构建视网膜色素变性疾病模型的方法、应用以及繁育方法与流程
本发明涉及基因编辑技术领域,具体而言,涉及一种构建视网膜色素变性疾病模型的方法、应用以及繁育方法。
背景技术:
视网膜色素变性(retinitispigmentosa,rp)是一种导致视力丧失的进行性、遗传性致盲眼底病,通常为视网膜光感受器异常所致,临床要表现为慢性进行性视野缺失,夜盲,色素性视网膜病变和视网膜电图异常,最终可导致失明。rp在中国人群的发病率可达1/3500,由于我国庞大的人口基数,rp患者可达三十万之众,给家庭和社会带来了沉重的负担。
目前针对rp尚无有效的治疗手段。rp的诊断和治疗面临许多困难,这主要归因于其在临床表型和遗传上具有高度的异质性,并且针对其病理机制系统研究不足。典型的rp患者最早由于视杆细胞功能缺陷而出现夜盲和视野狭窄,逐步发展为管状视野,直至失明;眼底检查可见视网膜色素沉着。在病理学方面,典型的rp主要影响视杆细胞,造成视杆细胞死亡并继发视锥细胞死亡,主要表现为光感受器受损、变性,视网膜外核层逐渐变薄直至消失,视网膜外网层及其他相关细胞层出现相应病理改变。并且导致rp的具体分子机制尚不清楚,这为rp的临床诊断和治疗带来极大阻碍,因此有必要对rp的详细致病机制进行深入研究。
而目前,rp疾病模型种类较少,需要更多的rp疾病模型和构建方法。
鉴于此,特提出本发明。
技术实现要素:
本发明的目的在于提供一种构建视网膜色素变性疾病模型的方法、应用以及繁育方法。本发明提供的方法可以构建出一种新的具有视网膜色素变性疾病特征的模型。该疾病模型可用于视网膜色素变性疾病的发病过程及机制等相关研究,也可以用于视网膜色素变性疾病的相关药物筛选,具有广阔的应用前景。
本发明是这样实现的:
一方面,本发明提供一种构建视网膜色素变性疾病模型的方法,其包括:使目标动物的视网膜视杆细胞中的同源染色体上的mettl14基因具有相同的修饰,且所述修饰导致mettl14基因产生如下效果中的至少一种:
(a):mettl14基因不表达;
(b):mettl14基因的表达被抑制;
(c):mettl14基因表达的蛋白不具有正常的生物活性功能。
mettl14蛋白(methyltransferaselike14)作为甲基转移酶复合体的一部分参与rna的m6a甲基化修饰过程,其广泛分布于各种类型的组织细胞中。m6a甲基化的形成过程主要是由甲基转移酶复合体(mettl3,mettl14和wtap等组成)、去甲基酶(fto和alkbh5)以及阅读蛋白(ythdf1/2/3,ythdc1)来进行动态调控,与基因表达调控密切相关。其次,m6a可能参与了mrna转录、选择性剪切、出核转运、翻译以及降解等生物学过程,而导致rna功能紊乱,进而影响一系列的动物生命活动。目前针对mettl14蛋白功能的研究逐渐增多,包括在肿瘤发生、机体发育等方面的影响,但其详细作用机制以及其在视网膜中的生物学功能还不甚明了,限制了其开发应用。
本发明的发明人创造性地发现,通过对目标动物的视网膜视杆细胞中的同源染色体上的mettl14基因进行修饰使mettl14基因不表达;mettl14基因的表达被抑制;或mettl14基因表达的蛋白不具有正常的生物活性功能;均可以使得目标动物表现出视网膜色素变性疾病的特征,例如视杆细胞死亡,主要表现为光感受器受损、变性,视网膜外核层逐渐变薄直至消失,视网膜外网层及其他相关细胞层出现相应病理改变等。因此,具有上述修饰的目标动物可以作为视网膜色素变性疾病模型;本发明为视网膜色素变性疾病研究提供了新的模型基础,可以帮助阐明视网膜色素变性疾病的发病过程及机制,并为该疾病的治疗或预防提供新的靶标;该视网膜色素变性疾病模型也可以用于筛选用于预防或治疗视网膜色素变性疾病的药物。
