大麦耐湿调控基因HvERF2.11、蛋白及其在育种中的应用的制作方法
本发明属于基因工程技术领域,尤其涉及一个大麦耐湿调控基因hverf2.11及其育种应用。
背景技术:
湿害(waterlogging)是指土壤水分过多造成的缺氧环境,对作物正常生长发育所产生的危害。湿害在世界各地均有发生,尤其在北美、澳大利亚、中国、印度等国较为严重。随着全球生态环境的恶化,极端天气出现的频率不断增加,旱涝灾害发生频繁,湿害已经成为影响农作物生产的主要非生物胁迫因素之一。据统计,在世界范围内,大约有16%耕地均受到不同程度的湿涝危害,湿害往往导致作物减产约20%-30%,严重的湿害甚至导致绝收。我国是世界上受湿涝影响较为严重的国家之一,每年大约有30%的耕地面积受到不同程度洪涝影响,且呈逐年增加的趋势。仅在2013年,我国因为洪涝造成农田受灾面积达1.19×107hm2,造成了3000亿元以上的直接经济损失。
植物在长期进化过程中形成了多个基因协同表达调控模式,以适应逆境胁迫。近年来,对植物响应逆境胁迫研究已从单个功能基因的克隆及功能鉴定转向对转录因子的鉴定及调控功能研究。一个与逆境胁迫相关的转录因子,可同时调控下游一系列相关功能基因的表达变化。因此,转录因子对作物抗逆性改良的效果可能比单个功能基因更好。水稻、拟南芥、猕猴桃等植物中克隆的耐湿调控基因均属于转录因子erf-viis亚族基因,目前大麦中作为转录因子的耐湿调控基因尚未见报道。
技术实现要素:
本发明的目的是提供一个大麦耐湿基因hverf2.11及其育种应用,克隆并构建hverf2.11基因表达载体,利用转基因技术将hverf2.11基因导入拟南芥,对转基因拟南芥进行湿害胁迫处理,确认hverf2.11具有耐湿活性,为大麦耐湿品种改良提供基因资源及理论依据。
通过生物信息学分析,我们在大麦中鉴定到54个属于erf基因家族;系统进化树分析发现hverf2.11与拟南芥中低氧胁迫相关基因hre1、hre2、rap2.12及rap2.2进化关系最近。通过qrt-pcr技术分析大麦不同器官、不同条件下hverf2.11基因表达水平,克隆了耐湿大麦tf58中hverf2.11基因,并将hverf2.11基因转入拟南芥,验证了该基因的功能,以期为大麦品种耐性分子标记辅助选择育种奠定基础。
为了实现上述发明目的,本发明所采用的技术方案为:一种大麦耐湿调控相关蛋白,所述的蛋白为如下(a)或(b):
(a)由seqidno:2所示的氨基酸序列组成的蛋白质;具体序列为mcggailagfippsaaaaaakaaatakkkqqqrsvtadslwtglrkkadeedfeadfrdferdsseeeddeveevppppapatagfafaaaaevalraparrdaavqhdgpaakqvkrvrknqyrgirqrpwgkwaaeirdpskgvrvwlgtydtaeeaaraydaearkirgkkakvnfpedaptvqkstlkptaaksaklapppkacedqpfnhlsrgdndlfamfafsdkkvpakptdsvdsllpvkhlapteafgmnmlsdqssnsfgstdfgwddeamtpdytsvfvpsaaampaygepaylqggapkrmrnnfgvavlpqgngaqdipafdnevkyslpyvesssdgsmdnlllngamqdgassgdlwsldelfmaaggy;
(b)将seqidno:2的氨基酸序列经过一个或几个氨基酸残基的取代和/或缺失和/或添加且与大麦耐湿调控相关的由seqidno:2衍生的蛋白质。
本发明还提供编码前述蛋白的基因。所述基因是如下(a1)-(a3)中任何一种的dna分子;
(a1)seqidno:1所示的dna分子;
