SlNAC29基因在提高番茄细菌性叶斑病抗性和耐旱性中的应用的制作方法
本发明涉及生物技术领域,尤其涉及slnac29基因在提高番茄细菌性叶斑病抗性和耐旱性中的应用。
背景技术:
近年来,我国蔬菜播种面积逐年上涨,蔬菜已成为市场化程度最大的经济作物。据2017年中国统计年鉴数据显示,在农作物种植结构中蔬菜的种植比例由2000年的9.7%上升到2016年的13.4%,居各类经济作物之首(http://www.stats.gov.cn/tjsj/ndsj/2017/indexch.htm)。
其中,番茄是栽培最为广泛的蔬菜作物之一,由于番茄及其制品的独特食用、保健及辅助治疗的功效,在世界农业生产中占有重要的地位。但是,温室效应导致全球气候变暖,干旱发生频率高、持续时间长,影响范围广、后续影响大,成为影响我国农业生产最严重的气象灾害,各种细菌性、病毒性以及真菌性病害发生十分猖獗,干旱和病害对农业产量及品质造成了极大的威胁。
细菌性叶斑病是由丁香假单胞杆菌引起的,给农业生产造成了巨大的经济损失。番茄细菌性叶斑病在辽宁、福建、内蒙古、新疆、广西、河北、甘肃、吉林、天津等省市区均有发生。该病可造成5%-75%的产量损失。干旱导致植物光合电子传递链过度还原,产生过量活性氧,损坏植物细胞膜系统,影响番茄正常生长发育,产量和品质下降,因此,增强番茄的抗病性以及耐旱性对番茄生产具有重要的现实意义。
目前,在我国使用化学农药防治病虫害是最为常见的防治手段。大量的化学农药投入到农业生态体系中严重破坏了农田生态系统,打破了农业生态平衡,影响了生物多样性,不利于农业的可持续发展。大量化学农药施用也频频引发食品安全问题,对人们的身体健康造成威胁。因此,寻找出新的、有效的、经济无害的方法,有效提高番茄应对不良环境的能力,是目前蔬菜生产中亟待解决的难题之一,也是当今蔬菜生产和科研工作中的研究热点,培育抗性品种具有重要的实际生产意义。
培育抗性品种,有两种常用方法,一种是将作物与具有抗性的亲本进行杂交及多代回交等过程进行筛选,一般需要花费较长时间,且需要大量人力物力;另一种方式是转基因育种,该方式最为高效快速,且可以实现跨物种的基因发掘,拓宽遗传资源的利用范围,实现已知功能基因的定向高效转移。这种对基因进行精确定向操作的育种方式效率更高,针对性更强,在缓解资源约束,保障粮食安全,保护生态环境,拓展农业功能等方面有巨大潜力。利用这种方法,可以导入一个广谱抗病调控基因,或者一种植物中可同时导入多个抗病调控基因,因此具有培育广谱、持久抗病品种等优点,或者导入抗旱基因,培育耐旱品种。转基因育种,需要对基因功能有所了解。
nac转录因子为植物六大转录家族成员之一,为植物所特有,nac家族成员众多,在番茄中有74个。有研究显示,nac基因家族中的很多成员在植物对病菌侵染或抗病相关信号分子处理后表现出差别表达特征,反映了nac基因家族在植物抗病反应中具有一定的生物学功能。有报道表明nac家族成员在防御某些植物病害中起正调控作用。例如,水稻osnac6基因表达受机械损伤和稻瘟病所诱导,过量表达osnac6基因可以激活过氧化物酶基因的表达,进而提高水稻对稻瘟病的抗性(nakashimak等,“functionalanalysisofanac-typetranscriptionfactorosnac6involvedinabioticandbioticstress-responsivegeneexpressioninrice.”theplantjournal,(2007)51,617–630);也有nac家族成员能通过增强穿透阻力而阻止病菌进入。