赤霉素在延缓秋葵采后软化衰老的功效及应用
1.本发明属于蔬菜保存技术领域,具体涉及赤霉素在延缓秋葵采后软化衰老的功效及应用。
背景技术:
2.秋葵是一年生草本植物,属锦葵科,广泛生长在热带和亚热带国家。果实肉质柔嫩,口感润滑,具有独特的风味,因其特有的高维生素c、膳食纤维、钙以及低饱和脂肪酸被认为是理想的健康食品,并具有助消化、降血脂、庇护肝脏、延缓衰老、保护视力等功效。目前随着社会对于绿色、有机蔬菜关注度的不断提高,秋葵作为兼备营养与保健功能的蔬菜之一,已经获得了越来越多的重视。黄秋葵隶属于锦葵科秋葵属,作为果菜类,因其形态纤细、颇似手指被誉为“仙女的手指”,具有较高的商品潜在价值和加工附加值。黄秋葵是一种非呼吸跃变型果实,本身水分含量高、表面积大,极易失水、衰老、软化,严重影响其商品价值。因此,研究黄秋葵的贮藏保鲜,延缓采后软化衰老具有一定的实际应用价值。
3.果实发生软化表明果实已经发育成熟甚至已经开始衰老。已有研究表明,黄秋葵在贮藏过程中硬度不断下降,失重率不断升高,细胞壁发生降解导致果实软化衰老。赤霉素(ga)也被报道对延缓某些果实软化具有积极作用,如cn111657344a公开了一种采用赤霉素处理维持草莓硬度、保鲜的方法,但目前尚无采用赤霉素防止秋葵采后软化衰老的相关报道,更没有关于赤霉素延缓黄秋葵贮藏过程中失重、软化、细胞壁降解,及对细胞壁降解和果实软化有关酶基因在黄秋葵贮藏过程中的调控作用的相关报道。
技术实现要素:
4.为解决上述问题,本发明提供了赤霉素在延缓秋葵采后软化衰老的功效及应用,并提供了赤霉素在维持秋葵的硬度、失重率,抑制秋葵细胞壁降解方面的用途,提供了赤霉素在维持秋葵总果胶含量、或抑制可溶性果胶生成制剂的用途,提供了赤霉素在抑制秋葵多聚半乳糖醛酸酶aepg1、aepg2、aepg3、aepg4、aepg5、aepg6基因、秋葵果胶果酯酶aepme1或aepme2基因、秋葵果胶裂解酶aepl1、aepl2、aepl3、aepl4、aepl5基因、秋葵纤维素酶aecx基因、秋葵α-l-阿拉伯呋喃糖苷酶aeα-af1、aeα-af2、aeα-af3、aeα-af4、aeα-af5基因、秋葵β-半乳糖苷酶aeβ-gal1、aeβ-gal2、aeβ-gal3、aeβ-gal4、aeβ-gal5、aeβ-gal6基因的表达的用途。本发明以赤霉素处理的黄秋葵,通过测定黄秋葵在贮藏过程中的硬度、失重率及细胞壁组分含量变化,并利用qt-pcr技术研究黄秋葵贮藏期间细胞壁降解基因的表达情况,并筛选出参与黄秋葵细胞壁降解的重要基因,并分析这些基因在赤霉素处理组与对照组间表达差异,为赤霉素处理在延缓黄秋葵贮藏过程中软化衰老,延长保鲜时间提供理论依据。
5.一方面,本发明提供了赤霉素用于制备抑制秋葵细胞壁降解制剂的用途。
6.本发明还提供了赤霉素用于制备维持秋葵硬度、或维持失重率的用途。
7.进一步地,所述赤霉素的质量百分含量为0.001%~0.05%(10mg/l~500mg/l)。
8.大多数的果实在软化时表现为硬度的快速下降;果实失水失重率升高;果实表面
颜色发生变化;原果胶降解,水溶性果胶增加的现象。近年来对采后果实软化的研究主要集中在细胞壁物质含量及有关酶的作用上,许多研究表明果实贮藏过程中细胞壁所含糖类及其他物质会不断降解导致细胞壁结构遭到破坏,最终导致果实软化腐烂。果胶、纤维素和半纤维素是构成细胞壁的主要成分,这三者通过糖苷键和氢键交联在一起,能维持细胞壁结构保持相对稳定的状态,果实在贮藏过程中这些物质会被水解从而导致细胞壁结构发生变化,继而引起果实的软化衰老。
9.黄秋葵在黄秋葵嫩荚中含有大量果胶物质,测量不同品种和不同组织部位黄秋葵果胶含量发现最高品种果胶含量可达24.05%。黄秋葵在贮藏过程中硬度不断下降,失重率不断升高,细胞壁发生降解导致果实软化衰老。
10.可以理解,本发明提供的赤霉素对各种秋葵都具有抑制秋葵细胞壁降解、维持秋葵硬度、或维持失重率的用途。在一些实施方式中,所述秋葵优选采用黄秋葵。
11.本发明经大量研究发现,经赤霉素处理后的黄秋葵,在贮藏期间能抑制水分的流失,使秋葵细胞具有较好的持水能力,能有效控制果实的失重,并能有效缓解硬度的降低和果实的衰老,大幅增加黄秋葵的贮藏保鲜时间;并且赤霉素还能延缓黄秋葵细胞壁的降解,抑制秋葵在贮藏期间细胞壁中纤维素和总果胶含量的降低,抑制可溶性果胶的生成,维持细胞壁的完整性,从而延缓果实的软化衰老。
12.另一方面,本发明提供了赤霉素用于制备维持秋葵纤维素含量制剂的用途。
13.进一步地,所述赤霉素的质量百分含量为0.001%~0.05%。
14.研究证明,经赤霉素处理后的秋葵,能有效抑制细胞壁的降解,维持细胞壁中的纤维素的含量,抑制纤维素的降解。
15.再一方面,本发明提供了赤霉素用于制备维持秋葵总果胶含量、或抑制可溶性果胶生成制剂的用途。
16.进一步地,所述赤霉素的质量百分含量为0.001%~0.05%。
17.黄秋葵在贮藏过程中硬度不断下降,失重率不断升高,原果胶不断降解,可溶性果胶不断升高,细胞壁发生降解导致果实软化衰老。
18.本发明经大量研究发现,赤霉素处理在一定程度上能抑制原果胶的降解和可溶性果胶的生成,延缓了细胞壁的降解,从而维持细胞壁的完整性,延缓果实在贮藏过程中的软化衰老。
19.再一方面,本发明提供了赤霉素用于制备抑制秋葵多聚半乳糖醛酸酶aepg1、aepg2、aepg3、aepg4、aepg5、aepg6基因中的任意一种或多种基因表达制剂的用途。
20.进一步地,所述aepg1、aepg2、aepg3、aepg4、aepg5、aepg6基因的序列分别如seq id no.1、seq id no.2、seq id no.3、seq id no.4、seq id no.5、seq id no.6所示;所述赤霉素的质量百分含量为0.001%~0.05%。
21.参与细胞壁代谢的酶多达30多种,但一般认为pg、pme、pl、cx、α-af、β-gal等与细胞壁的降解和果实的软化有着密切的联系。pg、pme和pl合称为果胶酶,三者的协同作用被广泛认为能导致采后果蔬中果胶的分解,在决定果胶含量中起重要作用。
22.黄秋葵的多聚半乳糖醛酸酶pg是与秋葵果实软化相关的主要酶类,在果实采后贮藏过程中,该酶活性的变化与果肉细胞壁结构的变化密切相关,进而影响到果实的硬度变化,测定酶活性对揭示果实采后软化机理具有重要意义,pg酶基因表达变化趋势与pg酶活
性变化趋势一致。
