TGL基因在麦蛾防控中的应用的制作方法
本发明属于农业害虫防控技术领域,具体涉及tgl基因在麦蛾防控中的应用。
背景技术:
麦蛾s.cerealellaolivier是一种世界性的粮食害虫,也是我国三大重要储粮害虫之一,麦蛾每年危害导致小麦、稻谷的质量损失率可达65%左右(李光涛等,2008;王殿轩等,2017)。目前对麦蛾的防治方法主要依赖化学防治,但由于麦蛾抗药性的增强及幼虫具有粮粒内为害的特点,化学防治方法难以取得理想效果,因此对麦蛾成虫的防控是降低该害虫为害的最有效方式。
三酰甘油在细胞形成和能量代谢中起重要作用,包括向飞行肌提供代谢燃料,促进卵巢发育,性激素合成和精子发育等(beenakkersetal.1985;canavosoetal.2001;arreseandsoulages2010;wanetal.2015)。三酰甘油在生物体内的利用需要tag酯酶(tagligase,tgl)的参与(yenetal2008),在哺乳体内,hsl和atgl在三酰甘油分解代谢上发挥重要作用,atgl是催化三酰甘油水解第一步的重要脂肪酶(holmetal.1988;zimmermannetal.2004)。在脂肪细胞中,过表达atgl会促进脂肪分解,抑制该基因表达则脂肪分解显著下降,说明atgl对脂肪分解至关重要(黄文钰等2013)。而在昆虫体内,昆虫取食时,脂肪酸以甘油三酯的形式储存在脂肪体的脂滴中,同时为昆虫非进食期提供能量(wolinsetal.2006,arreseandsoμlages2010)。昆虫储存的脂肪酸对昆虫的生长发育必不可少。脂肪酸作为蜡和信息素合成的前体,并作为表皮脂质的成分参与细胞膜的生物合成和磷脂的合成(downerandmatthews1976)。烟草天蛾(manducasexta)三酰甘油脂肪酶(tgl)是唯一经过纯化和鉴定的昆虫三酰甘油水解酶(arreseandwells1994)。
精包是鳞翅目昆虫在交配过程中,雄虫通过射精把附腺以及双射精管的内容物传递给雌虫交配囊的过程中形成的(周祖琳1989)。精包内容物成分复杂包括,糖类、脂类、蛋白质类以及激素类等,其对于精子的活化发育、卵子发育、雌虫再交配等生理生化活动至关重要(何建平等2006)。tgl作为脂代谢通路上的限速酶,对于脂类分解成脂肪酸和甘油至关重要,是昆虫生命活动(如精子和卵子活化发育)必不可缺的物质。但目前对于昆虫tgl基因功能研究较少,具体机制仍不明确。
技术实现要素:
有鉴于此,本发明的目的在于提供tgl基因在麦蛾防控中的应用,抑制tgl基因的表达,会降低三酰甘油的分解速度,从而导致雄虫体内无法合成足够的交配内容物,所以在交配过程中没有传递给雌虫更多的内容物,使得在交配囊内无法形成正常的精包,最终使得成熟卵子不能成功受精。
为了实现上述发明目的,本发明提供以下技术方案:
本发明提供了tgl基因在麦蛾防控中的应用,所述tgl基因的核苷酸序列如seqidno.1所示。
优选的,所述麦蛾防控包括抑制正常精包形成和降低麦蛾产卵量。
有益效果:本发明提供了tgl基因在麦蛾防控中的应用,当抑制tgl基因的表达时,会降低三酰甘油的分解速度,从而导致雄虫体内无法合成足够的交配内容物,所以在交配过程中没有传递给雌虫更多的内容物,使得交配后的雌虫无法获得正常的精包,最终使得成熟卵子不能成功受精。
本发明利用rnai技术抑制麦蛾三酰甘油脂肪酶基因(tgl)的表达,探究tgl基因对麦蛾成虫生殖能力的调控作用。通过向初羽化麦蛾雄成虫体内注射dstgl,再与未处理雌虫交配。结果表明:注射dstgl的雄虫与雌虫成功交配后,交配囊在交配过程中能够被充气,正常鼓起,但其内形成的精包却因雄虫传递给雌虫的交配内容物相对较少,从而导致精包填充不够多,不饱满,进而导致雌虫产卵量下降。
附图说明
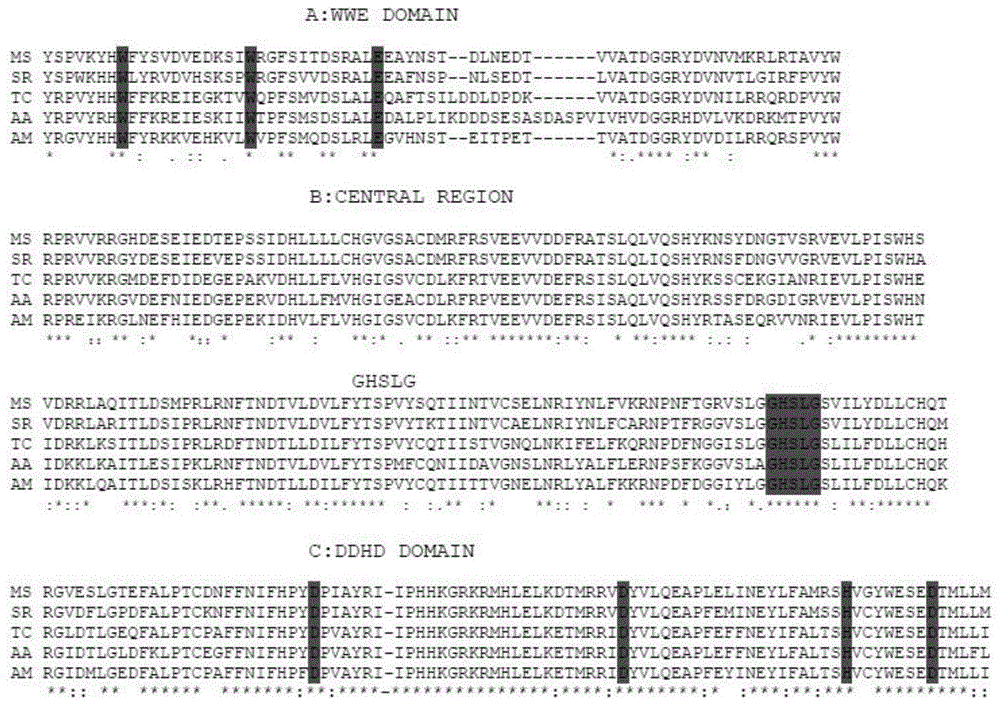
图1为麦蛾tgl氨基酸序列与其他昆虫的多重比对图;
图2为麦蛾tgl基因在雄虫不同发育时期的表达量;
图3为麦蛾tgl基因在不同组织中的表达量;
图4为注射不同体积rnase-freeh2o雄虫6天内存活率;
图5为注射不同溶液雄虫6天内存活率;
图6为注射dstgl7天内沉默效果图;
图7为rna干扰麦蛾雄虫tgl基因后精包形态图;
图8为rna干扰tgl后精包形态统计图;
图9为rna干扰tgl后产卵量。
具体实施方式
本发明提供了tgl基因在麦蛾防控中的应用,所述tgl基因的核苷酸序列如seqidno.1所示。
