针对α-突触核蛋白的抗体及其用途的制作方法
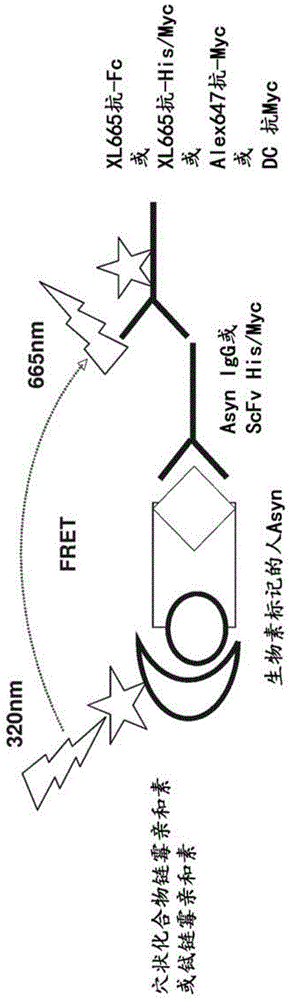
此申请要求2016年6月2日提交的美国临时申请号62/344,746的优先权权益。上述申请通过提述以其整体并入本文。序列表本申请包含序列表,其已经以ascii格式电子提交,并通过提述以其整体并入本文。将所述ascii版本(创建于2017年5月30日)命名为1848081-0002-091-wo1_sl.txt,且大小为48,099个字节。背景本发明涉及α-突触核蛋白抗体和它们在预防或治疗疾病,特别是α-突触核蛋白病(alpha-synucleinopathy),且更特别地是帕金森氏病(pd)中的用途。α-突触核蛋白病(也称为路易体疾病(lewybodydisease,lbd))是神经退行性疾病家族,它们都在其核心具有α-突触核蛋白作为关键的病理学标志(jellinger,movdisord(2003),18suppl6:s2-12;和spillantini和goedert,annnyacadsci(2000),920:16-27;其中二者通过提述并入本文)。α-突触核蛋白病包括是帕金森氏病(pd)、路易体痴呆(dlb)和多系统萎缩症(msa)。pd是一种缓慢进展的与年龄相关的活动病症,其影响1%以上的65岁以上的人。pd是阿尔茨海默病后第二常见的神经退行性病况。α-突触核蛋白病的定义性标志病理是路易体和路易神经突,其是在死后的组织病理学检查中揭示的在脑的神经元内部发现的聚集蛋白质的不溶性内含物。非运动脑区(如基底前脑、中脑桥(mesopontine)系统、杏仁核、新皮质、迷走神经的背侧运动核、嗅球、蓝斑和脑干)中路易病理和神经元丢失的存在可引起认知缺陷和痴呆、嗅觉减退、睡眠障碍(包括快速眼动睡眠行为病症(rbd)、情绪病症(包括抑郁和焦虑)、自主神经功能紊乱(包括心血管和胃肠道问题,如便秘、疲劳和嗜睡)。这些非运动症状中的一些看起来表征疾病的前运动或前驱阶段(kalia等lancet(2015),386(9996):896-912;其通过提述并入本文)。运动脑区中路易病理和神经元丢失(包括最显著地黑质中多巴胺能神经元的死亡)的存在可引起静息性震颤、僵硬、运动迟缓和姿势不稳定(spillantini和goedert,annnyacadsci(2000),920:16-27;其通过提述并入本文)。α-突触核蛋白(alpha-synuclein)(也称为“α-突触核蛋白(α-synuclein)”或α-syn)蛋白质是路易体和路易神经突的主要结构组分。α-突触核蛋白是由140个氨基酸组成的小酸性蛋白质(14kda)。人天然野生型α-突触核蛋白具有如根据uniprotkb登录号p37840所述的氨基酸序列seqidno:1。除非上下文另有明文,否则提及的α-突触核蛋白或其片段包括上述指示的天然的人野生型氨基酸序列和其人等位基因变体,特别是与路易体疾病相关的那些(例如e46k、a30p、h50q、g51d和a53t,其中第一个字母指示seqidno:1中的氨基酸,数字是seqidno:1中的密码子位置,以及第二个字母是等位基因变体中的氨基酸)。这些变体可任选地单独存在或以任何组合存在。诱导的突变e83q、a90v、a76t(其增强α-突触核蛋白聚集)也可单独存在或彼此组合存在和/或与人等位基因变体e46k、a30p、h50q、g51d和a53t组合存在。在结构水平上,α-突触核蛋白包含三个独特的区域:具有脂质和膜结合特性的两亲性n-末端α-螺旋域(残基1-60),编码斑块的非淀粉样蛋白-β组分(nac)的中心疏水性淀粉样蛋白结合域(残基61-95),和酸性富含脯氨酸的c-末端尾(残基96-140)。认为所述nac域的残基71-82是通过使蛋白质从无规卷曲结构转变为β-片层结构的α-突触核蛋白聚集/原纤维形成性质的关键(bisaglia等fasebj(2009),23(2):329-40;其通过提述并入本文)。虽然c末端域没有明显的二级结构,但它在残基ser129处包含关键的磷酸化位点和多个在胞质α-突触核蛋白内含物中硝化的酪氨酸残基。也存在α-突触核蛋白的n-末端和c-末端截短形式。蛋白质的翻译后修饰影响α-突触核蛋白聚集和毒性(oueslati等progbrainres(2010),183:115-45;其通过提述并入本文)。α-突触核蛋白在中枢神经系统(cns)/脑中是丰富的,其中在细胞内神经元和神经胶质细胞中以及细胞外的脑脊髓液(csf)(mollenhauer等jneuraltransm(2012),119(7):739-46;其通过提述并入本文)和浸浴且包围脑细胞的组织间液(isf)(emmanouilidou等plosone(2011),6(7):e22225;其通过提述并入本文)中发现它。α-突触核蛋白是主要在新皮质、海马、黑质、丘脑和小脑的神经元中表达的突触蛋白(iwai等neuron(1995),14:467-475;其通过提述并入本文)。