一种密罗木基因MfbHLH145及其应用的制作方法
本发明属于植物基因工程
技术领域:
,具体涉及一种密罗木基因mfbhlh145及其应用。
背景技术:
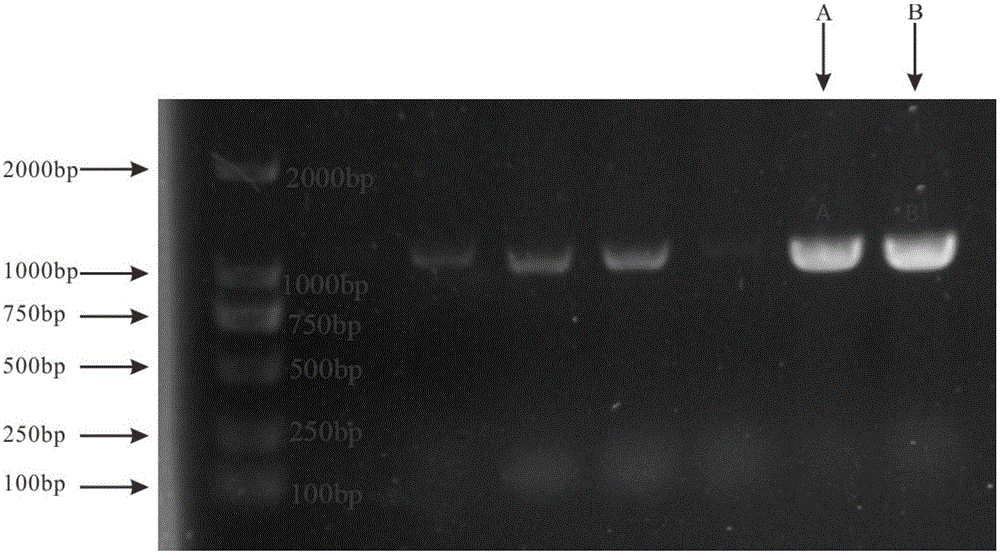
:干旱对植物生长发有着极其不利的影响。我国是一个受干旱影响严重的国家,干旱、半干旱面积约占国土面积的1/2。人口的增长,城镇建设的快速发展,不仅导致民用、工业、农业用水竞争日益激烈,也客观造成污染加剧,生态系统破坏,水资源短缺现象日益加剧。因此,发掘抗旱基因,培育抗旱、节水的植物新品种,对加快建设资源节约型、环境友好型社会,大力发展绿色经济,增强可持续发展能力,实现经济发展和生态文明的有机统一具有重要作用。基因工程技术是植物耐旱性改良的重要手段。目前,已有不少耐旱基因被报道和应用,其中转录因子基因通过与其调控的下游基因启动子区的顺式作用元件结合而调控靶基因的表达,以对外界不良环境做出响应。现今研究中认为高等植物中对染色体的分离、新陈代谢调节以及植物抗逆性相关的几大转录因子家族包括了wrky、myb、abf、b-zip以及bhlh等,而bhlh转录因子家族在真核植物中也已发现myb、myc、fbh等几个大类。其中,bhlh转录因子广泛存在于真核生物中,不仅在动物的细胞中出现,在高等植物中也有表达。其特殊的螺旋-环-螺旋结构域(basichelix-loop-helix,bhlh)是一段拥有近60个氨基酸的片段,因其上游富含碱性氨基酸而得名,其特殊之处在于除开hlh保守结构域之外,其序列中远端基序非常灵活,具有较高变异性。其n端与hlh基序相邻,包含6个共有氨基酸残基,属于dna结合域,而bhlh结构域的c端则包含hlh序列作为二聚体化功能域,它主要由含有两个亲水亲脂α螺旋的疏水残基组成的环形结构组成,两个螺旋之间被不同长度的环分开,从而形成三位一体结构域。bhlh的特殊结构使其不同转录因子或同源转录因子之间能够形成同源或异源二聚体,与靶基因启动子特异性结合,起到转录调控的作用。已有大量研究表明bhlh转录因子可受到多种逆境条件如干旱、盐、射线、低温、高温等能都诱导,并在植物对逆境的抗性反应发挥作用。密罗木(myrothamnusflabellifoliawelw.)主要分布在南非,是地球上为数不多的极耐旱复苏植物之一,也是唯一的木本复苏植物。它能够像种子一样耐受高度脱水干燥,在组织含水量仅存7%-11%时仍能保持遇水复活的特性。因此,密罗木是一种理想地研究植物抗旱性的材料。然而过去,研究人员主要通过形态学、生理、生化等方面研究了该植物的抗旱机理,但对于其中与脱水响应相关的基因及其耐旱分子机制还不清楚,也尚未见密罗木中抗旱bhlh转录因子及其应用的相关报道。技术实现要素:针对现有技术中的上述不足,本发明提供一种密罗木基因mfbhlh145及其应用,该基因能在干旱脱水期快速响应,提高作物的抗旱性能。为实现上述目的,本发明解决其技术问题所采用的技术方案是:一种密罗木基因mfbhlh145,该基因的编码序列如seqidno.1所示或seqidno.1所示的核苷酸序列经取代、缺失和/或添加一个或多个核苷酸,且能编码相同功能蛋白质的核苷酸序列。其中,序列表seqidno.1从密罗木叶片rna中通过rt-pcr的方法克隆,核心编码区长度为1020bp。采用上述基因编码的蛋白质,其氨基酸序列如seqidno.2所示或seqidno.2所示的氨基酸序列经取代、缺失和/或添加一个或多个氨基酸,且表达相同功能蛋白质的氨基酸序列。其中,seqidno.2由339个氨基酸组成,其中有一个位于241aa的核定位信号“pskkrkl”,对mfbhlh145氨基酸序列进行结构域分析,发现在196-209aa处含有一个低复杂度区域,hlh域位于284aa-333aa处,可以看出mfbhlh145具备hlh结构域,属于bhlh类转录因子。包含上述密罗木mfbhlh145基因的质粒。包含上述密罗木mfbhlh145基因的重组表达载体。包含上述密罗木mfbhlh145基因的转基因细胞系。包含上述密罗木mfbhlh145基因的工程菌。密罗木基因mfbhlh145在作物抗旱过程中的应用。本发明的有益效果为:1.获得了对干旱有较高耐受性的转基因拟南芥。2.密罗木基因mfbhlh145能在干旱脱水期快速响应,为利用该基因提高其他植物的干旱耐受性提供了理论依据及利用价值。附图说明图1为密罗木基因mfbhlh145的目的片段条带。