活性提高的磷酸转酮酶及在生产代谢物中的应用的制作方法
本发明属于基因工程领域,具体涉及活性提高的磷酸转酮酶,以及该磷酸转酮酶在生产代谢物中的应用,涉及的代谢物包括但不限于氨基酸(特别是源自乙酰辅酶a的氨基酸)、琥珀酸、柠檬酸等。
背景技术:
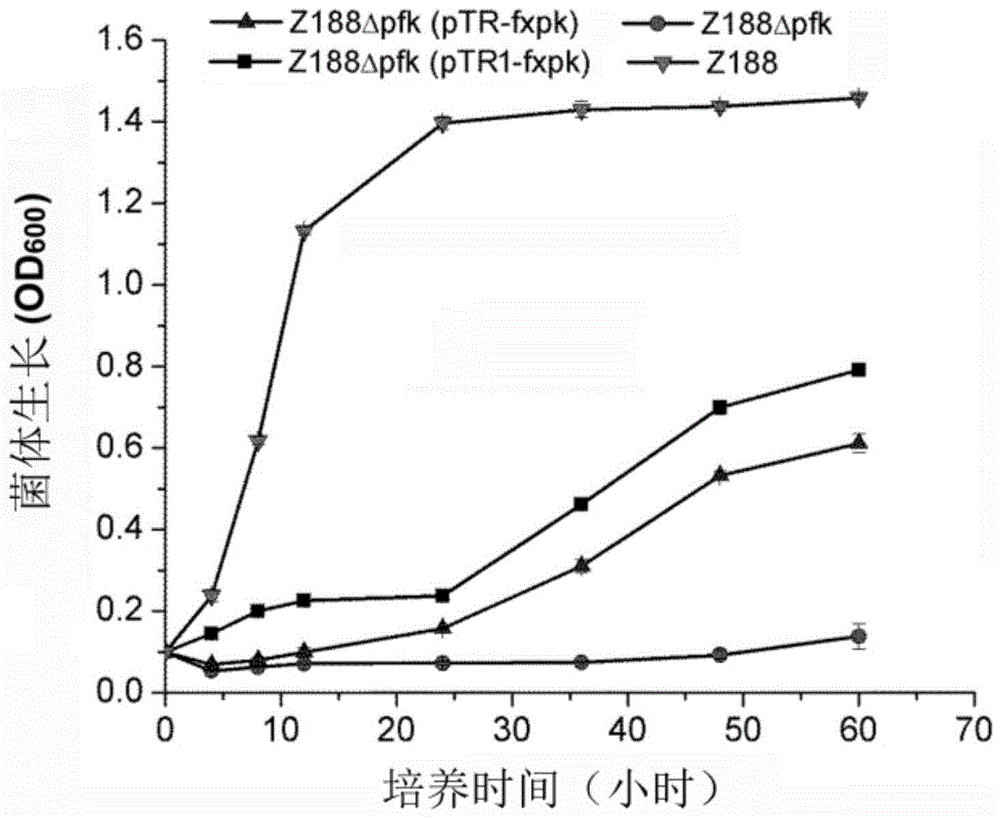
:氨基酸、有机酸,是人类和动物营养中最重要的成分,在医药、健康、食品、化工、动物饲料和化妆品等行业中有着十分重要的地位。当前,氨基酸、有机酸主要采用微生物发酵法来生产,已知的可生产氨基酸的微生物包括埃希氏菌属(escherichia),棒杆菌属(corynebavterium)、短杆菌属(brevibacterium)等。近年来,随着基因工程技术的发展,利用基因工程手段对菌株进行遗传改造,已经获得了大量的高效的氨基酸、有机酸生产工程菌株。磷酸转酮酶(phosphoketolase,f/xpk),包括6-磷酸果糖转酮酶(fpk)和5-磷酸木酮糖转酮酶(xpk)。6-磷酸果糖转酮酶催化的反应为:果糖-6-磷酸(f6p)+pi→乙酰磷酸(acp)+赤藓糖-4-磷酸(e4p)。5-磷酸木酮糖转酮酶催化的反应为:木酮糖-5-磷酸(x5p)+pi→乙酰磷酸(acp)+甘油醛-3-磷酸(g3p)。磷酸转酮酶催化的反应能够减少葡萄糖的氧化,但是通常使用的多种基因工程出发菌株(包括大肠杆菌和谷氨酸棒杆菌等),都不存在f/xpk,在这些菌株中引入f/xpk,能够增加源自乙酰辅酶a的代谢物的理论转化率。天然的f/xpk酶活性较低,需要大量过表达才能体现出效果,限制了其应用潜力。因此,本领域急需开发酶活性高的磷酸转酮酶。技术实现要素:本发明的目的是提供活性提高的磷酸转酮酶及在生产代谢物中的应用。本发明提供的蛋白质,为突变体蛋白,为将磷酸转酮酶进行如下(a1)至(a12)中任意一种或多种突变得到的蛋白质:(a1)对应于序列3的第2位氨基酸残基由t突变为a;(a2)对应于序列3的第6位氨基酸残基由i突变为t;(a3)对应于序列3的第14位氨基酸残基由n突变为d;(a4)对应于序列3的第20位氨基酸残基由e突变为d;(a5)对应于序列3的第120位氨基酸残基由t突变为a;(a6)对应于序列3的第231位氨基酸残基由e突变为k;(a7)对应于序列3的第260位氨基酸残基由h突变为y;(a8)对应于序列3的第342位氨基酸残基由e突变为k;(a9)对应于序列3的第397位氨基酸残基由k突变为r;(a10)对应于序列3的第676位氨基酸残基由d突变为g;(a11)对应于序列3的第785位氨基酸残基由f突变为l;(a12)对应于序列3的第801位氨基酸残基由w突变为r。所述多种突变具体可为以上突变中的任意两种、以上突变中的任意三种、以上突变中的任意四种、以上突变中的任意五种突变、以上突变中的任意六种、以上突变中的任意七种、以上突变中的任意八种、以上突变中的任意九种、以上突变中的任意十种、以上突变中的任意十一种或者以上突变的所有。本发明提供的突变体蛋白具体可为将磷酸转酮酶进行(a1)和(a2)两种突变得到的蛋白质。本发明提供的突变体蛋白具体可为将磷酸转酮酶进行特定突变和非特定突变得到的蛋白质。所述特定突变为(a1)和(a2)两种突变。所述非特定突变为如下突变中的任意一种或任意组合:(a3)、(a4)、(a5)、(a6)、(a7)、(a8)、(a9)、(a10)、(a11)、(a12)。本发明提供的突变体蛋白具体可为将磷酸转酮酶进行(a1)和(a2)和(a7)三种突变得到的蛋白质。本发明提供的突变体蛋白具体可为将磷酸转酮酶进行特定突变和非特定突变得到的蛋白质。所述特定突变为(a1)和(a2)和(a7)三种突变。所述非特定突变为如下突变中的任意一种或任意组合:(a3)、(a4)、(a5)、(a6)、(a8)、(a9)、(a10)、(a11)、(a12)。所述(a1)具体可为(b1)。所述(a2)具体可为(b2)。所述(a3)具体可为(b3)。所述(a4)具体可为(b4)。所述(a5)具体可为(b5)。所述(a6)具体可为(b6)。所述(a7)具体可为(b7)。所述(a8)具体可为(b8)。所述(a9)具体可为(b9)。所述(a10)具体可为(b10)。所述(a11)具体可为(b11)。所述(a12)具体可为(b12)。(b1)第2位氨基酸残基由t突变为a。(b2)第6位氨基酸残基由i突变为t。(b3)第14位氨基酸残基由n突变为d。(b4)第20位氨基酸残基由e突变为d。(b5)第120位氨基酸残基由t突变为a。(b6)第231位氨基酸残基由e突变为k。(b7)第260位氨基酸残基由h突变为y。(b8)第342位氨基酸残基由e突变为k。(b9)第397位氨基酸残基由k突变为r。(b10)第676位氨基酸残基由d突变为g。(b11)第785位氨基酸残基由f突变为l。(b12)第801位氨基酸残基由w突变为r。所述磷酸转酮酶具体可为序列表的序列3所示的蛋白质。所述磷酸转酮酶还可为与序列表的序列3所示的蛋白质具有90%,优选95%,更优选98%,最优选99%同源性的蛋白或多肽。示例性的,所述突变体蛋白可为实施例中的m21蛋白、m41蛋白、m71蛋白、m81蛋白、m82蛋白、t2a蛋白、i6t蛋白、n14d蛋白、e20d蛋白、t120a蛋白、e231k蛋白、h260y蛋白、e342k蛋白、k397r蛋白、d676g蛋白、f785l蛋白、w801r蛋白、t2a/i6t蛋白、t2a/h260y蛋白、i6t/h260y蛋白或t2a/i6t/h260y蛋白。编码以上任一所述突变体蛋白的多核苷酸(例如基因,该基因命名为突变体基因)也属于本发明的保护范围。示例性的,所述突变体基因具体可为将磷酸转酮酶基因的编码框进行如下(c1)至(c12)中任意一种或多种突变得到的dna分子:(c1)第4位核苷酸由a突变为g;(c2)第17位核苷酸由t突变为c;(c3)第40位核苷酸由a突变为g;(c4)第60位核苷酸由a突变为t;(c5)第358位核苷酸由a突变为g;(c6)第691位核苷酸由g突变为a;(c7)第778位核苷酸由c突变t为;(c8)第1024位核苷酸由g突变为a;(c9)第1190位核苷酸由a突变为g;(c10)第2027位核苷酸由a突变为g;(c11)第2353位核苷酸由t突变为c;(c12)第2401位核苷酸由t突变为c。本发明提供的突变体基因具体可为将磷酸转酮酶基因的编码框进行(c1)和(c2)两种突变得到的dna分子。本发明提供的突变体基因具体可为将磷酸转酮酶基因的编码框进行特定突变和非特定突变得到的dna分子。所述特定突变为(c1)和(c2)两种突变。所述非特定突变为如下突变中的任意一种或任意组合:(c3)、(c4)、(c5)、(c6)、(c7)、(c8)、(c9)、(c10)、(c11)、(c12)。本发明提供的突变体蛋白具体可为将磷酸转酮酶基因的编码框进行(c1)和(c2)和(c7)三种突变得到的dna分子。本发明提供的突变体蛋白具体可为将磷酸转酮酶基因的编码框进行特定突变和非特定突变得到的dna分子。所述特定突变为(c1)和(c2)和(c7)三种突变。所述非特定突变为如下突变中的任意一种或任意组合:(c3)、(c4)、(c5)、(c6)、(c8)、(c9)、(c10)、(c11)、(c12)。磷酸转酮酶基因的编码框具体可如序列表的序列4所示。具有以上任一所述多核苷酸的表达盒、重组载体、重组离体细胞或重组微生物均属于本发明的保护范围。具有以上任一所述突变体蛋白的融合蛋白也属于本发明的保护范围。所述融合蛋白可为将所述突变体蛋白与蛋白标签融合得到的蛋白质。蛋白标签可位于突变体蛋白的n端,也可位于突变体蛋白的c端。突变体蛋白和蛋白标签之间还可以具有间隔氨基酸残基,具体可具有10个以下间隔氨基酸残基。示例性的标签具体如表1所示。表1标签的序列标签残基序列poly-arg5-6(通常为5个)rrrrrpoly-his2-10(通常为6个)hhhhhhflag8dykddddkstrep-tagii8wshpqfekc-myc10eqkliseedlha9ypydvpdya所述融合蛋白自n端至c端依次由如下元件组成:突变体蛋白、间隔序列、蛋白标签。蛋白标签具体可为his6标签。间隔序列具体可由10个以下氨基酸残基组成。间隔序列具体可为“le”。示例性的,所述融合蛋白可为实施例中的fxpk-his6蛋白、m21-his6蛋白、m41-his6蛋白、m71-his6蛋白、m81-his6蛋白、m82-his6蛋白、t2a-his6蛋白、i6t-his6蛋白、n14d-his6蛋白、e20d-his6蛋白、t120a-his6蛋白、e231k-his6蛋白、h260y-his6蛋白、e342k-his6蛋白、k397r-his6蛋白、d676g-his6蛋白、f785l-his6蛋白、w801r-his6蛋白、t2a/i6t-his6蛋白、t2a/h260y-his6蛋白、i6t/h260y-his6蛋白或t2a/i6t/h260y-his6蛋白。编码所述融合蛋白的多核苷酸(例如基因)也属于本发明的保护范围。具有编码所述融合蛋白的多核苷酸的表达盒、重组载体、重组离体细胞或重组微生物均属于本发明的保护范围。示例性的,以上任一所述重组载体具体可为实施例中的重组质粒ptr-fxpk、重组质粒ptr1-fxpk、重组质粒ptr-m21、重组质粒ptr-m41、重组质粒ptr-m71、重组质粒ptr-m81、重组质粒ptr-m82、重组质粒ptr-t2a、重组质粒ptr-i6t、重组质粒ptr-h260y、重组质粒ptr-t2a/i6t、重组质粒ptr-t2a/i6t/h260y、重组质粒pet-fxpk、重组质粒pet-m21、重组质粒pet-m41、重组质粒pet-m71、重组质粒pet-m81、重组质粒pet-m82、重组质粒pet-t2a、重组质粒pet-i6t、重组质粒pet-n14d、重组质粒pet-e20d、重组质粒pet-t120a、重组质粒pet-e231k、重组质粒pet-h260y、重组质粒pet-e342k、重组质粒pet-k397r、重组质粒pet-d676g、重组质粒pet-f785l、重组质粒pet-w801r、重组质粒pet-t2a/i6t、重组质粒pet-t2a/h260y、重组质粒pet-i6t/h260y、重组质粒pet-t2a/i6t/h260y或质粒psil-fxpk。所述重组微生物可为将以上任一所述重组载体导入宿主微生物得到的重组微生物。示例性的,以上任一所述重组微生物具体可为实施例中的菌株z188△pfk(ptr-fxpk)或菌株z188△pfk(ptr1-fxpk),以及实施例5至实施例11中得到的各个重组菌。所述宿主微生物可为氨基酸生产菌株或有机酸生产菌株。所述宿主微生物包括但不限于谷氨酸棒杆菌、大肠杆菌或黑曲霉,等等。示例性的,所述宿主微生物可为谷氨酸棒杆菌z188、菌株z188△pfk、大肠杆菌bl21(de3)、大肠杆菌琥珀酸生产菌株(例如cgmccno.5107、cgmccno.5108或cgmccno.5109等)、谷氨酰胺生产菌株(例如实施例8中的谷氨酰胺生产菌株)、大肠杆菌脯氨酸生产菌株(例如dh5α(psw2))、大肠杆菌反式-4-羟基-l-脯氨酸生产菌株(例如dh5α(psw3))、黑曲霉(例如co827(cicc40347))。