在生理条件下,它位于神经元突触末端,并且在获得相关的突触重排期间在突触前端特异性上调(fortin等jneurosci(2005),25:10913-10921;其通过提述并入本文)。体外研究已经表明α-突触核蛋白单体可形成聚集过程的起始点。所述单体可聚集成各种小的寡聚物种类(然后通过β-片层相互作用使其稳定),继而形成可聚合成不可溶的纤维结构(其令人联想到路易体中鉴定的那些)的原纤维(cremades等cell(2012),149(5):1048-59;其通过提述并入本文)。在生理条件下,异常α-突触核蛋白聚集可为α-突触核蛋白病中可见的病理变化的关键(lashuel等nature(2002),418:291;和tsigelny等febsjournal(2007),274:1862-1877;其中二者通过提述并入本文)。体外和体内研究已经表明α-突触核蛋白神经毒性作用似乎是由小的可溶性低聚构象异构体或原纤维引起的(winner等procnatlacadsciusa(2011),108(10):4194-9;和danzer等jneurosci(2007),27(34):9220-32;其中二者通过提述并入本文)。虽然α-突触核蛋白的纤维状聚集体是pd的特征,但α-突触核蛋白的寡聚形式是有毒的种类(danzer等jneurosci(2007),27(34):9220-32;lashuel等nature(2002),418:291;和winner等procnatlacadsciusa(2011),108:4194-4199;其中每个通过提述并入本文)。α-突触核蛋白寡聚体可被释放到细胞外环境中并以“传播(propagation)”机制由相邻细胞摄取(angot和brundin,parkinsonismrelatdisord(2009),15suppl3:s143-147;desplats等procnatlacadsciusa(2009),106:13010-13015;和lee等jbiolchem(2010),285:9262-9272;其中每个通过提述并入本文)。α-突触核蛋白的聚集体通过朊病毒样扩散机制传播错误折叠(lee等natrevneurol(2010),6:702-706;luk等jexpmed(2012),209(5):975-86;和luk等science(2012),338(6109):949-53;其中每个通过提述并入本文)。因此,α-突触核蛋白可通过寡聚体毒性或传播和朊病毒样扩散诱导神经退变。现已充分地确定并接受包括神经元的细胞可在正常条件下以及在细胞应激条件下分泌各种形式的α-突触核蛋白(单体、寡聚体、聚集体),在细胞应激条件下提高α-突触核蛋白的单体和聚集形式的分泌,并且通过这种使α-突触核蛋白释放到细胞外环境中,α-突触核蛋白的病理可传递形式可在神经元之间传播(recasens和dehay,frontneuroanat(2014),8:159;其通过提述并入本文)。α-突触核蛋白在pd中的作用可超出对脆弱的神经细胞的即时损伤。像大多数神经退行性疾病一样,也观察到促炎性细胞反应(lee等jbiolchem(2010),285:9262-9272;其通过提述并入本文)。循环的α-突触核蛋白和/或活化的星形胶质细胞可激活小胶质细胞,这导致反应性氧种类、一氧化氮和细胞因子产生的增加,并且进一步加重神经退变(lee等jbiolchem(2010),285:9262-9272;其通过提述并入本文)。各种不同的实验模型已经证明了培养的细胞中α-突触核蛋白的细胞至细胞的传递,或α-突触核蛋白病理的体内扩散和传播。在将胚胎中脑神经元移植物治疗性移植到pd患者的纹状体中超过10年后,在胚胎中脑神经元移植物中观察到路易体病理。具体地,移植的神经元含有许多染色对于α-突触核蛋白呈阳性的路易体样内含物,其指示α-突触核蛋白病理的宿主至移植物的传递已经发生(li等natmed(2008),14(5):501-3;和kordower等natmed(2008),14(5):504-6;其中二者通过提述并入本文)。进一步地,可将预形成的重组α-突触核蛋白原纤维和α-突触核蛋白寡聚体由培养的细胞和神经元内化,并且已经证明了α-突触核蛋白从供体直接转移至已形成与路易病理相似的α-突触核蛋白内含物的受体细胞(danzer等jneurosci(2007),27(34):9220-32;volpicelli-daley等neuron(2011),72(1):57-71;和luk等procnatlacadsciusa(2009),106(47):20051-6;其中每个通过提述并入本文)。将预形成的合成α-突触核蛋白原纤维和含有自年老α-突触核蛋白转基因小鼠的脑中提取的物质的路易体样α-突触核蛋白注射到无症状受体小鼠的脑中促进该受体动物的宿主神经元中的路易体样病理的形成,伴随着神经退变和神经学缺陷(luk等jexpmed(2012),209(5):975-86;和luk等science(2012),338(6109):949-53;其中二者通过提述并入本文)。