图2为密罗木基因mfbhlh145的系统进化树;图3为密罗木基因mfbhlh145在不同脱水时期的表达量;图4为密罗木基因mfbhlh145亚细胞定位的结果;图5为转mfbhlh145基因植株的阳性鉴定结果;图6为野生型和转基因拟南芥植株在氯化钠胁迫处理后的生长状态;其中,图6a为野生型和转基因拟南芥植株在氯化钠胁迫处理后的植株幼苗生长状态;图6b为野生型和转基因拟南芥植株在氯化钠胁迫处理后成株的生长状态;图7为野生型和转基因拟南芥植株在干旱胁迫处理后的生长状态;其中,图7a为野生型和转基因拟南芥植株在干旱胁迫处理后的植株幼苗生长状态;图7b为野生型和转基因拟南芥植株在干旱胁迫处理后的成株生长状态;图8为野生型和转基因拟南芥植株失水率测定;图9为野生型和转基因拟南芥植株的脯氨酸含量测定;图10为野生型和转基因拟南芥植株的叶绿素测定;图11为野生型和转基因拟南芥植株的气孔测定;其中,图11a为野生型和转基因拟南芥植株的气孔形状检测图;图11b为野生型和转基因拟南芥植株的气孔数量条状图;图12为在干旱条件下,转mfbhlh145基因在拟南芥株系中相关基因的表达量;其中,图12a为nced3基因的表达量;图12b为p5cs基因的表达量;图12c为rab18基因的表达量;图12d为rd22基因的表达量;图12e为rd29a基因的表达量。图13为在盐胁迫条件下,转mfbhlh145基因在拟南芥株系中相关基因的表达量;其中,图13a为nced3基因的表达量;图13b为p5cs基因的表达量;图13c为rab18基因的表达量;图13d为rd22基因的表达量;图13e为rd29a基因的表达量。具体实施方式下面对本发明的具体实施方式进行描述,以便于本
技术领域:
的技术人员理解本发明,但应该清楚,本发明不限于具体实施方式的范围,对本
技术领域:
的普通技术人员来讲,只要各种变化在所附的权利要求限定和确定的本发明的精神和范围内,这些变化是显而易见的,一切利用本发明构思的发明创造均在保护之列。实施例1克隆密罗木mfbhlh145基因1、取密罗木叶片,液氮速冻,放置-80℃冰箱中保存以备提取总rna;总rna提取采用成都兰博生物公司购买的planttotalrnaisolationkit试剂盒来提取;密罗木cdna的合成使用大连宝生物科技公司的reversetranscriptasem–mlv(rnaseh-),按其产品说明书操作进行第一链合成。以上述试剂盒合成的cdna第一链为扩增模板,以设计的f:5’-catgccatgggaaaggactgtggatcc-3’(seqidno.3)和r:5’-gctagtgagagaatcaagtcctaaagctttg-3’(seqidno.4)为引物,为了后续的酶切及重组连接,在设计引物时加上ncoi和spei这两个酶切位点,利用rt-pcr进行cdna扩增,扩增体系见表1,扩增条件为:95℃预变性3min,95℃变性15s,58℃退火15s,72℃延伸1min,共35个循环,最后72℃延伸5min。表1pcr扩增体系试剂用量dna100~150ng引物f5pmol引物r5pmoltaqplus聚合酶1u10×pcr缓冲液2.5μlmg2+1.5mmddh2oupto25μl2、pcr结束后进行电泳分析,采用omega公司的dna回收试剂盒回收约1020bp的扩增片段(参考图1),将扩增片段连接到peasy-t1simple克隆载体,转化大肠杆菌dh5α感受态细胞,挑取白色菌落进行菌落pcr以鉴定阳性克隆,将阳性克隆送到成都擎科生物科技有限公司测定密罗木基因mfbhlh145的编码序列,其序如seqidno.1所示,再利用orffinder软件将所得基因的开放阅读框翻译成氨基酸序列,该氨基酸序列如seqidno.2所示。实施例2密罗木mfbhlh145的序列同源与同源性分析根据基因同源对比图谱可以看出,重合度较高的氨基酸并没有集中分布在预测的bhlh功能结构域内,且系统发育树中mfbhlh145单独分为一类,说明mfbhlh145基因特异性较高,对该类bhlh基因的研究信息较少。根据系统进化树可见(如图2),与mfbhlh145亲缘度较高的植物均为木本植物,且同属于被子植物门,双子叶植物纲。对同源度较高植物如可可(theobromacacao)等的生长环境分析发现,同源度较高植物大多与密罗木来源于相似的生长环境,这些植物偏好湿度不高,气温偏高的生长环境,且大多生长于亚热带、热带地区,符合生物进化过程中的规律。