CARNK细胞的制作方法
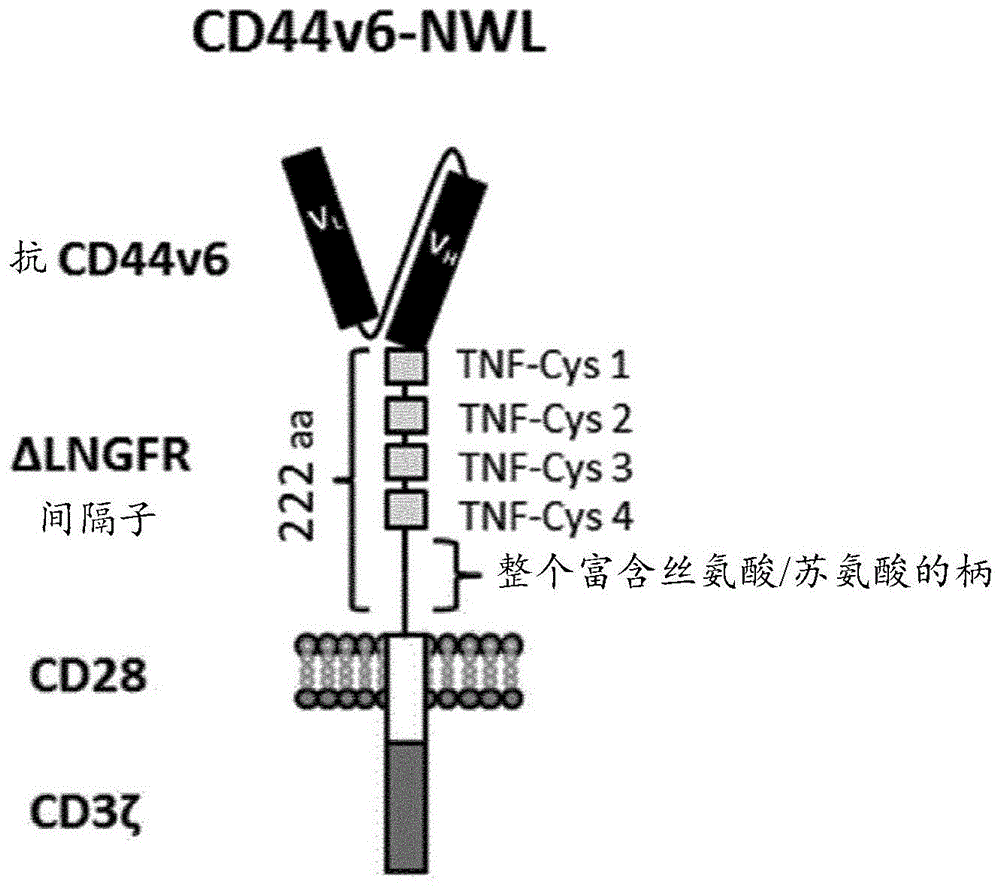
本发明涉及制造用慢病毒载体遗传修饰的自然杀伤(nk)细胞的领域,这些慢病毒载体携带编码嵌合抗原受体(car)的多核苷酸。本发明进一步涉及car-nk细胞,和这些car-nk细胞在医学中、特别是用于治疗癌症中的用途。
背景技术:
:本发明集中于开发自然杀伤细胞的制造方法,这些自然杀伤细胞是用携带至少一种编码car的多核苷酸的慢病毒载体遗传修饰的自然杀伤细胞(car-nk细胞)。本发明的制造方法允许获得以nk细胞和car的治疗效果之间的有效协同作用为特征的细胞群体。自然杀伤细胞(nk细胞)是具有高抗肿瘤、抗病毒和抗微生物活性的先天性免疫系统的细胞。在报道了nk细胞在患有癌症的患者中的成功过继性转移和体内扩增后,将nk细胞用于治疗癌症引起了关注[ruggeri等人(2005)curropinimmunol[免疫学当前观点]17:211-7;ren等人(2007)cancerbiotherradiopharm[癌症生物治疗和放射药物]22:223-34;koehl等人(2004)bloodcellsmoldis[血细胞分子和疾病]33:261-6.176;passweg等人(2004)leukaemia[白血病]18:1835-8]。通常,在这些研究中,供体nk细胞输注的耐受性良好且没有诱导gvhd的证据。然而,迄今为止,在患有癌症的患者中,只进行了少数研究过继性nk细胞输注的试验。主要障碍是仅相对少量nk细胞可以从常规白细胞去除术产物分离。这妨碍了患有癌症的人中nk细胞剂量依赖性抗肿瘤应答的临床试验[klingemann等人(2004)cytotherapy[细胞疗法]6:15-22;passweg等人(2006)bestpractresclinhaematol[临床血液学最佳实践和研究]19:811-824;mckenna等人(2007)transfusion[输液]47:520-528;koehl等人(2005)klinpadiatr[小儿科杂志]217:345-350;iyengar等人(2003)cytotherapy[细胞疗法]5:479-484;meyer-monard等人(2009)transfusion[输液]49:362-371]。因此,正在研究的用于扩增和激活nk细胞的离体方案,使得能够实现更高nk细胞剂量的临床试验,并且允许多次nk细胞输注[carlens等人(2001)humimmunol[人类免疫学]62:1092-1098;barkholt等人(2009)immunotherapy[免疫疗法]1:753-764;berg等人(2009)cytotherapy[细胞疗法]11:341-355;fujisaki等人(2009)cancerres[癌症研究]69:4010-4017;siegler等人(2010)cytotherapy[细胞疗法]12(6):750-63]。然而,大多数方案通过使用支持性饲养细胞系来处理技术缺点,这些支持性饲养细胞系对于大规模和多中心试验而言会导致产生nk细胞产物的调控问题。先前,我们已经描述了具有以高细胞数目、高纯度和功能性从cd34+造血干细胞产生临床相关的nk细胞产物的能力的替代性基于细胞因子的培养方法[spanholtz等人(2010)plosone[公共科学图书馆综合]5:e9221]。我们已经进一步优化了cd34+的富集,并且开发了导致高产率激活的cd34+细胞来源的nk细胞的可扩展程序。嵌合抗原受体(car)是识别在靶细胞上表达的特定蛋白质或抗原的重组受体。一旦car在t淋巴细胞中表达(则被称为car-t细胞)或免疫系统的其他细胞中表达,就能够针对表达其结合的抗原的所有细胞重定向特异性免疫应答。car研究最广泛的临床应用是癌症免疫疗法,该癌症免疫疗法包括输注携带靶向肿瘤抗原的car的免疫系统的细胞,例如t细胞或nk细胞。此类细胞能够针对表达由car靶向的抗原的细胞产生强抗肿瘤应答(sadelain等人,cancerdiscovery[癌症发现].2013.3(4):388-98)。car是由负责抗原识别的胞外结构域(通常源自单链可变片段(scfv))、间隔区、跨膜结构域、以及传递激活和共刺激信号至表达它们的细胞的胞内结构域组成的重组嵌合蛋白。取决于信号传导结构域的数量,将car分为第1代(一个)、第2代(两个)、或第3代(三个)car(dotti等人,immunolrev.[免疫学综述]2014年1月;257(1))。“间隔区”或“绞链区”是胞外结构域和跨膜结构域之间的连接序列。用作间隔子的最常见序列是恒定免疫球蛋白igg1铰链-ch2-ch3fc结构域。wo2016/042461披露了包含源自人低亲和力神经生长因子受体(lngfr)的胞外结构域的间隔区的car。现在有若干种car-t细胞候选物正在临床开发中,并且出现了不同的成功案例。美国食品药品监督管理局(fda)最近批准了替萨根微核(tisagenlecleucel)(第一种cart细胞疗法,适用于治疗上至25岁、患有难治性或处于二次复发或后期复发的b细胞前体急性成淋巴细胞性白血病(all)的患者)和axicabtageneciloleucel(适用于治疗在两个或更多个全身疗法线后仍患有复发性或难治性大b细胞淋巴瘤的成年患者)。大多数car-t细胞研究已经使用自体t细胞,因为同种异体t细胞具有发展移植物抗宿主病(gvhd)(这是一种严重且潜在致死的不良事件,包括同种异体t细胞针对宿主组织的攻击)的显著风险。自体car修饰的t细胞的制造是患者特异性的并且具有一些实践限制。经遗传修饰以表达car的nk细胞(car-nk细胞)的开发仍然处于早期开发阶段。使用靶向不同抗肿瘤剂的car-nk细胞进行的临床前研究和临床研究综述于:martin-antonio等人,intjmolsci.[国际分子科学杂志]2017年8月29日;18(9)。预期car-nk细胞与car-t细胞相比具有若干优点。在制造方面,由于nk细胞不引起gvhd,它们提供了产生可用于直接临床用途的成品同种异体产物的机会。此外,自体的和同种异体的nk细胞具有受限的体内持久性,这使得不太可能发生危及生命的毒性(像细胞因子释放综合征)。从功效角度,一旦将nk细胞用car工程化,应保留它们的天然受体,从而允许通过除由car特异性介导的机制以外的机制来介导抗肿瘤作用(rezvani等人,mol.ther.[分子疗法]2017年8月;25(8))。用于nk细胞工程化的不同策略目前还在开发中。迄今为止,公布的关于car-nk细胞的大多数临床前和早期临床研究已经利用了nk细胞系来表达car分子(martin-antonio等人,intjmolsci.[国际分子科学杂志]2017年8月29日;18(9)),其中对nk-92细胞系的研究最广泛。尽管nk-92细胞在制造方面存在一些潜在优点,但是nk-92细胞具有如下显著的临床缺点需要纳入考虑:潜在的致肿瘤性,这是因为它们源自受非霍奇金淋巴瘤、多种细胞遗传学异常、和被爱泼斯坦-巴尔病毒(ebv)潜在感染影响的患者。出于这些原因,在临床使用之前,对nk-92进行辐射处理。此类辐射对于car-nk细胞的体内增殖和持久性、以及明确地对其抗肿瘤活性具有负面影响。一些作者披露了从原代人nk细胞(源自外周血或脐带血)开始获得的、通过病毒转导或其他替代性非病毒转导方法而遗传修饰的car-nk细胞。由于不同供体中nk功能不同、以及转导方法和扩增策略影响,用这些方法观察到很大的可变性(rezvani等人,mol.ther.[分子疗法]2017年8月;25(8))。本发明集中于从干细胞开始制造car-nk细胞的方法的开发,这些干细胞纯化自生物样品,被遗传修饰以表达car并且然后分化成car-nk。发明人发现了用来进行制造方法的每个步骤的条件,以获得含有car-nk细胞的细胞群体,其中nk细胞抗肿瘤活性和car抗肿瘤活性协同地发挥作用。现有技术披露了按照类似方法的一些方案,但是现有出版物绝没有描述转导条件对中间体car-cd34+干细胞群体的性质和组成的影响、以及对最终car-nk细胞群体的治疗效果的影响。例如,lowe等人,met.mol.biol.[分子生物学方法]2016;1441:241-2511披露了用于从经修饰以表达嵌合抗原受体的脐带血来源的人造血干细胞(hsc)开始产生car-nk细胞的两个方案。在这两个方案中,在retronectin存在下,在补充有flt3、scf和tpo的培养基中进行lvv转导。然后将转导的干细胞与鼠op9-dl1细胞的饲养基质共培养(方案1),或在补充有scf、重组人flt-3配体、il-15和igf-1的aimv培养基中进行基于培养细胞的无饲养层方案(方案2)而分化成nk细胞。这些论文并没有披露本发明的制造方法以及使用的条件对car-nk细胞的最终群体的抗肿瘤作用的影响。ni等人,stemcells.[干细胞]2014年4月;32(4)披露了产生用于治疗hiv感染的car-nk细胞的另外的方案。在此情况下,将人胚胎干细胞或诱导性多能干细胞用携带car的lvv转导,然后在具有干细胞因子、血管内皮生长因子和骨形态发生蛋白4的bpel培养基中培养11天,并且然后在el08-1d2基质细胞的存在下分化成nk细胞。这些论文并没有披露本发明的方法的条件,并且有趣的是,这些论文清楚地表明尽管体外car-nk细胞能够比其未经修饰的对等物更有效地抑制cd4+t细胞中的hiv复制,但是与未经修饰的nk细胞相比,car修饰的nk细胞的体内功效没有显著增加。因此,在此方法中,在体内没有观察到nk细胞和car之间的协同作用。需要开发能够获得足够用于医学治疗的量和质量的car-nk细胞的制造方法,其特征在于nk细胞和car治疗效果之间的协同效应。本发明解决了这种需要。技术实现要素:本发明涉及开发car-nk细胞的制造方法,该方法允许获得以nk细胞和嵌合抗原受体(car)的治疗效果之间的有效协同作用为特征的细胞群体。制造方法包括第一阶段和第二阶段,该第一阶段旨在使用慢病毒载体转导来产生含有遗传修饰以表达至少一种编码car的多核苷酸的cd34+干细胞的中间体细胞群体,在该第二阶段中,将该中间体细胞群体扩增并且分化成含有car-nk细胞的细胞群体。现有技术披露了一些基于以下想法获得car-nk细胞的方法:将来自生物来源的cd34+干细胞进行病毒转导,随后分化成car-nk细胞。此类方法没有披露本发明的方法的条件。此外,通过比较不同方案,发明人出人意料地发现了不同转导和培养条件对于细胞的中间体群体的性质和组成具有影响,并且因此对于最终分化的car-nk细胞群体具有影响,从而导致不同的治疗效果。通过本发明的制造方法可获得的car-nk细胞群体在体内具有更强的治疗效果。本发明的方法包括两个阶段:在第一阶段中,将cd34+干细胞从生物样品纯化,并且通过用携带编码car的多核苷酸的慢病毒载体进行转导而进行遗传工程化。在包含细胞因子的集合的培养基存在下进行慢病毒转导,从而获得含有car-cd34+干细胞的中间体细胞群体,任选地将这些中间体细胞群体在包含该细胞因子的集合的培养基中进一步扩增。在第二阶段中,将中间体细胞群体(进一步)扩增和分化成含有car-nk细胞的细胞群体。优选地,将从阶段i获得的car-cd34+干细胞在不同于所述第一培养基的、具有细胞因子的集合的第二和第三培养基中进行培养,由此获得含有多种car-nk细胞或car-nk祖细胞或两者的培养细胞的集合。出人意料地发现,适用于进行本发明的方法的条件允许获得具有更强治疗效果(由nk细胞和car的固有抗肿瘤作用之间的协同作用导致)的car-nk细胞群体。附图说明图1.该图示出了含有cd44v6结合结构域、lngfr野生型长间隔子(包括四个tnfr-cys结构域和整个富含丝氨酸苏氨酸的柄)、cd28的跨膜和共刺激结构域、以及cd3ζ链的胞内结构域的carcd44v6野生型长(cd44v6-nwl)的示意性结构。图2.在条件a和条件b下获得的模拟物nk和cd44v6-carnk细胞的体外功能活性。该图示出了用k562、molt-4、k562-cd44v6或molt-4-cd44v6细胞刺激的,在条件a(左)或条件b(右)下获得的模拟物nk和carnk中cd107a阳性细胞(小图a;18小时)或tnf-α阳性细胞(小图b,6小时)的频率。在两个亚群(lngfr+细胞和lngfr-细胞)中进行carnk活性的分析。图3.在充分确立的疾病模型中,来自条件a和b的模拟物nk和cd44v6car-nk细胞的体内抗肿瘤作用。该图示出了针对每种条件的肝重量(克)。在条件b下(即根据本发明的方法)获得的cd44v6car-nk细胞导致具有强于对照的抗肿瘤作用。图4.a.针对k562、t98g和thp-1肿瘤细胞系(对于nk细胞的细胞毒性分别是敏感的、中间的或具有抗性的),来自条件a和b的模拟物nk(非t)和cd44v6car-nk(t)细胞的体外功能性数据(%7aad+)。b.在重构后,针对敏感性肿瘤细胞系(k562),来自条件a和b的模拟物nk(非t)和cd44v6car-nk(t)细胞的体外功能性数据(%7aad+)。c.在重构后,针对敏感性肿瘤细胞系(k562),来自条件a和b的模拟物nk(非t)和cd44v6car-nk(t)细胞的脱粒(%cd107a+)。图5.a在重构前,抑制和激活来自条件a的模拟物nk(非t)和cd44v6car-nk(t)细胞的受体表达。b.在重构后,抑制和激活来自条件a和b的模拟物nk(非t)和cd44v6car-nk(t)细胞的受体表达。具体实施方式现在将通过非限制性示例性实施例的方式,描述本发明的不同优选特征和实施例。本发明提供了用于制造用嵌合抗原受体(car)遗传修饰的nk细胞的群体的方法。本发明的方法允许获得其中nk细胞和car的治疗效果协同地发挥作用的细胞群体。发明人发现了用来获得此类协同作用的最佳条件。方法包括两个阶段:第一阶段旨在产生含有遗传修饰以表达car的干细胞的中间体细胞群体,即含有car-cd34+干细胞的中间体群体,以及在第二阶段中,将该中间体细胞群体扩增并分化成含有car-nk细胞的细胞群体。