23.本发明通过转录组测序筛选到黄秋葵多聚半乳糖醛酸酶pg的相关基因,包括aepg1、aepg2、aepg3、aepg4、aepg5和aepg6。研究发现赤霉素能抑制aepg1、aepg2、aepg3、aepg4、aepg5和aepg6的表达,从而延缓果实软化衰老的发生。
24.在一些方式中,用0.001%~0.05%的赤霉素溶液浸泡处理秋葵,都能抑制秋葵aepg1、aepg2、aepg3、aepg4、aepg5、aepg6中的任意一种或多种基因的表达。
25.再一方面,本发明提供了赤霉素用于制备抑制秋葵果胶果酯酶aepme1或aepme2基因表达制剂的用途。
26.进一步地,所述aepme1、aepme2基因的序列分别如seq id no.7、seq id no.8所示;所述赤霉素的质量百分含量为0.001%~0.05%。
27.黄秋葵的果胶果酯酶pme是细胞壁相关的果胶水解酶蛋白,通过使果胶去酯化产生水溶性的果胶酸和甲酯,使黄秋葵果实软化脱落和衰老。
28.本发明通过转录组测序筛选到黄秋葵果胶果酯酶pme的相关基因,包括aepme1、aepme2。研究发现赤霉素能抑制aepme1、aepme2的表达,从而延缓果实软化衰老的发生。
29.再一方面,本发明提供了赤霉素用于制备抑制秋葵果胶裂解酶aepl1、aepl2、aepl3、aepl4、aepl5基因中的任意一种或多种基因表达制剂的用途。
30.进一步地,所述aepl1、aepl2、aepl3、aepl4、aepl5基因的序列分别如seq id no.9、seq id no.10、seq id no.11、seq id no.12、seq id no.13所示;所述赤霉素的质量百分含量为0.001%~0.05%。
31.黄秋葵的果胶裂解酶pl能催化果胶分子链的消除裂解,通过反式消去作用裂解果胶聚合体,使黄秋葵果实软化衰老。
32.本发明通过转录组测序筛选到黄秋葵果胶裂解酶pl的相关基因,包括aepl1、aepl2、aepl3、aepl4、aepl5。研究发现赤霉素能抑制aepl1、aepl2、aepl3、aepl4、aepl5的表达,从而延缓果实软化衰老的发生。
33.再一方面,本发明提供了赤霉素用于制备抑制秋葵纤维素酶aecx基因表达制剂的用途。
34.进一步地,所述aecx基因的序列分别如seq id no.14所示;所述赤霉素的质量百分含量为0.001%~0.05%。
35.黄秋葵的纤维素酶cx能够降解羧甲基纤维素,在果实成熟软化中起重要作用,但在果实采后会使黄秋葵果实软化衰老。
36.本发明通过转录组测序筛选到黄秋葵纤维素酶cx的相关基因,包括aecx。研究发现赤霉素能抑制aecx的表达,从而延缓果实软化衰老的发生。
37.再一方面,本发明提供了赤霉素用于制备抑制秋葵α-l-阿拉伯呋喃糖苷酶aeα-af1、aeα-af2、aeα-af3、aeα-af4、aeα-af5基因中的任意一种或多种基因表达制剂的用途。
38.进一步地,所述aeα-af1、aeα-af2、aeα-af3、aeα-af4、aeα-af5基因的序列分别如seq id no.15、seq id no.16、seq id no.17、seq id no.18、seq id no.19所示;所述赤霉素的质量百分含量为0.001%~0.05%。
39.黄秋葵的α-l-阿拉伯呋喃糖苷酶α-af是降解细胞壁多糖的重要酶,在果实的成熟软化中有非常重要的作用,但在果实采后会使黄秋葵果实软化衰老。
40.本发明通过转录组测序筛选到黄秋葵α-l-阿拉伯呋喃糖苷酶α-af的相关基因,包括aeα-af1、aeα-af2、aeα-af3、aeα-af4、aeα-af5。研究发现赤霉素能抑制aeα-af1、aeα-af2、aeα-af3、aeα-af4、aeα-af5的表达,从而延缓果实软化衰老的发生。
41.再一方面,本发明提供了赤霉素用于制备抑制秋葵β-半乳糖苷酶aeβ-gal1、aeβ-gal2、aeβ-gal3、aeβ-gal4、aeβ-gal5、aeβ-gal6基因中的任意一种或多种基因表达制剂的用途。
42.进一步地,所述aeβ-gal1、aeβ-gal2、aeβ-gal3、aeβ-gal4、aeβ-gal5、aeβ-gal6基因的序列分别如seq id no.20、seq id no.21、seq id no.22、seq id no.23、seq id no.24、seq id no.15所示;所述赤霉素的质量百分含量为0.001%~0.05%。
43.黄秋葵的β-半乳糖苷酶β-gal能降解细胞壁组分,导致细胞壁结构破坏,引起果实软化,但在果实采后会使黄秋葵果实软化衰老。
44.本发明通过转录组测序筛选到黄秋葵β-半乳糖苷酶β-gal的相关基因,包括aeβ-gal1、aeβ-gal2、aeβ-gal3、aeβ-gal4、aeβ-gal5、aeβ-gal6。研究发现赤霉素能抑制aeβ-gal1、aeβ-gal2、aeβ-gal3、aeβ-gal4、aeβ-gal5、aeβ-gal6的表达,从而延缓果实软化衰老的发生。
45.再一方面,本发明提供了一种基因用于促进秋葵软化衰老的用途,所述基因包括aepg1、aepg2、aepg3、aepg4、aepg5、aepg6、aepme1、aepme2、aepl1、aepl2、aepl3、aepl4、aepl5、aecx、aeα-af1、aeα-af2、aeα-af3、aeα-af4、aeα-af5、aeβ-gal1、aeβ-gal2、aeβ-gal3、aeβ-gal4、aeβ-gal5、aeβ-gal6,其序列分别如seq id no.1~seq id no.25所示。
46.本发明先经大量转录测序工作,从秋葵的基因中筛选到抑制秋葵软化衰老的相关基因,明确相关基因的具体序列,并考察了秋葵贮藏期间各基因表达情况的变化趋势。
47.本发明提供的赤霉素在延缓秋葵采后软化衰老的应用具有以下有益效果:
48.1、发现了赤霉素在维持秋葵硬度、或维持失重率方面的用途,可用于制备维持秋葵硬度、或维持失重率的制剂;
49.2、发现了赤霉素在抑制秋葵细胞壁降解方面的用途,可用于制备抑制秋葵细胞壁降解的制剂;
50.3、发现了赤霉素在维持秋葵纤维素含量、维持黄秋葵总果胶含量、或抑制可溶性果胶生成方面的用途,可用于制备维持秋葵纤维素含量、维持秋葵总果胶含量、或抑制可溶性果胶生成的制剂;