实施例3密罗木不同脱水时期mfbhlh145的表达水平采用trizol法分别提取密罗木0,10%,25%脱水阶段叶片总rna,以提取的总rna为模板,使用大连宝生物科技公司的reversetranscriptasem-mlv(rnaseh-)进行反转录并获取cdna。通过引物设计软件设计引物,退火温度设置58℃,actin基因作为内参,每个反应3个技术重复,实时荧光定量pcr仪进行荧光定量pcr,结果分析使用程序自带软件完成。所用引物序列如seqidno.5、seqidno.6、seqidno.7和seqidno.8所示,结果如图3所示。actin引物序列:f:5’-atcaccattggggctgaacg-3’(seqidno.5);r:5’-gtcgacccaccactaaggac-3’(seqidno.6)。mfbhlh145引物序列:f:5’-cccatcatcgccaggctattt-3’(seqidno.7)r:5’-caaaagaactcaggccctccat-3’(seqidno.8)实施例4mfbhlh145的亚细胞定位根据亚细胞载体phb-yfp上的碱基和酶切位点情况,以及clonexpressii同源重组试剂盒的说明,选择hindiii和bamhi这两个酶切位点来设计引物;引物由成都擎科生物公司合成。采用已合成的带有同源臂的引物,以密罗木cdna为模板进行扩增,其引物的具体序列如下:f:5’-accagtctctctctcaagcttatgggaaaggactgtggatcc-3’(seqidno.9)r:5’-gctcaccatactagtggatccgagagaatcaagtcctaaagctttg-3’(seqidno.10)将扩增得到的pcr产物回收后,与经过hindiii和bamhi内切酶切除的phb载体相连,构建融合载体35s:mfbhlh145-phb-yfp,通过将转入phb-yfp质粒的野生型本氏烟草和转入35s:mfbhlh145-phb-yfp质粒的野生型本氏烟草叶片制成切片,在激光共聚焦显微镜下观察结果如图4所示;从图4中可看出,只转入phb-yfp的烟草叶片的荧光信号分布在烟草细胞内的各个部位,而转入mfbhlh145-phb-yfp质粒的烟草叶片的荧光信号则只出现在细胞核中,说明mfbhlh145基因定位于细胞核中,并在细胞核中起作用。实施例5:mfbhlh145基因的遗传转化将农杆菌lba4404感受态细胞置于冰上解冻,取5μl重组质粒35s:pgsa1403-mfbhlh145于100μl感受态中,用枪头轻轻混匀,冰浴30min,液氮速冻1min,37℃水浴5min,冰上放置2min后加入体积为800μl的lb液体培养基,在恒温培养箱中29℃,120r/min,培养2~4h。将上述菌液均匀涂布在含10μg/ml氯霉素的lb固体培养基中,置于28℃恒温培养箱中培养,然后挑取单菌斑接种于体积为5ml的lb液体培养基中,28℃,250r/min振荡培养24h。取1ml上述菌液转入体积为50ml,氯霉素浓度为10μg/ml的lb液体培养基中,于28℃,250r/min,振荡培养至od600≈1.5;将上述od值达标的菌液4℃,8000r/min离心10min收集菌体,弃掉上清,用等体积的5%蔗糖溶液将菌体重悬,在菌液中加入silwet-77并混匀,使溶液浓度达到0.02%且菌液od600≈0.8。转化前需剪掉成熟的果荚,颠倒植株让植株浸入农杆菌细胞悬浮液2min,然后将浸好的植株用滤纸擦干并用塑料薄膜包裹,暗处理24h后于光照培养箱中正常培养,一周后可重复浸染一次。待种荚黄化后收种,记为t0代。继续将t0代拟南芥种子放入含有50μg/ml卡那霉素的培养基里,转入外源基因的植株能在含有卡那霉素的抗性培养基上生长,而非转基因植株则不能正常生长。因此筛选出能正常生长且颜色呈深绿色的植株,对深绿色植株提取dna并进行pcr检测阳性(如图5),获得t1代转基因植株,重复筛选步骤直至筛选出阳性过表达纯系植株。实施例6:转基因拟南芥的胁迫处理对于幼苗处理,是将野生型和过表达t3代纯系(line-e,line-f)分别播种于配置含nacl(100mm,150mm)、甘露醇(200mm,250mm,300mm)的1/2ms培养基中,对照组只添加正常成分,记为ck。每个方形培养皿中三种株系分成两排点播,每排15株左右,每个处理重复三次,4℃暗处理2d后移入光照培养箱中垂直放置,继续培养2周,在根系分析仪下拍照并测量根长。