本发明还保护特定物质在制备代谢物中的应用;所述特定物质为:以上任一所述突变体蛋白、以上任一所述融合蛋白、以上任一所述突变体基因、以上任一所述编码所述融合蛋白的基因、以上任一所述表达盒、以上任一所述重组载体、以上任一所述重组离体细胞或以上任一所述重组微生物。所述代谢物具体可为氨基酸。示例性的,所述氨基酸具体可为源自乙酰辅酶a的氨基酸。示例性的,所述氨基酸包括但不限于谷氨酸、谷氨酰胺、脯氨酸、反式-4-羟基-l-脯氨酸等。所述代谢物具体可为有机酸。示例性的,所述有机酸包括但不限于琥珀酸、柠檬酸等。本发明还保护一种提高磷酸转酮酶酶活的方法,包括如下步骤:将磷酸转酮酶进行如下(a1)至(a12)中任意一种或多种突变:(a1)至(a12)同上。所述多种突变具体可为以上突变中的任意两种、以上突变中的任意三种、以上突变中的任意四种、以上突变中的任意五种突变、以上突变中的任意六种、以上突变中的任意七种、以上突变中的任意八种、以上突变中的任意九种、以上突变中的任意十种、以上突变中的任意十一种或者以上突变的所有。所述方法中,具体可进行(a1)和(a2)两种突变。所述方法中,具体可进行特定突变和非特定突变。所述特定突变为(a1)和(a2)两种突变。所述非特定突变为如下突变中的任意一种或任意组合:(a3)、(a4)、(a5)、(a6)、(a7)、(a8)、(a9)、(a10)、(a11)、(a12)。所述方法中,具体可进行(a1)和(a2)和(a7)三种突变。所述方法中,具体可进行特定突变和非特定突变。所述特定突变为(a1)和(a2)和(a7)三种突变。所述非特定突变为如下突变中的任意一种或任意组合:(a3)、(a4)、(a5)、(a6)、(a8)、(a9)、(a10)、(a11)、(a12)。所述(a1)具体可为(b1)。所述(a2)具体可为(b2)。所述(a3)具体可为(b3)。所述(a4)具体可为(b4)。所述(a5)具体可为(b5)。所述(a6)具体可为(b6)。所述(a7)具体可为(b7)。所述(a8)具体可为(b8)。所述(a9)具体可为(b9)。所述(a10)具体可为(b10)。所述(a11)具体可为(b11)。所述(a12)具体可为(b12)。所述磷酸转酮酶具体可为序列表的序列3所示的蛋白质。所述磷酸转酮酶还可为与序列表的序列3所示的蛋白质具有90%,优选95%,更优选98%,最优选99%同源性的蛋白或多肽。本发明还保护一种制备代谢物的方法,包括如下步骤:通过培养以上任一所述的重组微生物制备代谢物。所述方法还包括将代谢物从培养体系中分离纯化出来的步骤。所述代谢物具体可为氨基酸。示例性的,所述氨基酸具体可为源自乙酰辅酶a的氨基酸。示例性的,所述氨基酸包括但不限于谷氨酸、谷氨酰胺、脯氨酸、反式-4-羟基-l-脯氨酸等。所述代谢物具体可为有机酸。示例性的,所述有机酸包括但不限于琥珀酸、柠檬酸等。本发明还保护一种获得磷酸转酮酶酶活增高的蛋白质的方法,包括如下步骤:(1)将现有蛋白质与参照蛋白质进行序列比对;所述现有蛋白质为现有的具有磷酸转酮酶酶活的蛋白质;所述参照蛋白质为序列表的序列3所示的蛋白质;(2)根据比对结果,将参照蛋白质中的特定突变对应于现有蛋白质,然后将现有蛋白质进行突变,得到磷酸转酮酶酶活高于所述现有蛋白质的新蛋白质;所述特定突变为如下(a1)至(a12)中任意一种或多种突变:(a1)至(a12)同上。所述(a1)具体可为(b1)。所述(a2)具体可为(b2)。所述(a3)具体可为(b3)。所述(a4)具体可为(b4)。所述(a5)具体可为(b5)。所述(a6)具体可为(b6)。所述(a7)具体可为(b7)。所述(a8)具体可为(b8)。所述(a9)具体可为(b9)。所述(a10)具体可为(b10)。所述(a11)具体可为(b11)。所述(a12)具体可为(b12)。所述获得磷酸转酮酶酶活增高的蛋白质的方法还包括检测新蛋白质和现有蛋白质的磷酸转酮酶酶活,从而将酶活增高的蛋白质选择出来的步骤。以上任一所述磷酸转酮酶包括但不限于6-磷酸果糖转酮酶。以上任一所述磷酸转酮酶酶活包括但不限于6-磷酸果糖转酮酶酶活。所述重组离体细胞是将所述基因导入离体宿主细胞得到的。所述宿主细胞具体可为可以生产氨基酸、有机酸的宿主细胞。本文所用的术语“宿主细胞”是具有本领域普通技术人员通常理解的含义,即,含有本发明的磷酸转酮酶且能够生产氨基酸、有机酸特别是源自乙酰辅酶a的氨基酸、有机酸的细胞。换言之,本发明可以利用任何宿主细胞,只要所述细胞中的含有本发明所述的磷酸转酮酶且能够生产氨基酸、有机酸等化合物。所述重组微生物是将所述基因导入宿主微生物得到的。所述宿主微生物具体可为可以生产氨基酸、有机酸的宿主微生物。本文所用的术语“宿主微生物”是具有本领域普通技术人员通常理解的含义,即,含有本发明的磷酸转酮酶且能够生产氨基酸、有机酸特别是源自乙酰辅酶a的氨基酸、有机酸的微生物。换言之,本发明可以利用任何宿主微生物,只要所述宿主微生物中的含有本发明所述的磷酸转酮酶且能够生产氨基酸、有机酸等化合物。所述宿主微生物包括但不限于埃希氏菌属(escherichia)、棒状杆菌属(corynebacterium)、泛菌属(pantoea)、短杆菌属(brevibacteriumsp)、芽孢杆菌属(bacillus)、克雷伯氏菌属(klebsiella)、沙雷氏菌属(serratia)或弧菌属(vibrio)的微生物。所述宿主微生物优选为或谷氨酸棒状杆菌(corynebacteriumglutamicum),最优选为谷氨酸棒状杆菌。所述宿主微生物具体可为谷氨酸棒杆菌z188。本发明中,发明人敲除氨基酸生产菌株的6-磷酸果糖激酶(pfk)基因,将外源磷酸转酮酶的活性与菌株的生长速度偶联,从而在野生型fxpk蛋白的基础上通过生长富集发现了酶活提高的突变体蛋白以及可以增加现有磷酸转酮酶酶活的多个突变位点。与现有磷酸转酮酶相比,本发明提供的突变体蛋白的磷酸转酮酶酶活显著增加。采用本发明提供的突变位点对现有磷酸转酮酶进行单点或多点突变,可以显著提高其磷酸转酮酶酶活。本发明提供方案,可以实现更高的磷酸转酮酶活性,进而显著提高目标代谢物的产量。附图说明图1为实施例2中的结果。图2为实施例3中的结果。图3为实施例5中的结果。图4为实施例6中的结果。具体实施方式本发明的“磷酸转酮酶”包括6-磷酸果糖转酮酶(fpk)和/或5-磷酸木酮糖转酮酶(xpk),是指能够催化果糖-6-磷酸生成乙酰磷酸和赤藓糖-4-磷酸的酶和/或能够催化木酮糖-5-磷酸生成乙酰磷酸和甘油醛-3-磷酸的酶。“5-磷酸木酮糖转酮酶”可以是一种酶,其来自具有5-磷酸木酮糖转酮酶活性的细菌,包括但不限于乳酸菌属、甲醇同化细菌属、甲烷同化细菌属、链球菌属(streptococcus)等,可优选醋杆菌属(acetobacter)、双歧杆菌属(bifidobacterium)、乳杆菌属(lactobacillus)、硫杆菌属(thiobacillus)、链球菌属(streptoccus)、甲基球菌属(methylococus)、丁酸杆菌属(buryrivibrio)、丝状菌属(fibrobacter)的细菌,也可优选属于假丝酵母属(candida)、红酵母属(rhodotorula)、红冬孢酵母属(rhodosporidium)、毕赤酵母属(pichia)、汉逊酵母属(hansenula)、克鲁维酵母属(kluyveromyces)、酵母属(saccharomyces)、丝孢酵母属(trichosporon)、温酵母属(wingea)等的酵母。“6-磷酸果糖转酮酶”可以是一种酶,其来自于具有6-磷酸果糖转酮酶活性的细菌,包括但不限于醋杆菌属(acetobacter)、双歧杆菌属(bifidobacterium)、绿菌属(chlorobium)、布鲁氏菌属(brucella)、甲基球菌属(methylococus)、加德纳氏菌属(gardnerella)的细菌,包括但不限于属于红酵母属(rhodotorula)、假丝酵母属(trichosporon)、酵母属(saccharomyces)等的酵母。磷酸转酮酶也可能是一个酶,呈现出6-磷酸果糖转酮酶(fpk)和5-磷酸木酮糖转酮酶(xpk)两种活性。关于本发明,磷酸转酮酶可以来源于青春双歧杆菌。本发明中通过对序列表的序列3所示蛋白质进行突变获得了酶活提高的突变体蛋白,本领域技术人员可以任意进行突变,只要其相对于其自然状态酶活有所提高,也在本发明的保护范围内。磷酸转酮酶可以来源于各种物种,包括但不限于青春双歧杆菌、乳酸双歧杆菌(b.lactis)、戊糖乳杆菌(l.pentosus)、植物乳杆菌(l.plantarum)等。本文所用的术语“自然状态”是指微生物中多肽处于未修饰状态的活性,即自然状态下的活性。本文所用的术语“含有本发明的磷酸转酮酶”具有本领域技术人员常规理解的含义,并且可以通过本领域已知的方法实施,包括但不限于,如:将包含编码蛋白的多核苷酸序列的多核苷酸插入到染色体上,和/或将多核苷酸克隆到载体上引入微生物,和/或在染色体上行直接增加该多核苷酸的拷贝等方法来实现,也可以非限制性地包括任何已知的可以引入蛋白活性的方法。本发明所述“氨基酸、有机酸特别是源自乙酰辅酶a的氨基酸、有机酸”是指如l-谷氨酸、l-谷氨酰胺、l-脯氨酸、l-羟脯氨酸(反式-4-羟基-l-脯氨酸)、l-精氨酸、l-亮氨酸、l-异亮氨酸、l-半胱氨酸、柠檬酸、琥珀酸等以乙酰辅酶a为底物的氨基酸、有机酸。本发明所述“氨基酸、有机酸生产菌株”是指,当细菌在培养中被培养时能够产生氨基酸、有机酸并且能够积累氨基酸、有机酸,或者能够将氨基酸、有机酸分泌到培养基中,也就是能够得到胞外的游离氨基酸、有机酸,特别是指与野生型菌株或者亲本菌株相比,能够积累更多氨基酸、有机酸的能力。为了赋予菌株产氨基酸、有机酸的能力,可以采用传统的育种方法,比如培育营养缺陷型的突变株、抗类似物的菌株,或者能够产氨基酸、有机酸的代谢控制突变株,以及培育氨基酸、有机酸生物合成相关酶活性提高的重组菌株的方法,或者以上方法的组合。本领域技术人员知晓,为提升活性而对野生型多肽进行突变,找到能实现所需目的的位点更为重要。因此,基于本发明的教导,本领域技术人员会对序列3所示蛋白质的第2位、第6位、第14位、第20位、第120位、第231位、第260位、第342位、第397位、第676位、第785位、第801位的氨基酸残基进行突变,并检测突变体的相关活性。在具体的实施方式中,本发明的磷酸转酮酶突变体蛋白在对应于序列3所示蛋白质的第2位氨基酸残基为a,和/或第6位氨基酸残基为t,和/或第14位氨基酸残基为d,和/或第20位氨基酸残基为d,和/或第120位氨基酸残基为a,和/或第231位氨基酸残基为k,和/或第260位氨基酸残基为y,和/或第342位氨基酸残基为k,和/或第397位氨基酸残基为r,和/或第676位氨基酸残基为g,和/或第785位氨基酸残基为l,和/或第801位氨基酸残基为r。此外,本领域普通技术人员也不难知晓,在多肽的某些区域,例如非重要区域改变少数氨基酸残基基本上不会改变生物活性,例如,适当替换某些氨基酸得到的序列并不会影响其活性(可参见watson等,molecularbiologyofthegene,第四版,1987,thebenjamin/cummingspub.co.p224)。因此,本领域普通技术人员能够实施这种替换并且确保所得分子仍具有所需生物活性。因此,对本发明的磷酸转酮酶突变体作进一步突变而得到仍具备磷酸转酮酶的功能和活性的进一步突变体是显而易见的。