同源染色体上的mettl14基因具有相同的修饰是指,位于同源染色体上的两个不同位置的mettl14基因具有一样的修饰,即纯合修饰。具有如此修饰的目标动物,必然表现出视网膜色素变性疾病特征。
可选的,在本发明的一些实施方案中,所述修饰为突变、缺失和插入中的一种或几种的组合。
上述的修饰可以是突变、缺失和插入中的一种或几种的组合;本领域技术人员基于本发明公开的内容可以选择合适的核苷酸修饰方式。
当采用突变修饰时,可以是一个或多个核苷酸的突变,通过突变修饰可以是的相应的蛋白位点发生氨基酸改变,通过突变修饰至少可以实现mettl14基因表达的蛋白不具有正常的生物活性功能。
当采用缺失修饰时,可以是一个或多个核苷酸的缺失,例如可以优选为外显子上的一个或多个核苷酸的缺失;通过缺失修饰至少可以实现mettl14基因不表达、mettl14基因的表达被抑制以及mettl14基因表达的蛋白不具有正常的生物活性功能等效果。
当采用插入修饰时,可以是在mettl14基因中插入一个或多个核苷酸;例如在外显子中插入一个或多个核苷酸使其发生移码突变,导致表达的蛋白氨基酸序列和结构发生改变,进而可以实现mettl14基因表达的蛋白不具有正常的生物活性功能。
因此,无论选用何种修饰,只要使其mettl14基因产生如上效果(1)-(3)中的任意一种,使目标动物表现出视网膜色素变性疾病特征,即属于本发明的保护范围。
可选的,在本发明的一些实施方案中,所述修饰位于mettl14基因外显子序列上或位于mettl14基因上游的驱动mettl14基因表达的启动子序列上。
对于修饰的位置,本领域技术人员容易想到将其控制在mettl14基因的外显子上,但是可以将修饰控制在驱动mettl14基因表达的启动子序列上,例如删除原始启动子的全部或部分使其驱动表达的功能丧失即相当于发生缺失修饰,或者使用其他的特异启动子序列代替原始的启动子即相当于发生突变修饰,通过对启动子的修饰也可使得mettl14基因不表达或其表达被抑制;进而使得目标动物表现出视网膜色素变性疾病特征。
可选的,在本发明的一些实施方案中,所述修饰通过如下技术中的一种或几种的联用实现:基因敲除技术和基因编辑技术中的一种或几种的组合。
可选的,在本发明的一些实施方案中,所述基因敲除技术为cre-loxp基因敲除技术。
可选的,在本发明的一些实施方案中,所述基因编辑技术选自crispr/cas9技术、zfn技术和talen技术中的任意一种或几种的组合。
本领域技术人员可以采用本领域现有存在的或本申请公开后的多种基因修饰技术例如基因敲除技术(例如cre-loxp基因敲除技术)、基因编辑技术(例如crispr/cas9技术、人工核酸酶介导的锌指核酸酶技术(zinc-fingernucleases,zfn)、转录激活因子样效应物核酸酶技术(transceriptionactivator-likeeffectornucleases,talen)等)等技术中的一种或几种的组合来实现上述修饰,这对本领域技术人员来说是容易实现的。因此,无论采用何种基因修饰技术实现上述修饰以使目标动物表现出视网膜色素变性疾病特征,均属于本发明的保护范围。
可选的,在本发明的一些实施方案中,所述目标动物为非人哺乳动物。
