一种诱导斑点叉尾鮰性别分化的方法与流程
1.本发明涉及斑点叉尾鮰培育技术领域,具体涉及一种诱导斑点叉尾鮰性别分化的方法。
背景技术:
2.斑点叉尾鮰(ictalurus punctatus),属鲶形目(siluriformes)鮰科(ictaluridae),原产于北美,是美国最主要的淡水养殖品种,养殖产量占美国鱼类养殖产量的60%以上。自2013年起,我国的斑点叉尾鮰养殖产量超越美国,成为世界最大的斑点叉尾鮰养殖和消费国家。相较于斑点叉尾鮰雄鱼,雌鱼生长较为缓慢,且较大的卵巢组织一定程度上降低了其可食用比例,因此雄鱼在流通市场上更受欢迎。
3.性别二态性是自然界中的常见现象。在鱼类中,大多数种类由于存在生长、性成熟年龄、体型、体色等性别二态性现象,使得其中的一种性别具有更大的经济价值,雌雄个体经济价值的不同在一定程度上制约了其产业的发展。因此,越来越多的育种工作者开始探索性别控制育种,培育具有更大经济价值的单性品种。
技术实现要素:
4.为了克服现有技术中存在的不足,本发明提供一种诱导斑点叉尾鮰性别分化的方法。在斑点叉尾鮰全雄鱼的选育过程中,获得稳定遗传的xy伪雌鱼是最为核心的环节。本发明以培育斑点叉尾鮰全雄品种为对象,着力开展温度诱导其xy型雄鱼雌性化研究。在伪雌鱼性成熟后,通过与正常雄鱼杂交或采用雌核发育策略获得yy型超雄鱼,超雄鱼再与正常雌鱼杂交即可培育出全雄鱼。
5.为实现上述目的,本发明提供一种诱导斑点叉尾鮰性别分化的方法,其包括以下步骤,
6.选择卵黄囊苗在斑点叉尾鮰鱼卵流水孵化装置中孵化出苗,培育至30日龄,培育期间控制水体温度为36
±
0.5℃,上调foxl2基因的表达实现性别逆转;
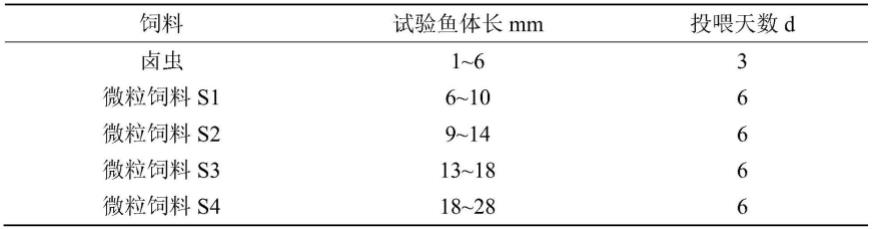
7.待卵黄囊褪去,仔鱼上游觅食后,先投喂卤虫开口饵料,然后根据体长投喂对应的微粒饲料;
8.30日龄试验鱼养殖至室外水泥池中,饲料更换为商品饲料,每日上、下午各投喂一次,日投喂量为鱼体重的3%,继续培育即可。
9.优选的,xy雌性斑点叉尾鮰dmrt1基因未被完全抑制。
10.优选的,所述微粒饲料,其成分为粗蛋白≥50%,粗脂肪≥8%,粗纤维≤3%,粗灰分≤16.5%,钙≤5%,总磷≥1%,水分≤12%,赖氨酸≥2%。
11.优选的,所述商品饲料,其成分为粗蛋白≥30%,水分≤12%,粗纤维≤10%,粗灰分≤13%,粗脂肪≥3%,总磷≥1%,赖氨酸≥1.5%。
12.本发明提供一种斑点叉尾鮰伪雌鱼。
13.本发明提供foxl2基因在诱导斑点叉尾鮰性别分化的应用,其为:上调foxl2基因
的表达。
14.优选的,抑制dmrt1基因的表达。
15.优选的,foxl2基因编码区的反应引物序列如seq id no:1、seq id no:2所示;dmrt1基因编码区的反应引物序列如seq id no:3、seq id no:4所示。
16.本发明提供用于鉴别斑点叉尾鮰性别的引物,其特征在于:引物序列如seq id no:1-seq id no:4所示。
17.本发明的有益效果如下:
18.本发明通过高温胁迫获得了斑点叉尾鮰雄性性别分化的控制方法。前期试验发现在斑点叉尾鮰中通过提高温度能够使其性腺向雌性化方向分化发育,这为斑点叉尾鮰的性控制研究提供了新的思路。斑点叉尾鮰的卵巢分化起始于受精后的第19d,而精巢的分化晚于卵巢,一般在受精后的第90~102d开始。在斑点叉尾鮰的卵黄囊苗时期提高养殖水体温度,以诱导斑点叉尾鮰向雌性化方向分化。比较不同处理组生长数据,并利用组织学切片技术观察其性腺分化与发育情况;同时,采用qrt-pcr方法分别检测雄性性别标志基因dmrt1基因和雌性性别标志基因foxl2基因在xx和xy伪雌鱼卵巢中的表达水平。本试验分别探究温度对斑点叉尾鮰生长及性腺分化的影响及不同温度对性腺分化影响的潜在机制,为温度影响斑点叉尾鮰分化机制奠定基础,同时为日后建立环境友好型的斑点叉尾鮰性别控制育种模式提供前期探索。
附图说明
19.图1为60日龄斑点叉尾鮰卵巢组织图像,其中,图a、c、g是t-30、t-33、t-36组中xx雌鱼的卵巢组织切片图,图e、i是t-33、t-36组中xy伪雌鱼的卵巢组织切片图;图b、d、h是t-30、t-33、t-36组中xx雌鱼的卵巢组织解剖图,图f、j是t-33、t-36组xy伪雌鱼的卵巢组织解剖图;
20.图2为斑点叉尾鮰性别分化基因表达图,其中图a为斑点叉尾鮰雄性性别分化基因dmrt1表达图,图b为斑点叉尾鮰雌性性别决定基因foxl2表达图。
具体实施方式
21.为更好的说明本发明的目的、技术方案和优点,下面将结合具体实施例对本技术作进一步说明。
22.实施例1:
23.试验所选用的斑点叉尾鮰亲鱼来自国家级江苏斑点叉尾鮰遗传育种中心。健康亲鱼交配产卵后,选取活力旺盛的受精卵,在斑点叉尾鮰鱼卵流水孵化装置中孵化出苗。使用加热棒进行水体加热,设置(30
±
0.5)℃(t-30,ck)、(33
±
0.5)℃(t-33)和(36
±
0.5)℃(t-36)3个温度组,初始水温为28℃,每隔12h升温1℃,至水温到达设定温度后持续保持温度恒定。每个试验组设置3个平行,每个平行放置800尾鱼苗(规格:52尾/g),试验从卵黄囊苗的培育开始,进行为期30d的温度诱导试验。待卵黄囊褪去,仔鱼上游觅食后,先投喂开口饵料,然后投喂微粒饲料(粗蛋白≥50%,粗脂肪≥8%,粗纤维≤3%,粗灰分≤16.5%,钙≤5%,总磷≥1%,水分≤12%,赖氨酸≥2%)(山东升索饲料科技有限公司),根据仔鱼的生长情况(体长)制定的开口饵料及微粒饲料调整方案如表1所示。30日龄试验鱼养殖至室外
水泥池(5m
×
5m
×
1m)中,饲料更换为商品饲料(粗蛋白≥30%,水分≤12%,粗纤维≤10%,粗灰分≤13%,粗脂肪≥3%,总磷≥1%,赖氨酸≥1.5%)(南京市澳华生物科技有限公司)。每日上、下午各投喂一次,日投喂量为鱼体重的3%。
24.表1开口饵料及微粒饲料调整方案
[0025][0026]
生长数据采集:在60日龄时,采用-氨基苯甲酸乙酯甲基磺酸盐(ms-222)(merck,德国)浸润麻醉,统计各组中试验鱼的存活情况,从各试验组中随机选取100尾试验鱼,测量试验鱼的体长、体重等生长数据。
[0027]
遗传性别鉴定:在60日龄时,从各试验组中随机选取20尾试验鱼,剪取试验鱼的尾鳍后,将其保存在无水乙醇中。取10~20mg的尾鳍组织,擦干剪碎后放置于1.5ml离心管中。基因组dna的提取采用dna isolation mini kit试剂盒(南京诺维赞生物科技有限公司),提取过程严格按照说明书进行,提取完成的基因组dna样本存放于-20℃冰箱中保存。采用课题组前期开发的斑点叉尾鮰遗传性别鉴定方法对试验鱼的遗传性别进行鉴定。
[0028]