本发明所述应用中,所述tgl基因的开放阅读框为1968bp,编码656个氨基酸(seqidno.2),预测分子量为72.966kda,等电点为5.54,脂类鉴定的可能性为77.98%,总的疏水性平均系数为-0.419,表明该编码蛋白为亲水性蛋白。本发明所述麦蛾tgl基因的编码蛋白存在脂类的三个保守区域,分别是a:n端58-128wwe保守结构域(pfam:pf02825);b:含有脂肪酶共识序列ghslg的中心区域;c:c末端ddhd保守结构域(pfam:pf02862);同时ser367、asp457、his590是该蛋白的活性位点。
本发明所述tgl基因的获得,优选以麦蛾总rna反转录后的第一链cdna为模板,以tgl1和tgl2引物进行pcr扩增得到麦蛾tgl基因片段tgl1和tgl2,然后将得到的进行拼接,即得所述tgl基因。本发明所述tgl1和tgl2引物序列分别如表1所示:
表1tgl基因扩增引物
本发明利用所述引物进行pcr扩增时,所述pcr扩增的体系以25μl计,包括:cdna1μl,引物f0.5μl,引物r0.5μl,primesrar酶mix
10μl和ddh2o13μl。本发明所述pcr扩增的程序优选为:95℃预变形5min;95℃变性10s,60℃退火10s,72℃延伸30s,35个循环。
本发明对所述基因片段tgl1和tgl2的拼接方法并没有特殊限定,利用本领域的常规技术手段即可。
本发明所述tgl基因在脂肪体中表达量最高,其次在精巢中表达量也是相较于其他组织较高(p<0.05)。tgl作为脂类分解代谢的重要酶,在脂肪体中高表达用于分解三酰甘油,但是在精巢组织中也高表达,揭示了tgl可能对于精巢组织的发育或者精子的发育有重要作用。且tgl基因在幼虫和成虫各个阶段表达量均比蛹期较高,tgl基因在幼虫期高表达用于分解三酰甘油参与机体能量代谢,从而促进生长发育;在成虫羽化后48h内表达量均高于幼虫期,羽化72h时表达量开始下降。tgl基因在成虫期也高表达说明,除了参与机体能量代谢外,可能也与生殖能力相关。
在本发明中,抑制所述tgl基因的表达,可以抑制正常精包形成(正常交配完的交配囊正常鼓起,内部精包整体呈珠光色饱满的球形,并且挤破精包有乳白色内容物流出)和降低麦蛾产卵量,从而起到麦蛾防控的效果。本发明对抑制所述tgl基因表达的方法并没有特殊限定,利用本领域的常规方法即可。
下面结合实施例对本发明提供的tgl基因在麦蛾防控中的应用进行详细的说明,但是不能把它们理解为对本发明保护范围的限定。
实施例1
tgl基因克隆及时空表达模式
1、材料与方法
1.1供试昆虫
实验所用麦蛾为农户储粮抓取室内同样条件饲养多代的麦蛾,于圆底透明玻璃瓶内饲养(高10cm,半径4cm),在瓶内装约2cm厚干净麦粒,置于温度28±1℃,相对湿度75±5%,光周期l:d=14:10人工气候箱饲养。挑选含有蛹的麦粒于指形管中单独饲养,逐日检查,取初羽化处女蛾,区分雌雄后备用。
1.2仪器设备及主要试剂
1.2.1仪器设备
neofuge13r台式高速冷冻离心机(上海力申科学仪器有限公司);凝胶成像仪geldocxrplus(bio-rad,usa);水平式核酸电泳仪powerstationae-8450;核酸蛋白测定仪biophotometer(eppendorf,germany);实时荧光定量pcr仪cfx96(bio-rad,usa)。
1.2.2主要试剂
trisol(北京全式金生物科技有限公司);反转录试剂盒(上海翊圣生物有限公司);胶回收试剂盒(axygen);质粒小提试剂盒(axygen);sybrgreensupermix(takara);primestarmaxdnapolymerase(takara)。
1.3实验方法
1.3.1tgl基因克隆
1.3.1.1rna提取
1、将麦蛾放入无酶的1.5ml离心管中,先用液氮处理,快速研磨组织,期间不断加入液氮,直至研磨成粉末状。
2、先加入500μltrizol到离心管中,用研磨棒混匀,再加500μltrizol清洗研磨棒,涡旋振荡5s,室温静置5~15min。
3、加入200μl氯仿,剧烈振荡15s,充分混匀,室温5min;4℃,12000rpm离心15min,这时会分为3层,(无色rna,白色蛋白层,有色的dna),吸取上清至新的离心管中。
4、加入等体积的异丙醇(500μl),上下颠倒,充分混匀(可放在-20℃),4℃,12000rpm,离心10min。
5、弃上清,加入1000μl,-20℃预冷的75%的乙醇,上下颠倒,清洗沉淀,12000rpm,4℃,离心5min。(此步骤可重复一次)
6、弃上清,会有少许残余,离心1min,用枪头吸取残液尽量吸净,至室温干燥,2~5min(不宜过长,一般为3min,在通风橱中进行)。
7、加入适量的depc水溶解沉淀(一般50μl,若沉淀较大则根据量选择80~100μl)。
8、检测rna浓度,跑胶检测rna质量,-80℃保存。
1.3.1.2反转录