51.4、发现了赤霉素在抑制秋葵多聚半乳糖醛酸酶aepg1、aepg2、aepg3、aepg4、aepg5、aepg6基因表达的用途,可用于制备抑制秋葵多聚半乳糖醛酸酶aepg1、aepg2、aepg3、aepg4、aepg5、aepg6基因表达的制剂;
52.5、发现了赤霉素在抑制秋葵果胶果酯酶aepme1、aepme2基因表达的用途,可用于制备抑制秋葵果胶果酯酶aepme1、aepme2基因表达的制剂;
53.6、发现了赤霉素在抑制秋葵果胶裂解酶aepl1、aepl2、aepl3、aepl4、aepl5基因表达的用途,可用于制备抑制秋葵果胶裂解酶aepl1、aepl2、aepl3、aepl4、aepl5基因表达的制剂;
54.7、发现了赤霉素在抑制秋葵纤维素酶aecx基因表达的用途,可用于制备抑制秋葵纤维素酶aecx基因表达的制剂;
55.8、发现了赤霉素在抑制秋葵α-l-阿拉伯呋喃糖苷酶aeα-af1、aeα-af2、aeα-af3、aeα-af4、aeα-af5表达的用途,可用于制备抑制秋葵α-l-阿拉伯呋喃糖苷酶aeα-af1、aeα-af2、aeα-af3、aeα-af4、aeα-af5表达的制剂;
56.9、发现了赤霉素在抑制秋葵β-半乳糖苷酶aeβ-gal1、aeβ-gal2、aeβ-gal3、aeβ-gal4、aeβ-gal5、aeβ-gal6表达的用途,可用于制备抑制秋葵β-半乳糖苷酶aeβ-gal1、aeβ-gal2、aeβ-gal3、aeβ-gal4、aeβ-gal5、aeβ-gal6表达的制剂;
57.10、为赤霉素处理在延缓黄秋葵贮藏过程中软化衰老、延长保鲜时间提供理论依据。
附图说明
58.图1为实施例1中赤霉素处理组与对照组对黄秋葵失重率影响对比图,其中*,**,***分别表示在0.05、0.01、0.001水平上差异显著;
59.图2为实施例2中赤霉素处理组与对照组对黄秋葵硬度影响对比图,其中*,**,***分别表示在0.05、0.01、0.001水平上差异显著;
60.图3为实施例3中赤霉素处理组与对照组对黄秋葵的细胞壁成分含量影响对比图,其中*,**,***分别表示在0.05、0.01、0.001水平上差异显著;
61.图4为实施例4中aepg1与木槿的多聚半乳糖醛酸酶pg相关基因序列比对结果;
62.图5为实施例4中aepg2与木槿的多聚半乳糖醛酸酶pg相关基因序列比对结果;
63.图6为实施例4中aepg3与木槿的多聚半乳糖醛酸酶pg相关基因序列比对结果;
64.图7为实施例4中aepg4与木槿的多聚半乳糖醛酸酶pg相关基因序列比对结果;
65.图8为实施例4中aepg5与木槿的多聚半乳糖醛酸酶pg相关基因序列比对结果;
66.图9为实施例4中aepg6与木槿的多聚半乳糖醛酸酶pg相关基因序列比对结果;
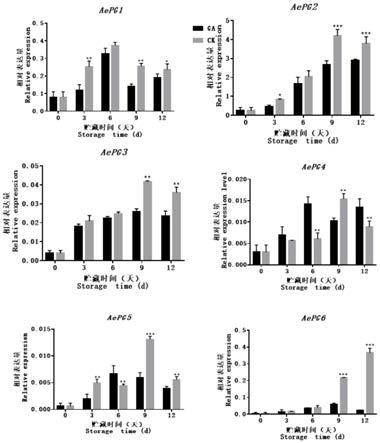
67.图10为实施例4中赤霉素处理组与对照组对黄秋葵aepg1、aepg2、aepg3、aepg4、aepg5和aepg6基因表达影响对比图,其中*,**,***分别表示在0.05、0.01、0.001水平上差异显著;
68.图11为实施例5中赤霉素处理组与对照组对黄秋葵aepme1和aepme2基因表达影响对比图,其中*,**,***分别表示在0.05、0.01、0.001水平上差异显著;
69.图12为实施例6中aepl2与木槿、棉花的果胶裂解酶pl相关基因序列比对结果;
70.图13为实施例6中aepl3与木槿、棉花的果胶裂解酶pl相关基因序列比对结果;
71.图14为实施例6中赤霉素处理组与对照组对黄秋葵aepl1、aepl2、aepl3、aepl4和aepl5基因表达影响对比图,其中*,**,***分别表示在0.05、0.01、0.001水平上差异显著;
72.图15为实施例7中赤霉素处理组与对照组对黄秋葵aecx基因表达影响对比图,其中*,**,***分别表示在0.05、0.01、0.001水平上差异显著;
73.图16为实施例8中aeα-af1与木槿、棉花的α-l-阿拉伯呋喃糖苷酶α-af相关基因序列比对结果;
74.图17为实施例8中aeα-af2与木槿、棉花的α-l-阿拉伯呋喃糖苷酶α-af相关基因序列比对结果;
75.图18为实施例8中aeα-af3与木槿、棉花的α-l-阿拉伯呋喃糖苷酶α-af相关基因序列比对结果;
76.图19为实施例8中aeα-af4与木槿、棉花的α-l-阿拉伯呋喃糖苷酶α-af相关基因序列比对结果;
77.图20为实施例8中aeα-af5与木槿、棉花的α-l-阿拉伯呋喃糖苷酶α-af相关基因序列比对结果;
78.图21为实施例8中赤霉素处理组与对照组对黄秋葵aeα-af1、aeα-af2、aeα-af3、aeα-af4和aeα-af5基因表达影响对比图,其中*,**,***分别表示在0.05、0.01、0.001水平上差异显著;
79.图22为实施例9中aeβ-gal1与木槿、棉花、榴莲、可可的β-半乳糖苷酶β-gal相关基因序列比对结果;
80.图23为实施例9中aeβ-gal2与木槿、棉花、榴莲、可可的β-半乳糖苷酶β-gal相关基因序列比对结果;
81.图24为实施例9中aeβ-gal3与木槿、棉花、榴莲、可可的β-半乳糖苷酶β-gal相关基因序列比对结果;
82.图25为实施例9中aeβ-gal4与木槿、棉花、榴莲、可可的β-半乳糖苷酶β-gal相关基因序列比对结果;
83.图26为实施例9中aeβ-gal5与木槿、棉花、榴莲、可可的β-半乳糖苷酶β-gal相关基因序列比对结果;
84.图27为实施例9中赤霉素处理组与对照组对黄秋葵aeβ-gal1、aeβ-gal2、aeβ-gal3、aeβ-gal4、aeβ-gal5和aeβ-gal6基因表达影响对比图,其中*,**,***分别表示在0.05、0.01、0.001水平上差异显著。
具体实施方式
85.下面结合附图对本发明的优选实施例作进一步详细描述,需要指出的是,以下实施例旨在便于对本发明的理解,而对其不起任何限定作用,本发明的实施例中公开的所有特征,或公开的所有方法或过程中的步骤,除了互相排斥的特征和/或步骤以外,均能够以任何方式组合。
86.实施例1赤霉素处理对黄秋葵失重率的影响
87.本实施例采用新采摘的黄秋葵(购自宁波市鄞州区联心菜市场),选择大小较为统一,个头饱满,没有黑斑及损伤的黄秋葵200根,分成2组(对照组和处理组),每组各100根。配制质量百分比含量为0.01%(100mg/l)的赤霉素水溶液,将处理组的黄秋葵在赤霉素溶液中浸泡15min。浸泡结束后,将2组黄秋葵置于常温下自然风干,最后放置于恒温恒湿培养箱中于25