大麦基因hvnac6在白粉病菌侵染早期,表达集中在表皮细胞内;在表皮细胞内沉默hvnac6,表皮细胞对白粉病菌的耐穿透力降低,而互补试验表明在表皮细胞瞬时过量表达hvnac6基因后能增强大麦表皮细胞对白粉病菌的耐穿透力(m.k.jensen等,thehvnac6transcriptionfactor:apositiveregulatorofpenetrationresistanceinbarleyandarabidopsis。plantmol.biol.,65(2007),pp.137-150)。然而,也有nac转录因子在抗病进程中起负调控作用,番茄slnac1能与番茄曲叶病毒(tomatoleafcurlvirus,tlcv)复制增强蛋白结合,过量表达slnac1后增加了tlcvdna的积累(selthla等,“anacdomainproteininteractswithtomatoleafcurlvirusreplicationaccessoryproteinandenhancesviralreplication.”,plantcell,2005,17(1):311–325)。
近年来的研究越来越显示出nac转录因子在转基因育种中的应用潜力,如过量表达snac1、snac2、onac10的转基因水稻植株抗盐能力增强,过量表达osnac06的转基因水稻能增强对稻瘟病的抗病性,过量表达hvnac6的转基因大麦能够增强对白粉病的抗性。因此,通过转基因技术改变nac基因表达水平或nac蛋白活性有可能获得抗病抗逆转基因作物新品种或新材料。
然而,对于番茄slnac29基因的研究却并未有报道。研究slnac29基因有利于深入解析番茄对细菌性叶斑病的抗性机制,以及用于解析番茄抗旱机制,可用于抗病抗旱番茄的培育,对研发抗性品种具有一定的理论指导意义;对于提高番茄应对不良环境的能力,培育抗性品种也具有重要一定的生产意义。
技术实现要素:
本发明提供了slnac29基因在提高番茄细菌性叶斑病抗性和抗旱性中的新用途,为培育抗细菌性叶斑病和耐旱的番茄品种提供依据。
具体技术方案如下:
本发明提供了slnac29基因在提高番茄细菌性叶斑病中的应用,其中,所述slnac29基因的核苷酸序列如seqidno.1所示。
该slnac29基因位于番茄的第五染色体上,其开放阅读框(orf)长849bp;slnac29基因编码一种转录因子,该转录因子由282个氨基酸组成,其序列如seqidno.2所示。
本发明通过转基因技术,构建了slnac29基因过表达载体,并利用组培技术,筛选获得了slnac29过表达纯合株系oe:slnac29-1,oe:slnac29-2;然后,通过接种番茄细菌性叶斑病菌后,统计发病率,并进行台盼蓝染色,以检验细胞是否死亡,证明了过表达的slnac29基因能够提高植物对细菌性叶斑病的抗性。同时,对过表达植株进行干旱处理,拍摄植株耐旱性表型,证明了过表达的slnac29基因能提高植物对干旱的耐受性。
此外,本发明还通过凝胶迁移实验(emsa),发现slnac29基因编码蛋白通过与碳酸酐酶基因βca3的启动子上的序列为cacg的特异位点结合,进而调控碳酸酐酶基因的转录;而在前人的研究中已发现,βca3基因过表达能增强番茄植株对番茄细菌性叶斑病的防御能力。
所以,进一步地,本发明提供了slnac29基因编码的转录因子在提高番茄细菌性叶斑病抗性中的应用。
进一步地,所述转录因子通过与碳酸酐酶基因的启动子特异性结合,调控βca3基因的表达,进而提高番茄细菌性叶斑病抗性。