解剖学观察:在60日龄时,从各试验组中随机采集100尾试验鱼,麻醉后割断鳃盖与腮丝间的动脉,血液放干后在试验鱼的肛门处做纵向切口,根据试验鱼规格调整切口长度(约3~4cm),观察试验鱼卵巢形成情况。同时,采集试验鱼的尾鳍组织,提取dna后鉴定遗传性别。根据鉴定结果统计各组中试验鱼的卵巢形成比例和性逆转率。
[0029]
组织学观察:取性腺保存在多聚甲醛溶液中,用于组织切片。性腺组织中的蛋白质经多聚甲醛保存后变性凝固,用低浓度到高浓度的酒精脱去性腺组织中的水分,置于二甲苯中透明后浸入融化的石蜡中,待石蜡浸入完全后进行包埋冷却。制作5~6μm的切片,采用he染色,最后滴上树胶封固。在光学显微镜(nikon,日本)下观察并拍照记录各试验组中性腺形成情况,在组织学水平观察t-30组中正常雌鱼,t-33组、t-36组中xy雌鱼的性腺发育情况。
[0030]
性别分化相关基因表达分析:采用qrt-pcr方法检测t-30组,t-36组雌雄性腺中foxl2和dmrt1基因mrna的相对表达量。采集150日龄t-30组雌雄试验鱼的精巢、卵巢组织,t-36组正常雌鱼和xy雌鱼的卵巢组织后,使用rna提取试剂盒(qiagen,德国)提取总rna,该过程严格按照说明书进行。使用prime script
tm rt master mix试剂盒(takara,日本)合成cdna,cdna合成后存放于-80℃冰箱。应用ncbi中primer-blast功能设计斑点叉尾鮰foxl2和dmrt1基因编码区的qrt-pcr反应引物,内参基因为α-tubulin,qrt-pcr反应引物如表2所示。qrt-pcr体系为25μl,包括12.5μl tb green染料(takara,日本),2μl cdna,上下游引物各1μl,5μl h2o。qrt-pcr程序为95℃30s,95℃15s,57℃30s,95℃15s,60℃15s,79℃5s,95℃15s,每个试验3次重复。采用2
‑△△
ct
值法计算foxl2和dmrt1基因在正常xx雌鱼、xy雌鱼卵巢中的相对表达量。
[0031]
表2 qrt-pcr反应引物表
[0032][0033][0034]
结果与分析
[0035]
温度对斑点叉尾鮰生长及性逆转的影响
[0036]
统计t-30、t-33和t-36组60日龄试验鱼的生长情况及卵巢形成情况发现,温度越高,试验鱼的存活率越低,当温度达到36℃时,试验鱼的存活率仅为82.67%,与其他两组呈显著性差异(p《0.05)。t-33组试验鱼的体长、体重均为三组中最高,t-36组试验鱼的体长、体重均为三组中最低,但是3组的生长数据无显著性差异(p》0.05)。t-30组卵巢形成比例为51%,雌雄鱼比例接近1:1,随着温度的提高,试验鱼的卵巢形成比例逐渐提高,性逆转率与温度呈正相关。各组在卵巢形成比例和性逆转率均呈极显著性差异(p《0.01)(如表3所示)。
[0037]
表3试验鱼生长特性和雌鱼比例
[0038][0039]
性逆转效果评估
[0040]
性逆转效果评估是在鉴定试验鱼遗传性别后,根据组织学观察和解剖学观察综合分析xy雌鱼卵巢发育情况(图1)。选取t-30、t-33、t-36组xx雌鱼和t-33、t-36组xy伪雌鱼的卵巢组织进行对比分析,结果发现各组的卵母细胞均呈圆形,t-30组xx雌鱼的部分卵母细胞发育至ⅲ期卵母细胞,而t-33组、t-36组中xx雌鱼的卵母细胞以ⅲ期卵母细胞为主,t-36组xy伪雌鱼中存在少量ⅲ期卵母细胞,而t-33组xy伪雌鱼中未见明显的ⅲ期卵母细胞。t-33组和t-36组xy伪雌鱼的卵母细胞数量少于xx雌鱼,且细胞内以1~2个核仁的卵母细胞为主,t-30组、t-33组和t-36组xx雌鱼的卵母细胞核内主要分布3个以上核仁。