±
1℃,80%~90%相对湿度下贮藏12天。分别在处理前(0天)和处理后贮藏3、6、9、12天取样,测定对照组和处理组黄秋葵的失重变化情况,计算失重率。
88.黄秋葵的失重率的检测方法:采用质量损失法。选取10个果实,每天同一时间取出用天平称取质量,重复三次测定。计算贮藏前、后的质量差占贮藏前样品质量的百分比,即失重率,计算公式如下:
[0089][0090]
两组黄秋葵的失重率变化情况如图1所示,其中*,**,***分别表示在0.05、0.01、
0.001水平上差异显著。
[0091]
由图1可知,黄秋葵在贮藏期间处理组和对照组的失重率都随着贮藏时间的增加不断升高,直至贮藏结束,处理组失重率上升至2.9%而对照组上升至3.3%,整个贮藏期间对照组失重率始终高于处理组,并在9d和12d时具有显著性(p﹤0.05)。因此,说明赤霉素处理在贮藏期间特别是贮藏后期能较好地抑制黄秋葵水分的流失,使黄秋葵细胞具有较好的持水能力,有效地控制果实的失重。
[0092]
本实施例还采用了0.001%、0.003%、0.005%、0.007%、0.01%、0.03%、0.05%赤霉素水溶液分别进行试验,都能起到一定的防止黄秋葵水分流失、延缓失重率上升的效果,因篇幅的限制,这里就不一一举例说明。
[0093]
实施例2赤霉素处理对黄秋葵硬度的影响
[0094]
本实施例采用新采摘的黄秋葵(购自宁波市鄞州区联心菜市场),选择大小较为统一,个头饱满,没有黑斑及损伤的黄秋葵200根,分成2组(对照组和处理组),每组各100根。配制质量百分比含量为0.01%的赤霉素水溶液,将处理组的黄秋葵在赤霉素溶液中浸泡15min。浸泡结束后,将2组黄秋葵置于常温下自然风干,最后放置于恒温恒湿培养箱中于25
±
1℃,80%~90%相对湿度下贮藏12天。分别在处理前(0天)和处理后贮藏3、6、9、12天取样,测定对照组和处理组黄秋葵的硬度变化情况。
[0095]
黄秋葵硬度测定方法:采用tms-touch物性分析仪(美国ftc公司)进行黄秋葵果实硬度的测定。挑选8个果实,对其果实腰部最大横径处进行测定,重复三次测定,结果以n表示。
[0096]
两组黄秋葵的硬度变化情况如图2所示,其中*,**,***分别表示在0.05、0.01、0.001水平上差异显著。
[0097]
由图2可见,对照组在0-12d贮藏期间硬度持续下降,贮藏末期的硬度为17.2n,与贮藏之前的硬度相比下降了32.1%。处理组在贮藏前期较好的保持了果实硬度,到贮藏末期时的硬度为20.4n,与贮藏之前的硬度相比下降了19.4%。处理组在整个贮藏过程中果实硬度始终显著高于对照组(p﹤0.05)。以上结果表明,赤霉素处理能有效延缓黄秋葵贮藏期间硬度的降低和果实的衰老,一定程度上增加黄秋葵贮藏保鲜时间。
[0098]
本实施例还采用了0.001%、0.003%、0.005%、0.007%、0.01%、0.03%、0.05%赤霉素水溶液分别进行试验,都能起到一定的延缓黄秋葵贮藏期间硬度降低和果实衰老的效果,因篇幅的限制,这里就不一一举例说明。
[0099]
实施例3赤霉素处理对黄秋葵细胞壁成分含量的影响
[0100]
本实施例采用新采摘的黄秋葵(购自宁波市鄞州区联心菜市场),选择大小较为统一,个头饱满,没有黑斑及损伤的黄秋葵200根,分成2组(对照组和处理组),每组各100根。配制质量百分比含量为0.01%的赤霉素水溶液,将处理组的黄秋葵在赤霉素溶液中浸泡15min。浸泡结束后,将2组黄秋葵置于常温下自然风干,最后放置于恒温恒湿培养箱中于25
±
1℃,80%~90%相对湿度下贮藏12天。分别在处理前(0天)和处理后贮藏3、6、9、12天取样,测定对照组和处理组黄秋葵细胞壁成分含量的变化情况。细胞壁成分含量的变化情况,主要包括黄秋葵贮藏期间的原果胶、可溶性果胶和纤维素含量的变化情况。黄秋葵贮藏期间原果胶会逐渐分解,并生成可溶性果胶,纤维素也逐渐降解,果实逐渐衰老软化。
[0101]
其中,黄秋葵的果胶类物质的检测方法为:采用咔唑比色法测定。以处理后的半乳
糖醛酸质量为横坐标,波长530nm处的吸光度值为纵坐标,绘制并计算果胶含量的标准曲线。分别将提取的原果胶和可溶性果胶1.0ml提取液加入到25ml刻度试管中,按照标准曲线的操作步骤测定样品中原果胶和可溶性果胶含量,重复三次测定,原果胶和可溶性果胶含量之和为总果胶含量。
[0102]
黄秋葵的纤维素含量测定方法为:采用蒽酮比色法测定。以处理后的纤维素含量为横坐标,波长620nm处的吸光度值为纵坐标,绘制并计算纤维素含量的标准曲线。将稀释后的1.0ml溶液加入到25ml刻度试管中,按照标准曲线的操作步骤测定样品中纤维素含量,重复三次测定。
[0103]
两组黄秋葵的细胞壁成分含量变化情况如图3所示,其中*,**,***分别表示在0.05、0.01、0.001水平上差异显著。
[0104]
由图3可以看出,黄秋葵在贮藏前期处理组和对照组中的原果胶含量下降缓慢,并无显著性差异(p>0.05),但在贮藏后期处理组和对照组原果胶含量都开始大幅度下降,对照组的下降速率远高于处理组,且在9d和12d时,处理组中原果胶含量显著高于对照组(p﹤0.05),贮藏结束时,处理组和对照组中原果胶含量分别下降了31.27%和50.04%,可见经赤霉素处理后,黄秋葵在贮藏期间能有效延缓原果胶含量的下降。
[0105]
黄秋葵在贮藏期间处理组和对照组中的可溶性果胶含量都呈逐渐上升趋势,对照组的上升速率大于处理组。在整个贮藏期间,对照组的可溶性果胶含量都大于处理组,且在6d、9d和12d时存在显著差异(p﹤0.05);贮藏结束时,处理组和对照组中可溶性果胶含量分别为5.24mg/g和7.08mg/g;可见经赤霉素处理后,黄秋葵在贮藏期间能有效抑制可溶性果胶的生成。
[0106]
两组黄秋葵的总果胶含量总体呈现小幅上升的趋势,但在9d时对照组中的总果胶含量突然发生了下降,在贮藏期间处理组和对照组的总果胶含量无显著差异(p>0.05)。这主要是因为总果胶为原果胶和可溶性果胶的总含量,黄秋葵贮藏期间,原果胶逐渐被降解为可溶性果胶,因此原果胶含量不断下降,可溶性果胶含量不断上升,果实逐渐软化衰老,但总果胶含量的变化并不明显,因此两组黄秋葵的总果胶含量无显著差异。
[0107]
