栾树体细胞胚胎发生及植株再生的培养方法与流程
本发明涉及植物体细胞胚胎发生及植株再生的培养方法,尤其涉及一种栾树体细胞胚胎发生及植株再生的培养方法,属于栾树体细胞胚及植株再生的培养领域。
背景技术:
栾树是栾树属植物中最主要的代表植物,在园林绿化应用中具有极其重要的作用。栾树属植物在分类学上共有四种,其中三种为中国特有的乡土树种,分别为栾树(koelreuteriapaniculatalaxm.)、台湾栾树(k.formosanahayata.)和复羽叶栾树(k.bipinnatafranch.)。其中在复羽叶栾树中还有一变种为全缘叶栾树(k.integrifoliolamerr.)。此外,还有一类栾树属植物,斐济栾树,分布于斐济群岛。与其他栾树属植物相比,栾树不具备完全的二回羽状复叶,且在小叶边缘有稍粗大、不规则的钝锯齿,近基部的齿常疏离而呈深缺刻状;蒴果圆锥形,顶端渐尖。由于栾树其本身抗逆能力强,比之其他栾树属植物分布范围更广,在中国大江南北都具有很好的适应性,因此常被栽培作庭园观赏树和街道绿化树。栾树作为绿化树种,不仅因其观赏价值高,其对环境修复也有一定的作用,既对so2抗性强,又能吸附烟尘。相关研究也表明,栾树对于重金属离子也有一定的吸附作用。此外,栾树体内富含次级代谢物,如没食子酸衍生物、氰基类化合物和类黄酮化合物等,具有一定的药用价值。栾树的叶片和种子也常常被用作抗菌剂和杀虫剂。
作为中国重要的乡土树种,栾树主要依靠种子繁殖。然而,栾树种子外种皮坚硬,有着强烈的内休眠作用,对其发芽率造成了极大的限制,且在栾树不同种群内,种子发芽率也有明显的差异。虽然众多的研究者们对其发芽率的提高已经进行了相关工作,但对于栾树育种工作的开展仍是远远不足的。此外,栾树花朵小,不易水培,常规的杂交育种工作很难应用于栾树的育种工作。栾树因体内富含次生代谢物,伤口极易褐化,嫁接和扦插成活率很低。
体细胞胚发生途径是植物再生的两种关键途径之一,在植物组织培养中占有重要地位(rosaybcj,belloccm,dornelasmc:species-dependentdivergentresponsestoinvitrosomaticembryoinductioninpassifloraspp.plantcell,tissueandorganculture(pctoc)2015,120(1):69-77.)。与器官发生途径相比,体细胞胚具有个体间遗传背景一致、繁殖数量快和结构完整等诸多优点,不仅可以作为植物快速繁殖的途径,对研究植物形态建成也有重要的意义(quiroz-figueroafr,rojas-herrerar,galaz-avalosrm,loyola-vargasvm:embryoproductionthroughsomaticembryogenesiscanbeusedtostudycelldifferentiationinplants.plantcell,tissueandorganculture2006,86(3):285;zimmermanjl:somaticembryogenesis:amodelforearlydevelopmentinhigherplants.theplantcell1993,5(10):1411-1423.),因此备受广大研究者的喜爱。