进一步地,所述slnac29基因编码的转录因子通过增强碳酸酐酶基因βca3的转录表达,进而提高对番茄细菌性叶斑病的防御能力。
进一步地,所述碳酸酐酶基因βca3启动子的核苷酸序列如seqidno.3所示。
本发明还提供了slnac29基因在提高番茄耐旱性中的应用,所述slnac29基因的核苷酸序列如seqidno.1所示。
本发明还提供了一种培育抗细菌性叶斑病和耐旱的番茄种质的方法,该方法包括以下步骤:
(1)制备slnac29基因的过表达载体;所述slnac29基因的核苷酸序列如seqidno.1所示;
(2)将所述过表达载体转入农杆菌感受态细胞中,构建含过表达载体的基因工程菌;
(3)将所述基因工程菌介导转化番茄子叶,培育得到转基因纯合株系。
进一步地,在过表达载体的制备过程中,采用的上游引物如seqidno.4所示,下游引物如seqidno.5所示。
进一步地,所述农杆菌为根癌农杆菌gv3101。
与现有技术相比,本发明具有以下有益效果:
(1)本发明经过系列试验发现了slnac29基因在提高番茄细菌性叶斑病抗性和耐旱性中的新用途,为培育抗细菌性叶斑病和耐旱的番茄品种提供了重要的基因资源。
(2)本发明利用转基因技术提供了一种抗细菌性叶斑病和耐旱的番茄种质的培育方法,得到了抗细菌性叶斑病和耐旱性强的转基因纯合植株。
附图说明
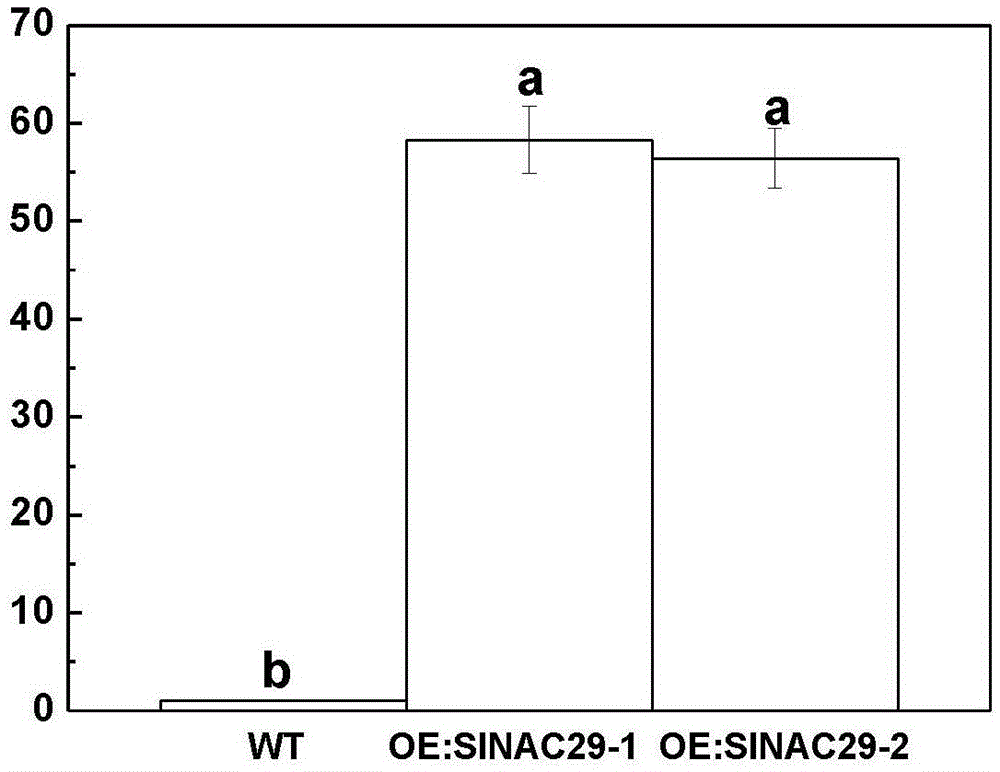
图1为slnac29基因在过表达植株中的表达模式分析;
其中,wt为未转基因的野生型番茄,oe:slnac29-1,oe:slnac29-2为筛选出的过表达纯合株系;小写字母a,b代表不同处理间在5%水平上的差异显著。
图2为番茄slnac29过表达植株接种细菌性叶斑病病原菌之后的病情指数;
其中,wt为未转基因的野生型番茄;oe:slnac29-1,oe:slnac29-2为筛选出的过表达纯合株系;小写字母a,b代表不同处理间在5%水平上的差异显著。
图3为番茄slnac29过表达植株接种细菌性叶斑病病原菌之后的台盼蓝染色图;
其中,wt为未转基因的野生型番茄,oe:slnac29-1,oe:slnac29-2为筛选出的过表达纯合株系。
图4为番茄slnac29过表达植株干旱胁迫下的表型;
其中,wt为未转基因的野生型番茄,oe:slnac29-1,oe:slnac29-2为筛选出的过表达纯合株系。
图5为slnac29蛋白结合在βca3基因启动子上的检测示意图,以及slnac29蛋白结合βca3基因启动子的凝胶电泳图。
其中,s1代表含有nac结合位点元件cacg的启动子片段,hotprobe表示生物素标记的含cacg元件的探针,mutantcoldprobe是生物素标记的含cacg突变的探针。
具体实施方式
下面结合具体实施例对本发明作进一步描述,以下列举的仅是本发明的具体实施例,但本发明的保护范围不仅限于此。
下述实施例中所用实验方法如无特殊说明,均为常规方法。
下述实施例中所用实验材料、试剂等,如无特殊说明,均可从商业途径得到。
实施例1番茄slnac29基因的克隆及载体构建
1.番茄总rna提取
采用tiangenplanttotalrnaextractionkit提取番茄幼嫩叶片的总rna,其步骤为:
(1)取0.1g叶片在液氮中磨碎,加1ml裂解液,后用匀浆仪(规格型号:上海净信jx-fstprp-48)处理;
(2)将匀浆样品在15-30℃放置5min,使得核酸蛋白复合物完全分离;
(3)4℃,12000rpm离心5min,去上清,转入一个新的无rnase的离心管中;
(4)加入200μl氯仿,盖好管盖,剧烈震荡15s,室温放置3min;