自接种到猕猴和小鼠的黑质或纹状体中的pd脑中分离的含有α-突触核蛋白的路易体提取物由宿主细胞快速摄取(24小时内),随后纹状体多巴胺能末端缓慢缺失,多于一年后细胞死亡明显(recasens等annneurol(2014),75(3):351-62;其通过提述并入本文)。相似地,将源自具有突触核蛋白病dlb或msa的患者的脑匀浆物接种小鼠触发该宿主小鼠中的α-突触核蛋白路易体样病理(watts等procnatlacadsciusa(2013),110(48):19555-60;和masuda-suzukake等brain(2013),136(pt4):1128-38;其中二者通过提述并入本文)。最后,已经在小鼠中证明了单体和寡聚体α-突触核蛋白二者从嗅球至相互连接的脑结构的转移和传递(rey等actaneuropathol(2013),126(4):555-73;其通过提述并入本文)。已经在多个临床前α-突触核蛋白病小鼠模型中测试了靶向α-突触核蛋白的抗体的被动免疫治疗方法(lawand等expertopinthertargets(2015):1-10;其通过提述并入本文)。具体而言,一项使用直接针对α-突触核蛋白的单克隆抗体(9e4)的研究已经显示α-突触核蛋白聚集体和病理的体内清除、行为运动改善和神经保护作用(wo2014/058924;其通过提述并入本文)。使用所述94e单克隆抗体被动免疫发展为pd/dlb实验模型的α-突触核蛋白转基因小鼠的进一步研究已经显示所述抗体清除α-突触核蛋白病理、降低突触和轴突缺陷、消除纹状体酪氨酸羟化酶纤维的损失,并显著减少记忆缺陷和运动功能损伤(games等jneurosci(2014),34(28):9441-54;bae等jneurosci(2012),32(39):13454-69;和masliah等plosone(2011),6(4):e19338;其中每个通过提述并入本文)。进一步地,已经证明在纹状体内注射合成的α-突触核蛋白预形成的原纤维(pffs)的野生型小鼠中抗α-突触核蛋白单克隆抗体的被动施用导致路易病理的稳固减少,防止黑质中多巴胺神经元的缺失,并且显著改善pffs处理后的小鼠模型中显现出的运动损伤(tran等cellrep(2014),7(6):2054-65;其通过引用并入本文)。此外,与用大分子疗法(如抗体)治疗cns的病症相关的主要挑战之一是将这些药物送入受影响的组织中。大分子进入脑和脊髓的通路很大程度上受血脑屏障(bbb)的限制。bbb保护和调节脑的动态平衡,并且防止分子自由进入脑的大部分区域,由此限制许多脑部疾病的治疗。经由调节穿过脑内皮细胞通路的一系列特异性转运蛋白和受体,实现必需分子(如营养素、生长因子和激素)的转运。因此,将生物制品和其他药物递送至脑代表着一项重大的挑战。此外,似乎存在快速从脑中去除抗体的转运机制,其可能是由于fc与促进促炎反应的效应配体结合而防止炎症反应。在过去的十年中,穿过bbb的抗体转运的报道已经出现,其中与转运蛋白分子的胞外域的结合促进受体抗体复合物穿过内皮细胞层的转胞吞作用。bbb主要由脑毛细血管内皮细胞组成,所述细胞具有特殊的特征(如紧密连接)以限制将分子转运入脑(reese等1967,j.cellbiol.34:207-217;brightman等1969,j.cellbiol.40:648-677;rubin等1999,ann.rev.neurosci.22:11-28),尽管其他细胞类型(如周皮细胞、星形胶质细胞和神经元细胞)也在bbb功能中起重要作用。通常,少于0.1%的外周给药的抗体到达脑部(boado等2010,mol.pharm.7:237-244;pepinsky等2011,nat.neurosci.8:745-751)。bbb作为物理、代谢和免疫的屏障起作用(gaillard等2003,microvasc.res.65:24-31)。穿过bbb的抗体转运可通过触发脑内皮细胞上受体介导的转胞吞作用来增强。通过此过程,抗原在内皮细胞的腔侧上的结合可诱导穿过细胞的抗体内化和穿梭,然后其随后释放到组织中。针对pd的现有药物疗法主要集中在治疗所述疾病的运动相关的症状。目前,没有能治疗或预防α-突触核蛋白病的市售或可用的疗法。因此,本领域中需要用于治疗α-突触核蛋白病(特别是在人体内)的疗法。技术实现要素:本发明涉及针对人α-突触核蛋白的分离的抗体。本发明提供具有aslo0452ngl-3抗体的一种或多种功能特性的抗体或其抗原结合片段。例如:本发明提供特异性结合人α-突触核蛋白的c末端区的抗体或其抗原结合片段。本发明提供特异性结合包含人α-突触核蛋白(例如seqidno:1)的约氨基酸102至约氨基酸130的区域的抗体或其抗原结合片段。在一些实施方案中,所述抗体或其抗原结合片段特异性结合至包含人α-突触核蛋白(例如seqidno:1)的约氨基酸120至约氨基酸130的区域。在一些实施方案中,所述抗体或其抗原结合片段结合至与所述9e4抗体所结合的表位不同的表位。