[0041]
通过解剖学观察发现,t-30组、t-33组和t-36组之间卵巢发育无异,t-36组中xy伪雌鱼的卵巢发育正常,但t-33组中xy伪雌鱼的卵巢发育缓慢,卵巢纤细,且与体腔黏膜相连,轮廓不清晰。
[0042]
温度对性别分化相关基因表达量的影响
[0043]
对150日龄t-30组中雌鱼、雄鱼的卵巢和精巢组织,以及t-36组中xy伪雌鱼和xx雌鱼卵巢组织提取总mrna后,通过qrt-pcr方法检测雄性性别决定基因dmrt1和雌性性别决定基因foxl2在上述组织中的相对表达量(图2)。结果显示,dmrt1基因在正常雄鱼的精巢中的表达量最高,与t-30、t-36组中雌鱼的卵巢组织呈极显著性差异(p《0.01),与t-36组中xy伪雌鱼的卵巢组织无显著性差异。foxl2基因在t-36组xy雌鱼中被激活,与t-36组雌鱼,t-30组正常雌鱼、雄鱼的性腺组织呈显著性差异(p《0.05)。
[0044]
在已报道的大部分硬骨鱼类中,高温通常诱导其性别比例偏向于雄性,本试验中斑点叉尾鮰在高温诱导下性腺向雌性化方向分化。前人研究表明,当水温在19~31℃时,斑点叉尾鮰的特定生长率(sgr)和相对增重率(wgr)与温度呈正相关,并基于研究推断31℃并未达到斑点叉尾鮰的最适生长温度。在本试验中,t-33组试验鱼的生长数据最佳,这可能与33℃更贴近斑点叉尾鮰的最适生长温度有关。
[0045]
foxl2和dmrt1基因被认为是硬骨鱼类雌雄性别分化的标志基因,其在性别决定和分化中互为拮抗作用,foxl2基因对卵巢的分化和促进卵母细胞的发育有着重要作用,而dmrt1基因主要参与雄性的性别决定和精巢的分化,抑制卵母细胞发育。foxl2和dmrt1基因通过调控cyp19a1、sox9等下游性别分化相关基因的表达,使雌雄硬骨鱼体内雌二醇和睾酮等类固醇激素含量出现差异,从而达到调控雌雄性别分化的目的。研究发现,dmrt1基因对雄性斑马鱼的性腺维持起着重要作用,dmrt1基因的突变会导致雄性斑马鱼性逆转为可育雌性;使用crispr/cas9技术敲除异育银鲫(carassius gibelio)foxl2基因,导致其卵母细胞发育缓慢甚至发生性逆转。
[0046]
在本发明中,foxl2基因在xy伪雌鱼卵巢中的表达量显著高于其他3个组织,造成这一现象的可能原因是高温通过上调xy伪雌鱼foxl2基因的表达实现其性别逆转。然而,xy伪雌鱼的dmrt1基因在卵巢中的表达量仅低于正常雄鱼精巢,其表达并没有完全被抑制。因此,推测即使xy雌性斑点叉尾鮰dmrt1基因虽未被完全抑制,但是受高表达foxl2基因的影响,其性腺仍旧会发育成卵巢。
[0047]
最后所应当说明的是,以上实施例仅用以说明本发明的技术方案而非对本发明保护范围的限制,尽管参照较佳实施例对本发明作了详细说明,本领域的普通技术人员应当理解,可以对本发明的技术方案进行修改或者等同替换,而不脱离本发明技术方案的实质和范围。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1