(5)4℃,12000rpm离心10min,样品会分为三层:黄色的有机相,中间层和无色的水相,rna主要在水相中,水相的体积约为所用裂解液rz试剂的50%。把水相转移到新管中,进行下一步操作;
(6)缓慢加入0.5倍体积的无水乙醇,混合均匀;将得到的溶液和沉淀一起转入吸附柱cr3,4℃,12000rpm离心30s,弃掉收集管中的废液;
(7)向吸附柱cr3中加入500μl去蛋白液rd,4℃,12000rpm离心30s,弃废液;
(8)向吸附柱cr3中加入600μl漂洗液rw,室温静止2min,4℃,12000rpm离心30s,弃废液;
(9)重复操作步骤(8);
(10)将吸附柱放入2ml收集管中,4℃,12000rpm离心2min,去除残余废液;
(11)将吸附柱cr3转入一个新的离心管中,加入50μlrnase-freeddh2o,室温放置2min,4℃,12000rpm离心2min;
(12)用紫外分光光度计测定od260/od280检验rna样品含量与纯度,浓度为1000ng/μl,od260/od280=2.12。
2.基因克隆
采用toyoborevertraaceqpcrrtkit,将1μg番茄总rna反转录成cdna。根据slnac29基因的编码序列,设计扩增出完整框的引物,前引物slnac29oef为ccgctcgagatggttggaaaaattagctc(seqidno.4),后引物slnac29oer为tcccccgggctgagattgaaatgttgg(seqidno.5),并在引物上分别加上限制性酶切位点(xho1和sma1);对slnac29基因进行扩增,然后酶切,分别连接到pfgc1008-ha得到过表达载体。由北京擎科生物科技有限公司测序,测序结果如seqidno.1所示,所编码的蛋白序列如seqidno.2所示,结果表明,所克隆的序列与pgsbplantgenomeandsystemsbiology(http://pgsb.helmholtz-muenchen.de/plant/tomato/reportsjsp/sequencereportcds.jsp?elementid=300776)中公布的序列(solyc05g007770.2)一致。
3.根癌农杆菌工程菌构建
将过表达载体转入根癌农杆菌gv3101中,获得含番茄slnac29基因过表达载体的根癌农杆菌工程菌。
实施例2构建番茄slnac29基因的过表达植株
1.培养无菌苗
番茄种子(solanumlycopersicummill.cv.condinered)用摇床于28℃,200rpm摇6-8h,然后用75%酒精消毒30sec,弃去酒精后在10%naclo中消毒15min(用摇床28℃,200rpm),灭菌蒸馏水冲洗3次并转移到灭菌器皿,接种于1/2ms培养基。25℃黑暗条件下培养至发芽,转入光照培养室,幼苗生长条件为25℃、16h光照/8h黑暗,光照强度1800lx。
2.准备外植体、培养农杆菌
种子发芽后6d,将无菌苗的子叶用刀切成长度约2-3cm的大小,置入看护培养基kcms中预培养1d(避光,过夜)。在含有氯霉素(34μg/ml)、庆大霉素(50μg/ml)、利福平(50μg/ml)的抗生素的lb平板上挑取slnac29基因过表达载体的农杆菌单菌落,置于2ml含有相同抗生素的lb中,28℃、200rpm摇24h后,再加入至20ml含有相同抗生素的lb中扩摇,28℃、200rpm过夜培养至对数中期(od600≈1.0,约16-24h)。
3.转化再生
3.1侵染