黄秋葵经外源ga处理后贮藏期间纤维素含量变化如图3所示。除处理组在3d时小幅升高外,在整个贮藏期间处理组和对照组的纤维素含量都呈现了不同程度的下降趋势,处理组的下降速率低于对照组,处理组在整个贮藏期间的纤维素含量也高于对照组,且在9d和12d时处理组中纤维素含量显著高于对照组(p﹤0.05),直至贮藏结束处理组和对照组的纤维素含量分别下降了19.5%和38.5%,可见经赤霉素处理后,黄秋葵在贮藏期间能有效抑制纤维素的降解。
[0108]
上述结果表明赤霉素处理在一定程度上能抑制原果胶的降解和可溶性果胶的生成,抑制纤维素的降解,延缓了细胞壁的降解,从而维持细胞壁的完整性,延缓果实在贮藏过程中的软化衰老。
[0109]
本实施例还采用了0.001%、0.003%、0.005%、0.007%、0.01%、0.03%、0.05%赤霉素水溶液分别进行试验,都能起到一定的抑制黄秋葵贮藏期间抑制原果胶的降解、可溶性果胶的生成,以及纤维素的降解的效果,因篇幅的限制,这里就不一一举例说明。
[0110]
实施例4赤霉素处理对黄秋葵多聚半乳糖醛酸酶pg相关基因表达的影响
[0111]
本实施例采用新采摘的黄秋葵(购自宁波市鄞州区联心菜市场),选择大小较为统
一,个头饱满,没有黑斑及损伤的黄秋葵200根,分成2组(对照组和处理组),每组各100根。配制质量百分比含量为0.01%的赤霉素水溶液,将处理组的黄秋葵在赤霉素溶液中浸泡15min。浸泡结束后,将2组黄秋葵置于常温下自然风干,最后放置于恒温恒湿培养箱中于25
±
1℃,80%~90%相对湿度下贮藏12天。分别在处理前(0天)和处理后贮藏3、6、9、12天取样,测定对照组和处理组黄秋葵的多聚半乳糖醛酸酶pg相关基因aepg1、aepg2、aepg3、aepg4、aepg5、aepg6表达的变化情况。
[0112]
本实施例通过转录组测序筛选到黄秋葵的多聚半乳糖醛酸酶pg的相关基因(委托广州基迪奥生物科技有限公司进行转录组测序),包括aepg1、aepg2、aepg3、aepg4、aepg5、aepg6,其序列分别如seq id no.1~seq id no.6所示。将aepg1、aepg2、aepg3、aepg4、aepg5、aepg6的序列分别在ncbi上跟棉花、木槿的多聚半乳糖醛酸酶pg相关基因序列进行比对,结果分别如图4~9所示,其中aepg1的比对结果见图4,aepg2的比对结果见图5,aepg3的比对结果见图6,aepg4的比对结果见图7,aepg5的比对结果见图8,aepg6的比对结果见图9,可以看出,比对结果达到高度相似,证明aepg1、aepg2、aepg3、aepg4、aepg5、aepg6基因确实为黄秋葵的多聚半乳糖醛酸酶pg相关基因。
[0113]
总rna提取和cdna合成方法为:称取约0.2g果实冻样,用柱式植物rna提取试剂盒2.0(北京天恩泽基因科技有限公司)提取黄秋葵的总rna。利用核酸蛋白仪与琼脂糖电泳仪来检测提取rna的质量。cdna采用hifiscript cdna synthesis kit试剂盒(北京康为生物有限公司)反转录而成。
[0114]
本实施例通过实时定量pcr分析秋葵中aepg1、aepg2、aepg3、aepg4、aepg5、aepg6表达的变化情况,aepg1、aepg2、aepg3、aepg4、aepg5、aepg6的特异性引物由beacon designer 7设计,以aeact为内参基因,引物序列见表1。采用美国赛默飞旗下的real-time pcr仪,按照美国赛默飞世尔公司的dynamo flash sybr green qpcr试剂盒说明书,进行三步法扩增。进行如下程序设置:95℃预变性7min;95℃,15s,45℃退火30s,75℃延伸15s,此步骤进行39个循环。每个样品进行4次生物学重复,相对表达量采用2-δct
方法进行计算分析。
[0115]
表1荧光定量pcr
[0116][0117]
两组黄秋葵的aepg1、aepg2、aepg3、aepg4、aepg5和aepg6基因表达情况如图10所示,其中*,**,***分别表示在0.05、0.01、0.001水平上差异显著。
[0118]
由图10可见,对照组在贮藏期间除aepg6呈逐渐上升的变化趋势外,aepg1、aepg2、aepg3、aepg4、aepg5都呈先上升后下降的趋势。处理组在贮藏期间aepgs基因的表达模式多
样,其中aepg1、aepg4在贮藏前期表达上升,贮藏中期表达下降,到贮藏末期再呈上升的趋势,aepg2呈不断上升趋势,而aepg3、aepg5、aepg6呈先上升后下降的趋势。赤霉素处理促进了aepg4和aepg5在贮藏期间某几个时间点的表达而抑制了aepg1、aepg2、aepg3、aepg6在整个贮藏期间的表达,并在9d和12d时都显著抑制表达(p﹤0.05),且在这四个基因中的表达强度远高于aepg4和aepg5,因此赤霉素处理黄秋葵,总体对黄秋葵的aepgs基因是起抑制表达作用。
[0119]
本实施例还采用了0.001%、0.003%、0.005%、0.007%、0.01%、0.03%、0.05%赤霉素水溶液分别进行试验,都能起到一定的抑制黄秋葵aepgs基因表达的效果,因篇幅的限制,这里不一一举例说明。
[0120]
实施例5赤霉素处理对黄秋葵果胶果酯酶pme相关基因表达的影响
[0121]
本实施例采用新采摘的黄秋葵(购自宁波市鄞州区联心菜市场),选择大小较为统一,个头饱满,没有黑斑及损伤的黄秋葵200根,分成2组(对照组和处理组),每组各100根。配制质量百分比含量为0.01%的赤霉素水溶液,将处理组的黄秋葵在赤霉素溶液中浸泡15min。浸泡结束后,将2组黄秋葵置于常温下自然风干,最后放置于恒温恒湿培养箱中于25
±
1℃,80%~90%相对湿度下贮藏12天。分别在处理前(0天)和处理后贮藏3、6、9、12天取样,测定对照组和处理组黄秋葵的果胶果酯酶pme相关基因aepme1或aepme2表达的变化情况。
[0122]
本实施例通过转录组测序筛选到黄秋葵的果胶果酯酶pme的相关基因(委托广州基迪奥生物科技有限公司进行转录组测序),包括aepme1和aepme2,其序列分别如seq id no.7、seq id no.8所示。将aepme1和aepme2的序列在ncbi上跟棉花、可可、木槿的果胶果酯酶pme相关基因序列进行比对,对比结果显示达到高度相似,证明aepme1和aepme2基因确实为黄秋葵的果胶果酯酶pme相关基因。