本发明提供结合人α-突触核蛋白但不结合人β-突触核蛋白或人γ-突触核蛋白的抗体或其抗原结合片段。本发明提供结合至人、大鼠或食蟹猴α-突触核蛋白的抗体或其抗原结合片段。本发明提供以高亲和力结合至人α-突触核蛋白的抗体或其抗原结合片段。在一个实施方案中,本发明的抗体或其抗原结合片段以如例如使用octet分析测量的小于500皮摩尔(pm)、小于400pm、小于300pm、小于200pm、小于150pm、小于120pm、小于110pm或106pm或更小的kd结合至α-突触核蛋白。在一个实施方案中,本发明的抗体或其抗原结合片段以如例如使用kinexa分析测量的小于400皮摩尔(pm)、小于300pm、小于250pm、小于200pm、小于150pm、小于120pm、小于110pm、小于100pm、小于80pm或74pm或更小的kd结合至α-突触核蛋白。本发明提供结合至天然内源性的人α-突触核蛋白的抗体或其抗原结合片段。本发明提供结合至人α-突触核蛋白的单体形式的抗体或其抗原结合片段。本发明提供结合至人α-突触核蛋白的聚集体的抗体或其抗原结合片段。本发明提供结合α-突触核蛋白的疾病相关的病理形式的抗体或其抗原结合片段。本发明提供减少脑组织间液中的α-突触核蛋白水平的本发明的抗体或其抗原结合片段。特别地,根据本发明的抗体或其抗原结合片段减少脑组织间液中游离未结合的α-突触核蛋白水平。本发明提供减少脑脊髓液中的α-突触核蛋白水平的本发明的抗体或其抗原结合片段。特别地,根据本发明的抗体或其抗原结合片段减少脑脊髓液中游离未结合的α-突触核蛋白水平。如本文所用,术语“游离未结合的α-突触核蛋白”指未结合至根据本发明的抗体或其抗原结合片段的α-突触核蛋白。所述游离未结合的α-突触核蛋白可应用于α-突触核蛋白的单体或寡聚体的形式或者聚集的形式。通常,这些术语应用于α-突触核蛋白的任何病理形式。本发明提供减少体内α-突触核蛋白扩散的抗体或其抗原结合片段。在一个实施方案中,本发明的抗体或其抗原结合片段与抗体aslo0452ngl-3竞争结合至人α-突触核蛋白。在一个实施方案中,本发明的抗体或其抗原结合片段结合至与aslo0452ngl-3相同的人α-突触核蛋白上的表位。在一个实施方案中,本发明的抗体或其抗原结合片段源自包含seqidno:2的氨基酸的可变重链区(vh)和seqidno:3的氨基酸的可变轻链区(vl)的抗体asyn0087,如本文所公开的。在特定的实施方案中,本发明的抗体或其抗原结合片段源自抗体asyn0087,其中所述抗体或抗原结合片段具有小于500mm的kd并且结合与抗体asyn0087、aslo0452ngl-3和aslo0543中任一个相同的表位,如本文所述。如本文所用,“h-cdr”代表着重链区上的互补决定区(cdr),以及“l-cdr”代表着抗体或其抗原结合片段的轻链区上的互补决定区(cdr)。在一个实施方案中,根据本发明的抗体或其抗原结合片段包含至少一个cdr,所述cdr选自下列:(i)seqidno:5的h-cdr1,(ii)seqidno:6的h-cdr2,(iii)seqidno:7的h-cdr3,(iv)seqidno:9的l-cdr1,(v)seqidno:10的l-cdr2,(vi)seqidno:11的l-cdr3。在进一步的实施方案中,根据本发明的抗体或其抗原结合片段的重链的cdr3是抗体aslo0452ngl-3的重链的seqidno:16的cdr3;和/或根据本发明的抗体或其抗原结合片段的轻链的cdr3是抗体aslo0452ngl-3的轻链的seqidno:21的cdr3。在一个实施方案中,本发明的抗体或其抗原结合片段具有选自抗体aslo0452ngl-3的cdr的cdr中的至少一个、至少两个、至少三个、至少四个、至少五个或全部,即选自seqidno:5、seqidno:15、seqidno:16、seqidno:20、seqidno:10、seqidno:21中任一个的至少一个cdr。在一个实施方案中,本发明的抗体或其抗原结合片段的重链的cdr3是抗体aslo0452ngl-3的重链的cdr3;和/或本发明的抗体或其抗原结合片段的轻链的cdr3是抗体aslo0452ngl-3的轻链的cdr3。在一个实施方案中,本发明的抗体或其抗原结合片段的重链的cdr3是抗体aslo0452ngl-3的重链的cdr3。在一个实施方案中,本发明的抗体或其抗原结合片段的轻链的cdr3是抗体aslo0452ngl-3的轻链的cdr3。在一个实施方案中,本发明的抗体或其抗原结合片段的重链的cdr3是抗体aslo0452ngl-3的重链的cdr3,且本发明的抗体或其抗原结合片段的轻链的cdr3是抗体aslo0452ngl-3的轻链的cdr3。本发明提供具有抗体aslo0452ngl-3的六个cdr的抗体或其抗原结合片段。因此,在一个实施方案中,根据本发明的抗体或其抗原结合片段包含:a)具有下列序列的三个重链cdr:(i)seqidno:5的h-cdr1,(ii)seqidno:15的h-cdr2;和(iii)seqidno:16的h-cdr3,和b)具有下列序列的三个轻链cdr:(i)seqidno:20的l-cdr1,(ii)seqidno:10的l-cdr2,和(iii)seqidno:21的l-cdr3。