将上述培养完成的根癌农杆菌工程菌菌液转到10ml离心管(封口膜封口),4℃,4000rpm离心10min;倒出培养基,加入悬浮培养基侵染液ms0.2,摇匀。将3-4皿的子叶转移到含有ms0.2培养基的培养皿中,倒入悬浮好的菌液至od600≈0.2-0.3(7-8mlms0.2,其中2-3ml倒入培养皿)暗下接种感染4-5min,并轻轻晃动培养皿。将外植体转移到灭菌滤纸上,吸干残留的菌液,回接到kcms上,22℃共培养2d(避光)。反面朝上。
3.2选择培养、再生
共培养后,将外植体小心的转入mrs1(2z+)上,2-3周后转mrs2(0.2z)中培养,选择培养的过程中要及时清理污染的材料,直至长出再生芽。
4.再生芽生根、移栽
待再生芽长至1cm左右时,将芽切下,放入生根培养基中生根。2周后对生根良好、长至5cm左右的转化苗进行炼苗,移栽至一次性塑料杯中,成活后移栽至花盆中,得到番茄slnac29基因过表达植株。
经筛选得到基因表达相对较高的过表达纯合株系oe:slnac29-1,oe:slnac29-2。过表达株系基因表达情况由图1可知,qrt-pcr结果表明,slnac29过表达植株两株系表达量上升约60倍。所用引物为nac29-rt-f:ccaaatagagcagctgtgtca(seqidno.6),nac29-rt-r:ttggtggtttgcctttgtaa(seqidno.7)。
实施例3细菌性叶斑病病原菌处理
对实施例2制备得到的番茄slnac29过表达植株进行接种细菌性叶斑病病原菌处理。
细菌性叶斑病病原菌接种于含25mg/l利福平的固体king’sb培养基(蛋白胨10g,k2hpo41.5g,甘油15ml,琼脂15g,h2o1l,ph7.0)中,于28℃培养箱中培养2天后得以活化,转板培养一天,用10mmmgcl2溶液重悬菌体,并调节细菌浓度至od600=0.1,加0.02%的有机硅,对番茄植株叶背进行喷施,使得细菌菌液能够浸润叶片,细菌悬浮液通过气孔进入细胞间隙。
3天后观察叶片发病情况,根据图2、3,可得知:相比野生型,两个过表达株系无论是病情指数还是台盼蓝染色发病病况明显减轻,表明slnac29基因能提高番茄对细菌性叶斑病的抗性。
实施例4干旱胁迫处理
对实施例2制备得到的番茄slnac29过表达植株进行干旱胁迫处理。
番茄slnac29基因过表达植株干旱胁迫处理时均以未转基因的野生型番茄作为对照,干旱胁迫处理的具体方法如下:
一组为正常水分供应对照组(包含wt对照和过表达植株),一组为干旱处理实验组(包含wt对照植株和过表达植株)。
当番茄长至五叶一心时,实验组和对照组同时浇水至饱和,之后对照组植株正常浇水,直至实验结束;实验组之后每隔1天用zigwsn记录仪测量一次wt对照植株和过表达植株相对土壤含水量(vwc),并根据失水量补充少量水分使wt对照植株与过表达植株之间的vwc保持一致,直至处理结束。实验组整个干旱胁迫过程中控制相对土壤含水量从100%缓慢降低至30%左右,干旱胁迫时间为13天。
实验结束后,进行拍照。结果如图4所示。
图4为番茄slnac29过表达植株干旱胁迫下的表型图。由图4可知,干旱处理后,野生型植株出现严重萎蔫,而两个过表达株系仅下部叶片黄化萎蔫,耐旱性明显高于wt植株,表明slnac29基因能提高番茄对干旱的耐性。
实施例5slnac29与βca3基因启动子的结合验证
具体内容如下:
构建原核表达载体,设计扩增出完整框的引物,前引物slnac29-32a-f为cgagctcatggttggaaaaattagctc(seqidno.8),后引物slnac29-32a-r为cccaagcttctgagattgaaatgttgg(seqidno.9),并在引物上分别加上限制性酶切位点sac1和hand3,利用pcr克隆slnac29基因的全长序列,连接所用原核表达载体为pet32a,抗性为氨苄青霉素,重组质粒构建成功后,42℃,15s热击转化bl21(de3)菌株。
蛋白的诱导采用异丙基硫代半乳糖苷(iptg)进行。菌液一般诱导体系为300ml,用含100mg/ml氨苄青霉素抗生素的lb液体培养基37℃摇菌,至od0.6~0.8,加入0.5m母液浓度的iptg60μl,16℃诱导过夜。4000g,4℃离心10min,收集菌体。
利用ni-nta亲和树脂来纯化重组蛋白(novagen),方法参照说明书。
分析βca3启动子发现在其atg上游920bp左右处有一个nac结合位点元件cacg元件,将含有该元件的βca3启动子片段设计引物。
野生型探针前引物:hotprobe-f(seqidno.10):tggtaggatacacacgaaaccttccaataattgatttca;
野生型探针后引物:hotprobe-r(seqidno.11):tgaaatcaattattggaaggtttcgtgtgtatcctacca;
野生型突变探针前引物:mutantcoldprobe-f(seqidno.12):gtggtaggatacattttaaaccttccaataattgatttca;
野生型突变探针后引物:mutantcoldprobe-r(seqidno.13):tgaaatcaattattggaaggtttaaaatgtatcctacca。
按照biotin3’enddnalabelingkit(pierce,89818)的说明对探针进行生物素末端标记,并且用以下程序将其退火为dna双链探针。具体退火程序为:95℃,5min,然后进行40个循环将温度从95℃降至55℃(每个循环降1℃,一个循环持续1分钟),最后4℃持续5min。
依据lightshiftchemiluminescentemsakit(thermofisherscientific,20148)说明书,利用生物素标记的探针进行slnac29-dna复合体的emsa实验。0.5μg的slnac29融合蛋白与生物素标记的探针,同时在20μl的反应体系中室温孵育20min,反应体系包括10mmtris-hcl,10mmdtt,50ng/μlpoly(di-dc),2.5%glycerol,0.05%nonidetp-40。利用6%的聚丙烯酰胺凝胶在100v下进行电泳分离,100v转膜,312nm紫外光下交联15min。用blockingbuffer封闭孵育15min,将66.7μl的链霉亲和辣根过氧化物酶共轭体稀释到blockingbuffer中(1:300),孵育15min。转皿,用10ml1×洗液洗膜。共5次,每次5min。转皿,加入15ml的底物平衡液,孵育5min。准备底物工作液,2ml鲁米诺增强因子,2ml双氧水等体积混合,化学发光仪检测。
经化学发光后,检测到slnac29蛋白与探针的结合带,而且slnac29蛋白与cacg突变的探针不能结合(图5);由此,用体外实验证明了slnac29与βca3基因启动子上的cacg元件特异的结合。
序列表
<110>浙江大学
<120>slnac29基因在提高番茄细菌性叶斑病抗性和耐旱性中的应用
<160>13
<170>siposequencelisting1.0
<210>1
<211>849
<212>dna
<213>番茄(solanumlycopersicuml.)