[0123]
总rna提取和cdna合成方法为:称取约0.2g果实冻样,用柱式植物rna提取试剂盒2.0(北京天恩泽基因科技有限公司)提取黄秋葵的总rna。利用核酸蛋白仪与琼脂糖电泳仪来检测提取rna的质量。cdna采用hifiscript cdna synthesis kit试剂盒(北京康为生物有限公司)反转录而成。
[0124]
本实施例通过实时定量pcr分析秋葵中aepme1和aepme2表达的变化情况,aepme1和aepme2的特异性引物由beacon designer 7设计,以aeact为内参基因,引物序列见表2。采用美国赛默飞旗下的real-time pcr仪,按照美国赛默飞世尔公司的dynamo flash sybr green qpcr试剂盒说明书,进行三步法扩增。进行如下程序设置:95℃预变性7min;95℃,15s,45℃退火30s,75℃延伸15s,此步骤进行39个循环。每个样品进行4次生物学重复,相对表达量采用2-δct
方法进行计算分析。
[0125]
表2荧光定量pcr
[0126][0127]
两组黄秋葵的aepme1和aepme2基因表达情况如图11所示,其中*,**,***分别表示
在0.05、0.01、0.001水平上差异显著。
[0128]
由图11可知,黄秋葵在贮藏期间处理组和对照组aepme1表达模式均呈先上升后下降的趋势,不同的是对照组上升至9d后开始下降而处理组上升至6d后就开始下降,且在整个贮藏期间aepme1在对照组中的表达量显著高于处理组(p﹤0.05),可见赤霉素处理对黄秋葵的aepme1基因表达具有明显的抑制作用。
[0129]
贮藏期间aepme2在处理组中的表达模式为随着贮藏时间的延长缓慢上升,在对照组的贮藏前期也表现为缓慢上升但到了贮藏后期则表现为急剧升高,使得贮藏后期aepme2在对照组中的表达显著高于处理组(p﹤0.05)。可见赤霉素处理对黄秋葵的aepme2基因表达具有明显的抑制作用。
[0130]
因此,赤霉素处理有效抑制了aepme1和aepme2的表达。
[0131]
本实施例还采用了0.001%、0.003%、0.005%、0.007%、0.01%、0.03%、0.05%赤霉素水溶液分别进行试验,都能起到一定的抑制黄秋葵aepme1和aepme2基因表达的效果,因篇幅的限制,这里就不一一举例说明。
[0132]
实施例6赤霉素处理对黄秋葵果胶裂解酶pl相关基因表达的影响
[0133]
本实施例采用新采摘的黄秋葵(购自宁波市鄞州区联心菜市场),选择大小较为统一,个头饱满,没有黑斑及损伤的黄秋葵200根,分成2组(对照组和处理组),每组各100根。配制质量百分比含量为0.01%的赤霉素水溶液,将处理组的黄秋葵在赤霉素溶液中浸泡15min。浸泡结束后,将2组黄秋葵置于常温下自然风干,最后放置于恒温恒湿培养箱中于25
±
1℃,80%~90%相对湿度下贮藏12天。分别在处理前(0天)和处理后贮藏3、6、9、12天取样,测定对照组和处理组黄秋葵的果胶裂解酶pl相关基因aepl1、aepl2、aepl3、aepl4、aepl5表达的变化情况。
[0134]
本实施例通过转录组测序筛选到黄秋葵的果胶裂解酶pl的相关基因(委托广州基迪奥生物科技有限公司进行转录组测序),包括aepl1、aepl2、aepl3、aepl4和aepl5,其序列分别如seq id no.9~seq id no.13所示。将aepl1、aepl2、aepl3、aepl4和aepl5的序列在ncbi上跟木槿、棉花的果胶裂解酶pl相关基因序列进行比对(因篇幅有限,这里列举部分对比结果如图12~13所示,其中aepl2的比对结果见图12,aepl3的比对结果见图13),结果都证明达到高度相似,证明aepl1、aepl2、aepl3、aepl4、aepl5基因确实为黄秋葵的果胶裂解酶pl相关基因。
[0135]
总rna提取和cdna合成方法为:称取约0.2g果实冻样,用柱式植物rna提取试剂盒2.0(北京天恩泽基因科技有限公司)提取黄秋葵的总rna。利用核酸蛋白仪与琼脂糖电泳仪来检测提取rna的质量。cdna采用hifiscript cdna synthesis kit试剂盒(北京康为生物有限公司)反转录而成。
[0136]
本实施例通过实时定量pcr分析秋葵中aepl1、aepl2、aepl3、aepl4和aepl5表达的变化情况,aepl1、aepl2、aepl3、aepl4和aepl5的特异性引物由beacon designer 7设计,以aeact为内参基因,引物序列见表3。采用美国赛默飞旗下的real-time pcr仪,按照美国赛默飞世尔公司的dynamo flash sybr green qpcr试剂盒说明书,进行三步法扩增。进行如下程序设置:95℃预变性7min;95℃,15s,45℃退火30s,75℃延伸15s,此步骤进行39个循环。每个样品进行4次生物学重复,相对表达量采用2-δct
方法进行计算分析。
[0137]
表3荧光定量pcr
[0138][0139]
两组黄秋葵的aepl1、aepl2、aepl3、aepl4和aepl5基因表达情况如图14所示,其中*,**,***分别表示在0.05、0.01、0.001水平上差异显著。
[0140]
由图14可以看出,aepl3、aepl4和aepl5在对照组中均呈先上升后下降的趋势,但在处理组中aepl3在贮藏前期表达水平几乎不变但在12d时急剧上升,且在6d、9d和12d时显著低于对照组(p﹤0.05),aepl4基因表达水平呈先缓慢上升至9d后又急剧下降,在3d、6d和12d时显著低于对照组(p﹤0.05),aepl5在对照组中的表达模式与处理组相同,也呈先上升后下降的趋势,且在6d和9d时显著低于对照组(p﹤0.05),可见赤霉素处理对黄秋葵的aepl3、aepl4和aepl5基因表达具有明显的抑制作用。
[0141]
aepl1在对照组中呈先上升后下降再上升的表达模式,在对照组中呈逐渐上升的表达模式,在整个贮藏期间aepl1表达量均为对照组高于处理组,并在9d和12d时存在显著性(p﹤0.05),可见赤霉素处理对黄秋葵的aepl1基因表达具有明显的抑制作用。
[0142]