采用翊圣生物科技有限公司的定量专用反转录试剂盒hifairii1ststrandcdnasynthesissupermixforqpcr(gdnadigesterplus)合成cdna。操作步骤如下:
1)残留基因组dna去除(10μl),体系如表2所示:
表2残留基因组dna去除体系
混匀后42℃,孵育2min;
2)逆转录反应体系(20μl),体系如表3所示:
表3逆转录体系
3)反应程序
25℃5min,42℃30min,85℃5min,-20℃保存。
1.3.1.3tgl基因扩增
基于麦蛾无参转录组数据,筛选注释为tgl基因的序列,并放到ncbi上比对序列,选取相似度最高的序列,运用primer5.0在线软件设计如seqidno3~6的引物扩增麦蛾tgl基因片段。
pcr反应体系(25μl),如表4所示:
表4pcr扩增体系
反应条件:
95℃预变形5min;95℃变性10s,60℃退火10s,72℃延伸30s,35个循环。
1.3.1.4pcr产物回收
pcr产物经低熔点琼脂糖凝胶电泳后进行切胶,利用axygen胶回收试剂盒说明书进行产物纯化,回收完成后将产物送至擎科生物公司进行测序。将序列结果进行blast比对分析、序列拼接获得完整的tgl基因的开放阅读框,并进行生物信息学分析。
利用primestar酶成功获得tgl1和tgl2基因片段,其中tgl1为1104bp,tgl2为888bp。对两个片段进行拼接,得到麦蛾tgl基因的开放阅读框为1968bp,编码656个氨基酸,预测分子量为72.966kda,等电点为5.54,脂类鉴定的可能性为77.98%,总的疏水性平均系数为-0.419,表明该蛋白为亲水性蛋白。将推测的氨基酸序列和其他昆虫进行多重比对,发现麦蛾tgl也存在脂类的三个保守区域分别是a:n端58-128wwe保守结构域(pfam:pf02825);b:含有脂肪酶共识序列ghslg的中心区域;c:c末端ddhd保守结构域(pfam:pf02862)。同时ser367、asp457、his590是该蛋白的活性位点。同其他昆虫相似,氨基酸序列相似度也较高。具体比对结果如图1所示,其中ms:烟草天蛾(acr61720.1;manducasexta);sr:麦蛾(sitootrogacerealella);tc:赤拟谷盗(xp_008200243.1;triboliumcastaneum);aa:埃及伊蚊(eat38149.1;aedesaegypti);am:意大利蜜蜂(xp_003251782.2;apismellifera)。a表示n末端wwe结构域的比对(pfam:pf02825);b表示含有脂肪酶共有序列ghslg的中心区域的比对;c表示与ddhd结构域(pfam:pf02862)同源的c末端结构域的比对。为了简洁起见,在比对中省略了部分序列。完全一致的用星号表示;完全保守的替换用两个点表示。部分保守的替换用单点表示,序列间隔用破折号表示。
1.3.2tgl基因时空表达模式
1.3.2.1取样
选取幼虫、蛹、初羽化0h、6h、12h、18h、24h、48h、72h的雄虫用于时间表达模式分析;选取雄虫附腺、精巢、头、脂肪体,雌虫卵巢用于组织表达模式分析。每个样品取三个重复,按照上节提取rna并反转录为cdna。
1.3.2.2引物特异性检测和表达模式分析
利用primerpremier5.0软件设计的qtgl基因引物(表5),将反转录得到的cdna稀释25倍作为模板,对三酰甘油脂肪酶基因tgl表达量进行qrt-pcr分析,每个样品重复4次,以gapdh作为内参基因。
表5麦蛾tgl和gapdh基因定量表达引物
荧光定量pcr体系(10μl),如表6所示:
表6荧光定量pcr体系
两步法pcr扩增标准程序:
步骤1:95℃预变性5min;步骤2:扩增反应95℃5s,60℃30s,40个循环;步骤3:融解曲线55℃30s,每个循环增加0.5℃,一直增加到95℃。
利用qrt-pcr技术检测麦蛾tgl基因在雄虫不同发育时期(图2)和不同组织中的表达量(图3),tgl在脂肪体中表达量最高,其次在精巢中表达量也是相较于其他组织较高(p<0.05)。tgl作为脂类分解代谢的重要酶,在脂肪体中高表达用于分解三酰甘油,但是在精巢组织中也高表达,揭示了tgl可能对于精巢组织的发育或者精子的发育有重要作用。且tgl基因在幼虫和成虫各个阶段表达量均比蛹期较高,tgl基因在幼虫期高表达用于分解三酰甘油参与机体能量代谢,从而促进生长发育;在成虫羽化后48h内表达量均高于幼虫期,羽化72h时表达量开始下降。tgl基因在成虫期也高表达说明,除了参与机体能量代谢外,可能也与生殖能力相关。
注:在图2和图3中,图中数值为平均值±标准误,不同小写字母表示经one-wayavova分析差异显著(p<0.05)。
实施例2
tgl基因的功能研究
1材料与方法
1.1供试昆虫