例如,本领域技术人员公知在多肽的任一端增加或减少数个氨基酸残基,例如优选1-20个、更优选1-15个、更优选1-10个、更优选1-3个、最优选1个氨基酸残基不会影响得到的突变体的功能。例如,为便于纯化,技术人员往往在得到的蛋白的任一端带上6×his标签,而这种蛋白与不具备6×his标签的蛋白具有相同的功能。因此,本发明应包括本发明的磷酸转酮酶的保守性突变体。这些保守性突变体可以根据,例如表2所示进行氨基酸替换而产生。表2初始残基代表性的取代残基优选的取代残基ala(a)val;leu;ilevalarg(r)lys;gln;asnlysasn(n)gln;his;lys;argglnasp(d)gluglucys(c)sersergln(q)asnasnglu(e)aspaspgly(g)pro;alaalahis(h)asn;gln;lys;argargile(i)leu;val;met;ala;pheleuleu(l)ile;val;met;ala;pheilelys(k)arg;gln;asnargmet(m)leu;phe;ileleuphe(f)leu;val;ile;ala;tyrleupro(p)alaalaser(s)thrthrthr(t)sersertrp(w)tyr;phetyrtyr(y)trp;phe;thr;serpheval(v)ile;leu;met;phe;alaleu本发明还提供了编码本发明多肽的多核苷酸。术语“编码多肽的多核苷酸”可以是包括编码此多肽的多核苷酸,也可以是还包括附加编码和/或非编码序列的多核苷酸。因此,本文所用的“含有”,“具有”或“包括”包括了“包含”、“主要由……构成”、“基本上由……构成”、和“由……构成”;“主要由……构成”、“基本上由……构成”和“由……构成”属于“含有”、“具有”或“包括”的下位概念。本文所用的术语“对应于”具有本领域普通技术人员通常理解的意义。具体地说,“对应于”表示两条序列经同源性或序列相同性比对后,一条序列与另一条序列中的指定位置相对应的位置。因此,例如,就“对应于序列3所示蛋白质的第40位的氨基酸残基”而言,如果在序列3所示蛋白质的一端加上6×his标签,那么所得突变体中对应于序列3所示氨基酸序列的第40位就可能是第46位。在具体的实施方式中,只要所述同源性或序列相同性在90%以上,优选95%以上,更优选96%、97%、98%、99%以上,且具有磷酸转酮酶活性(即6-磷酸果糖转酮酶活性和/或5-磷酸木酮糖转酮酶活性),也在本发明的保护范围内。本领域普通技术人员公知的测定序列同源性或相同性的方法包括但不限于:计算机分子生物学(computationalmolecularbiology),lesk,a.m.编,牛津大学出版社,纽约,1988;生物计算:信息学和基因组项目(biocomputing:informaticsandgenomeprojects),smith,d.w.编,学术出版社,纽约,1993;序列数据的计算机分析(computeranalysisofsequencedata),第一部分,griffin,a.m.和griffin,h.g.编,humanapress,新泽西,1994;分子生物学中的序列分析(sequenceanalysisinmolecularbiology),vonheinje,g.,学术出版社,1987和序列分析引物(sequenceanalysisprimer),gribskov,m.与devereux,j.编mstocktonpress,纽约,1991和carillo,h.与lipman,d.,siamj.appliedmath.,48:1073(1988)。测定相同性的优选方法要在测试的序列之间得到最大的匹配。测定相同性的方法编译在公众可获得的计算机程序中。优选的测定两条序列之间相同性的计算机程序方法包括但不限于:gcg程序包(devereux,j.等,1984)、blastp、blastn和fasta(altschul,s,f.等,1990)。公众可从ncbi和其它来源得到blastx程序(blast手册,altschul,s.等,ncbinlmnihbethesda,md.20894;altschul,s.等,1990)。熟知的smithwaterman算法也可用于测定相同性。下面结合具体实施例,进一步阐述本发明。应理解,这些实施例仅用于说明本发明而不用于限制本发明的范围。下列实施例中未注明具体条件的实验方法,通常按照常规条件如sambrook等人,分子克隆:实验室手册(newyork:coldspringharborlaboratorypress,1989)中所述的条件,或按照制造厂商所建议的条件。下述实施例中所用的试验材料,如无特殊说明,均为自常规生化试剂公司购买得到的。以下实施例中的定量试验,均设置三次重复实验,结果取平均值。记载有质粒pk18mobsacb的文献(plasmidpk18mobsacb),记载于如下文献:schafera,taucha,jagerw,kalinowskij,thierbachg,puhlera(1994)smallmobilizablemultipurposecloningvectorsderivedfromtheescherichiacoliplasmidspk18andpk19-selectionofdefineddeletionsinthechromosomeofcorynebacteriumglutamicum.gene145(1):69-73.https://doi.org/10.1016/0378-1119(94)90324-7。质粒ptrcmob(plasmidptrcmob),记载于如下文献:liu,q.,etal.(2007).journalofbiotechnology132(2007)273–279。液体cgxii无机盐培养基的配方见表3(调节ph为7.0)。表3培养基成分含量(每升)葡萄糖50g(nh4)2so420g尿素5gkh2po41gk2hpo41gmgso40.25g3-吗啉丙磺酸42gcacl20.01gfeso4·7h200.01gmnso4·h200.01gzns04·7h200.001g原儿茶酸0.03gcuso40.2mgnicl2·6h200.02mg生物素0.2mg维生素b10.1mg实施例1、构建敲除pfk基因的菌株,即菌株z188△pfk谷氨酸棒杆菌z188,即谷氨酸棒杆菌(corynebacteriumglutamicum)z188。谷氨酸棒杆菌z188的全基因组见genbankaccessionnumber:nz_akxp00000000.1(https://www.ncbi.nlm.nih.gov/nuccore/nz_akxp00000000)。pfk基因即6-磷酸果糖激酶基因。谷氨酸棒杆菌z188的基因组dna中,pfk基因的编码框及其上下游各1000bp部分的核苷酸序列见序列表的序列2(序列表的序列2中,第1001-2041位核苷酸为编码框,编码序列表的序列1所示的蛋白质)。δpfk-f1:cctcgaattcggatgctgccaatggaatggtgcccagtg;δpfk-r1:cttcaaggttaaattcattgctggctgtgc;δpfk-f2:caatgaatttaaccttgaaggaagttccattc;δpfk-r2:tctactgcagggaatgatgacaccgatggtgtctgtcctcgac。1、以谷氨酸棒杆菌z188的基因组dna为模板,采用δpfk-f1和δpfk-r1组成的引物对进行pcr扩增,得到pcr扩增产物。2、以谷氨酸棒杆菌z188的基因组dna为模板,采用δpfk-f2/δpfk-r2组成的引物对进行pcr扩增,得到pcr扩增产物。3、将步骤1的pcr扩增产物和步骤2的pcr扩增产物同时作为模板,采用δpfk-f1/δpfk-r2组成的引物对进行pcr扩增,得到pcr扩增产物。4、取步骤3得到的pcr扩增产物,采用限制性内切酶ecori和psti进行双酶切,回收酶切产物。5、取质粒pk18mobsacb,采用限制性内切酶ecori和psti进行双酶切,回收载体骨架。6、将步骤4的酶切产物和步骤5的载体骨架连接,得到重组质粒。7、利用步骤6得到的重组质粒,根据文献(niebischandbott,(2001).archmicrobiol175(4):282-294.schaferetal.,(1994).gene145(1):69-73)中记载的两步rec重组的方法敲除谷氨酸棒杆菌z188的pfk基因,得到敲除pfk基因的菌株,将其命名为菌株z188△pfk。菌株z188△pfk进行测序。与谷氨酸棒杆菌z188的基因组dna相比,菌株z188△pfk的基因组的差异仅在于,缺失了序列表的序列2所示dna分子的第1100-1983位核苷酸所示的区段。实施例2、重组菌的制备以及生长性能来源于青春双歧杆菌的磷酸转酮酶如序列表的序列3所示。对序列表的序列3所示的蛋白质的编码基因进行全密码子优化,优化后基因如序列表的序列4所示。将序列表的序列3所示的磷酸转酮酶又称为fxpk蛋白或f/xpk蛋白。将编码fxpk蛋白的基因又称为fxpk基因。一、制备重组质粒1、合成序列表的序列4所示的双链dna分子;将合成的双链dna分子作为模板,采用f1和r1组成的引物对进行pcr扩增,回收pcr扩增产物。f1:ctacgaattcgaaggagatatacatatg;r1:tcagggatcctcattcgttgtcacccgcggtc。2、取步骤1得到的pcr扩增产物,采用限制性内切酶ecori和bamhi进行双酶切,回收酶切产物。3、取质粒ptrcmob,采用限制性内切酶ecori和bamhi进行双酶切,回收载体骨架。4、将步骤2的酶切产物和步骤3的载体骨架连接,得到重组质粒ptr-fxpk。经测序验证,重组质粒ptr-fxpk中具有特异dna分子甲。所述特异dna分子甲自上游至下游依次由如下五个元件组成:序列表的序列5所示的dna分子(序列5中第1-246位核苷酸组成启动子)、ecori酶切识别序列“gaattc”、核糖体结合位点“gaaggagatatacat”、序列表的序列4所示的dna分子、bamhi酶切识别序列“ggatcc”。5、以重组质粒ptr-fxpk为模板,采用ptrc-1和ptrc-2组成的引物对进行单点突变,得到重组质粒ptr1-fxpk。本步骤的目的是在启动子区引入单点突变,从而增加启动子的启动活性,促进基因的表达。ptrc-1:gagcggataacaatctcacacaggaaacag;ptrc-2:ctgtttcctgtgtgagattgttatccgctc。重组质粒ptr1-fxpk进行测序验证。与重组质粒ptr-fxpk相比,重组质粒ptr1-fxpk的差异仅在于用序列表的序列6所示的dna分子取代了序列表的序列5所示的dna分子。二、制备重组菌将重组质粒ptr-fxpk导入菌株z188△pfk,得到重组菌,命名为菌株z188△pfk(ptr-fxpk)。将重组质粒ptr1-fxpk导入菌株z188△pfk,得到重组菌,命名为菌株z188△pfk(ptr1-fxpk)。三、比较菌株的生长性能供试菌株分别为:谷氨酸棒杆菌z188、菌株z188△pfk、菌株z188△pfk(ptr-fxpk)、菌株z188△pfk(ptr1-fxpk)。