyrsergluthralaarg
355360
<210>5
<211>39
<212>dna/rna
<213>引物s169d-f(unknown)
<400>5
cagtggcacggtgctccggatcgtactcttgttgctact39
<210>6
<211>39
<212>dna/rna
<213>引物s169d-r(unknown)
<400>6
agtagcaacaagagtacgatccggagcaccgtgccactg39
<210>7
<211>35
<212>dna/rna
<213>引物a259d-f(unknown)
<400>7
gtggcttactgataaagatgatcgtaacaacgcta35
<210>8
<211>35
<212>dna/rna
<213>引物a259d-r(unknown)
<400>8
tagcgttgttacgatcatctttatcagtaagccca35
- 还没有人留言评论。精彩留言会获得点赞!