在一个实施例中,提供了用于制造用嵌合抗原受体(car)遗传修饰的nk细胞的群体的方法,该方法包括:(i)第一阶段:根据以下步骤产生含有cd34+细胞的中间体细胞群体,这些细胞携带编码car的至少一种多核苷酸:a)获得选自由以下组成的组的生物起始样品:骨髓、外周血、胎盘或脐带血,b)从此类生物样品分离造血干细胞,c)在培养基i的存在下,培养分离的造血干细胞,d)通过将培养的干细胞与携带编码car的至少一种多核苷酸的慢病毒载体一起孵育来转导此类细胞,从而获得含有car-cd34+干细胞的中间体细胞群体,以及e)任选地,在培养基i中培养该含有car-cd34+干细胞的中间体细胞群体至少一天,其中培养基i是包含细胞因子的集合的基本培养基,其中所述细胞因子的集合包含白介素-7,和干细胞因子(scf)、flt-3配体(flt-3l)、血小板生成素(tpo)中的一种或多种,以及粒细胞-巨噬细胞集落刺激因子(gm-csf)、粒细胞集落刺激因子(g-csf)和白介素-6(il-6)中的两种或更多种,(ii)第二阶段,该第二阶段包括:任选的预备步骤:在培养基ii中进一步扩增来自该第一阶段的该含有car-cd34+干细胞的中间体细胞群体,由此获得含有car-cd34+干细胞和car-nk祖细胞的细胞群体,其中培养基ii是包含细胞因子的集合的基本培养基,其中所述细胞因子的集合包含scf、flt-3l白介素-15(il-15)和il-7中的两种或更多种,以及gm-csf、g-csf和il6中的两种或更多种,以及将收集自阶段i的该含有car-cd34+干细胞的中间体细胞群体、或收集自该任选的预备步骤的该含有car-cd34+干细胞和car-nk祖细胞的细胞群体扩增和分化成含有car-nk细胞的细胞群体的步骤,该步骤包括在培养基iii中培养来自阶段i的含有car-cd34+干细胞的中间体细胞群体、或来自该任选的预备步骤的含有car-cd34+干细胞和car-nk祖细胞的细胞群体,由此获得含有nk细胞的细胞群体,其中该培养基iii是包含细胞因子的集合的基本培养基,其中所述细胞因子的集合包含scf、il-7、il-15和白介素-2(il-2)中的两种或更多种,以及gm-csf、g-csf、和il-6中的两种或更多种。总共使用了以下三种不同培养基进行培养和转导:培养基i是包含细胞因子的集合的基本培养基,其中所述细胞因子的集合包含白介素-7(il-7),和干细胞因子(scf)、flt-3配体(flt-3l)、血小板生成素(tpo)中的一种或多种,以及粒细胞-巨噬细胞集落刺激因子(gm-csf)、粒细胞集落刺激因子(g-csf)和白介素-6(il-6)中的两种或更多种。优选地,该细胞因子的集合包含il-7,和scf、flt-3l和tpo中的两种或更多种,更优选地,该细胞因子的集合包含scf、flt-3l、tpo和il-7。在一个优选实施例中,该细胞因子的集合包含gm-csf、g-csf、和il-6。特别优选的是,该培养基i包含scf、flt-3l、tpo、il-7、gm-csf、g-csf、和il-6。培养基ii是包含细胞因子的集合的基本培养基,其中所述细胞因子的集合包含scf、flt-3l白介素-15(il-15)和il-7中的两种或更多种,以及gm-csf、g-csf、和il-6中的两种或更多种。优选地,该细胞因子的集合包含scf、flt-3l、il-15和il-7中的三种或更多种,更优选地,该细胞因子的集合包含scf、flt-3l、il-15和il-7。在一个优选实施例中,该细胞因子的集合包含gm-csf、g-csf、和il-6。特别优选的是,该培养基ii包含scf、flt-3l、il-15、il-7、gm-csf、g-csf、和il-6。培养基iii是包含细胞因子的集合的基本培养基,其中所述细胞因子的集合包含scf、il-7、il-15和白介素-2(il-2)中的两种或更多种,以及gm-csf、g-csf、和il-6中的两种或更多种。优选地,该细胞因子的集合包含scf、il-7、il-15、和il-2中的三种或更多种,更优选地,该细胞因子的集合包含scf、il-7、il-15和il-2。在一个优选实施例中,该细胞因子的集合包含gm-csf、g-csf、和il-6。特别优选的是,该培养基iii包含scf、il-7、il-15、il-2、gm-csf、g-csf、和il-6。第一阶段:产生含有car-cd34+干细胞的中间体群体(i)本发明的方法的第一阶段提供了产生含有cd34+干细胞的中间体细胞群体的条件,这些干细胞携带至少一种编码car的多核苷酸。根据以下步骤产生该中间体细胞群体:a)获得选自由以下组成的组的生物起始样品:骨髓、外周血、胎盘或脐带血,b)从此类生物样品分离造血干细胞,c)在培养基i的存在下,培养分离的造血干细胞,d)通过将培养的干细胞与携带编码car的至少一种多核苷酸的慢病毒载体一起孵育来转导此类细胞,从而获得含有car-cd34+干细胞的中间体细胞群体,以及e)任选地,在培养基i中培养该含有car-cd34+干细胞的中间体细胞群体至少一天,其中培养基i是包含细胞因子的集合的基本培养基,其中所述细胞因子的集合包含白介素-7,和干细胞因子(scf)、flt-3配体(flt-3l)、血小板生成素(tpo)中的一种或多种,以及粒细胞-巨噬细胞集落刺激因子(gm-csf)、粒细胞集落刺激因子(g-csf)和白介素-6(il-6)中的两种或更多种。本发明的方法中使用的起始材料是含有成体(胚后期)干细胞(也称为体干细胞)的生物样品。如本文所用,术语生物样品意指源自人类的样品。在一个优选实施例中,待使用的起始材料是脐带血。根据本发明的方法,从生物样品分离cd34+干细胞。本领域已知用于cd34+分离的不同方案,包括基于免疫磁性选择或细胞分选的方法。如本文所用,免疫磁性选择是指将抗体偶联至磁性颗粒,从而使得能够通过使用磁体分离抗原结构。在一个优选实施例中,首先使用梯度分离或离心技术使生物样品富集单核细胞,并且然后通过用缀合至磁珠的特异性抗cd34+抗体标记细胞和使用磁柱纯化cd34+细胞来进行免疫磁性选择。在一个另外的优选实施例中,使用midimacstm分离器、plus仪器或clinimacs装置进行免疫磁分离。在本发明的方法的第一阶段期间,首先培养分离的cd34+干细胞,并且然后在包含细胞因子和生长因子的混合物的基本培养基存在下进行转导。许多基本培养基是已知的。以下给出了选择,但是还有许多可能是适合的。基本培养基包括但不限于bem(基本伊格尔培养基)、dmem(杜氏改良的伊格尔培养基)、格拉斯哥极限必需培养基、m199基础培养基、hamf-10、hamf-12、伊思考夫(iscove)dmem、rpmi、莱博维茨氏(leibovitz)l15、mcdb、mccoy5a、stemspan和stemlineitm和stemlineiitm、格雷克斯迪姆(glycostem)基础生长培养基(gbgmtm);x-vivo10tm、x-vivo15tm和x-vivo20tm干细胞生长培养基等。还可以使用这些基本培养基的组合。优选地,在添加和/或不添加人血清的培养开始的时间点,使用无血清配制品,例如stemlineitm和stemlineiitm、stemspan或x-vivo10tm、gbgm、x-vivo15tm和x-vivo20tm。可替代地,或此外,基本培养基可以补充有其他(人)血液衍生物,例如血小板裂解物、自体的或同种异体的供体血清、(富血小板)血浆、白蛋白、或血清的合成替代物(例如像球蛋白富集的蛋白补充物)。本文给出的量典型地适合培养。这些量可以适合于开始培养时与其一起的细胞的不同量。在根据本发明的不同培养步骤中使用的培养基的血清含量可以变化,优选地,与细胞因子的不同组合一起在根据本发明的限定时间点处提供扩增培养基或分化培养基,或可替代地,扩增+分化培养基。在分离时,按从0,5x106至2x106个细胞/ml的浓度范围,将cd34+干细胞接种在容器中,例如板、烧瓶、细胞工厂或袋中。在一个优选实施例中,按浓度1x106个细胞/ml接种细胞。在一个实施例中,将细胞接种在容器中,这些容器预先涂覆有纤连蛋白的片段,例如片段ch-296或功能衍生物。当涂覆在细胞容器的表面时,显著增强了病毒载体介导的基因转导入哺乳动物细胞。在一个优选实施例中,细胞容器涂覆有一旦接种干细胞,在培养基i中培养这些干细胞。在一个实施例中,可以进行培养阶段,持续至少12-48小时(更优选地,22+/-2小时)的时间段。在一个优选实施例中,培养基i包含以下浓度范围的细胞因子中的一种或多种:2-100pg/ml(优选地,5-50pg/ml,最优选地约10pg/ml)的gm-csf,100至1000pg/ml(优选地,150至500pg/ml,最优选地约250pg/ml)的g-csf,4ng/ml至300ng/ml(优选地,10至100ng/ml,最优选地约25ng/ml)的scf,4ng/ml至300ng/ml(优选地,10至100ng/ml,最优选地约25ng/ml)的flt3-l,4ng/ml至100ng/ml(优选地,10至50ng/ml,最优选地约25ng/ml)的tpo,5-500pg/ml(优选地,25-100pg/ml,最优选地,约50pg/ml)的il-6,和/或4ng/ml至100ng/ml(优选地,10至50ng/ml,最优选地约25ng/ml)的il7。在本发明的一个更优选方面,培养基i包含浓度为约25ng/mlscf,约25ng/mlflt3-l、约25ng/mltpo、约250pg/mlg-csf、约10pg/mlgm-csf、约50pg/mlil-6、和约25ng/mlil7的细胞因子。在此背景下,“约”意指约20%,优选地10%、更优选地5%、最优选地2%的偏差。在一个具体的优选实施例中,提供了根据本发明的方法,其中培养基i包括:浓度为4ng/ml至300ng/ml的scf、或浓度为4ng/ml至300ng/ml的flt3-l、或浓度为4ng/ml至100ng/ml的tpo、或浓度为4ng/ml至50ng/ml的il7、或指定浓度范围的这些细胞因子的任何组合。在一个更优选实施例中,提供了根据本发明的方法,其中培养基i包括:浓度为2pg/ml至100pg/ml的gm-csf、或浓度为100pg/ml至1000pg/ml的g-csf、或5-500pg/ml的il6、或按指定浓度范围的这些细胞因子的任何组合。优选地,培养基i包含4-100μg/ml肝素、优选地10-40μg/ml肝素、更优选地约20μg/ml肝素。优选地,培养基i包含0.5%-25%血清、更优选地1%-20%血清、最优选地约10%血清。优选地,血清是人血清。在另一个优选实施例中,培养基i、ii和iii彼此独立,包含另一种(人)血液衍生物,例如血小板裂解物、自体的或同种异体的供体血清、(富血小板)血浆、白蛋白、或血清的合成替代物(例如像球蛋白富集的蛋白补充物)。此类血清替代物的最佳浓度可以由技术人员容易地确定。在培养步骤后,将cd34+干细胞遗传修饰以表达至少一种编码car的多核苷酸。用来进行干细胞的遗传工程化的技术是慢病毒转导。如本文所用,术语转导或慢病毒转导是指使用慢病毒载体将外源多核苷酸引入到细胞的基因组中。术语慢病毒载体用于指介导核酸转移的慢病毒颗粒。根据本发明的方法,通过在培养基i存在下,将cd34+干细胞与携带至少一种编码car的多核苷酸的慢病毒载体一起孵育来进行转导。在一个实施例中,通过用含有携带编码car的多核苷酸的慢病毒载体的新鲜培养基i取代至少一半的培养基来进行孵育。在另一个实施例中,通过将cd34+干细胞重新悬浮于含有携带编码car的多核苷酸的慢病毒载体的新鲜培养基i中,然后以1x106至10x106个细胞/ml(优选地2x106至5x106个细胞/ml、最优选地约2x106个细胞/ml)的浓度范围将此重新悬浮液接种到细胞容器中来进行孵育。如本文所用,术语“转导运行”是指cd34+干细胞与新鲜慢病毒载体一起孵育的单个循环。孵育的时间段可以根据慢病毒载体的性质而变化。在一个优选实施例中,在转导运行期间,cd34+干细胞可以与慢病毒载体一起孵育10至24小时(优选地13至20个小时、更优选地约15个小时)的时间段。在此背景下,“约”意指约20%,优选地10%、更优选地5%、最优选地2%的偏差。根据本发明的方法,转导可以包括一次或多次转导运行。在本发明的另一个方面,细胞容器可以涂覆有可以在增强转导功效的另外的添加剂(例如聚凝胺、硫酸鱼精蛋白或硫酸软骨素)存在下进行转导。取决于载体的性质,慢病毒载体可以与不同浓度的cd34+干细胞一起孵育。优选地,通过孵育10至200的感染复数(moi)范围的慢病毒载体来进行转导。moi是慢病毒载体颗粒数量与限定的空间内存在的靶细胞数量的比率。在一个优选实施例中,按约moi100孵育慢病毒载体。转导导致产生含有car-cd34+干细胞的中间体细胞群体,这些干细胞携带至少一种编码car的多核苷酸。可以将如此获得的含有car-cd34+干细胞的中间体细胞群体在新鲜培养基i存在下再培养至少一天,然后移至制造方法的第二阶段。在另一个实施例中,可以在转导结束时冷冻含有car-cd34+干细胞的中间体细胞群体。在另一个实施例中,可以将中间体细胞群体在新鲜培养基i存在下再培养至少一天,并且然后可以冷冻。可以在转导结束后或在培养一天或多天后,或在冷冻的含有car-cd34+干细胞的中间体细胞群体解冻后,进行制造的第二阶段。优选地,在培养基i中,在总共至少7个培养日后,进行第二阶段。在本发明的一个实施例中,提供了通过本发明的方法的第一阶段可获得的含有car-cd34+干细胞的中间体细胞群体。在另一个实施例中,本发明的方法的第一阶段可以进一步包括选择car-cd34+干细胞从而获得car-cd34+干细胞的纯群体的步骤。技术人员可以应用本领域已知的不同方法来选择car-cd34+干细胞。在一个实施例中,可以用慢病毒载体转导cd34+干细胞,该慢病毒载体携带一种编码car的多核苷酸和编码通常并不由cd34+细胞表达的可选择蛋白质(待用作免疫选择的标记)的另外的多核苷酸。适合用作标记的可选择蛋白质的实例是截短形式的受体蛋白,例如像人低亲和力神经生长因子(δlngfr)、人表皮生长因子受体、cd19或cd20的胞外结构域。以这种方式,转导的car-cd34+干细胞将共表达car和可选择蛋白质,并且因此可以通过用靶向可选择标记的特异性抗体(此类抗体缀合至磁珠)标记细胞并且使用磁柱纯化car-cd34+细胞进行免疫磁性选择方法来选择。在一个另外的优选实施例中,使用midimacstm分离器、plus仪器或clinimacs装置进行免疫磁分离。在另一个优选实施例中,可以通过免疫磁性选择方法(其中抗体靶向car分子的可选择区域)选择car-cd34+干细胞。例如,可以用携带一种编码car的多核苷酸的慢病毒载体转导cd34+干细胞,此类car含有用作免疫选择的标记的间隔子结构域,即可以通过缀合至磁珠的抗体而特异性识别,从而允许使用磁柱纯化car-cd34+细胞的间隔子结构域。优选地,使用midimacstm分离器、plus仪器或clinimacs装置进行免疫磁分离。优选地,用作标记的间隔子结构域源自通常并不由cd34+细胞表达的蛋白质的胞外结构域。在一个另外的优选实施例中,可以用携带一种编码car的多核苷酸的慢病毒载体转导cd34+干细胞,其中该car含有由人低亲和力神经生长因子(δlngfr)之一的胞外结构域衍生的间隔子结构域。