本发明提供本发明的抗体或其抗原结合片段,其包含与由seqidno:13所定义的核苷酸序列具有至少80%、85%、90%、95%、96%、97%、98%、99%或100%同一性的可变重链,以及与由seqidno:18所定义的核苷酸序列具有至少80%、85%、90%、95%、96%、97%、98%、99%或100%同一性的可变轻链。本发明提供本发明的抗体或其抗原结合片段,其包含与由seqidno:14所定义的核苷酸序列具有至少80%、85%、90%、95%、96%、97%、98%、99%或100%同一性的可变重链,以及与由seqidno:19所定义的核苷酸序列具有至少80%、85%、90%、95%、96%、97%、98%、99%或100%同一性的可变轻链。本发明提供本发明的抗体或其抗原结合片段,其包含具有seqidno:14的氨基酸序列的可变重链和具有seqidno:19的氨基酸序列的可变轻链。在特定的实施方案中,根据本发明的抗体或其抗原结合片段包含具有由seqidno:4所定义的序列的可变重链和具有由seqidno:8所定义的序列的可变轻链。在进一步的特定实施方案中,根据本发明的抗体或其抗原结合片段包含具有由seqidno:4所定义的序列的可变重链以及具有由seqidno:8所定义的序列的可变轻链,并且以小于500pm的kd结合至人α-突触核蛋白,并且结合与asyn0087、aslo0452ngl-3或aslo0543相同的表位。在另一个实施方案中,根据本发明的抗体或其抗原结合片段包含与由seqidno:14所定义的序列具有至少80%、85%、90%、95%、96%、97%、98%、99%或100%同一性的可变重链和与由seqidno:19所定义的序列具有至少80%、85%、90%、95%、96%、97%、98%、99%或100%同一性的可变轻链。在特定的实施方案中,根据本发明的抗体或其抗原结合片段包含具有与由seqidno:14所定义的序列的可变重链和具有由seqidno:19所定义的序列的可变轻链。在进一步的实施方案中,根据本发明的抗体或其抗原结合片段包含与由seqidno:14所定义的序列具有至少80%、85%、90%、95%、96%、97%、98%、99%或100%同一性的可变重链和与由seqidno:19多定义的序列具有至少80%、85%、90%、95%、96%、97%、98%、99%或100%同一性的可变轻链,并且进一步包含:a)具有下列序列的三个重链cdr:(i)seqidno:5的h-cdr1,(ii)seqidno:15的h-cdr2;和(iii)seqidno:16的h-cdr3,和b)具有下列序列的三个轻链cdr:(i)seqidno:20的l-cdr1,(ii)seqidno:10的l-cdr2;和(iii)seqidno:21的l-cdr3。本发明还提供包含可变重链和可变轻链的抗体或其抗原结合片段,所述可变重链具有由seqidno:13所定义的核苷酸序列,所述轻链具有由seqidno:18所定义的核苷酸序列。本发明提供包含可变重链和可变轻链的抗体或其抗原结合片段,所述可变重链具有由seqidno:14所定义的氨基酸序列,所述轻链具有由seqidno:19所定义的氨基酸序列。还提供本发明的包含重链和轻链的抗体或其抗原结合片段,其中所述重链具有由seqidno:12所定义的氨基酸序列,所述轻链具有由seqidno:17所定义的氨基酸序列。在另一个实施方案中,根据本发明的抗体或其抗原结合片段包含与由seqidno:24所定义的序列具有至少80%、85%、90%、95%、96%、97%、98%、99%或100%同一性的可变重链和与由seqidno:30所定义的序列具有至少80%、85%、90%、95%、96%、97%、98%、99%或100%同一性的可变轻链。在特定的实施方案中,根据本发明的抗体或其抗原结合片段包含可变重链和可变轻链,所述可变重链具有由seqidno:24所定义的序列,所述轻链具有由seqidno:30所定义的序列。在进一步的实施方案中,根据本发明的抗体或其抗原结合片段包含与由seqidno:24所定义的序列具有至少80%、85%、90%、95%、96%、97%、98%、99%或100%同一性的可变重链和与由seqidno:30所定义的序列具有至少80%、85%、90%、95%、96%、97%、98%、99%或100%同一性的可变轻链,并且进一步包含:c)具有下列序列的三个重链cdr:(i)seqidno:25的h-cdr1,(ii)seqidno:26的h-cdr2;和(iii)seqidno:27的h-cdr3,和d)具有下列序列的三个轻链cdr:(i)seqidno:31的l-cdr1,(ii)seqidno:32的l-cdr2;和(iii)seqidno:33的l-cdr3。