mettl14基因存在于所有的哺乳动物中,在不同的哺乳动物中均具有相似的功能,因此,本发明所述的目标动物可以是非人哺乳动物中的任意动物,例如可以是小鼠、大鼠、马、牛、羊、兔、狗、猪、猴、猿和猩猩中的任意一种。本领域技术人员可以根据需要或兴趣,选择合适的非人哺乳动物作为目标动物,以构建视网膜色素变性疾病模型。因此,无论选择何种非人哺乳动物作为目标动物,均属于本发明的保护范围。
可选的,在本发明的一些实施方案中,所述非人哺乳动物选自小鼠、大鼠、马、牛、羊、兔、狗、猪、猴、猿和猩猩中的任意一种。
可选的,在本发明的一些实施方案中,所述目标动物为小鼠,所述修饰为缺失,所述缺失为mettl14基因中的一个或多个外显子序列被删除。
小鼠mettl14基因(mgi:1919014),该基因位于小鼠3号染色体123368295-123385990bp,全长17.7kb,其cdna全长2741bp,包含11个外显子。
当所述目标动物为小鼠时,可以通过删除其mettl14基因中的一个或多个外显子序列来构建小鼠视网膜色素变性疾病模型。
可选的,在本发明的一些实施方案中,所述缺失为mettl14基因中第2外显子-第10外显子序列被删除。
本发明提供的视网膜色素变性疾病模型具有典型的视网膜色素变性疾病特征,具有多方面的用途,例如用于视网膜色素变性疾病研究和筛选用于预防或治疗视网膜色素变性疾病药物等。
另一方面,本发明提供由如上任一项所述的视网膜色素变性疾病模型的方法所得到的视网膜色素变性疾病模型在视网膜色素变性疾病研究中的应用,所用应用不以疾病的诊断或治疗为目的。
另一方面,本发明提供由如上所述的构建视网膜色素变性疾病模型的方法所得到的视网膜色素变性疾病模型在筛选用于预防或治疗视网膜色素变性疾病药物中的应用。
可选的,在本发明的一些实施方案中,所述应用包括:向所述视网膜色素变性疾病模型施用候选药物,如果所述视网膜色素变性疾病模型在施用所述候选药物前后发生如下变化中的至少一种,则指示所述候选药物可以作为预防或治疗视网膜色素变性性疾病的药物:
(1)施用所述候选药物后,相较于施用所述候选药物前,所述视网膜色素变性疾病模型的视力提高;
(2)施用所述候选药物后,相较于施用所述候选药物前,所述视网膜色素变性疾病模型的视网膜外核层变厚;
(3)施用所述候选药物后,相较于施用所述候选药物前,所述视网膜色素变性疾病模型的视网膜外节增长。
另一方面,本发明提供一种视网膜色素变性疾病模型的繁育方法,其包括:以如上任一项所述的构建视网膜色素变性疾病模型的方法所得到的视网膜色素变性疾病模型为亲本进行杂交。
在通过上述构建方法得到初代视网膜色素变性疾病模型后,为了获取更多数量后代视网膜色素变性疾病模型,本领域技术人员容易想到通过将上述的初代视网膜色素变性疾病模型进行相互交配的育种方法以获得更多数量的后代视网膜色素变性疾病模型,这种繁育视网膜色素变性疾病模型的方法也是属于本发明的保护范围。
另一方面,本发明提供由如上任一项所述的构建视网膜色素变性疾病模型的方法所得到的视网膜色素变性疾病模型的鉴定方法,其包括:检测待鉴定动物的视网膜视杆细胞中的同源染色体上的mettl14基因是否具有相同的所述修饰。
通过上述鉴定方法可以分别出待鉴定动物是否属于本发明提供的构建视网膜色素变性疾病模型,如果存在所述修饰,则该待鉴定动物为本发明提供的视网膜色素变性疾病模型,如果不存在上述修饰则该待鉴定动物不属于本发明提供的视网膜色素变性疾病模型。
对于具体如何鉴定是否存在上述修饰,本领域技术人员可以采用本领域常规的技术例如测序技术、pcr技术以及蛋白印迹技术等实现,这对本领域技术人员来说是容易实现的,无论采用何种技术进行鉴定,均属于本发明的保护范围。
附图说明