体细胞胚发生与诱导器官发生相比具有显著的特点:1)具有两极性。可以直接形成再生植株,在无性繁殖上具有一定的优越性。胚性细胞进行一次不均等的分裂产生两个大小不等的细胞(相当于合子分裂产生的基细胞和顶细胞),然后由较小的细胞继续分裂形成多细胞原胚,较大的细胞进行1-2次分裂成为胚柄,在发育的早期从方向相反的两端分化出胚根和胚芽,在形态上具有明显的极性。不定芽和不定根都表现为单向极性。2)存在生理隔离现象。steward(1958)指出体胚发生的前提条件是细胞与其周围组织之间的生理隔离,体胚原始细胞在开始分裂时常常形成清晰的界限。体胚在发生和发育过程中与周围的组织保持相对独立性,体胚的维管组织与母体植物或外植体的维管组织在结构上没有联系,呈独立的“丫”字形,作为体胚形成的标志。而不定芽或不定根的维管组织总是与母体植株或愈伤组织的维管组织相连接,不易分开。3)遗传相对稳定。通过体胚发生途径再生植株的变异性小于通过器官发生途径形成的再生植株,因为只有那些没有变异或者变异很小的细胞才能形成体胚,因此通过体胚发生途径再生植株的遗传性状相对稳定。4)经历了与合子胚形态发生相似的阶段:体细胞胚胎发生途径经历了球形期胚、心形期胚、鱼雷形胚、子叶期胚等,最后发育成完整的植株。不定芽的产生没有经历这些复杂过程,比较简单。
此外,体细胞发生途径在应用上具有着自己独特的优势。首先,体细胞胚单细胞起源方式可以有效减少嵌合体的存在(rosaybcj,belloccm,dornelasmc:species-dependentdivergentresponsestoinvitrosomaticembryoinductioninpassifloraspp.plantcell,tissueandorganculture(pctoc)2015,120(1):69-77;corredoirae,ballestera,ibarram,vieitezam:inductionofsomaticembryogenesisinexplantsofshootculturesestablishedfromadulteucalyptusglobulusande.saligna×e.maideniitrees.treephysiology2015,35(6):678-690.)。众多研究者认为体胚途径的起源方式为单细胞起源,可以有效的减少遗传转化中嵌合体现象的存在,更有利于基因工程育种工作的开展。此外,体胚发生途径还可应用于人工种子的生产。经由体细胞胚途径产生的子叶胚与合子胚具有类似的形态结构和功能,可用于人工种子的生产(quiroz-figueroafr,rojas-herrerar,galaz-avalosrm,loyola-vargasvm:embryoproductionthroughsomaticembryogenesiscanbeusedtostudycelldifferentiationinplants.plantcell,tissueandorganculture2006,86(3):285.)。生产的人工种子更有利于植株的储存、运输和商品化应用。同时体细胞胚还可作为研究植物合子胚发育的模型。体细胞胚经历了与合子胚形态发生相似的阶段,为研究人员研究合子胚的发育提供了体外模型。
体胚发生途径受多种因素调节,外植体的选择、植物生长调节剂的种类和浓度以及其他环境因子的刺激在体胚发育过程中都发挥了各自的影响作用。