(a2)在严格条件下与(a1)限定的dna序列杂交且编码大麦耐湿调控相关蛋白的dna分子;
(a3)与(a1)限定的dna序列至少具有70%、至少具有75%、至少具有80%、至少具有85%、至少具有90%、至少具有95%、至少具有96%、至少具有97%、至少具有98%或者至少具有99%同源性且编码大麦耐湿调控相关蛋白的dna分子。
本发明还提供含有前述基因的表达盒、重组载体、重组微生物或转基因细胞系。
本发明的另一个目的是提供(b1)或(b2)或(b3)的应用:
(b1)所述蛋白,或,所述基因,或,含有所述基因的表达盒、重组载体、重组微生物或转基因细胞系,在抗大麦湿害胁迫中的应用;
(b2)所述蛋白,或,所述基因,或,含有所述基因的表达盒、重组载体、重组微生物或转基因细胞系,在培育大麦、拟南芥耐湿新品种中的应用;
(b3)所述蛋白,或,所述基因,或,含有所述基因的表达盒、重组载体、重组微生物或转基因细胞系,在抗拟南芥湿害胁迫中的应用。
本发明还提供一种培育植物耐湿品种的方法,将所述的基因转入目的植物,或所述的表达盒、重组载体转化目的植物,得到转基因植物;所述转基因植物的湿害胁迫抗性高于所述目的植物。所述目的植物为大麦、拟南芥。所述的转基因植物表达所述的大麦耐湿相关蛋白。
hverf2.11基因所述dna片段如序列表seqidno.1所示,或者基本相当于seqidno.1所示的dna序列,或者其功能相当于seqidno.1所示序列的部分片段。对该基因进行序列分析,表明hverf2.11基因编码区全长为1158bp、编码385个氨基酸,分子量为41.38kda、等电点为5.14,含有保守的ap2结构域。超表达序列表seqidno.1所示序列可以增强拟南芥和大麦等植物对湿害的耐性。
上述基因可以应用于改良大麦的耐湿能力,具体操作如下:
(1)基因的获得:通过生物信息学分析,获得大麦中hverf2.11基因的全长基因序列,提取耐湿品系tf58根系总rna,利用rt-pcr扩增到hverf2.11的基因序列,将扩增产物连接到pgem-teasy载体上,经测序获得具有hverf2.11基因的克隆。
(2)植物表达载体构建与遗传转化:利用invitrogen公司的gateway技术构建大麦hverf2.11基因超表达载体。利用bpclonase™enzyme,将hverf2.11基因与pdonr221载体连接,构建入门载体,然后利用lrclonase™enzyme,将入门载体与pb2gw7载体连接,构建含有hverf2.11基因的超表达载体。通过液氮冻融法将含有hverf2.11基因的超表达载体导入农杆菌gv3101中。利用农杆菌介导的遗传转化法,将hverf2.11编码序列导入拟南芥中表达。通过抗生素筛选、rt-pcr等筛选阳性转基因拟南芥植株。
(3)转基因植株耐湿性分析:将正常生长35天的转基因和野生型拟南芥株系进行淹水处理。淹水处理2周后,观察湿害胁迫下转基因植株与野生型植株的表型差异,从而证明hverf2.11基因对改良拟南芥耐湿性的能力。
本发明为增强植物对湿害的抗性提供了一种新的方法,通过基因工程手段改良作物的耐湿性,不仅克服传统育种周期短,而且操作简单、容易获得耐湿性材料。本发明首次克隆得到了大麦耐湿调控基因hverf2.11,并通过农杆菌介导的方法将该基因转入拟南芥中,经过湿害鉴定分析,证明转基因植株较野生型植株的耐湿能力有明显提高,证实本发明的基因在提高大麦耐湿性方面具有广泛的应用前景。
附图说明
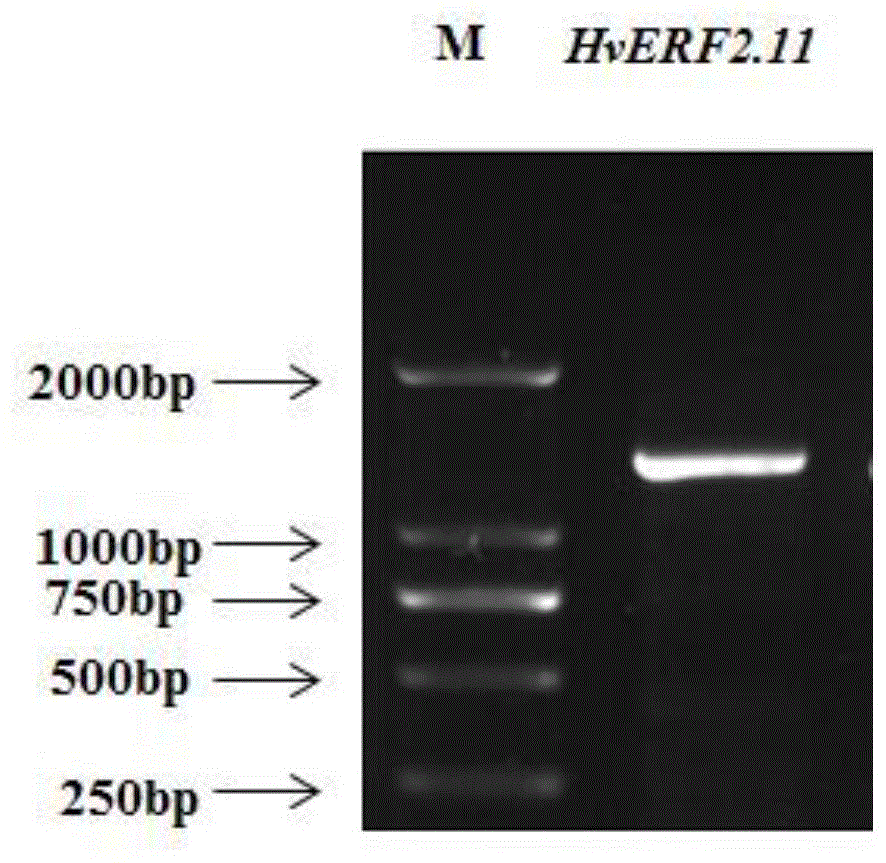
图1大麦hverf2.11基因序列rt-pcr扩增产物,
marker:dl2000dnamarker(大连宝生物),由2000bp、1000bp、750bp、500bp、250bp以及100bp六条dna片段组成;