如前所述,优选地,其中发生转导的培养条件(即在培养基i中培养)继续持续总共至少7天,即包括在转导前培养、在转导运行1和2期间培养、和在转导后培养。此后,开始第二阶段,该第二阶段包括扩增并且分化成nk细胞。第二阶段:将含有car-cd34+干细胞的中间体细胞群体扩增并且分化成含有car-nk细胞的细胞群体本发明的方法的第二阶段提供了其中将中间体细胞群体(或进一步扩增的细胞群体(参见下文))扩增并且分化成含有car-nk细胞的细胞群体的条件,包括在培养基iii中培养来自第一阶段的含有car-cd34+干细胞的中间体(或进一步扩增的)细胞群体,由此获得含有nk细胞的细胞群体,其中该培养基iii是包含细胞因子的集合的基本培养基,其中所述细胞因子的集合包含scf、il-7、il-15和白介素-2(il-2)中的两种或更多种,以及gm-csf、g-csf、和il-6中的两种或更多种。在培养基iii中培养中间体细胞群体(或进一步扩增的细胞群体)至少7天,由此获得含有多种car-nk细胞或car-nk祖细胞或两者的培养细胞的集合。在本发明的另外的方面,该方法在该第二阶段进一步包括预备步骤:在培养基ii中培养来自该第一阶段的该含有car-cd34+干细胞的中间体细胞群体,由此获得含有car-cd34+干细胞和car-nk祖细胞的细胞群体,其中培养基ii是包含细胞因子的集合的基本培养基,其中所述细胞因子的集合包含scf、flt-3l白介素-15(il-15)和il-7中的两种或更多种,以及gm-csf、g-csf和il6中的两种或更多种。使用培养基ii的预备培养步骤是任选的,并且旨在获得更多car-cd34+干细胞或car-nk祖细胞,从而进入旨在分化的用培养基iii的培养步骤。在此预备培养步骤中,将收集自阶段i的含有car-cd34+干细胞的中间体细胞群体在培养基ii中培养至少4天,由此获得含有定型为nk细胞谱系的多种car-nk祖细胞的培养的car-cd34+干细胞、car-nk祖细胞或两者的集合。在一个优选实施例中,培养基ii包含以下浓度范围的细胞因子中的一种或多种:50pg/ml至1000ng/ml(优选地,150至400ng/ml)的g-csf,2-100pg/ml(优选地,5-25ng/ml)的gm-csf,4-300ng/ml(优选地,10-100ng/ml)的scf,4ng/ml至300ng/ml(优选地,10ng/ml至100ng/ml)的flt3-l,4ng/ml至300ng/ml(优选地,10ng/ml至100ng/ml)的il15,2pg/ml至500pg/ml(优选地,20pg/ml至200pg/ml)的il-6,和/或4ng/ml至100ng/ml(优选地,10ng/ml至50ng/ml)的il7。在本发明的一个更优选方面,培养基ii包含浓度为约10pg/mlgm-csf、约250pg/mlg-csf、约25ng/mlscf、约25ng/mlflt3-l、约20ng/mlil-15、约50pg/mlil-6、和约25ng/mlil7的细胞因子。在此背景下,“约”意指约20%,优选地10%、更优选地5%、最优选地2%的偏差。优选地,培养基ii包含4-100μg/ml肝素、优选地10-40μg/ml肝素、更优选地约20μg/ml肝素。优选地,培养基ii包含0.5%-25%血清、更优选地1%-20%血清、最优选地约10%血清。优选地,血清是人血清。在一个优选实施例中,培养基iii包含以下浓度范围的细胞因子中的一种或多种:50pg/ml至1000ng/ml(优选地,150至400ng/ml)的g-csf,2pg/ml至100pg/ml(优选地,5ng/ml至25ng/ml)的gm-csf,4ng/ml至300ng/ml(优选地,10ng/ml至100ng/ml)的scf,200u/ml至5000u/ml(优选地,500至2000u/ml)的il-2,4ng/ml至300ng/ml(优选地,10ng/ml至100ng/ml)的il15,2pg/ml至500pg/ml(优选地,20pg/ml至200pg/ml)的il-6,和/或4ng/ml至100ng/ml(优选地,10ng/ml至50ng/ml)的il7。在本发明的一个更优选方面,培养基iii包含浓度为约10pg/mlgm-csf、约250pg/mlg-csf、约20ng/mlscf、约25ng/ml、约20ng/mlil-15、约50pg/mlil-6、约1000u/mlil-2、和约20ng/mlil7的细胞因子。在此背景下,“约”意指约20%,优选地10%、更优选地5%、最优选地2%的偏差。优选地,培养基iii包含0.5%-10%血清、更优选地1%-5%血清、最优选地约2%血清。优选地,血清是人血清。嵌合抗原受体用于本发明的方法的慢病毒载体携带至少一种编码car的多核苷酸。用于本发明的方法的car是重组嵌合受体,这些重组嵌合受体包含:(i)抗原特异性靶向结构域;(ii)间隔子结构域;(iii)跨膜结构域;(iv)任选地至少一个共刺激结构域;以及(v)胞内信号传导结构域。用于本发明的方法的car的胞外结构域包含具有结合目的靶抗原的功能的抗原特异性靶向结构域。抗原特异性靶向结构域可以是特异性结合靶抗原的任何天然存在的、合成的、半合成的、或重组技术产生的分子、蛋白质、肽或寡肽。可能的抗原特异性靶向结构域的实例包括抗体或抗体片段或衍生物,靶向受体的分子的合成的或天然存在的配体,包括分子、受体或结合蛋白的结合结构域或胞外结构域。在一个优选实施例中,所述抗原特异性靶向结构域是抗体或源自抗体。抗体是源自能够结合抗原的免疫球蛋白的蛋白质、或多肽序列。如本文所用的抗体包括多克隆抗体或单克隆抗体、多链抗体或单链抗体以及免疫球蛋白,无论源自天然来源还是重组来源。源自抗体的靶向结构域可以是抗体的片段或抗体的一个或多个片段的遗传工程化的产物,该片段涉及与抗原结合。实例包括可变区(fv)、互补决定区(cdr)、fab、单链抗体(scfv)、重链可变区(vh)、轻链可变区(vl)和骆驼抗体(vhh)。在一个优选实施例中,结合结构域是单链抗体(scfv)。scfv可以是鼠的、人的或人源化的scfv。在一个优选实施例中,靶抗原是肿瘤抗原。如本文所用,术语肿瘤抗原包括在肿瘤细胞(包括生物标记,或肿瘤细胞上发现的、且基本不见于正常组织上或其表达限制在非重要正常组织中的细胞表面标记)上表达的抗原。在一个优选实施例中,用于本发明的方法的car靶向cd44v6肿瘤抗原。在一个另外的优选实施例中,本发明的方法的car中的抗原特异性靶向结构域是抗cd44v6scfv。抗cd44v6scfv可以源自us6’972’324中披露的抗cd44v6抗体。间隔子结构域用于本发明的方法的car包含将抗原特异性靶向结构域连接至跨膜结构域的胞外间隔子结构域。用作间隔子的最常见序列是恒定免疫球蛋白igg1铰链-ch2-ch3fc结构域。本领域披露了此间隔子的突变体和或变体,例如hombach等人,2010genether.[基因疗法]17:1206-1213披露了一种间隔子,其中用pva/a突变修饰人lgg1的ch2和ch3结构域以减少fc受体结合。在一个优选实施例中,用于本发明的方法的car的间隔子是源自人低亲和力神经生长因子(lngfr)的胞外结构域的片段,如wo2016/042461中披露。在属于此属的可能的序列中,wo2016/042461披露了四个特定种类:(i)lngfr的整个胞外结构域(即:包括四个tnfr-cys结构域和富含丝氨酸苏氨酸的柄);(ii)lngfr的整个胞外结构域的突变版本(此类突变由以下组成:被三个特定氨基酸取代的第四个tnfr-cys结构域的片段的缺失);(iii)仅包括lngfr的胞外结构域的四个tnfr-cys结构域的片段;(iv)包括lngfr的胞外结构域的前三个tnfr-cys结构域和第四个的突变版本(此类突变由以下组成:被三个特定氨基酸取代的第四个tnfr-cys结构域的片段的缺失)的片段。lngfr并不在大多数人造血细胞上表达,因此,源自lngfr的间隔子单元可以用于协助遗传工程化以表达car的细胞的选择,和通过免疫荧光的转导的基因表达进行定量分析。在本发明的另外的方面,制造方法的第一阶段可以进一步包括在转导后进行car-cd34+干细胞的免疫磁性选择的步骤。例如,在转导步骤结束后,可以将细胞与缀合至磁珠并且能够识别lngfr来源的间隔子的抗体一起孵育,从而允许使用磁柱纯化car-cd34+干细胞。以下报道了lngfr来源的间隔子的实例:间隔子lngfr野生型长(nwl):蛋白质序列(seqidno:1):keacptglythsgecckacnlgegvaqpcganqtvcepcldsvtfsdvvsatepckpctecvglqsmsapcveaddavcrcaygyyqdettgrceacrvceagsglvfscqdkqntvceecpdgtysdeanhvdpclpctvcedterqlrectrwadaeceeipgrwitrstppegsdstapstqepeappeqdliastvagvvttvmgssqpvvtrgttdn。核苷酸序列(seqidno:2):aaagaggcctgccccaccggcctgtacacccacagcggagagtgctgcaaggcctgcaacctgggagagggcgtggcccagccttgcggcgccaatcagaccgtgtgcgagccctgcctggacagcgtgaccttcagcgacgtggtgtccgccaccgagccctgcaagccttgcaccgagtgtgtgggcctgcagagcatgagcgccccctgcgtggaagccgacgacgccgtgtgtagatgcgcctacggctactaccaggacgagacaaccggcagatgcgaggcctgtagagtgtgcgaggccggcagcggcctggtgttcagttgtcaagacaagcagaataccgtgtgtgaagagtgccccgacggcacctacagcgacgaggccaaccacgtggacccctgcctgccctgcactgtgtgcgaggacaccgagcggcagctgcgcgagtgcacaagatgggccgacgccgagtgcgaagagatccccggcagatggatcaccagaagcaccccccctgagggcagcgacagcaccgcccctagcacccaggaacctgaggcccctcccgagcaggacctgatcgcctctacagtggccggcgtggtgacaaccgtgatgggcagctctcagcccgtggtgacacggggcaccaccgacaat。间隔子lngfr野生型短(nws):蛋白质序列(seqidno:3):keacptglythsgecckacnlgegvaqpcganqtvcepcldsvtfsdvvsatepckpctecvglqsmsapcveaddavcrcaygyyqdettgrceacrvceagsglvfscqdkqntvceecpdgtysdeanhvdpclpctvcedterqlrectrwadaecee。核苷酸序列(seqidno:4):aaagaggcctgccccaccggcctgtacacccacagcggagagtgctgcaaggcctgcaacctgggagagggcgtggcccagccttgcggcgccaatcagaccgtgtgcgagccctgcctggacagcgtgaccttcagcgacgtggtgtccgccaccgagccctgcaagccttgcaccgagtgtgtgggcctgcagagcatgagcgccccctgcgtggaagccgacgacgccgtgtgtagatgcgcctacggctactaccaggacgagacaaccggcagatgcgaggcctgtagagtgtgcgaggccggcagcggcctggtgttcagttgtcaggacaagcagaacaccgtgtgtgaagagtgccccgacggcacctacagcgacgaggccaaccacgtggacccctgcctgccctgcactgtgtgcgaggacaccgagcggcagctgcgcgagtgcacaagatgggccgacgccgagtgcgaggaa。间隔子lngfr突变型长(nml):蛋白质序列(seqidno:5):keacptglythsgecckacnlgegvaqpcganqtvcepcldsvtfsdvvsatepckpctecvglqsmsapcveaddavcrcaygyyqdettgrceacrvceagsglvfscqdkqntvceecpdgtysdeaaraadaeceeipgrwitrstppegsdstapstqepeappeqdliastvagvvttvmgssqpvvtrgttdn。核苷酸序列(seqidno:6):aaagaggcctgccccaccggcctgtacacccacagcggagagtgctgcaaggcctgcaacctgggagagggcgtggcccagccttgcggcgccaatcagaccgtgtgcgagccctgcctggacagcgtgaccttcagcgacgtggtgtccgccaccgagccctgcaagccttgcaccgagtgtgtgggcctgcagagcatgagcgccccctgcgtggaagccgacgacgccgtgtgtagatgcgcctacggctactaccaggacgagacaaccggcagatgcgaggcctgtagagtgtgcgaggccggcagcggcctggtgttcagttgtcaagacaagcagaataccgtgtgtgaagagtgccccgacggcacctacagcgacgaagccgccagagccgccgacgccgagtgcgaagagatccccggcagatggatcaccagaagcaccccccctgagggcagcgacagcaccgcccctagcacccaggaacctgaggcccctcccgagcaggacctgatcgcctctacagtggccggcgtggtgacaaccgtgatgggcagctctcagcccgtggtgacacggggcaccaccgacaat。间隔子lngfr突变型短(nms):蛋白质序列(seqidno:7):keacptglythsgecckacnlgegvaqpcganqtvcepcldsvtfsdvvsatepckpctecvglqsmsapcveaddavcrcaygyyqdettgrceacrvceagsglvfscqdkqntvceecpdgtysdeaaraadaecee。