本发明还提供包含可变重链和可变轻链的抗体或其抗原结合片段,所述可变重链具有由seqidno:24所定义的核苷酸序列且所述轻链具有由seqidno:30所定义的核苷酸序列。本发明提供包含可变重链和可变轻链的抗体或其抗原结合片段,所述可变重链具有由seqidno:24所定义的氨基酸序列且所述轻链具有由seqidno:30所定义的氨基酸序列。还提供本发明的包含重链和轻链的抗体或其抗原结合片段,所述重链具有由seqidno:22所定义的氨基酸序列且所述轻链具有由seqidno:28所定义的氨基酸序列。在一个实施方案中,如上任意处所定义的,本发明的抗体或其抗原结合片段是iga、igd、ige、igm、igg如igg1、igg2、igg3或igg4抗体或其抗原结合片段。在另一个实施方案中,本发明的抗体或其抗原结合片段具有修饰的fc区。合适的修饰是本领域技术人员公知的,并且尤其可包括修饰以增加或减少半衰期、除去、降低或增强效应器功能、提供具有用于缀合的游离巯基的取代的半胱氨酸。此类修饰的实例是增加半衰期的yte和/或降低效应器功能的tm。在一些实施方案中,本文公开的任何抗体或其抗原结合片段包含所述抗体的fc区中的突变m252y/s254t/t256e(yte)(dall’acqua等,2006,j.biol.chem,281:23514-23524)。在一些实施方案中,本文公开的任何抗体或其抗原结合片段包含对应于oganesyan等actacrystallogrdbiolcrystallogr,(2008)64:700–704中公开的l234f/l235e/p331s突变的fc区中的三突变(本文中缩写为“tm”)。在一个实施方案中,本发明的抗体或其抗原结合片段可为igg1tm抗体或其抗原结合片段。在另一个实施方案中,本发明的抗体或其抗原结合片段可包含具有yte突变的fc区。在另一个实施方案中,本发明的抗体或其抗原结合片段可与血脑屏障(bbb)转运蛋白部分偶联,其中所述bbb转运蛋白部分能够转运所述抗体或其抗原结合片段穿过bbb。在实施方案中,bbb转运蛋白部分可为抗体。在实施方案中,bbb抗体可与抗α-突触核蛋白抗体或其抗原结合片段形成多特异性构建体。bbb转运蛋白部分可包含免疫球蛋白可变重链互补决定区-1(vh-cdr1)、免疫球蛋白可变重链互补决定区-2(vh-cdr2)、免疫球蛋白可变重链互补决定区-3(vh-cdr3)、免疫球蛋白可变轻链互补决定区-1(vl-cdr1)、免疫球蛋白可变轻链互补决定区-2(vl-cdr2)和免疫球蛋白可变轻链互补决定区-3(vl-cdr3);其中所述vh-cdr1包含seqidno.40或49,vh-cdr2包含seqidno.41或50,vh-cdr3包含seqidno.42或51,vl-cdr1包含seqidno.36、44或53,vl-cdr2包含seqidno.37、45或54且vl-cdr3包含seqidno.38、46或55。在一些实施方案中,所述转运蛋白部分包含免疫球蛋白可变重链(vh)区,其包含seqidno:47或seqidno:39。在一些实施方案中,所述转运蛋白部分包含免疫球蛋白轻链(vl)区,其包含seqidno:43。此外,所述转运蛋白部分可选自完整抗体、fv片段、fab片段、fab'片段、f(ab')2片段、二硫键连接的(dsfv)片段、单链fv(scfv)片段、sc(fv)2、双功能抗体(diabody)、三功能抗体(triabody)、四功能抗体(tetrabody)、微抗体(minibody)和单链抗体。在特定的实施方案中,所述转运蛋白部分包含scfv片段,其包含经由接头融合在一起的vh域和vl域。在一些情况下,所述接头可为(gly4ser)n(seqidno:56),其中n是选自由1、2、3、4、5、6、7、8、9和10组成的组的正整数。在一些实施方案中,如本文所述,本发明的任何转运蛋白分子可与本发明的任何α-突触核蛋白结合分子组合以提供本发明的多特异性结合分子。本发明提供如上任意处所定义的本发明的抗体或其抗原结合片段,其用作药物。本发明还提供如上任意处所定义的本发明的抗体或其抗原结合片段,其用于预防或治疗α-突触核蛋白病。在一个实施方案中,所述α-突触核蛋白病选自帕金森氏病(pd)、路易体痴呆(dlb)和多系统萎缩症(msa)。在一个实施方案中,所述α-突触核蛋白病是帕金森氏病(pd)。本发明提供治疗和预防患者中的疾病(特别是与中枢神经系统相关的疾病)的方法,所述方法包括向所述患者施用如上任意处所定义的本发明的抗体或其抗原结合片段。在一个实施方案中,所述疾病是α-突触核蛋白病。在一个实施方案中,所述α-突触核蛋白病选自帕金森氏病(pd)、路易体痴呆(dlb)和多系统萎缩症(msa)。在一个实施方案中,所述α-突触核蛋白病是帕金森氏病(pd)。本发明提供药物组合物,其包含如上任意处所定义的本发明的抗体或其抗原结合片段和药学上可接受的赋形剂。短语“药学上可接受的赋形剂”包括与药物施用相兼容的任何和所有溶剂、分散介质、涂层、抗细菌剂和抗真菌剂、等渗剂和吸收延迟剂等。