外植体的选择是植物体细胞胚诱导的一个重要因素。前人的研究表明,外植体的基因型、选取部位以及发育程度对体胚的诱导均有影响(rosaybcj,belloccm,dornelasmc:species-dependentdivergentresponsestoinvitrosomaticembryoinductioninpassifloraspp.plantcell,tissueandorganculture(pctoc)2015,120(1):69-77;corredoirae,ballestera,ibarram,vieitezam:inductionofsomaticembryogenesisinexplantsofshootculturesestablishedfromadulteucalyptusglobulusande.saligna×e.maideniitrees.treephysiology2015,35(6):678-690;nordinea,tlemcanicr,elmeskaouia:regenerationofplantsthroughsomaticembryogenesisinthymushyemalislange,apotentialmedicinalandaromaticplant.invitrocellular&developmentalbiology-plant2014,50(1):19-25;saboonin,shekafandeha:somaticembryogenesisandplantregenerationofblackberryusingthethincelllayertechnique.plantcell,tissueandorganculture(pctoc)2017,130(2):313-321.)。同一植物不同个体间诱导体胚的潜能是不一致的,不同的基因型间存在着显著的差异。在花生的研究中,乔利仙等就发现花生的不同品种诱导体胚的成功率是不一致的(乔利仙,隋炯明,谭玲玲,郭宝太,孙世孟,王晶珊:花生体胚诱导及植株再生.农学学报2012(10):9-13.),在白云杉的研究中也取得了类似的结果(parkys,barrettjd,bongajm:applicationofsomaticembryogenesisinhigh-valueclonalforestry:deployment,geneticcontrol,andstabilityofcryopreservedclones.invitrocellular&developmentalbiology-plant1998,34(3):231-239.)。因此在体细胞胚胎的研究中,基因型间的差异应成为研究者们考虑的因素。此外,植物同一个体不同部位和不同生长阶段对体胚的诱导成功率也是有影响的。体胚的发生机理在于植物细胞的全能型,原理上植物的每个细胞都具有潜能,但不同部位的细胞全能型潜能不同。众多研究表明植物种子更具有诱导胚状体的潜能,且未成熟的种子比成熟种子更具有诱导潜能,即生理年龄越小,诱导形成胚状体的潜能越大。
外源激素在植物组织培养中发挥着重要的作用,它是一类含量少但作用大的物质,对组织培养的诱导方向有着重大决策作用(
另一方面,一些研究人员认为植物生调节剂在体细胞胚发生途径中不仅起到生长调节的作用,而且还是重要的压力刺激来源之一(dobrowolskai,busingee,abreuin,moritzt,egertsdotteru:metabolomeandtranscriptomeprofilingrevealnewinsightsintosomaticembryogerminationinnorwayspruce(piceaabies).treephysiology2017,37(12):1752-1766.)。研究者们认为外源的压力刺激是体胚发生的重要推动力。大量的研究表明多种环境因素在体胚发生途径中起到了压力刺激的作用,如植物生长调节剂、无机元素、水势以及氧化还原电势等(