盐胁迫下的幼苗表型如图6所示,其中,图6a为野生型和转基因拟南芥植株幼苗在氯化钠胁迫处理后的植株生长状态;图6b为野生型和转基因拟南芥植株成株在氯化钠胁迫处理后的生长状态。干旱胁迫下的幼苗表型如图7所示,其中,图7a为野生型和转基因拟南芥植株幼苗在干旱胁迫处理后的生长状态;图7b为野生型和转基因拟南芥植株成株在干旱胁迫处理后的生长状态。对于成株的处理,是将野生型拟南芥(wt)和过表达t3代纯系(line-e,line-f)种子播种在1/2ms圆形培养皿,4℃暗处理3d后移入光照培养箱中培养至四叶期后移入花盆中,培养3周左右,选择长势一致的植株进行抗逆性分析。盐胁迫处理:对照组不作处理,记为ck,其余组每隔3天浇一次300mm的氯化钠处理液。2周后观察wt与过表达植株表型差异(如图6);每个株系10株重复。干旱胁迫处理:对照组(记为ck)正常浇水管理,其余组均不浇水使其自然干旱失水,期间不断观察拍照并记录植株生长状况,失水3周左右开始复水,处理期间拍照观察(如图7);每个株系10株重复。通过研究野生型与过表达拟南芥幼苗和成株在干旱及氯化钠胁迫下的表型,发现过表达mfbhlh145基因株系line-e和line-f对干旱及盐的耐受性均明显强于wt,说明过表达mfbhlh145基因提高了拟南芥对盐及干旱胁迫的抗性。实施例7:相关生理指标的测定(1)拟南芥的培养将野生型拟南芥(wt)和过表达t3代纯系(line-e,line-f)种子播撒在1/2ms圆形培养皿,4℃暗处理3d后转入光照培养箱中培养7天,移栽至50孔穴盘内(已等量分装好营养土并浇足水),继续培养4周左右。(2)失水率的处理及测定将穴盘中培养的wt,line-e,line-f浇足水分,剪去莲座叶对其取样,在天平上用定量滤纸称取三个株系各0.5g叶片,此时叶片含水量为初始含水量,记为m0,此后将叶片放入恒温光照培养箱中让其自然干旱(25℃,60%相对湿度),在0.5h、1h、1.5h、2h、3h、4h、5h、6h、7h分别称取叶片即时重量。结果如图8所示。(3)脯氨酸含量的测定在天平上准确称取25mg脯氨酸,倒入小烧杯内用少量蒸馏水溶解后倒入250ml容量瓶中,加蒸馏水定容至刻度,此标准液中每毫升含脯氨酸100μg;取6个50ml容量瓶,分别盛入脯氨酸原液0.5ml、1.0ml、1.5ml、2.0ml、2.5ml、3ml,用蒸馏水定容至刻度并混匀,各瓶的脯氨酸浓度分别为1μg/ml、2μg/ml、3μg/ml、4μg/ml、5μg/ml、6μg/ml。拟南芥的培养同步骤(1),4周后对wt,line-e,line-f三种植株进行自然干旱和浇灌300mm氯化钠溶液处理(自然干旱处理14d,氯化钠处理2d),随后取正常条件和经过干旱、氯化钠处理的wt,line-e,line-f植株叶片鲜样各0.5g,加入少许石英砂和少许3%的磺基水杨酸溶液在研钵中磨碎,倒入10ml离心管并定容至10ml,4500r离心3min。取上清液并用3%的磺基水杨酸溶液定容至10ml。吸取2ml提取液至一洁净干燥玻璃试管中,加入2ml酸性茚三酮和2ml冰乙酸,100℃加热1h,放入冰水中终止反应,加入4ml甲苯,充分振荡15~20s,静置分层后用移液枪轻轻吸取上层红色溶液于比色皿中,以甲苯为对照,在分光光度计520nm波长处比色并求得吸光度值,此步骤重复两次,结果如图9所示。(4)盐胁迫下叶绿素含量的测定拟南芥的培养同步骤(1),4周后对wt,line-e,line-f三种植株浇灌200mm氯化钠溶液处理2天,随后取正常生长和经氯化钠处理的wt,line-e,line-f植株叶片鲜样各0.5g,将其叶中脉剪掉并剪碎叶片,加入到50ml的棕色容量瓶中,分别加入上述95%乙醇提取液50ml,将容量瓶加塞放入25℃的恒温培养箱中暗中提取48h。因为95%乙醇提取液在波长为649nm和665nm处具有最大吸光值a649和a665,所以用紫外分光光度计测量a649和a665,以95%乙醇提取液作为空白对照,此步骤重复两次;结果如图10所示。实施例8:甘露醇处理下气孔开度的测定将野生型拟南芥(wt)和过表达t3代纯系(line-e,line-f)种子播撒在1/2ms圆形培养皿,4℃暗处理3d后转入光照培养箱中培养7天,移栽至50孔穴盘内(已等量分装好营养土并浇足水),继续培养4周左右。