为了更清楚地说明本发明实施例的技术方案,下面将对实施例中所需要使用的附图作简单地介绍,应当理解,以下附图仅示出了本发明的某些实施例,因此不应被看作是对范围的限定,对于本领域普通技术人员来讲,在不付出创造性劳动的前提下,还可以根据这些附图获得其他相关的附图。
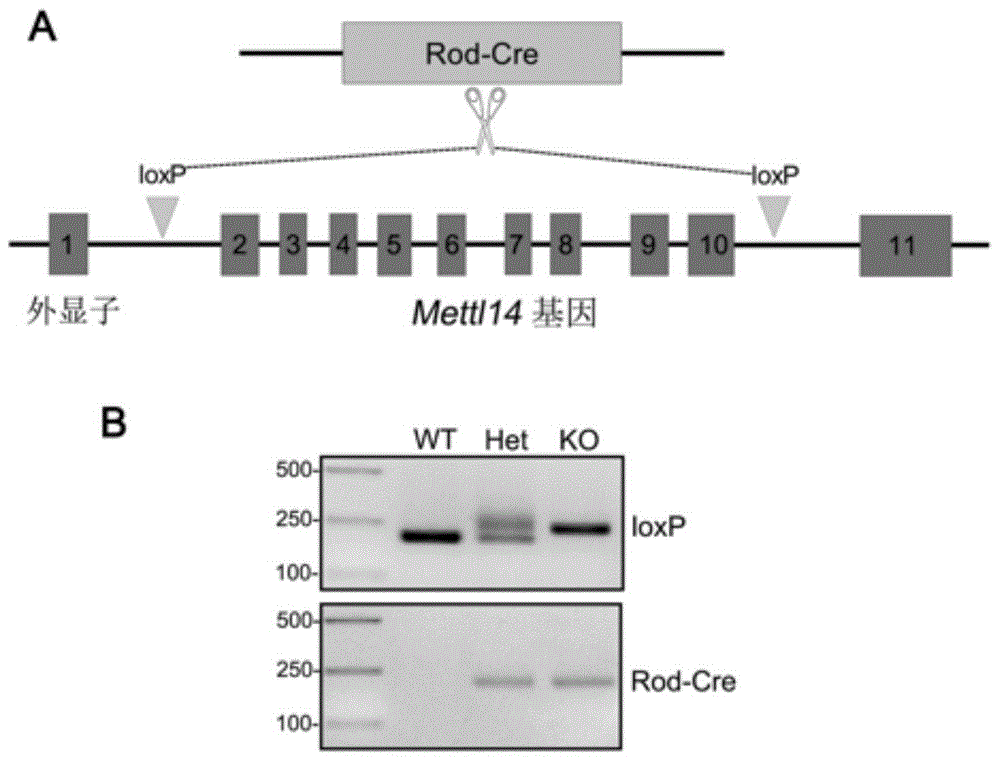
图1:视网膜视杆细胞特异敲除mettl14基因小鼠(ko)的构建和鉴定;
图中:
a:mettl14基因敲除小鼠的构建路线;
b:mettl14基因敲除小鼠的基因型鉴定结果。
图2:mettl14基因敲除效率检测;
a:westernblot实验分析mettl14敲除小鼠视网膜中基因敲除效率;
b:视网膜视杆细胞特异敲除mettl14基因小鼠ihc染色结果,证明mettl14在敲除小鼠视网膜视杆细胞中不再表达。
图3:暗适应视网膜电图(electroretinogram,erg)检测结果;
图中:
a:不同光强下mettl14基因敲除小鼠的暗适应视网膜电图a波统计;
b:不同光强下mettl14基因敲除小鼠的暗适应视网膜电图b波统计。
图4:视网膜视杆细胞特异敲除mettl14基因小鼠视网膜切片免疫组化染色结果;小鼠年龄:3.5个月和5个月;
图中:
a:视网膜视杆细胞特异敲除mettl14基因小鼠视网膜石蜡切片h&e染色结果,外核层及内核层均变薄。
b:对不同部位的mettl14敲除鼠视网膜外核层厚度统计。
图5:视网膜视杆细胞特异敲除mettl14基因小鼠ihc染色结果,表明在mettl14敲除鼠视网膜外节变短退化;小鼠年龄:5个月。
图6:视网膜视杆细胞特异敲除mettl14基因小鼠ihc染色结果,表明在mettl14敲除鼠视网膜出现损伤,引发炎症反应;小鼠年龄:5个月。
具体实施方式
为使本发明实施例的目的、技术方案和优点更加清楚,下面将对本发明实施例中的技术方案进行清楚、完整地描述。实施例中未注明具体条件者,按照常规条件或制造商建议的条件进行。所用试剂或仪器未注明生产厂商者,均为可以通过市售购买获得的常规产品。
以下结合实施例对本发明的特征和性能作进一步的详细描述。
实施例1
1视网膜色素变性疾病模型的构建
本实施例以小鼠为目标动物,对本发明实施例提供的视网膜色素变性疾病模型的构建方法进行说明,mettl14基因敲除的路线如图1中a所示,修饰方式为:删除小鼠mettl14基因的第2-10外显子;通过crispr/cas9技术和cre-loxp基因敲除技术的联用实现修饰。具体操作如下:
1)mettl14基因条件性敲除小鼠(mettl14em1flv,该小鼠mettl14基因的第2外显子上游和第10外显子下游插入了同向的loxp位点)由美国耶鲁大学richardflavell教授赠送;
2)将步骤1)得到的mettl14基因条件性敲除杂合子小鼠相互交配繁育,得到mettl14基因条件性敲除纯合子小鼠;
3)将步骤2)得到的mettl14基因条件性敲除纯合子小鼠与转rod-cre基因小鼠(b6.cg-pde6b+tg(rho-icre)1ck/boc,购自美国杰克森实验室,mgi:4417915)交配,rod驱动cre基因在视网膜视杆细胞中特异性表达,得到视网膜视杆细胞条件性敲除mettl14基因小鼠,通过鉴定后(方法见后文),将鉴定结果正确的小鼠作为视网膜色素变性疾病模型。
2视网膜色素变性疾病模型的鉴定方法如下:
1)剪待鉴定小鼠尾梢少许组织样本,置于干净的1.5ml离心管中;
2)在离心管中加入100μl裂解液(40mmnaoh,0.2mmedta溶液),并在金属浴100℃加热1h;