鉴于体细胞胚发生相比诱导器官发生存在诸多的优点,提供一种栾树体细胞胚胎发生以及植株再生的培养方法,将对栾树无性繁殖培养、分子辅助育种以及栾树优良种源的保存等奠定重要的基础。
技术实现要素:
本发明所要解决的技术问题是提供一种栾树体细胞胚胎发生以及植株再生的培养方法,该培养方法体胚发生率高,产生的体胚数量多,提高了栾树的再生植株数量。
为解决上述技术问题,本发明所采取的技术方案是:
本发明公开了一种栾树体细胞胎发生以及植株再生的培养方法,包括以下步骤:
(1)以栾树种子为外植体进行组织培养获得栾树无菌苗植株;(2)将栾树无菌苗的叶片及茎段接种到愈伤诱导培养基中培养获得胚性愈伤组织;(3)将胚性愈伤组织移接至体细胞胚发育培养基中进行体细胞胚的分化培养获得栾树体细胞子叶胚;(4)将栾树体细胞子叶胚在成熟发育培养基中进行子叶胚的成熟发育培养;(5)将发育成熟的子叶胚移至种子发芽培养基进行发芽培养获得完整植株;(6)将完整植株的茎杆的片段接种至生根培养基进行生根培养。
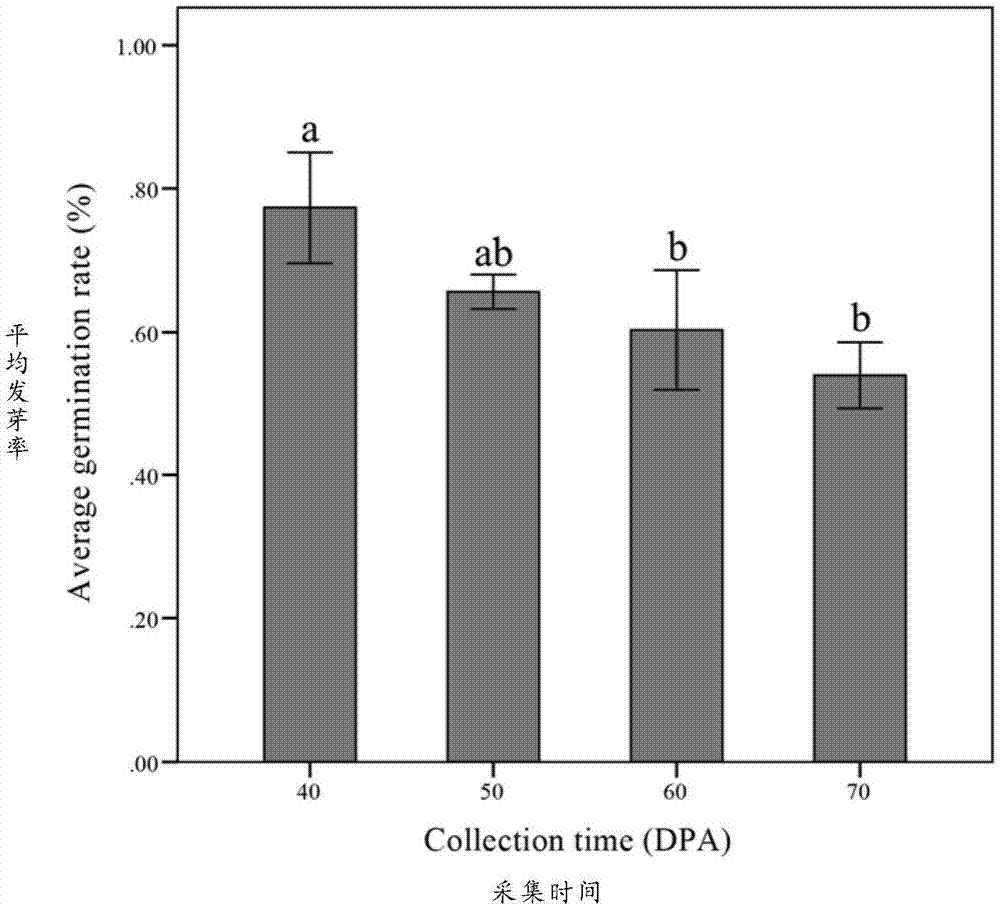
其中,优选的,步骤(1)选择种子胚发育完整后外种皮颜色为深绿色或稍有褐色的栾树种子为外植体材料进行消毒;其中,栾树种子的采集时间优选为每年10月-11月,即栾树开花后40-70天内。
步骤(1)中所述组织培养所采用的培养基为含有1.0mg/liba的1/2dkw培养基,其中,添加5.5g/l的琼脂和20g/l的蔗糖;所述培养的条件为温度为25±2℃,光周期为16/8h,光照强度为40μmolm-2s-1。
步骤(2)中所述愈伤诱导培养基以dkw培养基为基本培养基,其中含0.50-0.75mg/l6-ba、0.50-1.50mg/l2,4-d和0.25-0.50mg/lnaa,优选为含0.50-0.75mg/l6-ba、1.50mg/l2,4-d和0.25mg/lnaa,最优选为含0.50mg/l6-ba、1.50mg/l2,4-d和0.25mg/lnaa。所述愈伤诱导培养基中添加5.5g/l的琼脂和30g/l的蔗糖。
步骤(2)中将栾树无菌苗的叶片及茎段置于愈伤诱导培养基中暗培养,然后移接至相同培养基中进行孵育;优选的,所述暗培养的时间为2-3月;所述孵育的时间为2个月左右,其中每月进行一次继代,保持愈伤的良好状态。
步骤(3)所述体细胞胚发育培养基为含0-1.50mg/l2,4-d或0.10-0.20mg/lnaa的dkw培养基,优选为含0.10mg/lnaa的dkw培养基。步骤(3)每两个月进行一次新的继代培养,继代培养三次后丢弃尚未成功分化的愈伤材料,所获得的栾树体细胞胚为不同时期的栾树体细胞胚,包括球形胚、心形胚、肾形胚及子叶胚。
步骤(4)中所述的成熟发育培养基为含有0.10-0.20mg/lnaa的dkw培养基;其中,所述培养基中添加5.5g/l的琼脂和30g/l的蔗糖。暗培养一周后将体细胞胚移至光下培养,体胚的颜色由白色转变为嫩绿色后,进一步加深为深绿色,随后胚根与胚轴伸长生长,形成具有功能的根和茎。
步骤(5)中所述的发芽培养为将发育成熟的子叶胚移至含1.0mg/liba的1/2dkw的种子发芽培养基中,在光下进行完整植株的发育培养;其中,光周期为16/8h,光照强度为40μmolm-2s-1。所述种子发芽培养基中添加5.5g/l的琼脂和30g/l的蔗糖。