此类介质和试剂用于药物活性物质的用途在本领域中是公知的。所述组合物还可包含提供补充的、附加的或增强的治疗功能的其他活性化合物。所述药物组合物还可与施用说明书一起包括在容器、包装或分配器中。将本发明的药物组合物配置成以与其预期的施用途径相兼容。完成所述施用的方法是本领域普通技术人员所知的。所述施用可为(例如)静脉内的、腹膜内的、肌肉内的、腔内的、皮下的或经皮的。本发明提供编码如上任意处所定义的本发明的抗体或其抗原结合片段的分离的核酸分子。在特定的实施方案中,本发明提供包含seqidno:13和/或seqidno:18的分离的核酸分子。在另一个特定的实施方案中,本发明提供包含seqidno:23和/或seqidno:29的分离的核酸分子。一旦提供此信息,本领域技术人员可容易地获得编码本公开的抗体或其抗原结合片段的核酸分子。所述核酸可包含dna或rna,并且可完全地或部分地合成或重组。除非上下文另有需要,否则提及的核苷酸序列包含具有特定序列的dna分子,且包含具有特定序列的rna分子(其中u取代t)。本发明的核酸分子包含本文所公开的cdr、vh域和/或vl域的编码序列。本公开还提供质粒、载体、噬菌粒、转录或表达盒的形式的构建体,其包含至少一个编码如上任意处所定义的本发明的抗体或其抗原结合片段的核酸分子(特别是编码本文公开的cdr、vh域和/或vl域)。本公开进一步提供包含一个或多个如上的构建体的宿主细胞。还提供编码本文公开的任何一个或多个cdr(h-cdr1、h-cdr2、h-cdr3、l-cdr1、l-cdr2或l-cdr3)、vh或vl域的核酸,以及制备所述编码的产物的方法。所述方法包含自所述编码核酸表达编码的产物。可通过在适当的条件下培养包含所述核酸的重组宿主细胞来实现表达。通过表达生产后,可使用任何合适的技术分离和/或纯化vh或vl域或特异性结合成员,然后视情况使用。抗原结合片段、vh和/或vl域以及编码的核酸分子和载体可从它们的天然环境中以基本上纯的或同源的形式分离和/或纯化,或者在核酸的情况下,除了编码具有所需功能的多肽的序列外,没有或者基本上没有原始核酸或基因。在各种不同宿主细胞中克隆和表达多肽的系统是本领域中公知的。对于适合产生抗体的细胞,参见geneexpressionsystems,academicpress,eds.fernandez等,1999。简言之,适合的宿主细胞包括细菌、植物细胞、哺乳动物细胞和酵母和杆状病毒系统。本领域中可用于表达异源多肽的哺乳动物细胞系包括中国仓鼠卵巢细胞、hela细胞、幼仓鼠肾细胞、ns0小鼠骨髓瘤细胞和许多其它细胞。常见的细菌宿主是大肠杆菌(e.coli)。与本发明兼容的蛋白表达系统可用于产生本公开的抗体。适当的表达系统包括geneexpressionsystems,academicpress编fernandez等,1999中所述的转基因动物。可选择或构建适当的载体,使得它们包含合适的调控序列,所述调控序列包括启动子序列、终止子序列、多聚腺苷酸化序列、增强子序列、标记基因和其他适当的序列。载体可为质粒或病毒,例如噬菌体或噬菌粒,其视情况而定。对于进一步细节参见(例如)sambrook等,molecularcloning:alaboratorymanual,第2版,coldspringharborlaboratorypress,1989。用于操作核酸的许多已知技术和方案(例如制备核酸构建体,突变发生、测序、将dna导入细胞中和基因表达以及蛋白质分析)详细描述于currentprotocolsinmolecularbiology,第2版,ausubel等编,johnwiley&sons,1992。本公开的进一步的方面提供包含本文所公开的核酸的宿主细胞,特别是包含编码如上任意处所定义的本发明的抗体或其抗原结合片段的核酸分子的载体。又进一步的方面提供包含将此核酸导入宿主细胞的方法。所述导入可采用任何可用的技术。对于真核细胞,适当的技术可包括磷酸钙转染、deae葡聚糖、电穿孔、脂质体介导的转染以及使用逆转录病毒或其他病毒(如牛痘,或者对于昆虫细胞的杆状病毒)的转导。对于细菌细胞,适当的技术可包括氯化钙转化、电穿孔和使用噬菌体的转染。将核酸导入细胞中后可引起或允许例如通过在用于表达基因的条件下培养宿主细胞而从所述核酸的表达。附图和序列表的简要说明现在将会参考附图和序列表更详细地描述本发明,其中显示了:关键seqidno:附图说明:表1:在两个亲和力测量平台上进行的针对人α-syn的关键抗α-突触核蛋白抗体的亲和力测定。图1:测定的示意图图2:比较asyn0087、aslo0452ngl-3和aslo0543重链可变区(vh)(分别为seqidno:2、14和2)和轻链可变区(vl)(分别为seqidno:3、19和30)的氨基酸序列。加下划线的氨基酸对应于cdr。图3a-3d:aslo0452ngl-3的核苷酸和氨基酸序列。图3a和3b分别显示aslo0452ngl-3的可变重链和可变轻链的核苷酸和氨基酸序列。