核苷酸序列(seqidno:8):aaagaggcctgccccaccggcctgtacacccacagcggagagtgctgcaaggcctgcaacctgggagagggcgtggcccagccttgcggcgccaatcagaccgtgtgcgagccctgcctggacagcgtgaccttcagcgacgtggtgtccgccaccgagccctgcaagccttgcaccgagtgtgtgggcctgcagagcatgagcgccccctgcgtggaagccgacgacgccgtgtgtagatgcgcctacggctactaccaggacgagacaaccggcagatgcgaggcctgtagagtgtgcgaggccggcagcggcctggtgttcagttgtcaggacaagcagaacaccgtgtgtgaagagtgccccgacggcacctacagcgacgaggccgcccgggccgccgacgccgagtgcgaggaa。跨膜结构域用于本发明的方法的car包含间隔子结构域和信号传导结构域之间的跨膜结构域。该跨膜结构域可以源自天然来源或合成来源。源自天然来源的结构域可以包含来自任何膜结合蛋白或跨膜蛋白(包括类型i、类型ii或类型iii跨膜蛋白中的任一者)的跨膜序列。可用于car的跨膜区可以源自t细胞受体的α、β或ζ链,cd28、cd3ε、cd45、cd4、cd5、cd8、cd9、cd16、cd22、cd33、cd37、cd64、cd80、cd86、cd134、cd137、cd154、cd244(2b4)、dap10或dap12。源自合成来源的结构域将主要包含疏水序列(包括残基,例如亮氨酸和缬氨酸)。现有技术披露了可以用于car的跨膜结构域的实例,例如:1)cd28tm区(pule等人,molther[分子疗法],2005年11月;12(5):933-41;brentjens等人,ccr,2007年9月15日;13(18pt1):5426-35;casucci等人,blood[血液],2013年11月14日;122(20):3461-72);2)ox40tm区(pule等人,molther[血液],2005年11月;12(5):933-41);3)41bbtm区(brentjens等人,ccr,2007年9月15日;13(18pt1):5426-35);4)cd3ζtm区(pule等人,molther[分子疗法],2005年11月;12(5):933-41;savoldob,blood[血液],2009年6月18日;113(25):6392-402);5)cd8atm区(maher等人,natbiotechnol[自然生物技术],2002年1月;20(1):70-5;imaic,leukemia[白血病],2004年4月;18(4):676-84;brentjens等人,ccr,2007年9月15日;13(18pt1):5426-35;milone等人,molther[分子疗法],2009年8月;17(8):1453-64)。本领域技术人员可以将跨膜结构域的另外的实例应用至用于本发明的方法的car。在一个实施例中,用于本发明的方法的car包含选自以下任一个或多个的跨膜结构域:t细胞受体复合物的ζ链、cd28、cd8a、cd4的跨膜结构域或其组合。优选地,该跨膜结构域源自cd28。更优选地,cd28的跨膜结构域由以下组成:序列fwvlvvvggvlacysllvtvafiifwv(seqidno:9)。在一个实施例中,跨膜结构域和胞内信号传导结构域包含与seqidno:9的至少85%、90%、95%、97%、98%或99%同一性。共刺激结构域在胞质尾区,用于本发明的方法的car可以包括一个或多个共刺激结构域。此类结构域可以由以下组成:一个或多个共刺激蛋白受体(例如cd28、41bb、icos)的胞内信号传导结构域。共刺激结构域的功能是向细胞提供另外的信号,从而增强细胞扩增、细胞存活和记忆细胞的发育。用于本发明的方法的car可以包含选自由以下组成的组的一个或多个共刺激结构域:tnfr超家族、cd28、cd137(4-1bb)、cd134(ox40)、daplo、cd27、cd2、cd5、icam-1、lfa-1、lck、tnfr-1、tnfr-ii、fas、cd30、cd40、cd244(2b4)、dap10、dap12的胞内结构域或其组合。本领域技术人员可以将共刺激结构域的另外的实例用于car。在一个实施例中,共刺激结构域源自cd28的胞内结构域。在一个优选实施例中,跨膜结构域和共刺激结构域两者都源自cd28。在一个实施例中,跨膜结构域和胞内共刺激结构域包含以下序列:人cd28的跨膜部分和胞内部分(uniprot:p10747,cd28_人,位置153-220)fwvlvvvggvlacysllvtvafiifwvrskrsrllhsdymnmtprrpgptrkhyqpyapprdfaayrs(seqidno:10)在一个实施例中,跨膜结构域和共刺激结构域包含与seqidno:10的至少85%、90%、95%、97%、98%或99%同一性。在一个实施例中,car的胞内共刺激结构域源自cd28的胞内结构域并且包含序列rskrsrllhsdymnmtprrpgptrkhyqpyapprdfaayrs(seqidno:11)。胞内信号传导结构域用于本发明的方法的car还可以包含胞内信号传导结构域。此结构域可以是细胞质的,传递激活信号并且指导细胞进行其特化功能。胞内信号传导结构域的实例包括但不限于t细胞受体的ζ链或其同源物中任一个(例如,η链、fcεr1γ和β链、mb1(igα)链、b29(igβ)链等),cd3多肽(δ、δ和ε),syk家族酪氨酸激酶(syk、zap70等),src家族酪氨酸激酶(lck、fyn、lyn等)以及t细胞信号转导中涉及的其他分子,例如cd2、cd5和cd28。胞内信号传导结构域可以是人cd3ζ链、fcyriii、fcsri、fc受体的胞质尾区、具有胞质受体的基于免疫受体酪氨酸的活化基序(itam)、或其组合。在一个优选实施例中,信号传导结构域包括人cd3ζ链的胞内信号传导结构域。在一个实施例中,人cd3ζ链的胞内信号传导结构域包含以下序列:uniprot:p20963,cd3z_人,位置31-143rvkfsrsadapayqqgqnqlynelnlgrreeydvldkrrgrdpemggkpqrrknpqeglynelqkdkmaeayseigmkgerrrgkghdglyqglstatkdtydalhmqalppr(seqidno:12)在一个实施例中,胞内信号传导结构域包含与seqidno:12的至少85%、90%、95%、97%、98%或99%同一性。对于本领域技术人员而言,另外的胞内信号传导结构域是显而易见的,并且可以与本发明的替代实施例结合使用。多核苷酸如本文所用的术语多核苷酸被定义为形成dna或rna片段的核苷酸的聚合物。本领域技术人员具有以下公知常识:真核生物遗传密码的64个密码子编码仅20种天然存在的氨基酸,并且3个终止密码子相对于编码氨基酸残基而使遗传密码简并。鉴于遗传密码的简并性,不同多核苷酸序列可以编码相同的多肽。通过应用常规技术,可能详述含有核苷酸取代并且仍然编码本发明的多肽的不同多肽序列。用于修饰多核苷酸的方法是本领域已知的,并且可以由技术人员应用以改善多肽的活性或稳定性,或避免剪接现象。可以通过本领域可用的任何手段获得本发明的多核苷酸,这些手段包括但不限于重组手段,即使用普通克隆技术、pcrtm等克隆来自重组文库或细胞基因组的核酸序列,以及通过合成手段。用于本发明的多核苷酸可以是密码子优化的。密码子优化是本领域已知的技术(wo1999/41397和wo2001/79518),旨在增加或减少目的细胞中的蛋白表达。多种密码子通常编码相同氨基酸,但是在每种生物体中,密码子的优先使用是不同的。因此,在每种生物体中,对应于某些密码子的t-rna比对应于其他密码子的t-rna更丰富。可以通过使用与最丰富的简并trna匹配而不影响蛋白质的氨基酸序列的密码子,合成或修饰多核苷酸,从而增加宿主细胞中的蛋白质表达。慢病毒载体在一个优选实施例中,用于本发明的载体是慢病毒载体。如本文所用的慢病毒载体是指逆转录病毒载体家族内的属。在逆转录病毒载体中,由于慢病毒载体能够感染不分裂细胞,所以它们具有独特的特性。慢病毒载体提供了实现体内显著水平的基因转移的手段。慢病毒的详细清单可以见于coffin等人(“retroviruses[逆转录病毒]”1997coldspringharbourlaboratorypress[冷泉港实验室出版社]编辑:jmcoffin,smhughes,hevarmus758-763页)。简而言之,慢病毒可以分为灵长类动物组和非灵长类动物组。灵长类动物慢病毒的实例包括但不限于:人免疫缺陷病毒(hiv)(人获得性免疫缺陷综合征(aids)的病原体)和猿猴免疫缺陷病毒(siv)。非灵长类动物慢病毒组包括原型“慢病毒”维斯纳/梅迪病毒(visna/maedivirus,vmv),以及相关的山羊关节炎脑炎病毒(caev)、马传染性贫血病毒(eiav),和最近描述的猫免疫缺陷病毒(fiv)和牛免疫缺陷病毒(biv)。在一个优选实施例中,慢病毒载体源自hiv慢病毒,更优选地,慢病毒载体源自hiv-1慢病毒。用于本发明的方法的示例性慢病毒载体包含慢病毒基因组的至少以下部分:a)5’长末端重复序列(ltr);b)包装序列psi;c)rev应答元件(rre);d)可操作地连接至目的基因的启动子;e)3’长末端重复序列(ltr)。在一个优选实施例中,将5'ltr的u3区替换为选自由以下组成的组的异源启动子:巨细胞病毒(cmv)启动子、劳斯肉瘤病毒(rsv)启动子、或猿猴病毒40(sv40);由此使慢病毒转录tat独立。在一个另外的优选实施例中,3’ltr序列含有u3区的缺失(即载体是自失活载体或sin载体)。慢病毒载体可以进一步包含慢病毒中央多聚嘌呤区(cppt)和土拨鼠肝炎病毒(whp)转录后调控元件(wpre)。用于本发明的另外的示例性慢病毒载体是描述于以下中的载体:amendola等人,natbiotechnol.[自然生物技术]2005年1月;23(1):108-16,该载体包括双向启动子,用于以相反方向表达两个编码序列,从而使得能够实现有效的双基因转移。双向启动子由以下组成:来自人巨细胞病毒的最小核心启动子元件(mcmv),在上游并且以相反方向连接至源自人磷酸甘油酸激酶(pgk)或聚泛素ubi-c基因的有效启动子。掺入双向启动子的此慢病毒载体可以用于在一个单一构建体中表达car和自杀基因。在本发明的一个优选方面,自杀基因是单纯疱疹病毒胸苷激酶(hsv-tk),更优选地,是hsv-tk基因的无剪接变体,例如wo2005/123912中披露的那些。在一个另外的优选实施例中,自杀基因是由以下多核苷酸序列编码的hsv-tkmut2:在本发明的另一个方面,提供了包括双向启动子的慢病毒载体,该双向启动子允许能够在体内改善nk细胞存活和增殖的car和细胞因子的表达。在一个优选实施例中,能以这种程度使用细胞因子il15。可以使用瞬时的或稳定的包装系统进行慢病毒载体的制造。在瞬时系统中,至少用以下共转染包装细胞(例如hek293细胞或hek293thek293-sf、te671、ht1080或hela):编码慢病毒gag/pol的包装质粒、编码目的包膜蛋白的质粒、和携带如以上披露的必需慢病毒基因组元件和目的基因的转移质粒。在另外的瞬时系统中,慢病毒调控蛋白rev可以在第四个单独的质粒上反式表达。适合的env基因的实例包括但不限于vsv-genv、mlv4070env、rd114env、rd114-tr、rd114pro、杆状病毒5gp64env、galv或源自麻疹病毒的包膜蛋白。可以使用本领域熟知的方法,例如使用磷酸钙或可商购的配制品(例如聚乙烯亚胺(pei)或lipofectaminetm2000cd)进行共转染。用于产生慢病毒载体的适合的包装细胞系披露于例如:wo2012/028681,wo2004/022761,broussau等人,mol.ther.[分子疗法]2008;16(3),500-507或throm等人2009年5月21日;113(21):5104-10。细胞本发明进一步提供了包含通过本发明的方法可获得的car-nk细胞的组合物。出人意料地发现,适用于用携带嵌合抗原受体的慢病毒载体转导干细胞的条件对含有car-cd34+干细胞的中间体细胞群体的性质和组成具有影响,并且对源自扩增阶段和分化阶段的含有car-nk细胞的最终细胞群体具有影响。根据本发明的方法可获得的car-nk细胞呈现了nk细胞和car之间的协同治疗效果。此类效果导致强于用不同制造方法获得的效果。因此,本发明进一步提供了含有通过本发明的方法可获得的car-nk细胞的组合物,用于在医学中使用、更优选地用于在免疫疗法中使用、特别是用于治疗肿瘤和血液恶性肿瘤。术语“免疫疗法”表示使用人的免疫系统的某些部分来对抗疾病(例如癌症)的治疗。免疫系统的部分可以来自患有疾病的人,但还可以来自另一个人,称为“供体”,例如本发明中的情况。用于根据本发明使用的组合物优选地用于基于细胞的免疫疗法,其中将源自自体的、非单倍体相同的供体的免疫效应细胞施用至有需要的接受者。本发明优选使用由gmp顺应性培养系统产生的细胞用于例如从脐带血(ucb)来源的cd34+祖细胞(优选地没有t细胞污染)产生大批的免疫效应细胞。使用此类细胞是有利地,因为它们具有更高的一致性,使得例如调控过程容易得多。同时,本发明使得能够使用此类大批的免疫效应细胞,这是因为先前出于安全方面的考虑,必须基于与设想的接受者至少部分匹配而产生单个批次。然而本发明显示,如本发明定义的免疫效应细胞(超出单倍体相同的错配)可安全用于免疫疗法并且它们显示出功效。优选地,用于根据本发明使用的组合物进一步包含至少一种赋形剂,例如像用于输注的水、生理盐溶液(0.9%nacl)、或细胞缓冲液,优选地由经蛋白组分(例如人血清白蛋白(has))取代的生理盐溶液组成。为了使用本发明的组合物,例如在非单倍体相同的错配的情况下,优选的是用于根据本发明使用的组合物中b细胞或t细胞数目低,从而避免移植物抗宿主病。在一个优选实施例中,用于根据本发明使用的组合物并不导致移植物抗宿主病。优选地,该组合物包含不多于5%t细胞和不多于5%b细胞,更优选地不多于2%t细胞和不多于2%b细胞,最优选地少于1%t细胞和少于1%b细胞。在一个优选实施例中,提供了用于根据本发明使用的组合物,其中免疫效应细胞除了对car呈阳性外,还对神经细胞黏附分子(ncam)呈阳性。