步骤(6)所述生根培养基以1/4dkw培养基为基本培养基,其中含0.5-2.5mg/liba和0-0.3mg/lnaa,优选为含1.0-1.5mg/liba和0-0.3mg/lnaa,更优选为含1.0-1.5mg/liba,最优选为含1.5mg/liba。所述培养基中添加5.5g/l的琼脂和20g/l的蔗糖。所述茎杆为1-2cm长度的片段(至少包含一个茎节)。
本发明所述栾树体细胞胚胎发生及植株再生的培养方法中组织培养的温度为25±2℃,所有培养基的ph为5.8。
体胚发生途径受多种因素调节,包括外植体的选择、植物生长调节剂的种类和浓度以及其他环境因子等对于体细胞胚胎的发生均产生了重要的影响。
本发明对栾树种子发芽率与采集时间的关系分析表明,合理的采集时间是影响消毒接种成功率及发芽率的重要因素,选择栾树种子胚结构发育完整后,外种皮颜色为深绿色或稍有褐色的种子材料最为适宜。
本发明栾树胚性愈伤的诱导以栾树无菌苗的茎段和叶片为诱导材料进行。由种子获得的无菌苗生理年龄小,为栾树胚性愈伤的诱导提供了更高的潜能。胚性愈伤诱导培养基以dkw培养基为基本培养基。前期预实验表明,相比于b5和ms培养基的愈伤诱导状况,dkw培养基更适合培养栾树组织材料。不同浓度激素的处理结果表明,0.50mg/l或0.75mg/l6-ba+1.5mg/l2,4-d+0.25mg/lnaa是栾树胚性愈伤诱导和培养的较适宜配方,有利于栾树愈伤组织的增殖培养,栾树愈伤组织诱导率达70.25%-80.25%,且在愈伤继代培养的过程中愈伤生长状态正常。
将栾树愈伤组织移至体胚发育培养基内进行体细胞胚发育培养过程中,植物激素的种类及浓度显著影响了体细胞胚发育的进程。不同浓度的2,4-d对体细胞胚的发育产生了显著影响,浓度含量过低(0.5mg/l)或过高(1.5mg/l)均抑制了体胚的发育。相比于激素2,4-d,naa具有更好的体细胞胚诱导作用。其中,以含有0.1mg/lnaa的dkw培养基进行体胚发育培养,不仅获得了最高的体胚发生率(54.75%),而且产生的体胚数量多,极大的提高了栾树的再生植株数量。不同时期的体细胞胚在含有naa的培养基中再次孵育获得了进一步的成熟发育。不同浓度的naa在这一过程中并未产生明显的差异影响。
本发明栾树的无性生殖培养以1/4dkw培养基为基本培养基。前期预实验表明,培养基中大量元素的浓度对栾树的生根率产生了显著影响,dkw、1/2dkw培养基并未取得较好的生根效果。不同激素浓度的iba对栾树生根率具有显著影响。当激素iba浓度为1.0-1.5mg/l时,栾树具有较高的生根率;其中,使用1.5mg/liba的生根率最高,为52%。随着iba激素浓度的不断增加,茎段的伸长生长逐渐受到抑制;当激素浓度增加至2.5mg/l时,茎节极度缩短,不利于植株的生长。naa不适合用于栾树的生根培养,更有利于愈伤的产生。同时使用激素naa和iba不仅不能提高生根率,而且延长了植物响应所需的时间。在包含激素naa和iba的培养基中培养的茎段更倾向于产生愈伤组织,进而阻碍了根的产生。
本发明技术方案与现有技术相比,具有以下有益效果:
本发明以栾树未成熟的种子为初始材料,获得栾树无菌苗;然后以无菌苗的茎段和叶片为诱导材料,经历胚性愈伤的诱导、发育、成熟,获得由体胚途径再生的植株。本发明所述栾树体细胞胚培养的方法,体胚发生率高,而且产生的体胚数量多,极大的提高了栾树的再生植株数量,为栾树无性繁殖培养和分子辅助育种等提供了基础。