3)取出离心管,冷却至室温后,加入100μl的中和液(40mmtris-hcl,ph5.5),10000g离心2min后,取上清用于小鼠基因型鉴定。
4)pcr扩增:按照如下体系配置pcr反应体系
2×taqmix10μl;
尾巴组织裂解液2μl;
引物1(mettl14-loxp-forward或rod-cre-forward),1μl(浓度:10mm);
引物2(mettl14-loxp-reverse或rod-cre-reverse),1μl(浓度:10mm);
ddh2o6μl。
引物序列如下:
mettl14-loxp-forward序列:5’-agcggccacttacagttgac-3’;
mettl14-loxp-reverse序列:5’-cctgtcgccaatggtgaatg-3’;
rod-cre-forward序列:5’-tcagtgcctggagttgcgctgtgg-3’;
rod-cre-reverse序列:5’-cttaaaggccagggcctgcttggc-3’。
扩增程序:
pcr反应体系配制完后,在pcr仪上于95℃预加热5分钟使模板dna充分变性,然后进入扩增循环。在每一个循环中,先于95℃保持30秒钟使模板变性,然后将温度降到复性温度58℃,保持30秒,使引物与模板充分退火;在72℃保持30秒,使引物在模板上延伸,合成dna,完成一个循环。重复这样的循环25次,使扩增的dna片段大量累积。最后,在72℃保持5分钟,使产物延伸完整,4℃保存。
5)凝胶电泳
称取1g琼脂糖放于100mltaebuffer中,微波炉中融化后制成1%的琼脂糖胶。取10ulpcr产物于加样孔中,120v恒压琼脂糖电泳15min.用凝胶成像系统成像。
图1中b中显示了对野生型对照小鼠、杂合子小鼠和纯合子小鼠的对应的条带大小和分布,wt表示野生型对照,扩增loxp序列的条带大小为221bp,无扩增rod-cre基因的条带;
het表示杂合子小鼠,有三个条带,有两个扩增loxp序列的条带,分布是221bp和253bp,有一个扩增rod-cre基因的条带大小约为232bp;
ko表示纯合子小鼠,有两个条带,扩增loxp序列的条带大小为253bp,扩增rod-cre基因的条带大小约为232bp,具有如此条带分布和大小的结果,说明该待鉴定小鼠的同源染色体上的mettl14基因具有相同的修饰,即为视网膜色素变性疾病模型。
实施例2
1免疫印迹(westernblot)实验分析rod-cre敲除小鼠视网膜中基因敲除效率。
方法:
1)分别获取对照和敲除小鼠视网膜,充分研磨后加入200μl蛋白裂解液ripa。
2)超声破碎细胞后,在冰上裂解20min。
3)4℃,16000g离心10min后,取上清转移至另一干净离心管,加入50μl的蛋白上样液,混匀后95℃加热5min。
4)待样本冷却后,分别取20μl,160v电压进行聚丙烯酰胺凝胶电泳(sds-page)以分离蛋白。
5)sds-page结束后,根据需要,裁剪适当大小的硝酸纤维素膜,按顺序铺上滤纸、胶、硝酸纤维素膜及滤纸,并赶去气泡,转膜槽放入冰水浴中,采用恒流0.28a的电流进行转膜,转膜2h。
6)转膜完毕后,纯水冲洗硝酸纤维素膜一遍,晾干并标记。然后用8%的脱脂牛奶封闭2h。
7)封闭完成后,加入一定量的按一定比例(按照抗体使用说明书)稀释于封闭液的一抗,4℃孵育过夜。
8)回收一抗,1×tbst缓冲液洗膜4次,每次10min,根据一抗来源,选择合适二抗,用1×tbst稀释辣根过氧化氢酶(hrp)标记的二抗,室温于摇床上孵育2h。