图2大麦hverf2.11基因在大麦不同器官及拟南芥中的表达差异,
a:湿害胁迫后,大麦hverf2.11基因在tf58不同器官中的表达差异,l:叶片;ar:不定根;sr:种子根;nr:根节;s:穗子。b:hverf2.11基因在不同转基因拟南芥株系与野生型株系中表达差异;
图3淹水处理2周及对照条件下转基因拟南芥株系与野生型株系的耐湿性表型差异,(a)植株生长表型;(b)株高;(c)茎叶鲜重;(d)茎叶干重;(e)根系长度;(f)存活率;
图4不同淹水时期转基因株系与野生型株系叶片生理指标的变化,
a-e分别sod,cat,pod,adh酶活性及脯氨酸含量,数据表示平均值±标准差,试验3次生物学重复。*和**分别代表转基因株系与野生型间的差异达到显著和极显著水平(*p<0.05;**p<0.01);
图5不同湿害胁迫时期转基因拟南芥与野生型耐湿相关基因的表达差异,
a-f分别代表atsod1基因,atcat1基因,atpod1基因,atadh1基因,atpdc1及ataco1基因,拟南芥的atactin基因作为不同基因表达的内参基因。数值由3个重复的平均值±标准差来表示。*和**分别代表转基因株系与野生型间的差异达到显著和极显著水平。
具体实施方式
下面结合具体实施例对本发明作进一步详细说明。
实施例1hverf2.11基因的克隆
设计大麦hverf2.11基因的特异引物p1正向引物:5'-ccagtcagagatggtcaagg-3'(seqidno.3)和p2反向引物:5'-catccatcgtcttgctgag-3'(seqidno.4),采用ctab法提取耐湿材料tf58根系总rna,反转录合成cdna,以反转录cdna为模板,利用引物p1和p2扩增出如seqidno:1所示的大麦耐湿基因的cds序列,基因hverf2.11的cds序列全长1158bp(图1)。
具体步骤如下:
(1)向离心管中加入ctab(十六烷基三甲基溴化铵)提取缓冲液[2%(w/v)ctab,nacl1.4mol/l,edta(乙二胺四乙酸)20mmol/l,tris·hcl100mmol/l,2%(w/v)pvp]和10%β-巯基乙醇,在水浴锅中预热;
(2)将大麦根系液氮冷却研磨,加入提取液中,混匀,65℃水浴10分钟;
(3)加入等体积的氯仿:异戊醇(体积比24:1)混合液,颠倒混匀,静置10min,4℃12000g离心10min;
(4)取上清,重复步骤(3);
(5)取上清,加入终浓度为2mol/l的licl,冰浴10-12小时,11000rpm,4℃离心15min,弃上清,用75%乙醇清洗沉淀两次,溶于适量的depc(焦碳酸二乙酯)处理水中待用;
(6)从大麦品系tf58中提取根系总rna为模板,利用反转录酶(购自thermofisherscientific公司)将其反转录合成cdna第一条链,反应条件为:65℃5min,42℃50min,70℃10min;
(7)利用上述引物p1和p2从rna反转录得到的cdna中扩增出大麦乙烯转录因子基因hverf2.11的cds序列;
反应条件:94℃预变性4min;94℃30sec,55℃30sec,72℃1min,33个循环;72℃延伸10min。将扩增获得的pcr产物连入pmd18-t载体(购自宝生物工程大连有限公司),转化大肠杆菌感受态细胞,筛选阳性克隆并测序,获得所需的全长基因。从阳性克隆中提取携带有hverf2.11基因cds序列的质粒。
实施例2hverf2.11基因过表达载体的构建与遗传转化