同实施例1,取初羽化处女雄蛾,备用。
1.2仪器设备及主要试剂
1.2.1仪器设备
szm系列体视显微镜(舜宇仪器有限公司);neofuge13r台式高速冷冻离心机(上海力申科学仪器有限公司);凝胶成像仪geldocxrplus(bio-rad,usa);水平式核酸电泳仪powerstationae-8450;核酸蛋白测定仪biophotometer(eppendorf,germany);实时荧光定量pcr仪cfx96(bio-rad,usa);nanoliter2010显微注射仪(wpi,usa)。
1.2.2主要试剂
胶回收试剂盒(axygen);质粒小提试剂盒(axygen);感受态细胞:dh5αchemicallycompetentcell(上海翊圣生物有限公司);载体:pmd-19t(takara);sybrgreensupermix(takara);primestarmaxdnapolymerase(takara);t7riomaxtmexpressrnaisystem(promega)
1.3实验方法
1.3.1dstgl产物扩增,如表7所示:
表7合成dsrna引物
注:加粗序列表示t7启动子序列
以tgl基因pcr产物为模板,用dstgl1和dstgl2的引物分别扩增两段双链rna。
pcr扩增产物体系(25μl),如表8所示:
表8pcr扩增产物体系
扩增程序:
95℃预变性5min;95℃变性10s,60℃退火10s,72℃延伸10s,35个循环;16℃保存1min。
利用上述引物扩增的到的片段大小分别为dsegfp462bp,dstgl1614bp;dstgl2504bp。
1.3.2胶回收
具体步骤同上。
1.3.3连接转化、鉴定与测序
产物:dstglpcr胶回收产物
载体:pmd19t(takara),-20℃保存
感受态:dh5αchemicallycompetentcell(上海翊圣生物有限公司),-80℃保存。
连接体系(10μl)如表9所示:
表9连接体系
反应条件:16℃,30min
反应结束后,立即将离心管置于冰上。
(2)转化
1.加连接产物于100μl感受态细胞中(在感受态细胞刚解冻时立即加入),轻弹混匀,冰浴30min。
2.42℃水浴热激45s,立即置于冰上1min。
3.加入500μllb液体培养基(无amp),200rpm,37℃培养1h
4.取100μl菌液均匀地涂在平板上,在37℃培养箱中过夜培养(为得到较多克隆,1500g离心1min,弃掉部分上清,保留100-150μl,轻弹悬浮菌体,取全部菌液涂板)。
(3)鉴定和测序
1.挑选lb培养基上的菌落,接种于含有amp的lb液体培养基中,37℃,200r/min振荡培养6-8h至液体混浊。以菌液为模板,通用引物m13为引物进行pcr,产物经凝胶电泳鉴定后,挑选符合预测大小的菌液测序。
保种和摇菌
将测序正确的菌株取100μl加入含有抗生素amp的新鲜lb培养基6ml重新摇菌,获取新鲜菌液,将1ml菌液和35%的甘油按照1:1比例混合,保存于-80℃。剩余的菌液提取质粒。(每100ml培养基加入100μl氨苄)。
1.3.4质粒的提取
试剂盒:easypureplasmidminiprepkit,具体方法参照说明书。
1.3.5dsrna模板的制备
以带有靶标基因片段的质粒为模板,用primestar酶扩增,检测有无条带以及条带大小,扩增体系(25μl),如表10所示:
表10dsrna扩增体系
扩增程序:95℃预变性5min;95℃变性10s,60℃退火10s,72℃延伸10s,35个循环;16℃保存1min。
以质粒为模板进行pcr扩增,各自扩增10管25μl体系。
1.3.6利用酚氯仿抽提法提取pcr产物的dna
1.3.7合成dsrna
以pcr产物抽提产物为模板,t7dsrna体外合成试剂盒(promega),70%乙醇,95%乙醇,3m(ph5.2)乙酸钠。
rnasesolution需要先稀释200倍;ezymemix需要在冰上融化;除了72℃要用水浴外,其余都在pcr仪上进行。
1、第一步:分别合成两个单链rna(ssrna),反应前若2xbuffer有沉淀,先在37℃溶解后使用。反应体系如表11所示:
表11反应体系
反应条件:42℃,30min
2、将两个合成的单链rna(ssrna)混合,72℃,水浴10min,并在水中自然冷却至室温。
3、消化反应
rnasesolution稀释200倍,且不保存,不重复利用。消化反应体系如表12所示:
表12消化反应体系
反应条件:37℃,30min
4、加0.1个体积的3m乙酸钠(4.4μl,2.5个体积的95%乙醇1100μl,混匀后冰浴5min,然后在4℃,13000g离心10min。