9)二抗孵育结束后,用1×tbst洗膜3次,每次10min,用thermo的elc发光试剂盒检测蛋白,所用仪器为bio-rad的化学发光凝胶成像系统。
2免疫组化(ihc)实验分析rod-cre敲除小鼠视网膜中基因敲除效率。
视网膜冰冻切片免疫染色:取3月龄的实施例1构建得到的视网膜视杆细胞特异敲除mettl14基因小鼠断颈处死后,快速取眼球,并放入4%的pfa中,冰上固定15min后,在角膜上剪口,然后继续冰上固定。2h后,pbs缓冲液冲洗3遍,然后将眼球置于30%蔗糖溶液中脱水2h,然后解剖镜下剪去角膜及晶体,oct包埋并迅速置于-80℃冰箱冷冻。大约10min后,取出oct包埋的眼球,置于冰冻切片机-25℃平衡约30min后即可切片。切片厚度为12μm。
切片完成后,选取质量较高的片子于37℃烘箱放置30min,然后免疫组化笔在有视网膜组织的地方画圈,pbs洗三遍以去除oct,然后5%的nds(含有0.25%triton)封闭通透2h,孵育一抗,4℃过夜。第二天,pbs清洗三次后,孵育相应的荧光二抗,然后再用pbs清洗三次,封片,观察。
结果见图2,在小鼠3月龄时,通过视网膜冰冻组织切片染色mettl14抗体后发现,相较于野生型小鼠,敲除型小鼠视网膜外核层以及视杆细胞内节mettl14表达消失,表明其在视网膜视杆细胞中被特异性敲除。
实施例3
对5月龄的mettl14基因敲除小鼠进行erg视力检测:
1)暗适应动物应整夜暗适应,环境应做到绝对没有光线;
2)次日麻醉:称重,腹腔内注射;宜深度麻醉;
3)动物固定及散瞳:麻醉完成后,在暗红光照明下将小鼠用胶带固定在动物试验平台的前方:需保证小鼠正"趴",即相对于闪光刺激器的刺激口,双眼高度一致,充分暴露,滴加散瞳剂。
4)电极安装:将视网膜电图仪(espionvisualelectrophysiologysystem,diagnosysllc,lit-tleton,ma,usa)预热后,在电极涂上导电膏,夹住小鼠尾巴,插入放大器“地”接口;双头的针电极插入后颈皮(大约在两耳中间),同时接两个通道的“负”接口;金环电极夹在动物实验平台的电极支架上,仔细调整其角度,轻微的接触角膜的中心顶端。一通道正极接右眼,二通道正极接左眼。通过针管对双眼滴生理盐水,改善金环电极及角膜的接触效果。保证两个金环电极以相同的角度、方式接触两眼角膜中心正端的相同位置。
5)记录示波信号确认无误后,关闭暗红光。可以先尝试记录一下暗适应光强为0.003cd/s·m2的erg检测,确认一下信号的质量:如果双眼的振幅出现了与预期不同的较大差异,建议再次检查金环电极的安装位置。然后依次记录暗适应光强为0.03/0.3/3.0/20.0cd/s·m2的信号,记录后系统将自动打开背景光。
结果发现在5个月时,相较于wt(野生)小鼠,ko(视网膜视杆细胞敲除)小鼠的a波和b波在暗适应条件下均明显降低,表明mettl14在视杆细胞敲除后导致视力受损(图3)。
实施例4
视网膜石蜡切片h&e染色:
对3.5和5月龄小鼠的视网膜进行石蜡切片、苏木精-伊红染色法(h&e染色方法)染色,具体操作如下:
1)快速取小鼠眼球组织,并置于固定液中固定24h;
2)石蜡包埋,切片,厚度为4μm;
3)切片常规用二甲苯脱蜡,经多级乙醇至水洗:二甲苯(i)5min→二甲苯(ⅱ)5min→100%乙醇2min→95%的乙醇1min→80%乙醇1min→75%乙醇1min→蒸馏水洗2min;
4)苏木素染色5分钟,自来水冲洗;
5)盐酸乙醇分化30秒;
6)自来水浸泡15分钟;
7)置伊红液2分钟。