aepl2在对照组和处理组中的表达模式均为随着贮藏时间的延长不断上升,不同的是对照组从6d开始就急剧上升,而处理组是缓慢上升至9d后才急剧上升,整个贮藏期间aepl2表达量均为对照组高于处理组,并在6d、9d和12d时存在显著性(p﹤0.05),可见赤霉素处理对黄秋葵的aepl2基因表达具有明显的抑制作用。。
[0143]
因此,赤霉素处理在贮藏期间尤其是贮藏中后期对aepls基因具有明显的抑制作用。
[0144]
本实施例还采用了0.001%、0.003%、0.005%、0.007%、0.01%、0.03%、0.05%赤霉素水溶液分别进行试验,都能起到一定的抑制黄秋葵aepme1和aepme2基因表达的效果,因篇幅的限制,这里就不一一举例说明。
[0145]
实施例7赤霉素处理对黄秋葵纤维素酶cx相关基因表达的影响
[0146]
本实施例采用新采摘的黄秋葵(购自宁波市鄞州区联心菜市场),选择大小较为统一,个头饱满,没有黑斑及损伤的黄秋葵200根,分成2组(对照组和处理组),每组各100根。配制质量百分比含量为0.01%的赤霉素水溶液,将处理组的黄秋葵在赤霉素溶液中浸泡15min。浸泡结束后,将2组黄秋葵置于常温下自然风干,最后放置于恒温恒湿培养箱中于25
±
1℃,80%~90%相对湿度下贮藏12天。分别在处理前(0天)和处理后贮藏3、6、9、12天取样,测定对照组和处理组黄秋葵的纤维素酶cx相关基因aecx表达的变化情况。
[0147]
本实施例通过转录组测序筛选到黄秋葵的纤维素酶cx的相关基因(委托广州基迪奥生物科技有限公司进行转录组测序),包括aecx,其序列分别如seq id no.14所示。将aecx的序列在ncbi上跟木槿的纤维素酶cx相关基因序列进行比对,比对结果达到高度相似,证明aecx基因确实为黄秋葵的纤维素酶cx相关基因。
[0148]
总rna提取和cdna合成方法为:称取约0.2g果实冻样,用柱式植物rna提取试剂盒2.0(北京天恩泽基因科技有限公司)提取黄秋葵的总rna。利用核酸蛋白仪与琼脂糖电泳仪来检测提取rna的质量。cdna采用hifiscript cdna synthesis kit试剂盒(北京康为生物有限公司)反转录而成。
[0149]
本实施例通过实时定量pcr分析秋葵中aecx表达的变化情况,aecx的特异性引物由beacon designer 7设计,以aeact为内参基因,引物序列见表4。采用美国赛默飞旗下的real-time pcr仪,按照美国赛默飞世尔公司的dynamo flash sybr green qpcr试剂盒说明书,进行三步法扩增。进行如下程序设置:95℃预变性7min;95℃,15s,45℃退火30s,75℃延伸15s,此步骤进行39个循环。每个样品进行4次生物学重复,相对表达量采用2-δct
方法进行计算分析。
[0150]
表4荧光定量pcr
[0151][0152]
两组黄秋葵的aecx基因表达情况如图15所示,其中*,**,***分别表示在0.05、0.01、0.001水平上差异显著。
[0153]
由图15可知,赤霉素处理后黄秋葵贮藏期间对照组aecx基因的表达模式为随贮藏时间的增加持续上调,处理组aecx基因表达在0-3d略有下降后也随着贮藏时间的增加持续上调。整个贮藏期间对照组aecx基因的表达量都显著高于处理组(p﹤0.05)。因此,赤霉素处理能有效的抑制黄秋葵aecx基因的表达。
[0154]
本实施例还采用了0.001%、0.003%、0.005%、0.007%、0.01%、0.03%、0.05%赤霉素水溶液分别进行试验,都能起到一定的抑制黄秋葵aecx基因表达的效果,因篇幅的限制,这里就不一一举例说明。
[0155]
实施例8赤霉素处理对黄秋葵α-l-阿拉伯呋喃糖苷酶α-af相关基因表达的影响
[0156]
本实施例采用新采摘的黄秋葵(购自宁波市鄞州区联心菜市场),选择大小较为统一,个头饱满,没有黑斑及损伤的黄秋葵200根,分成2组(对照组和处理组),每组各100根。配制质量百分比含量为0.01%的赤霉素水溶液,将处理组的黄秋葵在赤霉素溶液中浸泡15min。浸泡结束后,将2组黄秋葵置于常温下自然风干,最后放置于恒温恒湿培养箱中于25
±
1℃,80%~90%相对湿度下贮藏12天。分别在处理前(0天)和处理后贮藏3、6、9、12天取样,测定对照组和处理组黄秋葵的α-l-阿拉伯呋喃糖苷酶α-af相关基因aeα-af1、aeα-af2、aeα-af3、aeα-af4和aeα-af5表达的变化情况。
[0157]
本实施例通过转录组测序筛选到黄秋葵的α-l-阿拉伯呋喃糖苷酶α-af的相关基因(委托广州基迪奥生物科技有限公司进行转录组测序),包括aeα-af1、aeα-af2、aeα-af3、aeα-af4和aeα-af5,其序列分别如seq id no.15~seq id no.19所示。将aeα-af1、aeα-af2、aeα-af3、aeα-af4和aeα-af5的序列在ncbi上跟木槿、棉花的α-l-阿拉伯呋喃糖苷酶α-af相关基因序列进行比对,结果如图16~20所示,其中aeα-af1的比对结果见图16,aeα-af2的比对结果见图17,aeα-af3的比对结果见图18,aeα-af4的比对结果见图19,aeα-af5的比对结果见图20,达到高度相似,证明aeα-af1、aeα-af2、aeα-af3、aeα-af4和aeα-af5基因确实为黄秋葵的α-l-阿拉伯呋喃糖苷酶α-af相关基因。
[0158]
总rna提取和cdna合成方法为:称取约0.2g果实冻样,用柱式植物rna提取试剂盒2.0(北京天恩泽基因科技有限公司)提取黄秋葵的总rna。利用核酸蛋白仪与琼脂糖电泳仪来检测提取rna的质量。cdna采用hifiscript cdna synthesis kit试剂盒(北京康为生物有限公司)反转录而成。
[0159]
本实施例通过实时定量pcr分析秋葵中aeα-af1、aeα-af2、aeα-af3、aeα-af4和aeα-af5表达的变化情况,aeα-af1、aeα-af2、aeα-af3、aeα-af4和aeα-af5的特异性引物由beacon designer 7设计,以aeact为内参基因,引物序列见表5。采用美国赛默飞旗下的real-time pcr仪,按照美国赛默飞世尔公司的dynamo flash sybr green qpcr试剂盒说明书,进行三步法扩增。进行如下程序设置:95℃预变性7min;95℃,15s,45℃退火30s,75℃延伸15s,此步骤进行39个循环。每个样品进行4次生物学重复,相对表达量采用2-δct