神经细胞黏附分子(ncam)是在神经元、神经胶质、骨骼肌和自然杀伤细胞的表面上表达的免疫球蛋白(ig)超家族的糖蛋白。ncam(也称为cd56)已经被认为在细胞-细胞黏附、神经突增生、突触可塑性、以及学习和记忆方面具有作用。优选地,ncam用于定义用于根据本发明使用的分化的免疫效应细胞的群体,并且可以用于将输注的效应细胞与患者外周血中的自然杀伤细胞区分。优选地,用于根据本发明使用的组合物包含多于90%cd56+细胞,更优选地多于95%cd56+细胞,最优选地多于98%cd56+细胞。典型地,本发明的组合物包含多种细胞。组合物中的所有细胞不是必须具有本发明定义的特征和效果。然而,优选的是在用于根据本发明使用的组合物中具有如本发明中定义的至少某一百分比的免疫效应细胞,从而在效率(在生产期间)和功效(在临床中)之间达到正确的平衡。在一个优选实施例中,提供了用于根据本发明使用的组合物,其中该组合物包含多种细胞,其特征在于该多种细胞中30%-100%、优选地30%-90%、更优选地30%-80%、更优选地30%-70%、更优选地30%-60%、更优选地30%-50%、最优选地30%-40%是如本发明定义的car-nk细胞。优选地,包含多种细胞的组合物的特征在于该多种细胞中40%-100%、更优选地50%-100%、更优选地60%-100%、更优选地70%-100%、更优选地80%-100%、最优选地90%-100%是如本发明定义的car-nk细胞。在用于根据本发明使用的组合物内,如本发明定义的car-nk细胞的其他优选范围是:40%-90%、50%-90%、60%-90%、70%-90%、80%-90%、40%-80%、50%-80%、60%-80%、40%-70%、40%-60%、50%-60%或40%-50%。出于生产效率,如本发明定义的更低百分比的car-nk细胞是希望的,而另一方面,出于临床功效和调控原因,如本发明定义的更高百分比的car-nk细胞是希望的。从调控角度而不是从效率角度来看,优选的是从单个供体获得用于根据本发明使用的组合物。甚至更优选的是,单个供体提供了多于一个治疗剂量,使得大规模批次可以生产、批准或认证,并且在必须用根据本发明使用的组合物治疗随机个体的情况下可以成品使用。在一个优选实施例中,提供了用于根据本发明使用的组合物,其中多种细胞源自从单个供体获得的细胞。优选地,该多种细胞源自脐带血和骨髓(因为这些是cd34阳性干细胞和/或祖细胞的丰富来源)中的至少一种。由于可能使用包含car-nk细胞的成品组合物,用于根据本发明使用的组合物将细胞过继性疗法从个性化医学向更通用的药物进一步推进,因为不再需要寻找单个供体来匹配单个接受者。这还有益地影响了治疗的成本。如本文所用的“成品(off-the-shelf)”意指制备并且储存以在需要时直接使用的组合物。具体地,可用于“成品”的组合物不是针对特定接受者产生,而是通常可以在不同时间点用于不同接受者。例如,如本发明定义的组合物可以冷冻,并且当需要时,如本发明定义的解冻和使用。如本发明定义的组合物使得能够实现大规模生产gmp产生的免疫效应细胞,这些免疫效应细胞理论上可以在任何随机接受者需要时的几分钟内提供。在一个实施例中,本发明提供了包含car-nk细胞的组合物,其中在用于制造用嵌合抗原受体(car)遗传修饰的nk细胞的群体的方法中离体产生该组合物,该方法包括:(i)第一阶段:根据以下步骤产生含有cd34+细胞的中间体细胞群体,这些细胞携带编码car的至少一种多核苷酸:a)获得选自由以下组成的组的生物起始样品:骨髓、外周血、胎盘或脐带血,b)从此类生物样品分离造血干细胞,c)在培养基i的存在下,培养分离的造血干细胞,d)通过将培养的干细胞与携带编码car的至少一种多核苷酸的慢病毒载体一起孵育来转导此类细胞,从而获得含有car-cd34+干细胞的中间体细胞群体,以及e)任选地,在培养基i中培养该含有car-cd34+干细胞的中间体细胞群体至少一天,其中培养基i是包含细胞因子的集合的基本培养基,其中所述细胞因子的集合包含白介素-7,和干细胞因子(scf)、flt-3配体(flt-3l)、血小板生成素(tpo)中的一种或多种,以及粒细胞-巨噬细胞集落刺激因子(gm-csf)、粒细胞集落刺激因子(g-csf)和白介素-6(il-6)中的两种或更多种,(ii)第二阶段,该第二阶段包括:任选的预备步骤:在培养基ii中进一步扩增来自该第一阶段的该含有car-cd34+干细胞的中间体细胞群体,由此获得含有car-cd34+干细胞和car-nk祖细胞的细胞群体,其中培养基ii是包含细胞因子的集合的基本培养基,其中所述细胞因子的集合包含scf、flt-3l白介素-15(il-15)和il-7中的两种或更多种,以及gm-csf、g-csf和il6中的两种或更多种,以及将来自阶段i的该含有car-cd34+干细胞的中间体细胞群体、或来自该任选的预备步骤的该含有car-cd34+干细胞和car-nk祖细胞的细胞群体扩增和分化成含有car-nk细胞的细胞群体的步骤,该步骤包括在培养基iii中培养来自阶段i的含有car-cd34+干细胞的中间体细胞群体、或来自该任选的预备步骤的含有car-cd34+干细胞和car-nk祖细胞的细胞群体,由此获得含有nk细胞的细胞群体,其中该培养基iii是包含细胞因子的集合的基本培养基,其中所述细胞因子的集合包含scf、il-7、il-15和白介素-2(il-2)中的两种或更多种,以及gm-csf、g-csf、和il-6中的两种或更多种。可以按任何可能的方式获得包含造血干细胞和/或祖细胞的样品组合物,该方式例如像获得或收集含有干细胞和/或祖细胞的细胞来源,例如骨髓、脐带血、胎盘材料、外周血、用干细胞动员剂治疗的人的外周血、使用细胞培养步骤从胚胎干细胞或其任何衍生物离体产生的外周血、或使用细胞培养步骤从诱导性多能干细胞及其任何衍生物离体产生的外周血。可以使用亲和纯化方法,从含有此类干细胞和/或祖细胞的细胞来源进一步纯化造血干细胞和/或祖细胞。术语“离体”意指过程或方法并不是在活的个体内进行的,而是例如在能够培养细胞的装置内、优选地是开放或者封闭的细胞培养装置(例如培养瓶、一次性袋或生物反应器)中进行。术语“cd34+干细胞”意指在细胞表面上表达cd34抗原的多能干细胞,优选地是能够发育成所有特定类型的血液细胞的干细胞,并且更优选地是可以产生血统的谱系特异性祖细胞的细胞。术语“cd34+祖细胞”意指在细胞表面上表达cd34抗原的多能祖细胞,优选地是能够发育成各种类型的血液细胞的祖细胞,并且更优选地是可以产生某些血统的谱系特异性祖细胞的细胞。如本文所用的术语“亲和纯化”意指通过以下方法标记待纯化的细胞:例如出于分离目的靶向特定目的表位,例如用与适合由分离方法检测的药剂偶联的抗体靶向抗原,使用例如与用于纯化方法(例如荧光激活细胞分选(facs))的荧光染料偶联的抗体,和/或使用例如与用于磁性选择程序的磁性颗粒偶联的抗体。亲和纯化方法是本领域已知的,并且例如可以是基于(例如抗原和抗体之间的、酶和底物之间的、或受体和配体之间的)高特异性相互作用分离生物化学混合物的任何方法。如本文所用的术语“扩增”意指由于由细胞培养步骤引起的细胞分裂事件造成的细胞扩增,优选地基本上不改变细胞的表型(这通常称为分化)。短语“基本上不改变细胞的表型”意指细胞优选地并不改变其功能、其细胞表面标记和/或其形态学。如本文所用的术语“分化”意指改变细胞的表型,这意味着在细胞培养过程期间改变某些细胞表面分子的表达、改变细胞功能和/或改变细胞的形态学,其中优选地,细胞由于添加了细胞培养基而仍然可以扩增。如先前所示,发明人已经表明,用于如本发明定义的免疫疗法的组合物对于治疗肿瘤而言是特别有用的。根据一个优选实施例,用于根据本发明使用的组合物被用于治疗肿瘤。在本发明的意义内,肿瘤包括造血系统肿瘤或实体肿瘤。肿瘤可以是恶性的或良性的。用于在根据本发明的免疫疗法中使用的组合物可以用于治疗肿瘤的不同阶段,特别是在治疗造血系统肿瘤(例如像急性骨髓性白血病(aml))中。例如,如本发明例示的,组合物优选地可以在无法进行骨髓移植的那些(老年)患者中用作巩固疗法。另外,如使用另一种治疗的其他实例所示,根据本发明的免疫效应细胞疗法优选地可以用于诱导疗法未完全缓解的患者(难治性患者)或在诱导疗法后很短时间复发的那些患者(复发性患者)。将免疫效应细胞疗法并入其他巩固疗法也是可行并且优选的,例如在同种异体hstc方案中,另外使用如本发明定义的免疫效应细胞。在一个优选实施例中,提供了用于根据本发明使用的组合物,其中待在一次治疗中施用的组合物包含至少5x105个细胞,并且不多于5x1011个细胞。可以通过任何可接受的方法施用本发明的组合物,条件是免疫效应细胞能够达到其在个体中的靶标。例如,可以经由静脉内途径或经由局部途径(包括但不限于眼、皮肤、肺、颊和鼻内途径)施用本发明的组合物。如本文所用的局部途径还意指任何直接局部施用(例如像在骨髓中),但也指直接注射入例如实体肿瘤中。在特定情况下,例如,如果免疫疗法旨在影响胃肠道的粘膜层,则可以使用口服途径。优选地,提供了用于根据本发明使用的组合物,其中通过静脉内途径或通过局部途径或通过口服途径或通过这些途径的任何组合来施用该组合物。如本文所用的术语“局部”意指局部地、优选地在肿瘤部位(可以位于任何解剖学部位,更特别地,肿瘤可以位于骨髓或任何其他器官)施用免疫效应细胞。可以施用一次用于根据本发明使用的组合物,但是如果认为有必要,则可以多次施用该组合物。这些可以是一天多次、一周多次或甚至一月多次。还可能的是,首先等待第一次施用(例如输注)的临床结果,并且如果认为有必要,在组合物无效的情况下进行第二次施用,并且甚至进行第三次施用、第四次施用等等。在一个优选实施例中,提供了用于根据本发明使用的组合物用于治疗肿瘤,其中该肿瘤是造血系统肿瘤或淋巴肿瘤,或其中该肿瘤是实体肿瘤。术语“血液肿瘤”、“造血系统肿瘤”或“淋巴肿瘤”意指这些是造血组织或淋巴组织的肿瘤。造血系统恶性肿瘤和淋巴恶性肿瘤是影响血液、骨髓、淋巴、和淋巴系统的肿瘤。在肿瘤是造血系统肿瘤或淋巴肿瘤的那些情况下,提供了用于根据本发明使用的组合物,其中该肿瘤是以下中的一种或多种:白血病、淋巴瘤、骨髓增生异常综合征或骨髓瘤,优选地是选自以下的白血病、淋巴瘤或骨髓瘤:急性骨髓性白血病(aml)、慢性骨髓性白血病(cml)、急性t细胞白血病、急性成淋巴细胞性白血病(all)、慢性成淋巴细胞性白血病(cll)、急性单核细胞白血病(amol)、套细胞淋巴瘤(mcl)、组织细胞性淋巴瘤或多发性骨髓瘤,优选地aml。在肿瘤是实体肿瘤的那些情况下,提供了用于根据本发明使用的组合物,其中该肿瘤是以下中的一个:腺癌、鳞状细胞癌、腺鳞癌、未分化癌、大细胞癌或小细胞癌、肝细胞癌、肝母细胞瘤、结肠腺癌、肾细胞癌、肾细胞腺癌、结肠直肠癌、结肠直肠腺癌、胶质母细胞瘤、神经胶质瘤、头颈癌、肺癌、乳腺癌、默克尔细胞癌、横纹肌肉瘤、恶性黑色素瘤、表皮样癌、肺癌、肾癌、肾腺癌、乳房癌、乳房腺癌、乳腺导管癌、非小细胞肺癌、卵巢癌、口腔癌、肛门癌、皮肤癌、尤因肉瘤、胃癌、尿道癌、子宫癌、子宫肉瘤、阴道癌、外阴癌、维尔姆斯瘤、瓦尔登斯特伦巨球蛋白血症、胰腺癌、胰腺腺癌、子宫颈癌、鳞状细胞癌、髓母细胞瘤、前列腺癌、结肠癌、结肠腺癌、移行细胞癌、骨肉瘤、导管癌、大细胞肺癌、小细胞肺癌、卵巢腺癌、卵巢畸胎瘤、膀胱乳头状瘤、成神经细胞瘤、多形性成胶质细胞瘤、胶质母细胞瘤、星形细胞瘤、上皮样癌、黑色素瘤或视网膜母细胞瘤的恶性肿瘤或转移性诱导性继发性肿瘤。在一个优选实施例中,提供了用于根据本发明使用的组合物,其中该实体肿瘤选自宫颈癌的恶性肿瘤或转移性诱导性继发性肿瘤,这些宫颈癌选自:腺癌、鳞状细胞癌、腺鳞癌、子宫颈癌、小细胞癌、和黑色素瘤。在另一个优选实施例中,提供了用于根据本发明使用的组合物,其中该实体肿瘤选自结肠直肠癌症的恶性肿瘤或转移性诱导性继发性肿瘤,这些结肠直肠癌症选自:腺癌、鳞状细胞癌、结肠腺癌、结肠直肠癌、结肠直肠腺癌、结肠癌、和黑色素瘤。相对于迄今为止已知的治疗选择,本发明的组合物具有若干优点。本发明的组合物是有利地,不依赖于hpv类型、肿瘤病史、肿瘤egfr表达和kras状态。除此之外,本发明的免疫效应细胞还克服了hla-e、hla-g和(ido)抑制,由此导致针对肿瘤(尤其是针对宫颈癌和结肠直肠癌症)的增强的抗肿瘤效果。如通常所述,术语“表皮生长因子受体”或egfr是指在几乎所有健康组织中广泛表达的细胞表面蛋白。egfr蛋白由跨膜糖蛋白编码,并且是蛋白激酶家族的成员。egfr的过表达及其下游信号传导途径中的突变与若干实体肿瘤(像结肠、肺和子宫颈)中的不良预后相关联。术语柯尔斯顿(kirsten)大鼠肉瘤病毒癌基因(kras)是指主动参与调节正常组织信号传导(egfr下游信号传导途径的一部分)的基因。然而,已经报道了kras基因在结肠、直肠和肺的实体肿瘤的肿瘤细胞中的突变。发生在多于50%的结肠直肠癌症患者中的此激活突变帮助肿瘤细胞逃避egfr靶向药物,像西妥昔单抗和帕尼单抗。如本文所用的术语“人乳头状瘤病毒(hpv)”是指在女性中引起宫颈癌的病毒组。hpv病毒影响嘴、喉、外阴、子宫颈和阴道周围的皮肤和湿膜。hpv感染引起会导致子宫颈中癌症的异常细胞变化。如本文所用的术语吲哚胺2,3双加氧酶(ido)是指在将氨基酸l-色氨酸降解为n-甲酰犬尿氨酸中充当催化剂的酶。报道了ido通常在前列腺、胃、卵巢、子宫颈和结肠的实体肿瘤中过表达,使得肿瘤细胞能够逃避被细胞毒性t细胞和nk细胞杀死。在以下非限制性实例中更详细地描述了本发明。实例实例1-lngfr间隔的cd44v6car的描述本实例中使用的car构建体是wo2016/042461中披露的cd44v6-nwl,该文献包括此类构建体的序列的详细描述。从创造者处购买了编码carcd44v6-nwl的cdna。在图1中显示了car构建体cd44v6-nwl的示意性结构图。carcd44v6-nwl由以下组成:cd44v6结合结构域、lngfr野生型长间隔子(包括四个tnfr-cys结构域和整个富含丝氨酸苏氨酸的柄)、cd28的跨膜和共刺激结构域、以及cd3ζ链的胞内结构域。以下报道了carcd44v6-nwl的蛋白质序列(序列idn°14):图例粗体:cd44v6特异性单链片段斜体加下划线:lngfr间隔子粗体加下划线:cd28跨膜和共刺激结构域斜体:cd3ζ链信号传导结构域实例2-阶段1:产生中间体car-cd34+干细胞群体本发明的方法的第一阶段提供了产生含有cd34+干细胞的中间体细胞群体的条件,这些干细胞携带至少一种编码car的多核苷酸。通过以下进行干细胞的工程化:用tat独立的sin慢病毒载体(携带含有人磷酸甘油酸激酶(hpgk)启动子的表达盒)转导,以驱动实例1中披露的carcd44v6-nwl的组成型表达。