将供试菌株接种至液体cgxii无机盐培养基(初始体系的od600nm值=0.1),30℃、850rpm振荡培养,不同时间取样(各200μl),利用酶标仪检测600nm下的吸光度(od600nm值)。结果见图1。菌株z188△pfk在液体cgxii无机盐培养基中几乎不生长。菌株z188△pfk(ptr-fxpk)和菌株z188△pfk(ptr1-fxpk)都可以正常生长,fxpk基因表达水平越高,菌株生长速度越快。结果表明,fxpk酶活/含量的增加能够促进菌株的生长,从而可以通过生长富集的方式从突变体库中筛选酶活提高的突变蛋白。实施例3、获得酶活性提高的突变蛋白一、构建fxpk基因突变体库1、以重组质粒ptr-fxpk为模板,采用f1和r1组成的引物对,进行易错pcr扩增。易错pcr扩增,采用dna聚合酶(北京全式金生物)。分别设置三个反应体系,分别含有0.2mm、0.5mm或0.8mm的氯化锰。完成易错pcr扩增后,将三个反应体系合并。2、取步骤1的产物,采用限制性内切酶ecori和bamhi进行双酶切,回收酶切产物。3、取重组质粒ptr-fxpk,采用限制性内切酶ecori和bamhi进行双酶切,回收载体骨架。4、将步骤2的酶切产物与步骤3的载体骨架连接,得到重组质粒。5、将步骤4得到的重组质粒导入菌株z188△pfk,得到重组菌。由于易错pcr扩增产生了多种fxpk基因的突变基因,在步骤4中得到了多种重组质粒(约1万种),形成fxpk基因突变重组质粒库,相应的,在步骤5中得到了多种重组菌,形成fxpk基因突变重组菌库。采用相同的方法构建了9个fxpk基因突变重组菌库,依次命名为m1重组菌库至m9重组菌库。二、通过生长富集获得fxpk酶活性提高的突变体利用液体cgxii无机盐培养基在24深孔板中培养9个fxpk基因突变重组菌库(每孔1ml培养基),30℃、850rpm振荡培养,每隔48小时转接一次(接种量为1%)。每次培养后,取200μl到微孔板中,利用酶标仪检测600nm下的吸光度(od600nm值)。将菌株z188△pfk(ptr-fxpk)作为对照菌株。结果见图2。随着连续传代培养,m2重组菌库、m4重组菌库、m7重组菌库和m8重组菌库中均富集到能够生长加快的菌株。对m2重组菌库、m4重组菌库、m7重组菌库和m8重组菌库分别进行测序分析,从m2重组菌库、m4重组菌库、m7重组菌库中各筛选到一个fxpk基因的突变基因,从m8重组菌库中筛选到2个fxpk基因的突变基因。从而,得到5个fxpk蛋白的突变蛋白。将各个突变蛋白分别命名为m21蛋白、m41蛋白、m71蛋白、m81蛋白和m82蛋白。与fxpk蛋白相比各个突变蛋白的突变氨基酸,以及与fxpk基因(序列表的序列4中第26-2503位核苷酸所示)相比各个突变基因的突变核苷酸,见表4。表4(1-14)表示核苷酸突变导致对应的氨基酸发生突变,(-)表示核苷酸突变没有导致氨基酸突变发生。实施例4、用于表达各个蛋白质的各个重组质粒的构建fxpk蛋白如序列表的序列3所示。fxpk基因如序列表的序列4所示。一、分别制备各个重组质粒,并测序验证。重组质粒ptr-m21。与重组质粒ptr-fxpk相比,重组质粒ptr-m21的差异仅在于用m21d基因取代了fxpk基因。与fxpk基因相比,m21d基因发生了如下5个核苷酸的突变:第17位核苷酸由t突变为c,第358位核苷酸由a突变为g,第691位核苷酸由g突变为a,第1190位核苷酸由a突变为g,第2027位核苷酸由a突变为g。m21d基因编码m21蛋白。与fxpk蛋白相比,m21蛋白发生了如下5个氨基酸残基的突变:第6位氨基酸残基由i突变为t,第120位氨基酸残基由突变t为a,第231位氨基酸残基由e突变为k,第397位氨基酸残基由k突变为r,第676位氨基酸残基由d突变为g。重组质粒ptr-m41。与重组质粒ptr-fxpk相比,重组质粒ptr-m41的差异仅在于用m41d基因取代了fxpk基因。与fxpk基因相比,m41d基因发生了如下2个核苷酸的突变:第4位核苷酸由a突变为g,第2353位核苷酸由t突变为c。m41d基因编码m41蛋白。与fxpk蛋白相比,m41蛋白发生了如下2个氨基酸残基的突变:第2位氨基酸残基由t突变为a,第785位氨基酸残基由f突变为l。重组质粒ptr-m71。与重组质粒ptr-fxpk相比,重组质粒ptr-m71的差异仅在于用m71d基因取代了fxpk基因。与fxpk基因相比,m71d基因发生了如下3个核苷酸的突变:第40位核苷酸由a突变为g,第1481位核苷酸由g突变为a,第2401位核苷酸由t突变为c。m71d基因编码m71蛋白。与fxpk蛋白相比,m71蛋白发生了如下3个氨基酸残基的突变:第14位氨基酸残基n由突变为d,第494位氨基酸残基由r突变为h,第801位氨基酸残基由w突变为r。重组质粒ptr-m81。与重组质粒ptr-fxpk相比,重组质粒ptr-m81的差异仅在于用m81d基因取代了fxpk基因。与fxpk基因相比,m81d基因发生了如下4个核苷酸的突变:第60位核苷酸由a突变为t,第778位核苷酸由c突变为t,第1024位核苷酸由g突变为a,第1401位核苷酸由g突变为a。m81d基因编码m81蛋白。与fxpk蛋白相比,m81蛋白发生了如下4个氨基酸残基的突变:第20位氨基酸残基由e突变为d,第260位氨基酸残基由h突变为y,第342位氨基酸残基由e突变为k,第467位氨基酸残基m由突变为i。重组质粒ptr-m82。与重组质粒ptr-fxpk相比,重组质粒ptr-m82的差异仅在于用m82d基因取代了fxpk基因。与fxpk基因相比,m82d基因发生了如下3个核苷酸的突变:第60位核苷酸由a突变为t,第778位核苷酸由c突变为t,第1024位核苷酸由g突变为a。m82d基因编码m82蛋白。与fxpk蛋白相比,m82蛋白发生了如下3个氨基酸残基的突变:第20位氨基酸残基由e突变为d,第260位氨基酸残基由h突变为y,第342位氨基酸残基由e突变为k。重组质粒ptr-t2a。与重组质粒ptr-fxpk相比,重组质粒ptr-t2a的差异仅在于用t2a基因取代了fxpk基因。与fxpk基因相比,t2a基因发生了如下1个核苷酸的突变:第4位核苷酸由a突变为g。t2a基因编码t2a蛋白。与fxpk蛋白相比,t2a蛋白发生了如下1个氨基酸残基的突变:第2位氨基酸残基由t突变为a。重组质粒ptr-i6t。与重组质粒ptr-fxpk相比,重组质粒ptr-i6t的差异仅在于用i6t基因取代了fxpk基因。与fxpk基因相比,i6t基因发生了如下1个核苷酸的突变:第17位核苷酸由t突变为c。i6t基因编码i6t蛋白。与fxpk蛋白相比,i6t蛋白发生了如下1个氨基酸残基的突变:第6位氨基酸残基由i突变为t。重组质粒ptr-h260y。与重组质粒ptr-fxpk相比,重组质粒ptr-h260y的差异仅在于用h260y基因取代了fxpk基因。与fxpk基因相比,h260y基因发生了如下1个核苷酸的突变:第778位核苷酸由c突变为t。h260y基因编码h260y蛋白。与fxpk蛋白相比,h260y蛋白发生了如下1个氨基酸残基的突变:第260位氨基酸残基由h突变为y。重组质粒ptr-t2a/i6t。与重组质粒ptr-fxpk相比,重组质粒ptr-t2a/i6t的差异仅在于用t2a/i6t基因取代了fxpk基因。与fxpk基因相比,t2a/i6t基因发生了如下2个核苷酸的突变:第4位核苷酸由a突变为g,第17位核苷酸由t突变为c。t2a/i6t基因编码t2a/i6t蛋白。与fxpk蛋白相比,t2a/i6t蛋白发生了如下2个氨基酸残基的突变:第2位氨基酸残基由t突变为a,第6位氨基酸残基由i突变为t。重组质粒ptr-t2a/i6t/h260y。与重组质粒ptr-fxpk相比,重组质粒ptr-t2a/i6t/h260y的差异仅在于用t2a/i6t/h260y基因取代了fxpk基因。与fxpk基因相比,t2a/i6t/h260y基因发生了如下3个核苷酸的突变:第4位核苷酸由a突变为g,第17位核苷酸由t突变为c,第778位核苷酸由c突变为t。t2a/i6t/h260y基因编码t2a/i6t/h260y蛋白。与fxpk蛋白相比,t2a/i6t/h260y蛋白发生了如下3个氨基酸残基的突变:第2位氨基酸残基由t突变为a,第6位氨基酸残基由i突变为t,第260位氨基酸残基由h突变为y。二、制备重组质粒pet-fxpk。在质粒pet-30a(+)的ndei和xhoi酶切识别序列之间插入序列表的序列4中第4-2475位核苷酸所示的dna分子,得到重组质粒pet-fxpk。经测序验证,重组质粒pet-fxpk中具有特异dna分子乙。所述特异dna分子乙自上游至下游依次由如下五个元件组成:ndei酶切识别序列“catatg”(其中atg作为融合基因的起始密码子)、序列表的序列4中第4-2475位核苷酸所示的dna分子(即去除起始密码子和终止密码子的fxpk基因)、xhoi酶切识别序列“ctcgag”、his6标签编码序列“caccaccaccaccaccac”、终止密码子“tga”。重组质粒中的融合基因表达fxpk-his6蛋白。fxpk-his6蛋白自n端至c端依次由如下元件组成:fxpk蛋白、le(xhoi酶切识别序列编码)、his6标签。三、分别制备各个重组质粒,并测序验证。重组质粒pet-m21。与重组质粒pet-fxpk相比,重组质粒pet-m21的差异仅在于用去除终止密码子的m21d基因取代了去除终止密码子的fxpk基因。重组质粒中的融合基因表达m21-his6蛋白。m21-his6蛋白自n端至c端依次由如下元件组成:m21蛋白、le(xhoi酶切识别序列编码)、his6标签。重组质粒pet-m41。与重组质粒pet-fxpk相比,重组质粒pet-m41的差异仅在于用去除终止密码子的m41d基因取代了去除终止密码子的fxpk基因。重组质粒中的融合基因表达m41-his6蛋白。m41-his6蛋白自n端至c端依次由如下元件组成:m41蛋白、le(xhoi酶切识别序列编码)、his6标签。重组质粒pet-m71。与重组质粒pet-fxpk相比,重组质粒pet-m71的差异仅在于用去除终止密码子的m71d基因取代了去除终止密码子的fxpk基因。重组质粒中的融合基因表达m71-his6蛋白。m71-his6蛋白自n端至c端依次由如下元件组成:m71蛋白、le(xhoi酶切识别序列编码)、his6标签。重组质粒pet-m81。与重组质粒pet-fxpk相比,重组质粒pet-m81的差异仅在于用去除终止密码子的m81d基因取代了去除终止密码子的fxpk基因。重组质粒中的融合基因表达m81-his6蛋白。m81-his6蛋白自n端至c端依次由如下元件组成:m81蛋白、le(xhoi酶切识别序列编码)、his6标签。重组质粒pet-m82。与重组质粒pet-fxpk相比,重组质粒pet-m82的差异仅在于用去除终止密码子的m82d基因取代了去除终止密码子的fxpk基因。重组质粒中的融合基因表达m82-his6蛋白。m82-his6蛋白自n端至c端依次由如下元件组成:m82蛋白、le(xhoi酶切识别序列编码)、his6标签。重组质粒pet-t2a。与重组质粒pet-fxpk相比,重组质粒pet-t2a的差异仅在于用去除终止密码子的t2a基因取代了去除终止密码子的fxpk基因。