8)常规脱水,透明,封片:95%乙醇(i)1min→95%乙醇(ⅱ)1min→100%乙醇(i)1min→100%乙醇(ⅱ)1min→二甲苯石碳酸(3:1)1min→二甲苯(i)1min→二甲苯(ⅱ)1min→中性树脂封固。
9)显微镜下拍照。
结果发现在3.5个月时,相较于wt(野生)小鼠,ko(视网膜视杆细胞敲除)小鼠的视网膜外核层已经开始变薄,而5月龄时外核层厚度显著变薄,表明感光细胞死亡(图4)。
实施例5
视网膜冰冻切片免疫染色:取5月龄的实施例1构建得到的视网膜视杆细胞特异敲除mettl14基因小鼠断颈处死后,快速取眼球,并放入4%的pfa中,冰上固定15min后,在角膜上剪口,然后继续冰上固定。2h后,pbs缓冲液冲洗3遍,然后将眼球置于30%蔗糖溶液中脱水2h,然后解剖镜下剪去角膜及晶体,oct包埋并迅速置于-80℃冰箱冷冻。大约10min后,取出oct包埋的眼球,置于冰冻切片机-25℃平衡约30min后即可切片。切片厚度为12μm。
切片完成后,选取质量较高的片子于37℃烘箱放置30min,然后免疫组化笔在有视网膜组织的地方画圈,pbs洗三遍以去除oct,然后5%的nds(含有0.25%triton)封闭通透2h,孵育一抗,4℃过夜。第二天,pbs清洗三次后,孵育相应的荧光二抗,然后再用pbs清洗三次,封片,观察。
结果见图5,在小鼠3月龄时,通过视网膜冰冻组织切片染色外节抗体rhodopsin后发现,相较于野生型(wt)小鼠,敲除型(ko)小鼠视网膜外节明显缩短,出现明显退化表征。
实施例6
视网膜冰冻切片免疫染色:取5月龄的实施例1构建得到的视网膜视杆细胞特异敲除mettl14基因小鼠断颈处死后,快速取眼球,并放入4%的pfa中,冰上固定15min后,在角膜上剪口,然后继续冰上固定。2h后,pbs缓冲液冲洗3遍,然后将眼球置于30%蔗糖溶液中脱水2h,然后解剖镜下剪去角膜及晶体,oct包埋并迅速置于-80℃冰箱冷冻。大约10min后,取出oct包埋的眼球,置于冰冻切片机-25℃平衡约30min后即可切片。切片厚度为12μm。
切片完成后,选取质量较高的片子于37℃烘箱放置30min,然后免疫组化笔在有视网膜组织的地方画圈,pbs洗三遍以去除oct,然后5%的nds(含有0.25%triton)封闭通透2h,孵育一抗,4℃过夜。第二天,pbs清洗三次后,孵育相应的荧光二抗,然后再用pbs清洗三次,封片,观察。
结果见图6,在小鼠5月龄时,通过视网膜冰冻组织切片染色神经胶质细胞标记物gfap后发现,相较于野生型小鼠,敲除型小鼠视网膜出现明显胶质细胞增生,炎症反应增强,表明视网膜损伤。
综上,可以看出,本发明实施例以小鼠为例,通过crispr/cas9技术和cre-loxp敲除技术的联用在其视网膜视杆细胞中特异性敲除mettl14基因,使小鼠表现出视力受损,视细胞外节变短退化以及视细胞丢失等视网膜色素变性疾病典型表征。由此充分说明,在视网膜视杆细胞条件性敲除mettl14基因,可以使目标动物表现出视网膜色素变性疾病特征。视网膜视杆细胞条件性敲除mettl14基因的动物,可以作为视网膜色素变性疾病模型。该疾病模型可以用于视网膜色素变性疾病研究等领域中,为该疾病的研究例如发病过程、机制以及相关药物的筛选提供一种新的疾病模型基础。
以上所述仅为本发明的优选实施例而已,并不用于限制本发明,对于本领域的技术人员来说,本发明可以有各种更改和变化。凡在本发明的精神和原则之内,所作的任何修改、等同替换、改进等,均应包含在本发明的保护范围之内。
- 还没有人留言评论。精彩留言会获得点赞!