为了能更好地分析基因hverf2.11的生物学功能,进一步将hverf2.11基因在拟南芥中实现过表达,利用invitrogen公司的gateway技术构建大麦hverf2.11基因超表达载体。利用bpclonase™enzyme,将hverf2.11基因与pdonr221载体连接,构建入门载体,然后利用lrclonase™enzyme,将入门载体与pb2gw7载体连接,构建含有hverf2.11基因的超表达载体。通过液氮冻融法将含有hverf2.11基因的超表达载体导入农杆菌gv3101中。利用农杆菌介导的遗传转化法,将hverf2.11编码序列导入拟南芥中。通过抗生素筛选和rt-pcr等筛选阳性转hverf2.11基因植株。
本试验实用的遗传转化方法如下:
(1)农杆菌的培养
首先,在带有对应抗性选择的固体lb培养基(10g/l蛋白胨、5g/l酵母提取物、10g/l氯化钠、kan100mg/l、琼脂1.5g/l)上预培养携带有hverf2.11基因的农杆菌48小时,培养温度28℃;挑取预培养农杆菌单菌落,接种于对应抗性选择的液体lb培养基(10g/l蛋白胨、5g/l酵母提取物、10g/l氯化钠、kan100mg/l)中,于28℃200rpm摇床培养过夜,至菌液浓度od600值约为0.8~1.0。
(2)花序法侵染拟南芥
采用改良的花序侵染法转化拟南芥。挑取含有hverf2.11基因质粒的农杆菌单菌落,转入100mllb液体培养基中,置入28℃摇床中200r/min过夜培养,使农杆菌菌液浓度达到od600在0.8~1.0时,4000r/min离心10min,弃上清后,用100ml含有蔗糖5g、silwetl-7750μl的lb液体培养基重新悬浮农杆菌菌液。将拟南芥花盆倒扣于含有菌悬液的烧杯中,将拟南芥的花序浸在菌液中10~20s,黑暗下培养24h后,置于温度25℃、湿度为60%、光照强度3000~4000lx和16h/8h光周期的光照培养箱中培养,隔天浇水和培养植株,待开花后收获种子。将收获的拟南芥种子用8%naclo酒精溶液(95%酒精配制)中消毒5min后,在选择培养基(ms+50mg/l卡那霉素+7g/l琼脂+30g/l蔗糖)上进行筛选。
实施例3hverf2.11基因转基因t3代苗rt-pcr检测
为了验证转基因拟南芥t3代株系耐湿能力的改变是否与转入的hverf2.11基因有关,采用rt-pcr方法对部分转基因拟南芥植株中hverf2.11基因表达进行了检测,结果见图2,可知转基因拟南芥t3代株系耐湿能力的改变与转入的hverf2.11基因的过表达有关。
具体步骤如下:
采用trizol试剂(购自宝生物工程大连有限公司)从转基因拟南芥t3代3个株系中提取植株的总rna(提取方法参照trizol试剂说明书操作),利用反转录酶(购自thermofisherscientific公司)将其反转录合成cdna第一条链,反应条件为65℃5min,42℃50min,70℃10min。先用报道的内参基因actin对反转录得到的cdna进行检测和浓度调整,进行pcr检测,确保内参基因均能在对照野生型拟南芥和转基因拟南芥植株中均能扩出。然后,根据hverf2.11基因的序列,利用引物p1和p2,进行rt-pcr检测,反应条件为:94℃预变性4min;94℃30sec,55℃30sec,72℃1.5min,33个循环;72℃延伸10min。扩增产物的琼脂糖凝胶电泳结果表明(图2),3个转基因株系内均检测到hverf2.11基因的表达。
实施例4转hverf2.11拟南芥植株耐湿性的鉴定