重组质粒中的融合基因表达t2a-his6蛋白。t2a-his6蛋白自n端至c端依次由如下元件组成:t2a蛋白、le(xhoi酶切识别序列编码)、his6标签。重组质粒pet-i6t。与重组质粒pet-fxpk相比,重组质粒pet-i6t的差异仅在于用去除终止密码子的i6t基因取代了去除终止密码子的fxpk基因。重组质粒中的融合基因表达i6t-his6蛋白。i6t-his6蛋白自n端至c端依次由如下元件组成:i6t蛋白、le(xhoi酶切识别序列编码)、his6标签。重组质粒pet-n14d。与重组质粒pet-fxpk相比,重组质粒pet-n14d的差异仅在于用去除终止密码子的n14d基因取代了去除终止密码子的fxpk基因。与fxpk基因相比,n14d基因发生了如下1个核苷酸的突变:第40位核苷酸由a突变为g。n14d基因编码n14d蛋白。与fxpk蛋白相比,n14d蛋白发生了如下1个氨基酸残基的突变:第14位氨基酸残基n由突变为d。重组质粒中的融合基因表达n14d-his6蛋白。n14d-his6蛋白自n端至c端依次由如下元件组成:n14d蛋白、le(xhoi酶切识别序列编码)、his6标签。重组质粒pet-e20d。与重组质粒pet-fxpk相比,重组质粒pet-e20d的差异仅在于用去除终止密码子的e20d基因取代了去除终止密码子的fxpk基因。与fxpk基因相比,e20d基因发生了如下1个核苷酸的突变:第60位核苷酸由a突变为t。e20d基因编码e20d蛋白。与fxpk蛋白相比,e20d蛋白发生了如下1个氨基酸残基的突变:第20位氨基酸残基由e突变为d。重组质粒中的融合基因表达e20d-his6蛋白。e20d-his6蛋白自n端至c端依次由如下元件组成:e20d蛋白、le(xhoi酶切识别序列编码)、his6标签。重组质粒pet-t120a。与重组质粒pet-fxpk相比,重组质粒pet-t120a的差异仅在于用去除终止密码子的t120a基因取代了去除终止密码子的fxpk基因。与fxpk基因相比,t120a基因发生了如下1个核苷酸的突变:第358位核苷酸由a突变为g。t120a基因编码t120a蛋白。与fxpk蛋白相比,t120a蛋白发生了如下1个氨基酸残基的突变:第120位氨基酸残基由突变t为a。重组质粒中的融合基因表达t120a-his6蛋白。t120a-his6蛋白自n端至c端依次由如下元件组成:t120a蛋白、le(xhoi酶切识别序列编码)、his6标签。重组质粒pet-e231k。与重组质粒pet-fxpk相比,重组质粒pet-e231k的差异仅在于用去除终止密码子的e231k基因取代了去除终止密码子的fxpk基因。与fxpk基因相比,e231k基因发生了如下1个核苷酸的突变:第691位核苷酸由g突变为a。e231k基因编码e231k蛋白。与fxpk蛋白相比,e231k蛋白发生了如下1个氨基酸残基的突变:第231位氨基酸残基由e突变为k。重组质粒中的融合基因表达e231k-his6蛋白。e231k-his6蛋白自n端至c端依次由如下元件组成:e231k蛋白、le(xhoi酶切识别序列编码)、his6标签。重组质粒pet-h260y。与重组质粒pet-fxpk相比,重组质粒pet-h260y的差异仅在于用去除终止密码子的h260y基因取代了去除终止密码子的fxpk基因。重组质粒中的融合基因表达h260y-his6蛋白。h260y-his6蛋白自n端至c端依次由如下元件组成:h260y蛋白、le(xhoi酶切识别序列编码)、his6标签。重组质粒pet-e342k。与重组质粒pet-fxpk相比,重组质粒pet-e342k的差异仅在于用去除终止密码子的e342k基因取代了去除终止密码子的fxpk基因。与fxpk基因相比,e342k基因发生了如下1个核苷酸的突变:第1024位核苷酸由g突变为a。e342k基因编码e342k蛋白。与fxpk蛋白相比,e342k蛋白发生了如下1个氨基酸残基的突变:第342位氨基酸残基由e突变为k。重组质粒中的融合基因表达e342k-his6蛋白。e342k-his6蛋白自n端至c端依次由如下元件组成:e342k蛋白、le(xhoi酶切识别序列编码)、his6标签。重组质粒pet-k397r。与重组质粒pet-fxpk相比,重组质粒pet-k397r的差异仅在于用去除终止密码子的k397r基因取代了去除终止密码子的fxpk基因。与fxpk基因相比,k397r基因发生了如下1个核苷酸的突变:第1190位核苷酸由a突变为g。k397r基因编码k397r蛋白。与fxpk蛋白相比,k397r蛋白发生了如下1个氨基酸残基的突变:第397位氨基酸残基由k突变为r。重组质粒中的融合基因表达k397r-his6蛋白。k397r-his6蛋白自n端至c端依次由如下元件组成:k397r蛋白、le(xhoi酶切识别序列编码)、his6标签。重组质粒pet-d676g。与重组质粒pet-fxpk相比,重组质粒pet-d676g的差异仅在于用去除终止密码子的d676g基因取代了去除终止密码子的fxpk基因。与fxpk基因相比,d676g基因发生了如下1个核苷酸的突变:第2027位核苷酸由a突变为g。d676g基因编码d676g蛋白。与fxpk蛋白相比,d676g蛋白发生了如下1个氨基酸残基的突变:第676位氨基酸残基由d突变为g。重组质粒中的融合基因表达d676g-his6蛋白。d676g-his6蛋白自n端至c端依次由如下元件组成:d676g蛋白、le(xhoi酶切识别序列编码)、his6标签。重组质粒pet-f785l。与重组质粒pet-fxpk相比,重组质粒pet-f785l的差异仅在于用去除终止密码子的f785l基因取代了去除终止密码子的fxpk基因。与fxpk基因相比,f785l基因发生了如下1个核苷酸的突变:第2353位核苷酸由t突变为c。f785l基因编码f785l蛋白。与fxpk蛋白相比,f785l蛋白发生了如下1个氨基酸残基的突变:第785位氨基酸残基由f突变为l。重组质粒中的融合基因表达f785l-his6蛋白。f785l-his6蛋白自n端至c端依次由如下元件组成:f785l蛋白、le(xhoi酶切识别序列编码)、his6标签。重组质粒pet-w801r。与重组质粒pet-fxpk相比,重组质粒pet-w801r的差异仅在于用去除终止密码子的w801r基因取代了去除终止密码子的fxpk基因。与fxpk基因相比,w801r基因发生了如下1个核苷酸的突变:第2401位核苷酸由t突变为c。w801r基因编码w801r蛋白。与fxpk蛋白相比,w801r蛋白发生了如下1个氨基酸残基的突变:第801位氨基酸残基由w突变为r。重组质粒中的融合基因表达w801r-his6蛋白。w801r-his6蛋白自n端至c端依次由如下元件组成:w801r蛋白、le(xhoi酶切识别序列编码)、his6标签。重组质粒pet-t2a/i6t。与重组质粒pet-fxpk相比,重组质粒pet-t2a/i6t的差异仅在于用t2a/i6t基因取代了fxpk基因。重组质粒中的融合基因表达t2a/i6t-his6蛋白。t2a/i6t-his6蛋白自n端至c端依次由如下元件组成:t2a/i6t蛋白、le(xhoi酶切识别序列编码)、his6标签。重组质粒pet-t2a/h260y。与重组质粒pet-fxpk相比,重组质粒pet-t2a/h260y的差异仅在于用去除终止密码子的t2a/h260y基因取代了去除终止密码子的fxpk基因。与fxpk基因相比,t2a/h260y基因发生了如下2个核苷酸的突变:第4位核苷酸由a突变为g,第778位核苷酸由c突变为t。t2a/h260y基因编码t2a/h260y蛋白。与fxpk蛋白相比,t2a/h260y蛋白发生了如下2个氨基酸残基的突变:第2位氨基酸残基由t突变为a,第260位氨基酸残基由h突变为y。重组质粒中的融合基因表达t2a/h260y-his6蛋白。t2a/h260y-his6蛋白自n端至c端依次由如下元件组成:t2a/h260y蛋白、le(xhoi酶切识别序列编码)、his6标签。重组质粒pet-i6t/h260y。与重组质粒pet-fxpk相比,重组质粒pet-i6t/h260y的差异仅在于用去除终止密码子的i6t/h260y基因取代了去除终止密码子的fxpk基因。与fxpk基因相比,i6t/h260y基因发生了如下2个核苷酸的突变:第17位核苷酸由t突变为c,第778位核苷酸由c突变为t。i6t/h260y基因编码i6t/h260y蛋白。与fxpk蛋白相比,i6t/h260y蛋白发生了如下2个氨基酸残基的突变:第6位氨基酸残基由i突变为t,第260位氨基酸残基由h突变为y。重组质粒中的融合基因表达i6t/h260y-his6蛋白。i6t/h260y-his6蛋白自n端至c端依次由如下元件组成:i6t/h260y蛋白、le(xhoi酶切识别序列编码)、his6标签。重组质粒pet-t2a/i6t/h260y。与重组质粒pet-fxpk相比,重组质粒pet-t2a/i6t/h260y的差异仅在于用t2a/i6t/h260y基因取代了fxpk基因。重组质粒中的融合基因表达t2a/i6t/h260y-his6蛋白。t2a/i6t/h260y-his6蛋白自n端至c端依次由如下元件组成:t2a/i6t/h260y蛋白、le(xhoi酶切识别序列编码)、his6标签。实施例5、各个蛋白质的制备及其作为磷酸转酮酶的酶活测定将重组质粒导入大肠杆菌bl21(de3),得到重组菌。采用液体lb培养基培养重组菌,37℃、200rpm振荡培养至od600nm=0.6-0.8,然后加入iptg并使其在体系中的浓度为0.4mm,然后20℃、200rpm振荡培养18小时,然后4℃、7000g离心5min收集菌体。洗涤菌体,然后进行超声破碎,然后收集上清液,利用hisspintrapcolumns(购自ge公司,产品货号28-4013-53)纯化具有his6标签的蛋白质,然后将蛋白质的缓冲体系置换为ph7.2的pbs缓冲液。重组质粒pet-fxpk进行上述步骤制备得到fxpk-his6蛋白。重组质粒pet-m21进行上述步骤制备得到m21-his6蛋白。重组质粒pet-m41进行上述步骤制备得到m41-his6蛋白。重组质粒pet-m71进行上述步骤制备得到m71-his6蛋白。重组质粒pet-m81进行上述步骤制备得到m81-his6蛋白。重组质粒pet-m82进行上述步骤制备得到m82-his6蛋白。重组质粒pet-t2a进行上述步骤制备得到t2a-his6蛋白。重组质粒pet-i6t进行上述步骤制备得到i6t-his6蛋白。重组质粒pet-n14d进行上述步骤制备得到n14d-his6蛋白。重组质粒pet-e20d进行上述步骤制备得到e20d-his6蛋白。重组质粒pet-t120a进行上述步骤制备得到与t120a-his6蛋白。重组质粒pet-e231k进行上述步骤制备得到e231k-his6蛋白。重组质粒pet-h260y进行上述步骤制备得到h260y-his6蛋白。重组质粒pet-e342k进行上述步骤制备得到e342k-his6蛋白。重组质粒pet-k397r进行上述步骤制备得到k397r-his6蛋白。重组质粒pet-d676g进行上述步骤制备得到d676g-his6蛋白。重组质粒pet-f785l进行上述步骤制备得到f785l-his6蛋白。