<400>1
atggttggaaaaattagctctgatcttcctcctggatttaggtttcatccaacagatgag60
gaattaatcatgtactatcttcgatatcaagctacttcaaggccatgccctgtctcaatt120
atccccgaaattgatgtctacaagttcgatccctgggtattacctgaaaaggcagaattt180
ggagacaatgagtggtatttttttacccctagagatagaaaatatccaaatggtgttagg240
ccaaatagagcagctgtgtcaggttattggaaggctactggcactgataaagcaatttat300
agtgcaaataaatatgttggaattaaaaaagcacttgttttttacaaaggcaaaccacca360
aaaggtgtcaagactgattggatcatgcatgaatatcgtttgagtgattcaaagtctcaa420
acatctaagcaaagtggatctatgaggttagatgattgggtactatgtagaatttataag480
aagaaaaatttggggagaactatagagatgatgaaagttgaagaagaagaattagaggca540
caaaatgtgagtactacaaataatgaaattgaagttgttggtggaccacaaacaatgaaa600
ttaccaagaatttgttcattgtcacatctattggaattggattattttggatcaattcca660
caattactaagtgacaatttattatatgatgaccaaggttacacaatgaataatgtgaat720
aacacaagtaatgtggatcaagtatcatcacaacaacaaaatacaaataatattacaagc780
aataattgtaatattttcttcaattatcaacaaccactttttgtgaatccaacatttcaa840
tctcagtga849
<210>2
<211>282
<212>prt
<213>番茄(solanumlycopersicuml.)
<400>2
metvalglylysileserseraspleuproproglypheargphehis
151015
prothraspglugluleuilemettyrtyrleuargtyrglnalathr
202530
serargprocysprovalserileileprogluileaspvaltyrlys
354045
pheaspprotrpvalleuproglulysalaglupheglyaspasnglu
505560
trptyrphephethrproargasparglystyrproasnglyvalarg
65707580
proasnargalaalavalserglytyrtrplysalathrglythrasp
859095
lysalailetyrseralaasnlystyrvalglyilelyslysalaleu
100105110
valphetyrlysglylysproprolysglyvallysthrasptrpile
115120125
methisglutyrargleuseraspserlysserglnthrserlysgln
130135140
serglysermetargleuaspasptrpvalleucysargiletyrlys
145150155160
lyslysasnleuglyargthrileglumetmetlysvalglugluglu
165170175
gluleuglualaglnasnvalserthrthrasnasngluilegluval
180185190
valglyglyproglnthrmetlysleuproargilecysserleuser
195200205
hisleuleugluleuasptyrpheglyserileproglnleuleuser
210215220
aspasnleuleutyraspaspglnglytyrthrmetasnasnvalasn
225230235240
asnthrserasnvalaspglnvalserserglnglnglnasnthrasn
245250255
asnilethrserasnasncysasnilephepheasntyrglnglnpro
260265270
leuphevalasnprothrpheglnsergln
275280
<210>3
<211>2000
<212>dna
<213>番茄(solanumlycopersicuml.)