附图说明
图1为不同采集时间下栾树种子的平均发芽率,误差线标尺为平方差。dpa,开花后天数;字母代表tukey氏多重比较的结果,相同的字母代表在p=0.05的情况下无显著性差异。
图2为栾树体胚组织培养过程观察;a未完全成熟的种子;b由b5培养基诱导的愈伤;c-d由ms培养基诱导的愈伤;e由iem1培养基诱导的愈伤;f由iem2培养基诱导的愈伤;g由iem3培养基诱导的愈伤;h由iem4培养基诱导的愈伤;i多次继代培养后产生的的深色愈伤;jiem3培养基中愈伤产生的气生根(箭头所指处);k光下培养中球形胚的形态;l光下培养中子叶胚的形态;m体细胞胚产生的小植株;n种子产生的小植株;o暗培养中子叶胚的形态;p白色透明的胚性愈伤;q棕色紧实的胚性愈伤;r无性生殖中产生的愈伤(箭头所指处);s种子无菌苗。标尺,a-s1cm。
图3为不同浓度iba下栾树茎段生根率,误差线标尺为平方差。***,代表在p=0.01的情况下tukey氏多重比较的结果。
图4为栾树胚性愈伤和体细胞胚的形态观察;a-b,p表面光滑、有结节状突起的胚性愈伤;c-f由胚性愈伤产生的不同时期的体细胞胚;g-k,r-t融合的子叶胚;l单子叶胚;m-o,q正常的子叶胚。标尺a-o0.2cm;p-t方格大小为1*1cm2。
图5为栾树不同时期体细胞胚组织学观察;a栾树胚性愈伤(黑色箭头所指处)和非胚性愈伤(白色箭头所指处);b胚性愈伤上的结节状突起(箭头所指处);c-d由胚性愈伤分离的球形胚(箭头所指处);e,i肾形胚;f单子叶胚;g,n子叶胚;h心形胚;j鱼雷形胚;k子叶胚子叶横截面;l子叶胚中的维管束(箭头所指处);m子叶胚胚轴横截面。标尺,a-k,m100μm;l50μm。
具体实施方式
下面结合具体实施例来进一步描述本发明,本发明的优点和特点将会随着描述而更为清楚。但是应理解所述实施例仅是范例性的,不对本发明的范围构成任何限制。本领域技术人员应该理解的是,在不偏离本发明的精神和范围下可以对本发明技术方案的细节和形式进行修改或替换,但这些修改或替换均落入本发明的保护范围。
实施例1栾树体细胞胚胎发生及植株再生的培养
1、材料和方法
1.1实验材料
实验材料取自栾树未完全成熟的种子。采集时间为2016年10-11月份,供体栾树定植于北京林业大学(中国,北京市,海淀区)。
1.2实验方法
1.2.1外植体消毒
采集的栾树种子经由流水冲洗1-2h,清洗表面灰尘后,置于75%酒精中浸泡30s,再由无菌蒸馏水冲洗两次,后置于次氯酸钠溶液中振荡清洗5min,再由无菌蒸馏水冲洗5-6次,去除残留的次氯酸钠溶液。消毒完毕的种子经由无菌滤纸吸除多余的水分,在超净工作台内用剪刀镊子去除外种皮,置于含有1.0mg/liba的1/2dkw培养基中放于光下培养,培养基中添加了5.5g/l的琼脂和20g/l的蔗糖。
本发明所用的愈伤诱导、成熟及发芽培养基中均添加5.5g/l的琼脂和30g/l的蔗糖,无性系生根培养基中添加有5.5g/l的琼脂和20g/l的蔗糖。所有的培养基配制完成后由稀酸(hcl)或稀碱(naoh)调节ph至5.8,后经过高温高压(121℃,101kpa,15min)灭菌存储备用。组织培养的材料均置于25±2℃环境下培养,光下培养材料置于16/8h光周期环境,光照强度为40μmolm-2s-1。
1.2.2胚性愈伤的诱导
由种子发芽获得的栾树无菌苗植株是进行胚性愈伤诱导的初始材料。将栾树无菌苗的叶片及茎段剪切并置于愈伤诱导培养基(表1)中暗培养2-3月后,再次移接至相同培养基中进行孵育,每月进行一次继代,保持愈伤的良好状态。