取wt,line-e,line-f的莲座叶叶片,每个株系撕取10块下表皮,置于100ml的mes-kcl缓冲液(50mmkcl,0.1mmcacl2,10mmmes,ph6.15)中光诱导2.5h,然后每个株系取5块下表皮分别置于100ml不含以及含300mm甘露醇的mes-kcl缓冲液(50mmkcl,0.1mmcacl2,10mmmes,ph6.15)中光下处理2h,处理完后放在荧光显微镜(10*40)下观察,气孔闭合程度(长与宽的比)作为统计气孔开度的指标,每个表皮条随机选取4个视野,每个视野随机测量10个气孔,总共统计100个气孔细胞,结果如图11所示,其中,图11a为野生型和转基因拟南芥植株的气孔形状和气孔闭合程度;图11b为野生型和转基因拟南芥植株的气孔长宽比柱状图。实施例9:mfbhlh145转基因拟南芥中nced3、rd22、p5cs5、rd29a、rab18胁迫相关基因表达量测定提前一天将培养4周的野生型拟南芥(wt)和过表达t3代纯系(line-e,line-f)的穴盘里浇足水分,此时对wt和过表达株系的叶片进行取样,记为ck,干旱处理按照自然干旱处理12h,24h,36h,48h共5个时间点对三个株系的叶片取样;氯化钠处理按照浇足200mm的氯化钠后的12h、24h、30h、36h共5个时间点对三个株系的叶片取样。分别提取各转基因株系拟南芥总rna,利用反转录试剂盒将rna反转录成cdna,利用primer5设计内参actin以及nced3、rd22、p5cs5、rd29a、rab18基因的引物,具体引物序列如下:actin:f:5’-ggaaggatctgtacggtaac-3’;(seqidno.11)r:5’-tgtgaacgattcctggacct-3’;(seqidno.12)nced3:f:5’-cgagccgtggcctaaagtct-3’;(seqidno.13)r:5’-gctccgatgaatgtaccgtgaa-3’;(seqidno.14)rd22:f:5’-acttggtaaatatcacgtcagggct-3’;(seqidno.15)r:5’-ctgaggtgttcttgtggcatacc-3’;(seqidno.16)p5cs5:f:5’-ggtggaccaagggcaagtaagata-3’;(seqidno.17)r:5’-tcggaaaccatctgagaatcttgt-3’;(seqidno.18)rd29a:f:5’-ggcggagaaactgacaactgaa-3’(seqidno.19)r:5’-ccaccaaaccagccagatgat-3’(seqidno.20)rab18:f:5’-gcagtatgacgagtacggaaatcc-3’;(seqidno.21)r:5’-ccttgtccatcatccgagctaga-3’。(seqidno.22)然后进行实时荧光定量pcr检测,其干旱条件下检测结果见图12;其中,图12a为nced3基因的表达量;图12b为p5cs基因的表达量;图12c为rab18基因的表达量;图12d为rd22基因的表达量;图12e为rd29a基因的表达量。根据数据图表显示,在自然干旱处理条件下,atnced3在wt及过表达株系中,在没有进行干旱处理时,linee及wt表达量相当,而linef表达量则明显较低,经过干旱处理后,在过表达mfbhlh145株系linee中,于36h达到峰值,linef中在96h时达到峰值。且在linee中,12h时表达量显著高于wt及linef,在24h时表达量较wt略低,36h时过表达株系均显著高于wt,在48h时linee及linef中表达量依然高于wt,但在72h时linee的表达量明显较低,随着干旱处理进行到96h时,过表达株系中表达量明显高于wt。过表达mfbhlh145株系linee及linef中atp5cs5的表达量在进行干旱处理之前linee表达量较wt及linef更高,且在48h时表达量达到峰值,而linef则在整个干旱过程中表达量较linee及wt低,其atp5cs5的表达量在干旱处理72h时达到峰值。linee中atp5cs5的表达量明显较高,且随着干旱程度加深,其表达量有明确升高的趋势。过表达mfbhlh145株系linee及linef中atrab18的表达量本底水平较wt更高,在12h时linef中表达量有所下降,但依然高于wt,而linee则相较0h时有所升高,linee中表达量在12h时达到峰值,在24h时维持较高水平,而在干旱处理开始之后linef的表达量则较未处理时有显著下降,但依然高于wt,且在12h时达到峰值。