5、弃掉上清,用500μl,75%乙醇洗涤沉淀,4℃,13000g离心5min,并在室温下干燥10-15min,加2-5倍体积的depc水溶解沉淀,-80℃保存备用。
1.3.8麦蛾成虫的注射方法
利用显微注射仪nanoliter2010对麦蛾成虫进行注射。注射部位参考陆艳的方法:注射部位在腹部第5~6节节间膜。注射前先把成虫置于冰中2-3min,冻晕,然后把虫放在平板上,腹部朝上,用软镊子轻轻按住腹部两侧,用注射仪进行注射即可,注射完成后用毛笔将虫扫到玻璃管中,放回培养箱中。
设置总注射量为250nl和500nl,两种方式对麦蛾进行注射,每个注射体积每次注射30头虫,连续注射3次,统计雄虫存活率。在确定注射量后,同时注射dstgl和dsegfp,每种溶液每次注射30头虫,连续注射3次,统计雄虫存活率。
通过对初羽化成虫进行注射存活率统计,分别对雄性麦蛾进行50nl注射量注射10次,以及50nl注射量注射5次这两个不同的注射剂量,结果如图4所示,其中a(50nl注射量注射10次)注射剂量成虫存活率明显小于b(50nl注射量注射5次)注射剂量。同时对注射dstgl和注射dsegfp的成虫进行6天内的存活率统计,结果如图5所示,两组之间在6天内无明显差异(p>0.05),在5天内的存活率均高于50%,第6天略有降低。
由上述可知,最终注射浓度2000-2500ng/μl,注射剂量为50nl注射量注射5~6次,最终注射量为250~300ng为宜。
1.3.9定量检测tgl基因沉默效果
取初羽化的雄虫进行注射,每头虫注射1-1.5ngdsrna,注射完dsrna后,雄虫放置在温度28±1℃,相对湿度75±5%,光周期l:d=14:10人工气候箱中。分别取注射后1天、2天、3天、5天、7天的虫提rna,反转录cdna,qrt-pcr检测tgl基因的表达量。
通过对注射dstgl和注射dsegfp的成虫进行沉默效率统计,结果如图6所示,两段双链rna在3天内的沉默效果和dsegfp组相比均差异显著(p<0.01),说明注射双链tgl均能显著降低tgl基因的表达。但是在注射后的第5天tgl基因的表达开始上升,第7天比对照组略有上调,说明注射双链tgl在第5天开始被降解。
1.3.10雄虫精包统计
注射完dsrna后,雄虫放置在温度28±1℃,相对湿度75±5%,光周期l:d=14:10人工气候箱中两天,第三天取出雄虫与羽化12小时的处女雌虫配对,观察雌雄虫交配情况,待雄虫与雌虫交配结束后立即在pbs中取出精包,于解剖镜下观察和统计精包形态,每组统计20个精包,3个生物学重复。用lightool’s软件进行拍照。
通过解剖注射双链rna的雄虫交配的雌虫,结果如图7所示,交配后雌虫体内的精包(b和c),明显比对照组小(a),注射dstgl的精包b和c精包和注射dsegfp的a精包相比内容物填充不足,所以导致内部精包较小。对精包进行统计,结果如图8所示,注射dstgl后,会显著增加不正常精包的形成(p=0.001,df=4,f=0.072),注射dsegfp组的不正常精包率为0.1±0.02887,注射dstgl组的不正常精包率为0.5167±0.0441。两组之间正常精包率之间差异极显著(p=0.001,df=4,f=2.571),注射dsegfp组的不正常精包率为0.6667±0.01667,注射dstgl组的不正常精包率为0.2167±0.0441。同时无论是注射dsegfp还是dstgl均有一定概率会在交配后无精包,其中dsegfp组无精包率为0.2333±0.016667,dstgl组无精包率为0.2667±0.016667,两组之间无明显差异(p=0.23,df=4,f=0)。
1.3.11产卵量统计
取注射dsrna两天后的雄虫与未处理羽化12h的雌虫进行配对,观察交配情况,待成功交配后,置产卵纸于管中,5天后统计产卵量,以注射dsegfp的雄虫为对照组,每组统计20对,设置3个重复。
结果如图9所示,注射dstgl的雄虫在与未处理雌虫交配后,产卵量和注射dsegfp的对照组相比显著下降(p=0.001,df=120,f=2.343),注射dsegfp的雄虫在与雌虫交配后,5天内的产卵总量为52.77±5.38,注射dstgl的雄虫在与雌虫交配后,5天内的产卵总量为28.82±4.56。
通过统计麦蛾产卵量发现注射dstgl基因后麦蛾的产卵量显著下降,将图9与图8精包形成率相比,注射dstgl后,雌虫形成正常精包的概率也显著下降。因此tgl基因可能与麦蛾交配过程中形成精包有关,抑制tgl基因表达后,麦蛾形成正常精包的概率下降了,进而导致麦蛾产卵量下降。
以上所述仅是本发明的优选实施方式,应当指出,对于本技术领域的普通技术人员来说,在不脱离本发明原理的前提下,还可以做出若干改进和润饰,这些改进和润饰也应视为本发明的保护范围。