1.2.3体细胞胚的发育及成熟
将孵育3个月左右的栾树愈伤材料移接至体细胞胚发育培养基(表2)中进行体胚的分化培养,每两个月进行一次新的继代培养。继代培养三次后丢弃尚未成功分化的愈伤材料。由发育培养基获得不同时期的栾树体细胞胚(球形胚、心形胚、肾形胚及子叶胚)置于含有naa(0.1-0.2mg/l)的新培养基中进行子叶胚的成熟发育。后将发育成熟的子叶胚(胚根伸长,颜色转变为深绿色)移至含有1.0mg/liba的1/2dkw培养基中放于光下进行完整植株的发育。
1.2.4无性系植株的生根
获得由体细胞胚发育的完整植株后,在超净台内剪除枝叶,将茎杆切除为1-2cm长度的片段(至少包含一个茎节),后将其接种至生根培养基内。生根培养基以1/4dkw培养基为基本培养基,植物生长调节剂选择不同浓度的iba(0.5-2.5mgl)和naa(0.1-0.2mgl)单独使用或配合使用。
1.2.5组织学观察
将获得的栾树愈伤组织和不同阶段的体细胞胚材料经由固定液(甲醛:冰醋酸:乙酸=8:1:1)固定过夜。经由不同梯度的乙醇溶液(50、70、85、95和100%)进行脱水。脱水的材料由融化的石蜡进行包埋,后由石蜡切片机(rm2235,leica)将包埋的蜡块切除为8um的薄片。材料染色采用苯胺蓝染色法,后在荧光万能显微镜(bx61,olympus)下进行观察拍照。
1.2.6数据分析
每个阶段实验均进行了三次生物学重复,每个重复中单个处理材料为9个。茎段及叶片在愈伤诱导培养基上孵育2个月后统计各自的愈伤诱导率。获得的愈伤经由一个月的继代培养后移至发育培养基进行体胚的诱导发育,45天后统计各个培养基内体胚发生率。由于产生的体胚个数难以统计,诱导效果经由分级法显示。产生的体胚再次移接至新的培养基内进行暗培养一周后移至光下培养,体胚的发芽率则在培养2-3周后进行统计。无性生殖的生根率则在茎段培养2个月后进行统计。所有的统计数据经由单因素方差分析检验,由spss统计软件(spss19.0)进行分析,多重比较结果采取t检验(p≤0.5)。
2、实验结果
2.1种子发芽率与采集时间的关系
经由外植体消毒的栾树未成熟种子发芽率与其自身的发育程度存在着密切联系。随着采集时间的进行,种子的消毒难度逐渐增大,且因其种子果实的特性,在超净台内进行外种皮的剥离难度也随着成熟程度而逐渐变大。因此,选择种子胚发育完整后,外种皮颜色为深绿色或稍有褐色进行采集有利于种子的消毒接种。本发明在适宜的时期内共采集了四次种子,每次间隔10天,发芽率的统计结果显示种子的发芽率随着采集时间出现了略微下降(图1),推断有可能是随着栾树种子成熟程度的增加,种子逐渐进入内休眠造成的。此外,不同采集时间的种子最终发育为完整植株的概率也不相同。种子的幼嫩程度越大,其发育成为完整植株的困难程度越大,多数出现子叶扩展后停止发育。
2.2胚性愈伤的诱导及增殖
在四种胚性愈伤诱导培养基中,栾树叶片及茎段的愈伤诱导率并没有产生显著差异,均保持了较高的诱导率(表1)。由四种诱导培养基获得的初始愈伤组织在形态上也无显著性差异(图2e-h)。然而随着继代培养过程的进行,获得的栾树愈伤组织颜色逐渐加深(图2i),但在后续的实验中证明其仍是具有胚性潜能的。此外在愈伤继代培养的过程中,四种培养基出现了显著差异。经由iem3培养基培养的愈伤组织在继代培养2-3次后产生了气生根(图2j),不利于后续体细胞胚的发育培养。由iem4培养基培养的愈伤增殖速度慢,甚至出现死亡。因此选择含有较高浓度2,4-d的iem1和iem2培养基有利于栾树愈伤组织的增殖培养。