实验包括使用以下不同条件对从ucb分离的cd34+细胞进行转导:-条件a:基础细胞培养基(gmp干细胞生长培养基,获得自cellgenix股份有限公司(cellgenixgmbh),弗莱堡,德国),补充有细胞因子(300ng/mlflt3-l和scf、100ng/mltpo和60ng/mlil3)。条件a是应用除了本发明的那些条件以外的转导条件的对照方法。-条件b:基础细胞培养基:glycostem基础生长培养基(gbgm,获得自格雷克斯迪姆医疗公司(glycostemtherapeutics),奥斯,荷兰),补充有10%人血清、细胞因子(10pg/mlgm-csf、250pg/mlg-csf、50pg/mlil-6、25ng/mlscf、25ng/mlflt-3l、25ng/mltpo、25ng/mlil-7)和20μg/ml肝素。条件b是本发明的方法的阶段1的代表性实例。实验目的是鉴定最有效的条件,以获得含有在制造方法的第二阶段中待扩增并且分化成car-nk细胞的car-cd34+干细胞的中间体细胞群体。以下总结了完整实验方案:表1:实验条件表2中披露了用于cd34+转导方法的主要步骤。表2:实验程序2.2起始生物材料为了进行实验,从nhs血液和移植司(bloodandtransplant)提供的三种脐带血(ucb)中分离cd34+细胞(表3)。表3:ucb数据2.3cd34分离和表征对三种ubc(由nhs血液和移植司供应)进行独立加工;根据提供者的标准条件,使用人工密度梯度(用lymphopreptm)分离单核细胞(mnc)。另外,将免疫磁性选择应用于每个样品,以获得cd34+群体:根据提供者的条件,将细胞首先用缀合至磁珠的抗cd34抗体进行免疫标记,并且然后加载到miltenyilsmidimacs柱中。选择在cd34+细胞%方面具有高纯度的样品用于实验,并且将其他细胞冷冻。在表4中报道了在cd34+细胞恢复方面的细胞数目、活力和总加工产率。表4:细胞数目、活力和恢复在控制温度速率的冰箱中冷冻从ucb1和ucb3回收的cd34+细胞。通过facs分析评估ucb样品的免疫分型。在bdfacscanto2上进行获取,并且使用diva软件分析数据。表5示出了预操作以及ucb样品1、2和3在第0天的阴性级分的结果。表6示出了ucb样品1、2和3在第0天的阳性级分的结果。表5:ucb预操作和阴性级分(nf)的免疫分型数据表6:阳性级分的免疫分型数据选择来自ucb样品2的阳性级分用于转导。2.4cd34+接种在取样用于if分析和克隆生成测定后,将从ucb2回收的细胞按1.0x106个细胞/ml的接种浓度接种在非组织培养物处理的板(预先涂覆有retronectin)中(表7)。2.5cd34+转导在第1天,在22h45’孵育后,从板收集一半细胞悬浮液,离心并且用lvcar-cd44v6载体pde_b17100悬浮(第1次转导运行)。15小时的孵育后,收集细胞并且计数,然后将这些细胞离心并且在培养基1x细胞因子中按1.0x106个细胞/ml的接种浓度悬浮。将以下等式用于计算培养物的生长速率:表7:第1次转导运行时的细胞浓度、活力和生长速率(第2天)在第3天,收集细胞并且计数。还通过乘以个体生长速率计算从(转导结束)第0天至第3天获得的总体生长速率。表8:第2次转导运行时的细胞浓度、活力和生长速率(第3天)在转导结束时,收集条件a的细胞并且转移到离心管中;在用gmp干细胞生长培养基洗涤孔一次后,将细胞离心、悬浮在新鲜的gmp干细胞培养基中、计数并且取样用于测试。将剩余细胞冷冻在冷冻小瓶中的人血清+格雷克斯迪姆(glycostem)基础培养基14%dmso(1:1)中。收集条件b的细胞,计数并且取样用于测试,如表6中所述。将剩余细胞离心并且按0.4x106个细胞/ml的接种浓度重新悬浮在培养基1x细胞因子中。在第5天,将条件b的细胞计数,并且添加1体积的新鲜培养基。表9:样品b的细胞浓度、活力和生长速率(第5天)在第7天,收集条件b的细胞并且转移到离心管中;在用gly培养基洗涤孔一次后,将细胞离心、重新悬浮在新鲜的gly培养基中(使用两倍的细胞培养体积)用于ds配制品、计数并且取样用于测试,如表6中所述。将剩余细胞冷冻在冷冻小瓶中的人血清+gly培养基14%dmso(1:1)中。还通过乘以个体生长速率计算从第0天至第7天获得的总体生长速率。表10:样品b的细胞浓度、活力和生长速率(第7天)通过facs分析评估免疫分型。在bdfacscanto2上进行获取,并且使用diva软件分析数据。在某天(条件a和条件b)和第7天(条件b)分析细胞。表11:转导的细胞的免疫分型数据2.6cd34+细胞克隆形成能力根据药典专论ep2.7.28中披露的方法(人造血祖细胞的集落形成测定),造血祖细胞的克隆形成能力。在细胞解冻(第0天)时进行分析,用于确定基础克隆形成潜能,并且在转导阶段结束时(第3天)进行分析。表12:在第0天的克隆形成能力结果表13:在转导阶段结束时(第3天)的克隆形成能力结果2.6.1转导效率通过对ngfr标记的facs分析评估转导效率。在液体培养后并且针对来自克隆生成测定的集落群(pool)分析细胞。表14:lc后的转导效率表15:集落群的转导效率2.6.2vcn测定通过在转导的cd34+细胞的14天髓系培养后进行定量pcr,确定载体拷贝数。表16中汇总了结果。表16:vcn结果结论细胞活力和生长速率:在所有方法(≥96%)期间,细胞维持了良好活力,并且两种条件下的生长速率相似。免疫分型:通过分析发现,在干细胞表征方面没有差异,除了在根据条件a培养和转导的细胞中cd15标记的表达更高,这指示细胞群体更倾向于定型为髓系谱系。在根据条件b培养和转导的细胞(即根据本发明的方法产生的中间体细胞群体)中没有观察到这些表型。克隆形成能力:在第3天的cd34+细胞的克隆形成能力是类似的。转导效率和载体拷贝数:在条件a中的转导效率和vcn高于在条件b中。实例3-阶段2:car-cd34+干细胞的细胞扩增和分化成car-nk细胞3.1解冻在由补充有2.5mmmgcl2和0.13mg/mldna酶的人血清白蛋白组成的解冻缓冲液中,将来自条件a(第3天)和条件b(第7天)的冷冻保存的cd44v6转导的祖细胞和模拟物祖细胞解冻。在新鲜的扩增培养基i中,按0.97x10^6个细胞/ml(条件a)、2.11x10^6个细胞/ml(条件b,非t)和1.99x10^6个细胞/ml(条件b,t)的细胞浓度,将cd34+ucb细胞铺板在组织培养物处理的6孔板中。在冷冻-解冻循环后,cd34+ucb细胞恢复良好,如表17中描绘,其显示来自最初冷冻的总细胞中活细胞百分比。表17.第3天(条件a)和第7天(条件b)的祖细胞的重构。3.2扩增阶段在扩增阶段中测量细胞活力、cd34含量和lngfr表达的流式细胞术数据,以监测和提供最佳细胞培养条件。在新鲜的扩增培养基i(基础培养基(gbgm),其补充有10%人血清、低剂量细胞因子混合物(含有10pg/mlgm-csf,250pg/mlg-csf和50pg/mlil-6,25ng/mlscf、flt-3l、tpo、il-7)、以及20μg/ml肝素)中培养cd34+ucb细胞。在扩增培养基i中培养来自条件a和b的细胞直至第9天,从第4天(解冻来自条件a的细胞后)和第8天(来自条件b的细胞)进行计数。每周每2-3天更换扩增培养基i。在第10天,通过添加扩增培养基ii(在此用25mg/mltpo替换20ng/mlil-15)培养祖细胞。从解冻之日(扩增设置为1)开始,在第3天(条件a)或第7天(条件b)计算所有条件中的真实扩增,并且已经显示在条件bt和b非t之间没有差异。对于条件at和a非t,在a非t中测量到比at中更低的扩增。通过排除对扩增的冷冻-解冻作用,计算理论扩增,显示了针对所有条件的最佳扩增。所有条件之间的比较结果表明,条件a(特别是a非t)的扩增更低。在培养第10天至第21天期间,来自lngfr表达的结果显示天然cd271在细胞中表达。3.3分化阶段在分化期间,测量作为nk细胞的标记的cd56含量,而不是cd34含量以及细胞活力和lngfr表达。分化培养基由以下组成:补充有2%人血清、低剂量细胞因子混合物(含有10pg/mlgm-csf,250pg/mlg-csf和50pg/mlil-6,20ng/mlscf、il-15、il-7,和1000u/mlil-2(阿地白介素(proleukin)))的gbgm。每周两次更换分化培养基,直至培养结束。在cd44v6转导的ucb-nk细胞中的lngfr表达显示在条件a转导的nk细胞中的高ngfr表达(在第41天87%),并且介导条件b转导的nk细胞中的ngfr表达(在第45天37%)。来自条件acd44v6转导的ucb-nk的形态学结果表明,与所有其他条件相比,在培养物中的细胞扩大。3.4体外功能性测定针对k562、t98g和thp-1肿瘤细胞系(对于nk细胞的细胞毒性,分别是敏感的、中间的和具有抗性的),在来自条件a和b的模拟物和cd44v6转导的ucb-nk细胞的培养结束时进行体外功能性测定。按2x10^5个细胞/ml的细胞密度,用25nm佛波酯(phorbolmyristateacetate,pma)刺激thp-1人白血病单核细胞48h,并且用新鲜培养基再孵育24h,从而在已知表达cd44的巨噬细胞中分化thp-1细胞(图4a)。在37℃下,按1x10^7个细胞/ml的浓度,用5μm太平洋蓝琥珀酰亚胺基酯(pbse)标记靶细胞10分钟。将靶细胞在靶培养基中洗涤并且浓缩至5x10^5个细胞/ml。将nk细胞也浓缩至5x10^5个细胞/ml,并且在过夜测定中与靶细胞(100μl效应细胞+100μl靶标)共培养。对于脱粒测量,在孵育开始时添加抗cd107a,并且在孵育结束时添加用于nk细胞鉴别的抗cd56。在1:1的效应细胞:靶细胞比率下,基于凋亡7aad活力标记的流式细胞术读出来计算细胞毒性。与其他条件相比,针对敏感性肿瘤细胞系k562,观察到来自条件a的cd44v6转导的ucb-nk细胞的更低细胞毒性。3.5nk细胞中的受体表达在培养结束时,针对nk细胞杀伤机制中涉及的抑制(nkg2a)和激活(dnam-1、nkg2c、nkg2d、cd16)受体表达,筛选来自条件a的模拟物和cd44v6转导的ucb-nk细胞(图5a)。测量到cd44v6转导的ucb-nk细胞与模拟物ucb-nk细胞相比更低的dnam-1激活受体表达。3.6运送至莫尔米德公司(molmed)将来自条件a(第31天)和条件b(第35天)的新鲜的模拟物和cd44v6转导的ucb-nk细胞运送至莫尔米德公司(molmed),用于功能活性的进一步表征和用nsg小鼠的体内研究。在补充有50%人血清和7%dmso的最终浓度的gbgm中,在第34天(条件a)和第38天(条件b)冷冻剩余细胞,以研究解冻后的模拟物和cd44v6转导的ucb-nk细胞在重新获得其功能能力方面的恢复。3.7重构在解冻缓冲液中解冻来自条件a(第34天)和b(第38天)的冷冻保存的模拟物和cd44v6转导的ucb-nk细胞(描述于3.1中),并且在良好的恢复后用于如3.4中所述的体外功能性测定。针对敏感性肿瘤细胞系k562,使用来自条件a和b的模拟物和cd44v6转导的ucb-nk细胞的体外功能性测定的结果显示了与重构之前的功能性数据相当的结果(图4b)。针对k562,来自条件a的cd44v6转导的ucb-nk细胞中仍观察到更低细胞毒性。针对敏感性肿瘤细胞系k562,分析在流式细胞仪上通过百分比cd107a+测量的模拟物和cd44v6转导的ucb-nk细胞的脱粒(图4c)。为了比较在重构前和重构后的nk细胞杀伤机制中涉及的抑制和激活受体表达,针对相同重要的nk细胞激活和抑制性受体(添加kir2dl2/l3、kir3dl和kir2dl1抑制性受体),筛选重构后来自条件a和b的模拟物和cd44v6转导的ucb-nk细胞(图5b)。在重构后,在来自条件a的cd44v6转导的ucb-nk细胞中,测量到dnam-1和nkg2d的激活受体表达更低。实例4-模拟物nk和car-nk细胞的功能活性的体外表征为了评估cd44v6-carnk细胞的抗原特异性活性,我们进行了基于流式细胞术的效力测定,在用表达或不表达cd44v6抗原的靶细胞系刺激的细胞中测量了脱粒和细胞因子的产生。4.1实验设计在最后一次改变培养基后第3天,在条件b(即根据本发明的方法)下和在条件a(即对照)下产生的模拟物nk和cd44v6-carnk中,分析脱粒(作为cd107a+细胞的百分比)和细胞因子的产生(作为tnf-α+细胞的百分比)。作为靶细胞,将cd44v6阴性和阳性肿瘤细胞(两者都对nk细胞的细胞溶解敏感(k562,慢性骨髓性白血病)和具有抗性的(molt-4,急性成淋巴细胞性白血病))在1∶1的e∶t比率下使用。此外,将效应细胞保持未处理、或用佛波酯(pma)和离子霉素(iono)刺激,以分别检测基础细胞激活或非特异性激活。4.2材料在96孔圆底板中,按100μl的最终体积,进行效力测试。进行了效应细胞的facs分析,在表18中报道了抗体组。表18:用于nk细胞染色的抗体和活力染色组标记荧光克隆目录号公司cd271pehi100557196bd公司cd56percpcy5.5b159560842bd公司tnf-αpe-cy7mab11557647bd公司cd107aapch4a3130-100-314美天旎公司cd45apch72d1641417bd公司可固定的活力菌株510510n.a.564406bd公司为了产生cd44v6阳性k562和molt-4细胞系,我们用表达cd44v6抗原的逆转录病毒载体进行了亲本细胞系的转导。4.3结果在培养的第38天(条件a;82%lngfr+细胞)或在培养的第42天(条件b;33%lngfr+细胞),针对体外功能活性,对条件a(对照)和条件b(即根据本发明的方法)中获得的模拟物nk和cd44v6-carnk进行测试。将所有效应细胞和靶细胞计数,并且重新悬浮在补充有人igg的rpmi1640中,并且在冰上孵育20min。离心后,在rpmi+10%fbs(单独地或与不同靶细胞系以1:1的e:t比率)中培养效应细胞(0.2x106个细胞/条件)。对于细胞功能性,用pma/iono刺激细胞,作为阳性对照。在cd107aab、布雷菲德菌素和/或莫能菌素存在下,在6或18小时孵育后,将nk用适当组的荧光ab进行表面抗原染色,如表18中报道。然后根据制造商对胞内tnf-α染色的说明,将样品固定并透化,并且通过流式细胞术进行分析。用不同靶标刺激的活阳性效应细胞的百分比(在减去单独孵育的阳性效应细胞的百分比后)显示于图2、小图a(在18小时处,cd107a阳性细胞的百分比)、和小图b(在6小时处,tnf-α阳性细胞的百分比)中。