序列表
<110>华中农业大学
<120>tgl基因在麦蛾防控中的应用
<160>16
<170>siposequencelisting1.0
<210>1
<211>1971
<212>dna
<213>人工序列(artificialsequence)
<400>1
atggacggtaacgctgacgctaagagagacagtatgatggaacaggagaaggatgtagat60
ccccggaggtcgatcacggtgaattttgacagcagtgtggaggaggaatcaaatgaaggt120
gtcactgtgatgacagagaaccggtcctccctggagtactcgccagtgaagcaccactgg180
ttctaccgtgtggatgtggacagcaagtccacgtggcgagggttctccgtggtggactcc240
agggcgcttgaggaggcgtttaacagcccgaacctgagcgaagacaccctggtggcgaca300
gatggtggaaggtacgatgtgaacgtgacgggccggcttcggttccccgtctactggacc360
gacaagccgaccaatgttagacggtgcagttggttttacaagggtaccaccgacgccaga420
tatgtgccgtatactgaagcggttgctgagaagttggaggaggagtaccgccatggaatg480
acgacgggcgactggcaccgccgactagtgttaccgaacaacgaaatggtagtcatgcat540
gggccggctgtgatggtgcatttccttcagggcagcgccactgatgccttcagcgcgccg600
ccgcaatcaacaatgcgtcccagagtagtccgccgaggttacgacgagtctgagatcgag660
gaggtggaaccctccagcattgaccacctgctgttgctctgccatggagtagggtccgct720
tgtgatatgaggttccgatccgttgaggaggtggtcgacgacttcagagctaccagtctg780
cagctgattcagtcgcattatcggaattccttcgatagtggagttgttggaagagtggag840
gttctgccaatatcttggcacgcgaccctgcactctggcgagacgggcgtggaccggcgg900
ctggcccgcatcacgctggacagcatcccccgcctgaggaacttcaccaacgacactgtc960
ttggacgtgctgttctacaccagcccggtctacacgaagacgataatcaacacagtatgc1020
gccgagctgaatcgcatctacaatctgttctgcgcgcgcaatcctaccttccgtggtggg1080
gtgtcattaggggggcactcgctcggcagcgtcatattgtatgatctgttgtgtcatcag1140
atggggccggccccaatggcgccttcaacatcagacaagcgttacgtaagcggcgcggtg1200
ggtaccggactacccgttgtggactacccaccgctgcacttcacgcctactgcgctctac1260
gcgctgggcagtcctatagcaatattcgaatgcatccgcggcgtggacttcctcgggccg1320
gacttcgcgctgcccacctgcaagaatttcttcaacatcttccacccgtacgaccccatc1380
gcttacaggatagaaccgctgatcaaccctgaactgcgagacgtgaagccgttcctgata1440
ccccatcacaaagggaggaagaggatgcatctcgagctcaaagacacgatggctcgcgtg1500
ggtgcggacatcaaacagaaactcatcgagagcattaaaagcacttggtccagcatgtgg1560
aagacacagccgccgcccactgacagacagctcgaaaaggttgtggaagaagagattgaa1620
aaagagcaattggaagatgataaggaggaggtgcaacccgatgacaagactccgcccgaa1680
ctcctaggccgcttgaacagcggtcgtcgcgtggactacgtattacaagaggcgcccttc1740