方法进行计算分析。
[0160]
表5荧光定量pcr
[0161][0162][0163]
两组黄秋葵的aeα-af1、aeα-af2、aeα-af3、aeα-af4和aeα-af5基因表达情况如图21所示,其中*,**,***分别表示在0.05、0.01、0.001水平上差异显著。
[0164]
由图21可见,对照组在贮藏期间除aeα-af2呈先上升后下降再上升再下降的变化趋势外,其余都呈先上升后下降的趋势,其中aeα-af1、aeα-af4、aeα-af5都在9d达到表达水平的最高峰后下调,aeα-af3表达量则在6d后开始逐渐下降。处理组除aeα-af4在3d时小幅下降后逐渐上升外,其余表达模式均呈先上升后下降的趋势。除6d时aeα-af2基因表达量为处理组高于对照组,在整个贮藏时间aeα-afs基因表达量均为对照组大于处理组,且在9d和12d时对照组显著高于处理组(p﹤0.05)。因aeα-af2相比于其他aeα-afs表达水平较低,所以总体上看赤霉素处理总体对aeα-afs是起抑制表达作用的。
[0165]
本实施例还采用了0.001%、0.003%、0.005%、0.007%、0.01%、0.03%、0.05%赤霉素水溶液分别进行试验,都能起到一定的抑制黄秋葵aeα-afs基因表达的效果,因篇幅的限制,这里不一一举例说明。
[0166]
实施例9赤霉素处理对黄秋葵β-半乳糖苷酶β-gal相关基因表达的影响
[0167]
本实施例采用新采摘的黄秋葵(购自宁波市鄞州区联心菜市场),选择大小较为统一,个头饱满,没有黑斑及损伤的黄秋葵200根,分成2组(对照组和处理组),每组各100根。配制质量百分比含量为0.01%的赤霉素水溶液,将处理组的黄秋葵在赤霉素溶液中浸泡15min。浸泡结束后,将2组黄秋葵置于常温下自然风干,最后放置于恒温恒湿培养箱中于25
±
1℃,80%~90%相对湿度下贮藏12天。分别在处理前(0天)和处理后贮藏3、6、9、12天取
样,测定对照组和处理组黄秋葵的β-半乳糖苷酶β-gal相关基因aeβ-gal1、aeβ-gal2、aeβ-gal3、aeβ-gal4、aeβ-gal5和aeβ-gal6表达的变化情况。
[0168]
本实施例通过转录组测序筛选到黄秋葵的β-半乳糖苷酶β-gal的相关基因(委托广州基迪奥生物科技有限公司进行转录组测序),包括aeβ-gal1、aeβ-gal2、aeβ-gal3、aeβ-gal4、aeβ-gal5和aeβ-gal6,其序列分别如seq id no.20~seq id no.25所示。将aeβ-gal1、aeβ-gal2、aeβ-gal3、aeβ-gal4、aeβ-gal5和aeβ-gal6的序列在ncbi上跟木槿、棉花、榴莲、可可的β-半乳糖苷酶β-gal相关基因序列进行比对(因篇幅有限,这里列举部分比对结果如图22~26所示,其中aeβ-gal1的比对结果见图22、aeβ-gal2的比对结果见图23、aeβ-gal3的比对结果见图24、aeβ-gal4的比对结果见图25、aeβ-gal5的比对结果见图26),比对结果证明达到高度相似,证明aeβ-gal1、aeβ-gal2、aeβ-gal3、aeβ-gal4、aeβ-gal5和aeβ-gal6基因确实为黄秋葵的β-半乳糖苷酶β-gal相关基因。
[0169]
总rna提取和cdna合成方法为:称取约0.2g果实冻样,用柱式植物rna提取试剂盒2.0(北京天恩泽基因科技有限公司)提取黄秋葵的总rna。利用核酸蛋白仪与琼脂糖电泳仪来检测提取rna的质量。cdna采用hifiscript cdna synthesis kit试剂盒(北京康为生物有限公司)反转录而成。
[0170]
本实施例通过实时定量pcr分析秋葵中aeβ-gal1、aeβ-gal2、aeβ-gal3、aeβ-gal4、aeβ-gal5和aeβ-gal6表达的变化情况,aeβ-gal1、aeβ-gal2、aeβ-gal3、aeβ-gal4、aeβ-gal5和aeβ-gal6的特异性引物由beacon designer 7设计,以aeact为内参基因,引物序列见表6。采用美国赛默飞旗下的real-time pcr仪,按照美国赛默飞世尔公司的dynamo flash sybr green qpcr试剂盒说明书,进行三步法扩增。进行如下程序设置:95℃预变性7min;95℃,15s,45℃退火30s,75℃延伸15s,此步骤进行39个循环。每个样品进行4次生物学重复,相对表达量采用2-δct
方法进行计算分析。
[0171]
表6荧光定量pcr
[0172][0173]
两组黄秋葵的aeβ-gal1、aeβ-gal2、aeβ-gal3、aeβ-gal4、aeβ-gal5和aeβ-gal6基因表达情况如图27所示,其中*,**,***分别表示在0.05、0.01、0.001水平上差异显著。
[0174]
由图27可以看出,对照组在贮藏期间都呈先上升后下降的趋势,均在9d达到表达水平的最高峰后下调。与对照组相比,aeβ-gal1基因和aeβ-gal2基因整个贮藏期间在对照组中几乎不表达,所以在6d、9d和12d时对照组中aeβ-gal1和aeβ-gal2表达量显著高于处理组(p﹤0.05)。aeβ-gal3、aeβ-gal4和aeβ-gal6在处理组中呈上升趋势,且表达水平在9d和12d显著低于对照组(p﹤0.05)。aeβ-gal5基因在处理组中呈先急剧上升后缓慢下降的趋势,
并在9d时显著低于对照组的表达(p﹤0.05)。从整体上看,在黄秋葵整个贮藏期间尤其是贮藏后期,aeβ-gals在对照组中的表达高于处理组。因此,赤霉素处理在贮藏期间尤其是贮藏后期对aeβ-gals基因有抑制作用。
[0175]
本实施例还采用了0.001%、0.003%、0.005%、0.007%、0.01%、0.03%、0.05%赤霉素水溶液分别进行试验,都能起到一定的抑制黄秋葵aeα-afs基因表达的效果,因篇幅的限制,这里不一一举例说明。
[0176]
本发明的应用并不局限于此。如根据其在环境保护方面的应用范围均可做扩展。任何本领域技术人员,在不脱离本发明的精神和范围内,均可作各种更动与修改,因此本发明的保护范围应当以权利要求所限定的范围为准。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1