在lngfr阳性和阴性细胞的两个亚群中,进行carnk激活的分析。模拟物和转导的nk细胞两者(条件a和b两者)识别nk敏感性k562细胞系(虽然来自条件a的cd44v6-carnk细胞显示了相对于模拟物nk的功能降低),但是在使用k562cd44v6细胞的激活中,仅car-nklngfr+细胞显示了清楚的特异性增加(两者都分别考虑了cd107和tnf-α阳性)。通过nk细胞与抗性molt-4细胞系的共培养,证实了这种由cd44v6抗原的carnk导致的特异性激活。当与表达cd44v6抗原的molt-4细胞一起孵育时,可检测的激活(脱粒和tnf-α产生两者)仅发生在carnklngfr+细胞中(来自条件a和b两者)。实例5-nk和car-nk细胞的体内功效5.1实验设计在aml异种肿瘤模型中,评估cd44v6car-nk细胞的抗肿瘤活性。实验包括在nsg小鼠中的输注,该小鼠荷载thp1白血病细胞(最小残余疾病设置),根据实例2(在条件a或条件b下,脐带血来源的干细胞的转导)和实例3(扩增和分化成car-nk细胞或nk细胞)中描述的条件产生的nk细胞。该方法(包括在条件b下在实例2中的转导,随后在实例3中扩增和分化成nk细胞)是本发明的方法的代表性实例。在第0天,用thp1癌细胞输注小鼠,并且在三天后,用过继性免疫疗法治疗这些小鼠。实验包括七个实验组(表19)表19:实验条件如表20中所示,详细总结了实验程序表20:实验程序5.2材料thp1肿瘤细胞对于体内功效实验,将thp1细胞系(源自急性单核细胞白血病患者的人单核细胞系)输注到小鼠中,从而实现肿瘤移植。在rpmi+10%fbs、100u/ml青霉素、100u/ml链霉素、1mm丙酮酸钠和50μmβ巯基乙醇中,培养和扩增thp1肿瘤细胞12天。在第0天,在尾静脉中输注总计1.5e+06个细胞/小鼠。未转导的和cd44v6car转导的nk细胞(条件a和b)为了进行体内功效实验,产生在不同条件下(表18)培养的nk细胞,并且将这些细胞保持在基础培养基(gbgm)(补充有2%人血清、低剂量细胞因子混合物(含有10pg/mlgm-csf,250pg/mlg-csf和50pg/mlil-6,20ng/mlscf、il-15、il-7,和1000u/mlil-2(阿地白介素))中培养,并且输注到小鼠中,如下所示(表21)。表21:输注到小鼠中的细胞总数*在免疫分型分析的基础上计算nk和car-nk阳性细胞的数量;nk细胞被定义为cd45+/cd56+/cd3-/7aad-;carnk为cd271+/cd45+/cd56+/cd3-/7aad-nsg小鼠模型由查尔斯河实验室(charlesriverlaboratories)提供了8周龄的nod.cg-prkdcscidil2rgtm1wjl/szj(nsg)转基因小鼠。在第0天,用1.5e+06个thp1细胞输注小鼠用于肿瘤移植。输注两天后,将小鼠随机分为来自两个不同小鼠队列(雄性/雌性)的五个组(五个小鼠/组),并且用不同细胞制剂进行处理。在性别匹配的队列中,施用来自条件a(雄性队列)和b(雌性队列)的nk细胞和carnk。在第3-6-9天给予nk细胞(分别是第1、2和3次输注)。对照小鼠(雄性队列)保持未处理。在第3-6-9-12-15-18天,提供重组hil-15(il15)作为支持。重组hil-15(il15)按0.1mg/ml的浓度,用无菌水重构il-15(免疫工具公司(immunotool)),并且将溶液等分并且在-20℃下冷冻。(i)方法免疫分型和活力通过用台盼蓝染色液计数细胞来评估细胞活力。通过facs分析,针对cd45-cd3-cd8-cd56-cd16-cd271表达来表征细胞。通过排除7aad阳性细胞,确定cd56+和cd56+cd271+细胞的群体。在bdfacscanto2上进行获取,并且使用diva软件分析数据。在每个输注日分析来自条件a和b的细胞。体内thp1和治疗性细胞输注对于体内功效实验,收获thp1肿瘤细胞、nk细胞和t细胞,将这些细胞用台盼蓝染色液计数(针对细胞活力),并且通过在380g下离心10分钟、用pbs洗涤两次。按希望的浓度,将细胞重新悬浮在0.9%盐水溶液中,并且在小鼠尾静脉中输注。il15处理按5μg/ml的浓度,将il15解冻并且重新悬浮在0.9%盐水溶液中。如实验程序中所示(表20),腹腔内注射0.5μg的il15(100μl/小鼠)来处理小鼠。肝切除在第39天,根据动物实验的道德准则,在co2室中将小鼠安乐死。将肝切除、在分析天平上称重、记录数据,并且获得切除的肝的图像。5.4结果在充分确立的疾病模型中,评估来自条件a和b的模拟物nk和cd44v6car-nk细胞的体内抗肿瘤作用。图3示出了针对每种条件的肝重量(克)。用单向方差分析进行统计分析。在条件b(即根据本发明的方法)下获得的cd44v6car-nk细胞产生的抗肿瘤作用比在条件a下获得的模拟物细胞或cd44v6car-nk细胞获得的抗肿瘤作用更强。结论体内结果显示,本发明的制造方法允许获得如下细胞群体,该细胞群体的特征在于与未转导的nk细胞以及用不同条件下进行的方法获得的car-nk细胞相比具有更强的抗肿瘤作用。结果是本发明的方法允许获得以nk细胞和car两者的功能效果为特征的含有car-nk细胞的细胞群体。序列表<110>格雷克斯迪姆医疗私人有限公司(glycostemtherapeuticsb.v.)<120>制造方法<130>ip-7<150>ep18175055<151>2018-05-30<160>14<170>bissap1.3.6<210>1<211>222<212>prt<213>人工序列<220><223>蛋白质序列间隔子结构域(nwl)<400>1lysglualacysprothrglyleutyrthrhisserglyglucyscys151015lysalacysasnleuglygluglyvalalaglnprocysglyalaasn202530glnthrvalcysgluprocysleuaspservalthrpheseraspval354045valseralathrgluprocyslysprocysthrglucysvalglyleu505560glnsermetseralaprocysvalglualaaspaspalavalcysarg65707580cysalatyrglytyrtyrglnaspgluthrthrglyargcysgluala859095cysargvalcysglualaglyserglyleuvalphesercysglnasp100105110lysglnasnthrvalcysgluglucysproaspglythrtyrserasp115120125glualaasnhisvalaspprocysleuprocysthrvalcysgluasp130135140thrgluargglnleuargglucysthrargtrpalaaspalaglucys145150155160glugluileproglyargtrpilethrargserthrproproglugly165170175seraspserthralaproserthrglngluproglualaproproglu180185190glnaspleuilealaserthrvalalaglyvalvalthrthrvalmet195200205glyserserglnprovalvalthrargglythrthraspasn210215220<210>2<211>666<212>dna<213>人工序列<220><223>核苷酸序列间隔子结构域(nwl)<400>2aaagaggcctgccccaccggcctgtacacccacagcggagagtgctgcaaggcctgcaac60ctgggagagggcgtggcccagccttgcggcgccaatcagaccgtgtgcgagccctgcctg120gacagcgtgaccttcagcgacgtggtgtccgccaccgagccctgcaagccttgcaccgag180tgtgtgggcctgcagagcatgagcgccccctgcgtggaagccgacgacgccgtgtgtaga240tgcgcctacggctactaccaggacgagacaaccggcagatgcgaggcctgtagagtgtgc300gaggccggcagcggcctggtgttcagttgtcaagacaagcagaataccgtgtgtgaagag360tgccccgacggcacctacagcgacgaggccaaccacgtggacccctgcctgccctgcact420gtgtgcgaggacaccgagcggcagctgcgcgagtgcacaagatgggccgacgccgagtgc480gaagagatccccggcagatggatcaccagaagcaccccccctgagggcagcgacagcacc540gcccctagcacccaggaacctgaggcccctcccgagcaggacctgatcgcctctacagtg600gccggcgtggtgacaaccgtgatgggcagctctcagcccgtggtgacacggggcaccacc660gacaat666<210>3<211>162<212>prt<213>人工序列<220><223>蛋白质序列间隔子结构域(nws)<400>3lysglualacysprothrglyleutyrthrhisserglyglucyscys151015lysalacysasnleuglygluglyvalalaglnprocysglyalaasn202530glnthrvalcysgluprocysleuaspservalthrpheseraspval354045valseralathrgluprocyslysprocysthrglucysvalglyleu505560glnsermetseralaprocysvalglualaaspaspalavalcysarg65707580cysalatyrglytyrtyrglnaspgluthrthrglyargcysgluala859095cysargvalcysglualaglyserglyleuvalphesercysglnasp100105110lysglnasnthrvalcysgluglucysproaspglythrtyrserasp115120125glualaasnhisvalaspprocysleuprocysthrvalcysgluasp130135140thrgluargglnleuargglucysthrargtrpalaaspalaglucys145150155160gluglu<210>4<211>486<212>dna<213>人工序列<220><223>核苷酸序列间隔子结构域(nws)<400>4aaagaggcctgccccaccggcctgtacacccacagcggagagtgctgcaaggcctgcaac60ctgggagagggcgtggcccagccttgcggcgccaatcagaccgtgtgcgagccctgcctg120gacagcgtgaccttcagcgacgtggtgtccgccaccgagccctgcaagccttgcaccgag180tgtgtgggcctgcagagcatgagcgccccctgcgtggaagccgacgacgccgtgtgtaga240tgcgcctacggctactaccaggacgagacaaccggcagatgcgaggcctgtagagtgtgc300gaggccggcagcggcctggtgttcagttgtcaggacaagcagaacaccgtgtgtgaagag360tgccccgacggcacctacagcgacgaggccaaccacgtggacccctgcctgccctgcact420gtgtgcgaggacaccgagcggcagctgcgcgagtgcacaagatgggccgacgccgagtgc480gaggaa486<210>5<211>200<212>prt<213>人工序列<220><223>蛋白质序列间隔子结构域(nml)<400>5lysglualacysprothrglyleutyrthrhisserglyglucyscys151015lysalacysasnleuglygluglyvalalaglnprocysglyalaasn202530glnthrvalcysgluprocysleuaspservalthrpheseraspval354045valseralathrgluprocyslysprocysthrglucysvalglyleu505560glnsermetseralaprocysvalglualaaspaspalavalcysarg65707580cysalatyrglytyrtyrglnaspgluthrthrglyargcysgluala859095cysargvalcysglualaglyserglyleuvalphesercysglnasp100105110lysglnasnthrvalcysgluglucysproaspglythrtyrserasp115120125glualaalaargalaalaaspalaglucysglugluileproglyarg130135140trpilethrargserthrproprogluglyseraspserthralapro145150155160serthrglngluproglualaproprogluglnaspleuilealaser165170175thrvalalaglyvalvalthrthrvalmetglyserserglnproval180185190valthrargglythrthraspasn195200<210>6<211>600<212>dna<213>人工序列<220><223>核苷酸序列间隔子结构域(nml)<400>6aaagaggcctgccccaccggcctgtacacccacagcggagagtgctgcaaggcctgcaac60ctgggagagggcgtggcccagccttgcggcgccaatcagaccgtgtgcgagccctgcctg120gacagcgtgaccttcagcgacgtggtgtccgccaccgagccctgcaagccttgcaccgag180tgtgtgggcctgcagagcatgagcgccccctgcgtggaagccgacgacgccgtgtgtaga240tgcgcctacggctactaccaggacgagacaaccggcagatgcgaggcctgtagagtgtgc300gaggccggcagcggcctggtgttcagttgtcaagacaagcagaataccgtgtgtgaagag360tgccccgacggcacctacagcgacgaagccgccagagccgccgacgccgagtgcgaagag420atccccggcagatggatcaccagaagcaccccccctgagggcagcgacagcaccgcccct480agcacccaggaacctgaggcccctcccgagcaggacctgatcgcctctacagtggccggc540gtggtgacaaccgtgatgggcagctctcagcccgtggtgacacggggcaccaccgacaat600<210>7<211>140<212>prt<213>人工序列<220><223>蛋白质序列间隔子结构域(nms)<400>7lysglualacysprothrglyleutyrthrhisserglyglucyscys151015lysalacysasnleuglygluglyvalalaglnprocysglyalaasn202530glnthrvalcysgluprocysleuaspservalthrpheseraspval354045valseralathrgluprocyslysprocysthrglucysvalglyleu505560glnsermetseralaprocysvalglualaaspaspalavalcysarg65707580cysalatyrglytyrtyrglnaspgluthrthrglyargcysgluala859095cysargvalcysglualaglyserglyleuvalphesercysglnasp100105110lysglnasnthrvalcysgluglucysproaspglythrtyrserasp115120125glualaalaargalaalaaspalaglucysgluglu130135140<210>8<211>420<212>dna<213>人工序列<220><223>核苷酸序列间隔子结构域(nms)<400>8aaagaggcctgccccaccggcctgtacacccacagcggagagtgctgcaaggcctgcaac60ctgggagagggcgtggcccagccttgcggcgccaatcagaccgtgtgcgagccctgcctg120gacagcgtgaccttcagcgacgtggtgtccgccaccgagccctgcaagccttgcaccgag180tgtgtgggcctgcagagcatgagcgccccctgcgtggaagccgacgacgccgtgtgtaga240tgcgcctacggctactaccaggacgagacaaccggcagatgcgaggcctgtagagtgtgc300gaggccggcagcggcctggtgttcagttgtcaggacaagcagaacaccgtgtgtgaagag360tgccccgacggcacctacagcgacgaggccgcccgggccgccgacgccgagtgcgaggaa420<210>9<211>27<212>prt<213>人工序列<220><223>cd28的蛋白质序列跨膜结构域<400>9phetrpvalleuvalvalvalglyglyvalleualacystyrserleu151015leuvalthrvalalapheileilephetrpval2025<210>10<211>68<212>prt<213>人工序列<220><223>人cd28的蛋白质序列跨膜部分和胞内部分<400>10phetrpvalleuvalvalvalglyglyvalleualacystyrserleu151015leuvalthrvalalapheileilephetrpvalargserlysargser202530argleuleuhisserasptyrmetasnmetthrproargargprogly354045prothrarglyshistyrglnprotyralaproproargasppheala505560alatyrargser65<210>11<211>41<212>prt<213>人工序列<220><223>cd28的蛋白质序列胞内结构域<400>11argserlysargserargleuleuhisserasptyrmetasnmetthr151015proargargproglyprothrarglyshistyrglnprotyralapro202530proargaspphealaalatyrargser3540<210>12<211>113<212>prt<213>人工序列<220><223>人cd3ζ链的蛋白质序列信号传导结构域<400>12argvallyspheserargseralaaspalaproalatyrglnglngly151015glnasnglnleutyrasngluleuasnleuglyargarggluglutyr202530aspvalleuasplysargargglyargaspproglumetglyglylys354045proglnargarglysasnproglngluglyleutyrasngluleugln505560lysasplysmetalaglualatyrsergluileglymetlysglyglu65707580argargargglylysglyhisaspglyleutyrglnglyleuserthr859095alathrlysaspthrtyraspalaleuhismetglnalaleupropro100105110arg<210>13<211>1131<212>dna<213>人工序列<220><223>核苷酸序列自杀基因是hsv-tkmut2<400>13atggcttcgtacccctgccatcaacacgcgtctgcgttcgaccaggctgcgcgttctcgc60ggccatagcaaccgacgtacggcgttgcgccctcgccggcagcaagaagccacggaagtc120cgcctggagcagaaaatgcccacgctactgcgggtttatatagacggtcctcacgggatg180gggaaaaccaccaccacgcaactgctggtggccctgggttcgcgcgacgatatcgtctac240gtacccgagccgatgacttactggcaggtgctgggggcttccgagacaatcgcgaacatc300tacaccacacaacaccgcctcgaccagggcgagatatcggccggggacgcggcggtggta360atgacaagcgcccagataacaatgggcatgccttatgccgtgaccgacgccgttctggct420cctcatgtcgggggggaggctgggagttcacatgccccgcccccggccctcaccctcatc480ttcgaccgccatcccatcgccgccctcctgtgctacccggccgcgcgataccttatgggc540agcatgaccccccaggccgtgctggcgttcgtggccctcatcccgccgaccttgcccggc600acaaacatcgtgttgggggcccttccggaggacagacacatcgaccgcctggccaaacgc660cagcgccccggcgagcggcttgacctggctatgctggccgcgattcgccgcgtttacggg720ctgcttgccaatacggtgcggtatctgcagggcggcgggtcgtggtgggaggattgggga780cagctttcggggacggccgtgccgccccagggtgccgagccccagagcaacgcgggccca840cgaccccatatcggggacacgttatttaccctgtttcgggcccccgagttgctggccccc900aacggcgacctgtataacgtgtttgcctgggccttggacgtcttggccaaacgcctccgt960cccatgcacgtctttatcctggattacgaccaatcgcccgccggctgccgggacgccctg1020ctgcaacttacctccgggatggtccagacccacgtcaccaccccaggctccataccgacg1080atctgcgacctggcgcgcacgtttgcccgggagatgggggaggctaactga1131<210>14<211>658<212>prt<213>人工序列<220><223>蛋白质序列carcd44v6-nwl<400>14metglualaproalaglnleuleupheleuleuleuleutrpleupro151015aspthrthrglygluilevalleuthrglnserproalathrleuser202530leuserproglygluargalathrleusercysseralaserserser354045ileasntyriletyrtrpleuglnglnlysproglyglnalaproarg505560ileleuiletyrleuthrserasnleualaserglyvalproalaarg65707580pheserglyserglyserglythraspphethrleuthrileserser859095leugluprogluaspphealavaltyrtyrcysleuglntrpserser100105110asnproleuthrpheglyglyglythrlysvalgluilelysarggly115120125glyglyglyserglyglyglyglysergluvalglnleuvalgluser130135140glyglyglyleuvallysproglyglyserleuargleusercysala145150155160alaserglyphethrphesersertyraspmetsertrpvalarggln165170175alaproglylysglyleuglutrpvalserthrileserserglygly180185190sertyrthrtyrtyrleuaspserilelysglyargphethrileser195200205argaspasnalalysasnserleutyrleuglnmetasnserleuarg210215220alagluaspthralavaltyrtyrcysalaargglnglyleuasptyr225230235240trpglyargglythrleuvalthrvalserserglyaspprolysglu245250255alacysprothrglyleutyrthrhisserglyglucyscyslysala260265270cysasnleuglygluglyvalalaglnprocysglyalaasnglnthr275280285valcysgluprocysleuaspservalthrpheseraspvalvalser290295300alathrgluprocyslysprocysthrglucysvalglyleuglnser305310315320metseralaprocysvalglualaaspaspalavalcysargcysala325330335tyrglytyrtyrglnaspgluthrthrglyargcysglualacysarg340345350valcysglualaglyserglyleuvalphesercysglnasplysgln355360365asnthrvalcysgluglucysproaspglythrtyrseraspgluala370375380asnhisvalaspprocysleuprocysthrvalcysgluaspthrglu385390395400argglnleuargglucysthrargtrpalaaspalaglucysgluglu405410415ileproglyargtrpilethrargserthrproprogluglyserasp420425430serthralaproserthrglngluproglualaproprogluglnasp435440445leuilealaserthrvalalaglyvalvalthrthrvalmetglyser450455460serglnprovalvalthrargglythrthraspasnprolysphetrp465470475480valleuvalvalvalglyglyvalleualacystyrserleuleuval485490495thrvalalapheileilephetrpvalargserlysargserargleu500505510leuhisserasptyrmetasnmetthrproargargproglyprothr515520525arglyshistyrglnprotyralaproproargaspphealaalatyr530535540argserargvallyspheserargseralaaspalaproalatyrgln545550555560glnglyglnasnglnleutyrasngluleuasnleuglyargargglu565570575glutyraspvalleuasplysargargglyargaspproglumetgly580585590glylysproargarglysasnproglngluglyleutyrasngluleu595600605glnlysasplysmetalaglualatyrsergluileglymetlysgly610615620gluargargargglylysglyhisaspglyleutyrglnglyleuser625630635640thralathrlysaspthrtyraspalaleuhismetglnalaleupro645650655proarg当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1