在整个干旱处理过程中,wt中的atrab18的表达量随着时间推移而升高,而过表达株系中的atrab18则于12-48h时明显高于wt,在72-96h时表达量较wt低。atrd22在过表达株系linee及linef中表达量始终高于wt,其本底表达量显著高于wt。随着时间的推移wt中atrd22的表达量缓慢升高,而linee一直保持较高表达水平,在36h时出现低值,随后表达量随着时间的推移降低,在24h时达到峰值。linef在48h时达到最高表达量,且同样一直保持高于wt的表达水平。atrd29a在过表达株系中本底水平显著高于wt,且在12h时linee中及linef中表达量达到峰值。wt中,atrd29a的表达量在36h时达到峰值,且高于linee,随后其表达量逐渐下降,在72h-96h时表达量显著高于过表达株系linee及linef。在linee中,12h后表达量随着时间的推移下降,且在36h时明显低于wt,而linef中则在48h时表达量明显高于wt及linee,在36h后表达量逐渐下降。其盐胁迫条件下检测结果见图13;其中,图13a为nced3基因的表达量;图13b为p5cs基因的表达量;图13c为rab18基因的表达量;图13d为rd22基因的表达量;图13e为rd29a基因的表达量。在盐胁迫条件下,atnced3在未进行氯化钠处理时,过表达株系linee中的表达量显著高于wt及linef,linef的本底表达量则低于wt及linee。氯化钠处理进行到12h时,过表达株系linee及linef中的表达量提高,且在12h时达到峰值,随着时间的推移在48h时,过表达株系linee及linef中的atnced3表达量达到做最低值,随后在72h时,linee中atnced3表达量突然大量累积。atp5cs5在过表达株系linee及linef中的本底表达水平较wt高,但随着氯化钠处理的进行,linee中表达量站在12h及24h处理时表达量有所下降,在36-72h时表达量显著高于wt及linef,且于72h时达到峰值。在linef中表达量在处理时间点中均低于wt,且于12h时达到最低值。wt中atp5cs5的表达量呈现稳定升高趋势,于72h时达到峰值。atrab18在过表达株系linee及linef中,本底表达水平高于wt,在氯化钠处理进行到12-24h时表达量降低,且低于wt,在24h后表达量有所升高,与36h时达到峰值,其后linef中表达量持续降低,而linee中表达量则在72h时有所升高,且高于wt。而wt中atrab18的表达量于12h时达到峰值,随后缓慢降低,在24-72h中表达量稳定偏低。atrd22在过表达株系中的本地表达水平显著高于wt,在经过氯化钠处理后,linee及linef的表达水平有相对下降趋势,并在24-72h时呈现较为稳定的表达量,atrd22在过表达株系中表达水平始终较为稳定,与wt中atrd22表达量相当。过表达株系linef则随着氯化钠处理时间的推移,其表达量开始稳步升高,于72h时达到峰值,且显著高于wt。在wt中,atrd22表达量没有明显的波动形势,其表达量较为稳定。atrd29a的表达量在过表达株系linee及linef中本底表达水平高于wt。随着氯化钠处理加深,linee中atrd29a的表达量在72h时达到峰值,且显著高于wt及linef,而linef中atrd29a的表达量在36h时达到峰值,且显著高于wt及linee。在wt中atrd29a的表达量则始终处于较低水平,在36h处理时达到峰值。序列表<110>四川农业大学<120>一种密罗木基因mfbhlh145及其应用<160>22<170>siposequencelisting1.0<210>1<211>1020<212>dna<213>密罗木(myrothamnusflabellifoliawelw)<400>1atgggaaaggactgtggatcctggtttccccagcggcattattgtgagcaatcacccgat60ctggataggttgtgtcctccccttcatttgggacagcaaaatatgatcccatcctacatg120aacccttactctaattcggtttctacatatgggattttgccaatgttacaatctcctgag180tctctccatttgacggcaaaccaagcggatgaacctcatggatggttttattgtttgccc240catcatcgccaggctatttcccctggatcgaagtcagttatcaaagatgaagttccttct300agcctttataaagcagatacagatcagaggaggttcttagtttttgatcaatctggcgat360caaacaaccttcatgttcagtccagctattaggacttccatggagggcctgagttctttt420ggtccaagtccttataatggtcataactttagtggggaaaggccaggtactgaaagagac480tcgcttcacaatgctggtcctattctaacagatggaaacgaaattaatgggactgatttg540gaaagtgagatgcatgaagacacagaagaacttaatgccttgctgtactcagatgacgag600aatgactactctgaggatgacgaagaaactagcaccggtcattcgcctagtacaatgaca660tctaacgaaagattgaaaagctttgaaggaagtgctgaagaagttgctagcactgctgag720ccaagtaaaaagcgaaaactatcactcatggacaccgcaacttctaggaaatcaatccag780ttcttgggttatgaagaagatgacgcagaatctagctgtggtaatggccttagtcaaggt840aatggaggggagggcagtaaaaggctgagaagggagagaattcgtgcaactgtgagcatt900ctacagagcataattcctaatggaatgggaaaggatgcagttgcggttcttgatgaagct960attaactactttagatctttgaagctcaaagccaaagctttaggacttgattctctctaa1020<210>2<211>339<212>prt<213>密罗木(myrothamnusflabellifolia)<400>2metglylysaspcysglysertrppheproglnarghistyrcysglu151015glnserproaspleuaspargleucysproproleuhisleuglygln202530glnasnmetileprosertyrmetasnprotyrserasnservalser354045thrtyrglyileleuprometleuglnserprogluserleuhisleu505560thralaasnglnalaaspgluprohisglytrpphetyrcysleupro65707580hishisargglnalaileserproglyserlysservalilelysasp859095gluvalproserserleutyrlysalaaspthraspglnargargphe100105110leuvalpheaspglnserglyaspglnthrthrphemetpheserpro115120125alaileargthrsermetgluglyleuserserpheglyproserpro130135140tyrasnglyhisasnpheserglygluargproglythrgluargasp145150155160serleuhisasnalaglyproileleuthraspglyasngluileasn165170175glythraspleugluserglumethisgluaspthrglugluleuasn180185190alaleuleutyrseraspaspgluasnasptyrsergluaspaspglu195200205gluthrserthrglyhisserproserthrmetthrserasngluarg210215220leulysserphegluglyseralaglugluvalalaserthralaglu225230235240proserlyslysarglysleuserleumetaspthralathrserarg245250255lysserileglnpheleuglytyrglugluaspaspalagluserser260265270cysglyasnglyleuserglnglyasnglyglygluglyserlysarg275280285leuargarggluargileargalathrvalserileleuglnserile290295300ileproasnglymetglylysaspalavalalavalleuaspgluala305310315320ileasntyrpheargserleulysleulysalalysalaleuglyleu325330335aspserleu<210>3<211>27<212>dna<213>人工序列(artificialsequence)<400>3catgccatgggaaaggactgtggatcc27<210>4<211>30<212>dna<213>人工序列(artificialsequence)<400>4gctagtgagagaatcaagtcctaaagctttg31<210>5<211>20<212>dna<213>人工序列(artificialsequence)<400>5atcaccattggggctgaacg20<210>6<211>20<212>dna<213>人工序列(artificialsequence)<400>6gtcgacccaccactaaggac20<210>7<211>21<212>dna<213>人工序列(artificialsequence)<400>7cccatcatcgccaggctattt21<210>8<211>22<212>dna<213>人工序列(artificialsequence)<400>8caaaagaactcaggccctccat22<210>9<211>42<212>dna<213>人工序列(artificialsequence)<400>9accagtctctctctcaagcttatgggaaaggactgtggatcc42<210>10<211>46<212>dna<213>人工序列(artificialsequence)<400>10gctcaccatactagtggatccgagagaatcaagtcctaaagctttg46<210>11<211>20<212>dna<213>人工序列(artificialsequence)<400>11ggaaggatctgtacggtaac20<210>12<211>20<212>dna<213>人工序列(artificialsequence)<400>12tgtgaacgattcctggacct20<210>13<211>20<212>dna<213>人工序列(artificialsequence)<400>13cgagccgtggcctaaagtct20<210>14<211>22<212>dna<213>人工序列(artificialsequence)<400>14gctccgatgaatgtaccgtgaa22<210>15<211>25<212>dna<213>人工序列(artificialsequence)<400>15acttggtaaatatcacgtcagggct25<210>16<211>23<212>dna<213>人工序列(artificialsequence)<400>16ctgaggtgttcttgtggcatacc23<210>17<211>24<212>dna<213>人工序列(artificialsequence)<400>17ggtggaccaagggcaagtaagata24<210>18<211>24<212>dna<213>人工序列(artificialsequence)<400>18tcggaaaccatctgagaatcttgt24<210>19<211>23<212>dna<213>人工序列(artificialsequence)<400>19ggcggagaaactgacaactgaa22<210>20<211>23<212>dna<213>人工序列(artificialsequence)<400>20ccaccaaaccagccagatgat21<210>21<211>24<212>dna<213>人工序列(artificialsequence)<400>21gcagtatgacgagtacggaaatcc24<210>22<211>23<212>dna<213>人工序列(artificialsequence)<400>22ccttgtccatcatccgagctaga23当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1