gagatgatcaacgagtatttattcgcgatgagcagccatgtttgttactgggaatccgaa1800
gacacgatgctgctgatgctccgagagatctacgactcaatgggagtgcaacccgactgc1860
tcactcccgcagcacagcctcaccgtgcagcggacgcggctgcccaccccggaagaaact1920
ctcctactgagcacgactgaacatccgtcaaccagccgcggacccacgtga1971
<210>2
<211>656
<212>prt
<213>人工序列(artificialsequence)
<400>2
metaspglyasnalaaspalalysargaspsermetmetgluglnglu
151015
lysaspvalaspproargargserilethrvalasnpheaspserser
202530
valgluglugluserasngluglyvalthrvalmetthrgluasnarg
354045
serserleuglutyrserprovallyshishistrpphetyrargval
505560
aspvalaspserlysserthrtrpargglypheservalvalaspser
65707580
argalaleugluglualapheasnserproasnleusergluaspthr
859095
leuvalalathraspglyglyargtyraspvalasnvalthrglyarg
100105110
leuargpheprovaltyrtrpthrasplysprothrasnvalargarg
115120125
cyssertrpphetyrlysglythrthraspalaargtyrvalprotyr
130135140
thrglualavalalaglulysleuglugluglutyrarghisglymet
145150155160
thrthrglyasptrphisargargleuvalleuproasnasnglumet
165170175
valvalmethisglyproalavalmetvalhispheleuglnglyser
180185190
alathraspalapheseralaproproglnserthrmetargproarg
195200205
valvalargargglytyraspglusergluileglugluvalglupro
210215220
serserileasphisleuleuleuleucyshisglyvalglyserala
225230235240
cysaspmetargpheargservalglugluvalvalaspaspphearg
245250255
alathrserleuglnleuileglnserhistyrargasnserpheasp
260265270
serglyvalvalglyargvalgluvalleuproilesertrphisala
275280285
thrleuhisserglygluthrglyvalaspargargleualaargile
290295300
thrleuaspserileproargleuargasnphethrasnaspthrval
305310315320
leuaspvalleuphetyrthrserprovaltyrthrlysthrileile
325330335
asnthrvalcysalagluleuasnargiletyrasnleuphecysala
340345350
argasnprothrpheargglyglyvalserleuglyglyhisserleu
355360365
glyservalileleutyraspleuleucyshisglnmetglyproala
370375380
prometalaproserthrserasplysargtyrvalserglyalaval
385390395400
glythrglyleuprovalvalasptyrproproleuhisphethrpro
405410415
thralaleutyralaleuglyserproilealailepheglucysile
420425430
argglyvalasppheleuglyproaspphealaleuprothrcyslys
435440445
asnphepheasnilephehisprotyraspproilealatyrargile
450455460
gluproleuileasnprogluleuargaspvallyspropheleuile