表1不同浓度的6-ba、naa和2,4-d对栾树愈伤组织诱导的影响
2.3体细胞胚的发育及成熟
植物激素的种类及浓度显著影响了体细胞胚发育的进程(表2)。不同激素浓度的2,4-d对体细胞胚的发育产生了显著影响,浓度含量过低(0.5mg/l)或过高(1.5mg/l)均抑制了体胚的发育。此外,相比于激素2,4-d,naa显然具有更好的体细胞胚诱导作用。其中,体胚诱导率最高的培养基为含有0.1mg/lnaa的dkw培养基(54.75%)。
不同时期的体细胞胚在含有naa的培养基中再次孵育获得了进一步的成熟发育。不同浓度的naa在这一过程中并未产生明显的差异影响(90.83%,92.81%)。暗培养一周后,将获得的体细胞胚转移至光下培养后则出现颜色的转变,由白色转变为嫩绿色后进一步加深,最终转变为深绿色,并出现了胚根和茎段的伸长。将完全成熟的子叶胚移至含有1.0mg/liba的1/2dkw培养基内进一步培养获得完整植株。
表2不同浓度的2,4-d或naa对栾树体细胞胚发育的影响
注:*,代表每个培养基内产生的体细胞胚个数少于10个.**,代表每个培养基内产生的体细胞胚个数范围未10-30个.***,代表每个培养基内产生的体细胞胚个数大于30个;
字母代表tukey氏多重比较的结果,相同的字母代表在p=0.05的情况下无显著性差异。
2.4无性生殖
不同激素浓度的iba对栾树生根率具有显著影响。当激素iba浓度为1.0-1.5mg/l时,栾树具有较高的生根率(图3)。且随着iba激素浓度的不断增加,茎段的伸长生长逐渐受到抑制。当激素浓度增加至2.5mg/l时,茎节极度缩短,不利于植株的生长。此外,同时使用激素naa和iba并没有取得较好的效果。naa和iba的结合使用不仅不能提高生根率(表3),而且延长了植物响应所需的时间。在包含激素naa和iba的培养基中培养的茎段更倾向于产生愈伤组织(图2r),进而阻碍了根的产生。
表3不同浓度的iba和naa对栾树生根率的影响
2.5组织学观察
在栾树胚性愈伤组织的诱导过程中,常常伴随着非胚性组织的存在(图5a),属于体胚诱导方式中的间接体胚发生方式。与非胚性愈伤组织相比,胚性愈伤组织细胞有着明显的细胞界限,多数呈方形或卵圆形,而非胚性愈伤组织则呈现出杂乱无序的状态(图5a)。胚性愈伤组织表面光滑,有着结节状的突起(图4a-b,p)。这些突起随着发育过程的不断推进,逐渐演变为球形胚或子叶胚(图4c-f)。球形胚的出现伴随着表皮原的形成,并逐渐从愈伤组织上分离出来(图5c-d)。随后球形胚演变为肾形胚(图5e,i),出现两端极性的分化,开始径向伸长。随着发育过程的前进,叶原基分化产生两个子叶的分离,根原基极性生长,出现胚轴的伸长,在此过程中,体胚逐渐演变为心形胚(图5h)和鱼雷形胚(图5j)的形态。待子叶、胚轴、胚根发育成熟后,体细胞胚最终演变为子叶胚(图4m-o,q;图5g,n)。在成熟的子叶胚组织中,本发明也观察到了维管束组织的存在(图5l)。在对栾树胚性愈伤组织和体细胞胚的观察中,本发明同样也发现了单子叶胚(图4l;图5f)和融合子叶胚等畸形胚(图4g-k,r-t)的存在。
综上,本发明以栾树未成熟的种子为初始材料,获得无菌苗后以其茎段和叶片为诱导材料,经历胚性愈伤的诱导、发育、成熟,获得由体胚途径再生的植株,为栾树无性繁殖培养和分子辅助育种提供了基础。
- 还没有人留言评论。精彩留言会获得点赞!