为了能进一步分析大麦耐湿基因hverf2.11的生物学功能,将该基因在拟南芥中实现了过量表达,并拟从转基因植株耐湿性的表型特征来验证基因hverf2.11在湿害胁迫调控上的生物学功能。具体实验步骤如下:在前期筛选获得的hverf2.11基因转化拟南芥获得的t3代阳性转基因株系中,随机挑选各3个株系的种子,对照用拟南芥野生型种子(wt),在1/2ms培养基上培养至4叶期,将幼苗移栽到装有营养土的盆钵中,在人工气候室中培养35天。将转基因株系和野生型放入水箱中,进行淹水处理,淹水2周后,观察转基因材料与野生型材料耐湿性的差异,并取不同淹水时间段的材料进行耐湿相关基因的表达分析以及相关生理指标的测定,淹水结束后,测定不同材料的苗高、植株鲜重、干重、根系长度等性状。结果表明,野生型的株高下降了49.09%,而3个转基因株系分别下降了30.54%,27.41%,32.48%;野生型叶绿素含量下降了61.56%,而3个转基因株系分别下降了29.52%,35.26%,31.37%;野生型茎叶鲜重下降了65.8%,3个转基因株系分别下降了50%,19.67%,17.86%;野生型茎叶干重下降了51.01%,3个转基因株系分别下降了46.25%,40.5%,16.1%;野生型根长下降了74.87%,3个转基因株系分别下降了59.69%,41.47%,41.88%;湿害处理后,野生型拟南芥的存活率仅为27.61%,而转基因拟南芥的存活率高达78%以上(图3),表明过表达hverf2.11基因明显增强了拟南芥的耐湿性。
对照与湿害处理条件下野生型与转基因拟南芥抗氧化酶活性(sod、cat、pod)、乙醇脱氢酶(adh)活性和脯氨酸含量的变化如图4所示,正常生长条件下,转基因株系与野生型在生理生化水平方面没有明显差异;湿害处理3天,抗氧化酶和乙醇脱氢酶在野生型与转基因株系中均明显增加,湿害处理6天,增加达到最大值,转基因株系增加幅度明显要高于非转基因系;湿害处理9天后,抗氧化酶和乙醇脱氢酶活性开始下降(图4a,b,c,d)。湿害导致野生型与转基因株系的脯氨酸含量均明显增加,但野生型的增加幅度要明显大于转基因株系(图4e)。表明湿害胁迫下过表达hverf2.11基因增强了拟南芥中抗氧化酶系统清除有害物质的效率,提高了拟南芥对湿害胁迫的抗性。
为鉴定hverf2.11基因对湿害胁迫响应相关基因的调控效果,利用qrt-pcr对转基因拟南芥和野生型中与抗氧化酶活性、糖酵解和乙烯合成相关的基因(atsod1,atcat1,atpod1,atadh1,atpdc11,ataco1)进行基因表达分析。如图5所示,在正常生长条件下,atpod1和ataco1基因在野生型与转基因拟南芥间的表达差异达显著水平,其余基因均没有明显的表达差异;湿害胁迫后,6个基因的表达水平在野生型拟南芥和转基因拟南芥中均显著上调表达,且在转基因拟南芥中的上升幅度更大;湿害处理3天,atsod1、atpod1和ataco1基因表达水平达到最大值,处理6天和9天后开始逐渐降低(图5a,c,f);湿害处理3天,atcat1、atadh1和atpdc1基因表达水平上升速度较慢,湿害处理6天后,基因表达水平达到最高,湿害处理9天,表达水平开始降低(图5b,d,e)。表明hverf2.11基因可调控抗氧化酶、发酵、乙烯合成途径中与湿害胁迫相关的基因,增强拟南芥对湿害胁迫的抗性。
以上显示和描述了本发明的基本原理、主要特征以及本发明的优点。本行业的技术人员应该了解,本发明不受上述实施例的限制,上述实施例和说明书中描述的只是说明本发明的原理,在不脱离本发明精神和范围的前提下,本发明还会有各种变化和改进,这些变化和改进都落入要求保护的本发明范围内。本发明要求保护范围由所附的权利要求书及其等效物界定。
序列表
<110>扬州大学
<120>大麦耐湿调控基因hverf2.11、蛋白及其在育种中的应用
<130>xhx2019041602
<141>2019-04-16
<160>4
<170>siposequencelisting1.0
<210>1
<211>1158
<212>dna
<213>hordeumvulgarel.