重组质粒pet-w801r进行上述步骤制备得到w801r-his6蛋白。重组质粒pet-t2a/i6t/i6t进行上述步骤制备得到t2a/i6t-his6蛋白。重组质粒pet-t2a/h260y进行上述步骤制备得到t2a/h260y-his6蛋白。重组质粒pet-i6t/h260y进行上述步骤制备得到i6t/h260y-his6蛋白。重组质粒pet-t2a/i6t/h260y/i6t/h260y进行上述步骤制备得到t2a/i6t/h260y-his6蛋白。利用thermofisher公司的bca蛋白定量试剂盒进行蛋白定量。分别测定上述制备的各个具有his6标签的蛋白质对6-磷酸果糖的反应活性。反应原理:底物6-磷酸果糖被供试蛋白催化生成的乙酰磷酸,乙酰磷酸与羟胺生成羟肟酸,羟肟酸与氯化铁反应生成红色化合物,能够在505nm下被检测到。反应体系:49μl磷酸盐缓冲液(ph6.5、100mm),1μl10mm焦磷酸硫胺素水溶液,0.1μl100mmmgcl2水溶液,0.1μl0.7m半胱氨酸盐酸盐水溶液,30μl100mm果糖-6-磷酸水溶液,20μl供试蛋白溶液。先将反应体系置于30℃水浴中反应10分钟;然后加入80μl2mol/l羟胺水溶液,混匀后室温静置10分钟;然后加入55μl15g/100ml三氯乙酸水溶液、55μl4mol/l盐酸水溶液,55μl5g/100mlfecl3·6h2o水溶液,然后室温静置反应1分钟;然后取样200μl,检测505nm下的吸光值(od505nm值)。一个酶活单位(u)定义为:在1分钟的反应时间内,每生成1μmol的乙酰磷酸所需要的酶。酶活(u)除以蛋白量(mg),得到比酶活(u/mg)。上述制备的各个具有his6标签的蛋白质作为磷酸转酮酶的比酶活见图3。图3中,fxpk代表fxpk-his6蛋白,m21代表m21-his6蛋白,m41代表m41-his6蛋白,以此类推。各个突变体蛋白作为磷酸转酮酶的酶活均高于野生型的fxpk蛋白,t2a/i6t蛋白和t2a/i6t/h260y蛋白的酶活最高。实施例6、表达各个蛋白质的重组菌产谷氨酸的能力比较分别将重组质粒导入谷氨酸棒杆菌z188,得到各个重组菌。重组质粒分别为如下:重组质粒ptr-fxpk、重组质粒ptr-m21、重组质粒ptr-m41、重组质粒ptr-m71、重组质粒ptr-m81、重组质粒ptr-m82、重组质粒ptr-t2a、重组质粒ptr-i6t、重组质粒ptr-h260y、重组质粒ptr-t2a/i6t、重组质粒ptr-t2a/i6t/h260y。检测供试菌株发酵生产谷氨酸的能力。供试菌株分别为如下:谷氨酸棒杆菌z188以及上述制备的各个重组菌。种子培养基:葡萄糖50g/l,磷酸0.7g/l,七水硫酸镁0.8g/l,硫酸铵10g/l,3-(n-玛琳代)丙磺酸84g/l,玉米粉3g/l,尿素10g/l,蛋白胨1g/l,酵母粉0.5g/l,余量为水,用氢氧化钠调ph为7.0。发酵培养基与种子培养基的区别仅在于不加入蛋白胨和酵母粉。取供试菌株的单克隆,接种至5ml种子培养基中,30℃、850rpm振荡培养12小时,得到种子液。取96孔板,每孔加入150μl发酵培养基,然后以10%的接种量接种种子液,然后30℃、850rpm振荡培养33小时,然后利用sba-40d生物传感分析仪检测谷氨酸产量(每升体系中的谷氨酸含量)和葡萄糖消耗,计算从葡萄糖到谷氨酸的转化率。结果见图4。可以看出,相比野生型的fxpk蛋白,所测试的fxpk突变体蛋白都能够增加谷氨酸的产量和转化率。实施例7、各个蛋白质对大肠杆菌发酵生产琥珀酸的影响利用实施例4的步骤一中构建的各个重组质粒以及重组质粒ptr-fxpk,转化大肠杆菌琥珀酸生产菌株(cgmccno.5107、cgmccno.5108或cgmccno.5109,中国专利zl201110264353.9),以获得携带野生型fxpk基因及不同fxpk突变体基因的琥珀酸生产菌株。利用中国专利zl201110264353.9中所述的琥珀酸生产的发酵条件对菌株生产琥珀酸的能力进行测试。与野生型的fxpk相比,各个fxpk突变体蛋白酶活显著提高。与携带野生型fxpk基因的平行菌株相比,携带各个fxpk突变体基因的菌株生产琥珀酸的能力显著提高,糖酸转化率显著提高。实施例8、各个蛋白质对谷氨酸棒杆菌发酵生产谷氨酰胺的影响在谷氨酸生产菌株z188的基础上,根据文献报告,将其谷氨酰胺合成酶进行y405f突变,能够生产谷氨酰胺(liu,q.,etal.2008.applmicrobiolbiotechnol77(6):1297-1304.)。利用实施例4的步骤一中构建的各个重组质粒以及重组质粒ptr-fxpk,转化到构建的谷氨酰胺生产菌株中,利用文献(liu,q.,etal.2008.applmicrobiolbiotechnol77(6):1297-1304.)所述的谷氨酰胺发酵条件对菌株生产谷氨酰胺的能力进行测试。与野生型的fxpk相比,各个fxpk突变体蛋白酶活显著提高。与携带野生型fxpk基因的平行菌株相比,携带各个fxpk突变体基因的菌株生产谷氨酰胺的能力显著提高,糖酸转化率显著提高。实施例9、各个蛋白质对大肠杆菌发酵生产脯氨酸的影响大肠杆菌脯氨酸生产菌株dh5α(psw2)是携带有质粒psw2的大肠杆菌dh5α,其中质粒psw2通过将谷氨酸激酶prob74【prob(ncbi-gi:16128228)第107位asp突变为asn】的基因以及谷氨酸半醛脱氢酶proa(ncbi-gi:16128229)的基因连接到质粒puc19上构建而成(中国专利2014107400221中的实施例5)。利用实施例4的步骤一中构建的各个重组质粒以及重组质粒ptr-fxpk,转化大肠杆菌脯氨酸生产菌株dh5α(psw2)。利用中国专利2014107400221实施例5中所述的发酵条件对菌株生产脯氨酸的能力进行测试。与野生型的fxpk相比,各个fxpk突变体蛋白酶活显著提高。与携带野生型fxpk基因的平行菌株相比,携带各个fxpk突变体基因的菌株生产脯氨酸的能力显著提高,糖酸转化率显著提高。实施例10、各个蛋白质对大肠杆菌发酵生产反式-4-羟基-l-脯氨酸的影响大肠杆菌反式-4-羟基-l-脯氨酸生产菌株dh5α(psw3)是携带有质粒psw3的大肠杆菌dh5α,其中质粒psw3是在专利中质粒psw2的基础上进一步连接l-脯氨酸-4-羟基化酶构建而成的(中国专利2014107400221)。利用实施例4的步骤一中构建的各个重组质粒以及重组质粒ptr-fxpk,转化大肠杆菌反式-4-羟基-l-脯氨酸生产菌株dh5α(psw3)。利用中国专利2014107400221实施例5中所述的发酵条件对菌株生产反式-4-羟基-l-脯氨酸的能力进行测试。与野生型的fxpk相比,各个fxpk突变体蛋白酶活显著提高。与携带野生型fxpk基因的平行菌株相比,携带各个fxpk突变体基因的菌株生产反式-4-羟基-l-脯氨酸的能力显著提高,糖酸转化率显著提高。实施例11、各个蛋白质对黑曲霉发酵生产柠檬酸的影响以重组质粒ptr-fxpk为模板,采用fxpk-fm(fxpk-fm:attctcgagatgacctctccggttatcg)和fxpk-rm(fxpk-rm:aatgcatgctcattcgttgtcacccg)组成的引物对,扩增野生型的fxpk基因,同时分别在5′端与3′端添加xhoi与sphi的酶切位点。然后通过xhoi与sphi双酶切将基因插入与黑曲霉表达质粒psilent-1(genbankid:ab303070),获得以ptrpc为启动子、以ttrpc为终止子的fxpk基因的黑曲霉表达质粒psil-fxpk。实施例4的步骤一中构建的各个重组质粒,参照上述方法,制备突变体基因的黑曲霉表达质粒。将质粒分别转化至黑曲霉co827(cicc40347)的原生质体中。利用中国专利201710022533.3实施例中所述的发酵条件对重组黑曲霉生产柠檬酸的能力进行测试。与野生型的fxpk相比,各个fxpk突变体蛋白酶活显著提高。与携带野生型fxpk基因的平行菌株相比,携带各个fxpk突变体基因的菌株生产柠檬酸的能力显著提高,糖酸转化率显著提高。sequencelisting<110>中国科学院天津工业生物技术研究所<120>活性提高的磷酸转酮酶及在生产代谢物中的应用<130>gncyx190964<160>6<170>patentinversion3.5<210>1<211>346<212>prt<213>corynebacteriumglutamicum<400>1metgluaspmetargilealathrleuthrserglyglyaspcyspro151015glyleuasnalavalileargglyilevalargthralaserasnglu202530pheglyserthrvalvalglytyrglnaspglytrpgluglyleuleu354045alaaspargargvalglnleutyraspaspgluaspileaspargile505560leuleuargglyglythrileleuglythrglyargleuhisproasp65707580lysphelysalaglyileaspglnilelysalaasnleugluaspala859095glyileaspalaleuileproileglyglygluglythrleulysgly100105110alalystrpleuseraspasnglyileprovalvalglyvalprolys115120125thrileaspasnaspvalasnglythraspphethrpheglypheasp130135140thralavalalavalalathraspalavalaspargleuhisthrthr145150155160alagluserhisasnargvalmetilevalgluvalmetglyarghis165170175valglytrpilealaleuhisalaglymetalaglyglyalahistyr180185190thrvalileprogluvalpropheaspilealagluilecyslysala195200205metgluargargpheglnmetglyglulystyrglyileilevalval210215220alagluglyalaleuproarggluglythrmetgluleuargglugly225230235240hisileaspglnpheglyhislysthrphethrglyileglyglngln245250255ilealaaspgluilehisalaargleuglyhisaspvalargthrthr260265270valleuglyhisileglnargglyglythrprothralapheasparg275280285valleualathrargtyrglyvalargalaalaargalacyshisglu290295300glyserpheasplysvalvalalaleulysglygluargileglumet305310315320ilethrphegluaspalavalglythrleulysgluvalpropheglu325330335argtrpvalthralaglnalametphegly340345<210>2<211>3041<212>dna<213>corynebacteriumglutamicum<400>2accgaggaccagtggcaggcctccaagcgggagttcaatgagcaaaatgaggcagaagaa60ctcgtgctcagcaaggcataagggcaaggggttctagaaagaccaacggacgtgttttcc120cagctcccattggtttaaaaagctcatttcgaggcctgaacgtgcgaatcaggacaaaac180tcgagtagcccggaatgattttggtttccttctgcgagttcgccatgtggctgcggtaaa240actgccccggaaccggaatatctcgacgccacagaacgccatttgcgggccttaacaccc300cgtgggcataccttttaccgattccagatattcggtcgcttatattgcctagtgtgattc360caaacagaaaactggggcgacctctaataagagtcgccccgataagttttttaccgtaat420tattactgggagtcagatactgcgtaagcaatcgcagcagcgccagcggtcacagtaaga480actgcaggccacgcgccaatcttcttggcaagtgggtgggacaggccaaatgcaccaacg540taggttgtcagcaggccagtagctactgcaggacccttcttttcattccagcttcgtgca600gcaagcgctccggatgctgccaatggaatggtgcccagtgggcgaatgccggattcacgg660gcagtcaaccaaccgccgatcaaacctgctgcgacgacggtggcagtgctgacctgggat720gcctttttcaatttcatttccatggtgagccagtctagagacaaaatttttccgcggggt780tttcttgatctgatccgacaagccaatgggggaaaaaatgtgtccgaccaaaaattgtgc840agcacaccacatgcccgctcggacaatgtcgatttgttaatgaaactgcagctctggcga900ttaaataaggtggtcagagacaatttttcggccggtcaacccctgtgattttcttatttt960tgggtgattgttccggcgcgggtgttgtgatgggtttaatatggaagacatgcgaattgc1020tactctcacgtcaggcggcgactgccccggactaaacgccgtcatccgaggaatcgtccg1080cacagccagcaatgaatttggctccaccgtcgttggttatcaagacggttgggaaggact1140gttagccgatcgtcgcgtacagctgtatgacgatgaagatattgaccgaatcctccttcg1200aggcggcaccattttgggcactggtcgcctccatccggacaagtttaaggccggaattga1260tcagattaaggccaacttagaagacgccggcatcgatgcccttattccaatcggtggcga1320aggaaccctgaagggtgccaagtggctgtctgataacggtatccctgttgtcggtgtccc1380aaagaccattgacaatgacgtgaatggcactgacttcaccttcggtttcgatactgctgt1440tgcagtggctaccgacgctgttgaccgcctgcacaccaccgctgaatctcacaaccgtgt1500gatgattgtggaggtcatgggccgccacgtgggttggattgctctgcacgcaggtatggc1560cggcggtgctcactacaccgttatcccagaagttcctttcgatattgcagagatctgcaa1620ggcgatggaacgtcgcttccagatgggcgagaagtacggcattatcgtcgttgcggaagg1680tgcattgccacgcgaaggcaccatggaacttcgtgaaggccacattgaccagttcggcca1740caagaccttcaccggaatcggccagcagattgctgatgaaatccatgcgcgcctcggtca1800cgatgttcgtaccaccgttcttggccacattcagcgtggtggaaccccgactgctttcga1860ccgtgttctggccactcgttatggtgttcgggcagctcgtgcatgccatgagggaagctt1920tgacaaggttgttgccttgaagggtgagcgcatcgagatgatcacctttgaagatgccgt1980cggaaccttgaaggaagttccattcgagcgctgggttactgcccaggcaatgtttggata2040gtttttcgggcttttatcaacagccaataacagctctttcgcccattgaggtggaggggc2100tgttttttcatgccgtaaggaaggtgcaagtaagtgaaatcaagtggcatagatccattg2160atgcttagactgtgacctaggcttgactttcgtgggggagtggggataagttcatcttaa2220acacaatgcaatcgattgcatttacattccttatcccacaataggggtaccttccagaaa2280gttggtgaggagatggcttccgaaacctccagcccgaagaagcgggccaccacactcaaa2340gacatcgcgcaaacaacacagctttcagtcagcacggtgtcccgggcattggccaacaac2400gcgagcattccggaatccacacgcatccgagtggttgaagccgctcaaaagctgaactac2460cgtcccaatgcccaagctcgtgcattgcggaagtcgaggacagacaccatcggtgtcatc2520attccaaacattgagaacccatatttctcctcactagcagcatcgattcaaaaagctgct2580cgtgaagctggggtgtccaccattttgtccaactctgaagaaaacccagagctgcttggt2640cagactttggcgatcatggatgaccaacgcctcgatggaatcatcgtggtgccacacatt2700cagtcagaggaacaagtcactgacttggttaccaggggagtgccagtagtgctggcagac2760cgtagttttgttaactcgtctattccttcggttacctcagatccagttccgggcatgact2820gaagctgtggacttactcctggcagctgacgtgcaattgggctaccttgccggcccgcag2880gatacttccactggtcagctgcgtcttaacacttttgaaagactatgcgtggaccgcggc2940atcgtcggagcatctgtctattacggtggctaccgccaagaatctggatatgacggcatc3000aaggtgctgatcaaacagggagccaatgcgattatcgctgg3041<210>3<211>825<212>prt<213>bifidobacterium<400>3metthrserprovalileglythrprotrplyslysleuasnalapro151015valsergluglualailegluglyvalasplystyrtrpargalaala202530asntyrleuserileglyglniletyrleuargserasnproleumet354045lysgluprophethrarggluaspvallyshisargleuvalglyhis505560trpglythrthrproglyleuasnpheleuileglyhisileasnarg65707580leuilealaasphisglnglnasnthrvalileilemetglyprogly859095hisglyglyproalaglythralaglnsertyrleuaspglythrtyr100105110thrglutyrpheproasnilethrlysaspglualaglyleuglnlys115120125phepheargglnphesertyrproglyglyileproserhistyrala130135140progluthrproglyserilehisgluglyglygluleuglytyrala145150155160leuserhisalatyrglyalavalmetasnasnproserleupheval165170175proalailevalglyaspglyglualagluthrglyproleualathr180185190glytrpglnserasnlysleuileasnproargthraspglyileval195200205leuproileleuhisleuasnglytyrlysilealaasnprothrile210215220leuserargileseraspglugluleuhisgluphephehisglymet225230235240glytyrgluprotyrgluphevalalaglypheaspasngluasphis245250255leuserilehisargargphealagluleuphegluthrvalpheasp260265270gluilecysaspilelysalaalaalaglnthraspaspmetthrarg275280285prophetyrprometileilepheargthrprolysglytrpthrcys290295300prolyspheileaspglylyslysthrgluglysertrpargserhis305310315320glnvalproleualaseralaargaspthrglualahisphegluval325330335leulysasntrpleuglusertyrlysproglugluleupheaspglu340345350asnglyalavallysprogluvalthralaphemetprothrglyglu355360365leuargileglygluasnproasnalaasnglyglyargileargglu370375380gluleulysleuprolysleugluasptyrgluvallysgluvalala385390395400glutyrglyhisglytrpglyglnleuglualathrargargleugly405410415valtyrthrargaspileilelysasnasnproaspserpheargile420425430pheglyproaspgluthralaserasnargleuglnalaalatyrasp435440445valthrasnlysglntrpaspalaglytyrleuseralaglnvalasp450455460gluhismetalavalthrglyglnvalthrgluglnleusergluhis465470475480glnmetgluglypheleugluglytyrleuleuthrglyarghisgly485490495iletrpsersertyrgluserphevalhisvalileaspsermetleu500505510asnglnhisalalystrpleuglualathrvalarggluileprotrp515520525arglysproilesersermetasnleuleuvalserserhisvaltrp530535540argglnasphisasnglypheserhisglnaspproglyvalthrser545550555560valleuleuasnlyscyspheasnasnasphisvalileglyiletyr565570575pheprovalaspserasnmetleuleualavalalaglulyscystyr580585590lysserthrasnlysileasnalaileilealaglylysglnproala595600605alathrtrpleuthrleuaspglualaargalagluleuglulysgly610615620alaalaglutrplystrpalaserasnvallysserasnaspgluala625630635640glnilevalleualaalathrglyaspvalprothrglngluilemet645650655alaalaalaasplysleuaspalametglyilelysphelysvalval660665670asnvalvalaspleuvallysleuglnseralalysgluasnasnglu675680685alaleuseraspglugluphealagluleuphethrgluasplyspro690695700valleuphealatyrhissertyralaargaspvalargglyleuile705710715720tyraspargproasnhisaspasnpheasnvalhisglytyrgluglu725730735glnglyserthrthrthrprotyraspmetvalargvalasnasnile740745750aspargtyrgluleuglnalaglualaleuargmetileaspalaasp755760765lystyralaasplysileasngluleuglualapheargglngluala770775780pheglnphealavalaspasnglytyrasphisproasptyrthrasp785790795800trpvaltyrserglyvalasnthrasnlysglnglyalaileserala805810815thralaalathralaglyaspasnglu820825<210>4<211>2478<212>dna<213>bifidobacterium<400>4atgacctctccggttatcggtaccccgtggaaaaaactgaacgcgccggtttctgaagaa60gcgatcgaaggtgttgacaaatactggcgtgcggcgaactacctgtctatcggtcagatc120tacctgcgttctaacccgctgatgaaagaaccgttcacccgtgaagacgttaaacaccgt180ctggttggtcactggggtaccaccccgggtctgaacttcctgatcggtcacatcaaccgt240ctgatcgcggaccaccagcagaacaccgttatcatcatgggtccgggtcacggtggtccg300gcgggtaccgcgcagtcttacctggacggtacctacaccgaatacttcccgaacatcacc360aaagacgaagcgggtctgcagaaattcttccgtcagttctcttacccgggtggtatcccg420tctcactacgcgccggaaaccccgggttctatccacgaaggtggtgaactgggttacgcg480ctgtctcacgcgtacggtgcggttatgaacaacccgtctctgttcgttccggcgatcgtt540ggtgacggtgaagcggaaaccggtccgctggcgaccggttggcagtctaacaaactgatc600aacccgcgtaccgacggtatcgttctgccgatcctgcacctgaacggttacaaaatcgcg660aacccgaccatcctgtctcgtatctctgacgaagaactgcacgagttcttccacggtatg720ggttacgaaccgtacgagttcgttgcgggtttcgacaacgaagaccacctgtctatccac780cgtcgtttcgcggaactgttcgaaaccgttttcgacgaaatctgcgacatcaaagcggcg840gcgcagaccgacgacatgacccgtccgttctacccgatgatcatcttccgtaccccgaaa900ggttggacctgcccgaaattcatcgacggtaaaaaaaccgaaggttcttggcgttctcac960caggttccgctggcgtctgcgcgtgacaccgaagcgcacttcgaagttctgaaaaactgg1020ctggaatcttacaaaccggaagaactgttcgacgaaaacggtgcggttaaaccggaagtt1080accgcgttcatgccgaccggtgaactgcgtatcggtgaaaacccgaacgcgaacggtggt1140cgtatccgtgaagaactgaaactgccgaaactggaagactacgaagttaaagaagttgcg1200gaatacggtcacggttggggtcagctggaagcgacccgtcgtctgggtgtttacacccgt1260gacatcatcaaaaacaacccggactctttccgtatcttcggtccggacgaaaccgcgtct1320aaccgtctgcaggcggcgtacgacgttaccaacaaacagtgggacgcgggttacctgtct1380gcgcaggttgacgaacacatggcggttaccggtcaggttaccgaacagctgtctgaacac1440cagatggaaggtttcctggaaggttacctgctgaccggtcgtcacggtatctggtcttct1500tacgaatctttcgttcacgttatcgactctatgctgaaccagcacgcgaaatggctggaa1560gcgaccgttcgtgaaatcccgtggcgtaaaccgatctcttctatgaacctgctggtttct1620tctcacgtttggcgtcaggaccacaacggtttctctcaccaggacccgggtgttacctct1680gttctgctgaacaaatgcttcaacaacgaccacgttatcggtatctacttcccggttgac1740tctaacatgctgctggcggttgcggaaaaatgctacaaatctaccaacaaaatcaacgcg1800atcatcgcgggtaaacagccggcggcgacctggctgaccctggacgaagcgcgtgcggaa1860ctggaaaaaggtgcggcggaatggaaatgggcgtctaacgttaaatctaacgacgaagcg1920cagatcgttctggcggcgaccggtgacgttccgacccaggaaatcatggcggcggcggac1980aaactggacgcgatgggtatcaaattcaaagttgttaacgttgttgacctggttaaactg2040cagtctgcgaaagaaaacaacgaagcgctgtctgacgaagagttcgcggaactgttcacc2100gaagacaaaccggttctgttcgcgtaccactcttacgcgcgtgacgttcgtggtctgatc2160tacgaccgtccgaaccacgacaacttcaacgttcacggttacgaagaacagggttctacc2220accaccccgtacgacatggttcgtgttaacaacatcgaccgttacgaactgcaggcggaa2280gcgctgcgtatgatcgacgcggacaaatacgcggacaaaatcaacgaactggaagcgttc2340cgtcaggaagcgttccagttcgcggttgacaacggttacgaccacccggactacaccgac2400tgggtttactctggtgttaacaccaacaaacagggtgcgatctctgcgaccgcggcgacc2460gcgggtgacaacgaatga2478<210>5<211>252<212>dna<213>artificialsequence<400>5cgactgcacggtgcaccaatgcttctggcgtcaggcagccatcggaagctgtggtatggc60tgtgcaggtcgtaaatcactgcataattcgtgtcgctcaaggcgcactcccgttctggat120aatgttttttgcgccgacatcataacggttctggcaaatattctgaaatgagctgttgac180aattaatcatccggctcgtataatgtgtggaattgtgagcggataacaatttcacacagg240aaacagaccatg252<210>6<211>252<212>dna<213>artificialsequence<400>6cgactgcacggtgcaccaatgcttctggcgtcaggcagccatcggaagctgtggtatggc60tgtgcaggtcgtaaatcactgcataattcgtgtcgctcaaggcgcactcccgttctggat120aatgttttttgcgccgacatcataacggttctggcaaatattctgaaatgagctgttgac180aattaatcatccggctcgtataatgtgtggaattgtgagcggataacaatctcacacagg240aaacagaccatg252当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1