<400>3
tagaatattgtatgatgtatataattctaatataaatgtgtcaagtgatcaagaatttga60
aagttctaggagtcgacttaataaagataatattgatgattatttagtttcttatcttga120
actttctcacaataatagaaatgattttgatgcatcctactgacgatgttctagcatggt180
gaagaaatcgcggcaaagaatttccaaaatttcaaccgatggctcgaaatgtattagcta240
ttcaagcatcttcagtagcttcaaaagatgtttttagtgcagcaaagtttcaaattggat300
atcatagcattcactactagcagcagatagtttggagatatcggtattatttagagattg360
gattaatgtcgagagaagaaatttaggtcgtgaaccactgtcgatcaaatttcaaagtga420
cgttgatgaaataatgcaagattatagtgacgacgacattgaggcaatggagaatttatc480
tattcaaccaattcccgatcatgttactagagaaatgttaaatgatttaagtaaagatct540
atatagtagcatcgactattaaatttaaatgtttaattgtctataagtgtgagtatgata600
attcgataactaattcaaacggagccttccaagaaagtggttgtatatattatatctttt660
aatactctactaatatttttgtagatcaaactgtactatcaaattaatattaataaaagt720
ctaggccattcgtcttaattttttttattattgtattaaatttaatttttattaatgtga780
aatttaaaattttatagtttttaaattttgaaatttaaacttttacaacctcaaagtttc840
aattttgaaattttaaacttttaaagtttgaatttaaattttcaaacttttaaaatttaa900
aatttaaattttcaaacttttaaagttcaaatattaaattagaaatataaaatttcaaat960
ttatattcaaagtttcaaactaatatttaaattatatatttataatttgaaaaattgaat1020
tttataaattttatatttataattcgaaagattattttacatcgtggtaggatacacacg1080
aaaccttccaataattgatttcattgaattgaacaagataattcaaataaaacaaatcgg1140
tcaaagaaattcatttttttgtgataaacgttacatgtcgataatttgacaaagatggca1200
ctgtattgccttataatacatatttaaattgactaccacttttttatgtatacttttatc1260
aatatatagttttattggaggtggcagaagcatagttaggtagggtttataggtttagac1320
gaatctattaacttttcatagattttgtatttatattaaaaattaattagatatatatat1380
atatagagagagagacttttaaccgaataaattgttttagaatccccaaactcctaatct1440
taactcctctgatcggtgggacttgacataattaaggtagttaattgaattaaactttac1500
tttattataaaatttgagtaggattttcttaaaaaatgattatgttttttcaaacctgac1560
atatattttcttcaaaacatgttaaattcacttatactaattagactttattttaattct1620
cattcaatttctaatacgtatattagagtctgaatattttggactcgttccgaataaggc1680
ccattctattatgaattttttcatatccaagactcgaatttaaaatttctaattaagaaa1740
agaataacctcatccattgcgccaccagatgaacaaacagataaagatcaaagctctagc1800
taggtaacaatacattgaattatttaccttgaaatcattttcaacactattgcgctattt1860
ggctttatatataaaataaaaggtaacatacctaaaacgtgatacagagattaaatatga1920
taactcatttattagctaatcgagtagtataaatagaggctaaagaaacccaaatttctc1980
caaacaatttcaaattaatt2000
<210>4
<211>29
<212>dna
<213>人工序列(artificialsequence)
<400>4
ccgctcgagatggttggaaaaattagctc29
<210>5
<211>27
<212>dna
<213>人工序列(artificialsequence)
<400>5
tcccccgggctgagattgaaatgttgg27
<210>6
<211>21
<212>dna
<213>人工序列(artificialsequence)
<400>6
ccaaatagagcagctgtgtca21
<210>7
<211>20
<212>dna
<213>人工序列(artificialsequence)
<400>7
ttggtggtttgcctttgtaa20
<210>8
<211>27
<212>dna
<213>人工序列(artificialsequence)
<400>8
cgagctcatggttggaaaaattagctc27
<210>9
<211>27
<212>dna
<213>人工序列(artificialsequence)
<400>9
cccaagcttctgagattgaaatgttgg27
<210>10
<211>39
<212>dna
<213>人工序列(artificialsequence)
<400>10
tggtaggatacacacgaaaccttccaataattgatttca39
<210>11
<211>39
<212>dna
<213>人工序列(artificialsequence)
<400>11
tgaaatcaattattggaaggtttcgtgtgtatcctacca39
<210>12
<211>40
<212>dna
<213>人工序列(artificialsequence)
<400>12
gtggtaggatacattttaaaccttccaataattgatttca40
<210>13
<211>39
<212>dna
<213>人工序列(artificialsequence)
<400>13
tgaaatcaattattggaaggtttaaaatgtatcctacca39
- 还没有人留言评论。精彩留言会获得点赞!