465470475480
prohishislysglyarglysargmethisleugluleulysaspthr
485490495
metalaargvalglyalaaspilelysglnlysleuilegluserile
500505510
lysserthrtrpsersermettrplysthrglnproproprothrasp
515520525
argglnleuglulysvalvalgluglugluileglulysgluglnleu
530535540
gluaspasplysglugluvalglnproaspasplysthrproproglu
545550555560
leuleuglyargleuasnserglyargargvalasptyrvalleugln
565570575
glualapropheglumetileasnglutyrleuphealametserser
580585590
hisvalcystyrtrpglusergluaspthrmetleuleumetleuarg
595600605
gluiletyraspsermetglyvalglnproaspcysserleuprogln
610615620
hisserleuthrvalglnargthrargleuprothrproglugluthr
625630635640
leuleuleuserthrthrgluhisproserthrserargglyprothr
645650655
<210>3
<211>22
<212>dna
<213>人工序列(artificialsequence)
<400>3
atcatggacggtaacgctgacg22
<210>4
<211>20
<212>dna
<213>人工序列(artificialsequence)
<400>4
gagtgcccccctaatgacac20
<210>5
<211>21
<212>dna
<213>人工序列(artificialsequence)
<400>5
tcattaggggggcactcgctc21
<210>6
<211>22
<212>dna
<213>人工序列(artificialsequence)
<400>6
taattcacgtgggtccgcggct22
<210>7
<211>17
<212>dna
<213>人工序列(artificialsequence)
<400>7
gacggtgcagttggttt17
<210>8
<211>18
<212>dna
<213>人工序列(artificialsequence)
<400>8
catttcgttgttcggtaa18
<210>9
<211>24
<212>dna
<213>人工序列(artificialsequence)
<400>9
ccatcaagcaaaaggtaagggagg24
<210>10
<211>22
<212>dna
<213>人工序列(artificialsequence)
<400>10
ttgatgagtgcgtgtcaccgat22
<210>11
<211>44
<212>dna
<213>人工序列(artificialsequence)
<400>11
ggatcctaatacgactcactatagggaatcgcatctacaatctg44
<210>12
<211>42
<212>dna
<213>人工序列(artificialsequence)
<400>12
ggatcctaatacgactcactataggagctgtctgtcagtggg42
<210>13
<211>42
<212>dna
<213>人工序列(artificialsequence)
<400>13
ggatcctaatacgactcactatagggtgaagccgttcctgat42
<210>14
<211>42
<212>dna
<213>人工序列(artificialsequence)
<400>14
ggatcctaatacgactcactataggtgtgctgcgggagtgag42
<210>15
<211>42
<212>dna
<213>人工序列(artificialsequence)
<400>15
ggatcctaatacgactcactataggcaagttcagcgtgtccg42
<210>16
<211>43
<212>dna
<213>人工序列(artificialsequence)
<400>16
ggatcctaatacgactcactataggcttgatgccgttcttctg43
- 还没有人留言评论。精彩留言会获得点赞!