<400>1
atgtgcggcggagccatcctcgcgggattcatcccgccgtcggcggccgcggcggcggcc60
aaggcagcggcgacggccaagaagaagcagcagcagcgcagcgtgacggcagactcgctc120
tggacgggcctgcggaaaaaggcggacgaggaggacttcgaggccgacttccgcgacttc180
gagcgggactccagcgaggaggaggacgacgaggtcgaggaggtcccccctccgccggcg240
ccggcgacggccgggttcgccttcgccgccgcggccgaggtcgcgctcagggcccctgcc300
cgccgagatgctgctgttcaacatgatggacctgctgccaaacaagtaaagcgcgttcgg360
aagaatcagtacagagggatccgccagcgtccctgggggaaatgggcagctgaaatccgt420
gaccctagcaagggtgtccgggtttggctcgggacatacgacactgctgaggaggcagcc480
agggcatatgatgctgaggcccgcaagatccgtggcaagaaagccaaggtcaattttcct540
gaggatgctccgactgttcagaagtctaccctgaagccaactgctgctaaatcagcaaag600
ctagctccacctccgaaggcctgcgaggatcagcctttcaatcatctgagcagaggagac660
aatgatttgttcgcgatgtttgccttcagtgacaagaaggttcctgcaaagccaactgac720
agtgtggattcccttcttccagtgaaacaccttgcccccaccgaggcattcggaatgaac780
atgctctctgaccagagcagcaattcatttggctccactgactttgggtgggacgacgag840
gccatgaccccggactacacgtccgtcttcgtaccgagtgctgctgccatgccggcgtac900
ggcgagcccgcttacctgcaaggcggagctccaaagagaatgaggaacaactttggcgta960
gctgtgctgcctcagggaaatggtgcacaagacatccctgcttttgacaatgaggtgaag1020
tactcgttgccctacgttgagagcagctcggacggatctatggacaaccttttgctgaat1080
ggtgcgatgcaggatggggcaagcagtggggatctctggagcctcgatgagctgttcatg1140
gcagctggtggttattga1158
<210>2
<211>385
<212>prt
<213>hordeumvulgarel.
<400>2
metcysglyglyalaileleualaglypheileproproseralaala
151015
alaalaalaalalysalaalaalathralalyslyslysglnglngln
202530
argservalthralaaspserleutrpthrglyleuarglyslysala
354045
aspglugluasppheglualaasppheargaspphegluargaspser
505560
sergluglugluaspaspgluvalglugluvalproproproproala
65707580
proalathralaglyphealaphealaalaalaalagluvalalaleu
859095
argalaproalaargargaspalaalavalglnhisaspglyproala
100105110
alalysglnvallysargvalarglysasnglntyrargglyilearg
115120125
glnargprotrpglylystrpalaalagluileargaspproserlys
130135140
glyvalargvaltrpleuglythrtyraspthralagluglualaala
145150155160
argalatyraspalaglualaarglysileargglylyslysalalys
165170175
valasnpheprogluaspalaprothrvalglnlysserthrleulys
180185190
prothralaalalysseralalysleualaproproprolysalacys
195200205
gluaspglnpropheasnhisleuserargglyaspasnaspleuphe
210215220
alametphealapheserasplyslysvalproalalysprothrasp
225230235240
servalaspserleuleuprovallyshisleualaprothrgluala
245250255
pheglymetasnmetleuseraspglnserserasnserpheglyser
260265270
thrasppheglytrpaspaspglualametthrproasptyrthrser
275280285
valphevalproseralaalaalametproalatyrglygluproala
290295300
tyrleuglnglyglyalaprolysargmetargasnasnpheglyval
305310315320
alavalleuproglnglyasnglyalaglnaspileproalapheasp
325330335
asngluvallystyrserleuprotyrvalgluserserseraspgly
340345350
sermetaspasnleuleuleuasnglyalametglnaspglyalaser
355360365
serglyaspleutrpserleuaspgluleuphemetalaalaglygly
370375380
tyr
385
<210>3
<211>20
<212>dna
<213>人工序列(artificialsequence)
<400>3
ccagtcagagatggtcaagg20
<210>4
<211>19
<212>dna
<213>人工序列(artificialsequence)
<400>4
catccatcgtcttgctgag19
- 还没有人留言评论。精彩留言会获得点赞!