图3a按照出现的顺序分别公开seqidno:13和14。图3b按照出现的顺序分别公开seqidno:18和19。图3c和3d显示这些序列与最接近的人种系序列的比对。图3c显示aslo0452ngl-3可变重链域氨基酸序列(seqidno:14)与种系ighv3-23(seqidno:58)和jh6序列(seqidno:59)的比对。图3d显示aslo0452ngl-3可变轻链域氨基酸序列(seqidno:19)与种系iglv5-45(seqidno:60)和jl2(seqidno:61)(3个序列)的比对。互补决定区(cdr)加下划线并标记。来自所述种系的差异以粗体并突出显示。轻链框架区中的所有非-vernier残基是人种系氨基酸。未改变vernier残基(*)以匹配所述种系氨基酸。图4a和4b:使用一组α-syn截短物的表位结合先导分离克隆。用一系列商业可得的α-syn截短物包覆elisa孔,所述α-syn截短物代表所述蛋白质的多个定义的区:1-140:全长α-syn,1-60:仅n末端区,61-140:斑块的非淀粉样蛋白组分(nac)加c末端区,1-95:n末端和nac区,96-140:仅c末端区,δnac:缺失nac区,ncap:是缺失氨基酸103-129的α-syn的可替换剪接形式(rpeptide)。用于检测的第一抗体是asyn087(图4a);和aslo0452ngl-3(黑条)、aslo0543(浅灰条)和nip228同种型匹配的对照(深灰条)(图4b)。用抗人iggeu3+二抗(图4a)或抗人igg-hrp二抗(图4b)检测结合。图5:使用delfia表位竞争测定的相对于突触核蛋白家族成员的aslo0452ngl-3和aslo0543对α-syn的特异性。使用表位竞争htrf测定,通过滴定未标记的α-syn、β-syn和γ-syn来确定亲和力优化的aslo0452ngl-3和aslo0543克隆对α-syn的特异性。由此确定ic50值。图6:使用htrf表位竞争测定的aslo0452ng1-3和aslo0543对人、食蟹猴和大鼠α-syn的特异性。使用表位竞争htrf测定,以类似的测定通过滴定未标记的α-syn来确定亲和力优化的克隆的物种交叉反应性概况,并推导针对每个物种的α-syn的ic50值。图7a和7b:典型流式细胞术结果证明亲和力优化的克隆结合至人成神经细胞瘤细胞系中的天然人α-syn。图7a和7b的组a、c、e和g显示与α-syn阴性人乳腺癌细胞系bt20的结合。图7a和7b的组b、d、f和h显示与α-syn阳性人成神经细胞瘤细胞系shsy5y的结合。图7a:本研究中使用的第一人抗体是asyn0087和huigg对照。使用第二抗人igg-fitc(jackson)检测人抗体结合。图7b:在此研究中使用的第一人抗体是aslo0452ng-3、aslo0543和nip228同种型匹配的igg1tm对照。使用第二抗人igg-fitc(jackson)检测人抗体结合。使用的第一小鼠抗体是4d6(covance)和同种型匹配的阴性对照(r&dsystems)。使用第二抗小鼠igg-fitc(sigma)检测小鼠抗体结合。图8:通过delfiaelisa的优化的抗α-synigg对聚集的人α-syn的特异性。该图显示aslo0452ng1-3和aslo0543(两个高亲和力的α-syn特异性克隆)和先导抗体asyn0087检测到捕获的聚集形式的α-syn(黑条)但未检测到捕获的单体α-syn。图9a-9c:通过免疫组织化学的亲和力优化的克隆在疾病相关组织中的特异性。图9a、9b和9c显示分别用aslo0452ng1-3、asyn0087和aslo0543染色。组a、b和c显示了aslo0452ng1-3使pd脑组织中黑质的路易体(组a和b)和路易神经突(组c)二者染色。组d显示aslo0452ngl-3显示正常脑切片中来自颞叶皮层的细胞中的α-syn的低水平染色。组e、f和g显示aslo0452ngl-3使pd脑组织中杏仁核的的路易体、路易神经突和路易斑点染色。组h显示同种型匹配的对照抗体证明未使pd脑组织中的杏仁核染色。图i至m显示asyn0087染色pd脑组织中的蓝斑(locuscoeruleus);鉴定的病理学特征是路易体(组i和l)、神经元聚集体(组j)、路易神经突(组k)和苍白体(图m)。组n和o显示aslo0543使pd脑组织中黑质中的路易体和路易神经突染色。组p显示正常脑切片中来自颞叶皮层的细胞中的α-syn的aslo0543低水平染色。图10a和10b:全身施用aslo0452ngl-3快速地降低大鼠的前额叶皮质中的游离asyn水平。aslo0452ng-3(30mg/kg静脉内;实心符号)或运载体(空心符号)处理的大鼠的isf中的平均值±sem绝对的(图10a)或相对的(图10b)游离α-突触核蛋白浓度。图11a和11b:全身施用后aslo0452ngl-3剂量和时间依赖性地降低大鼠脑脊髓液中的游离asyn水平。aslo0452ngl-3(3、10、30、100mg/kg静脉内;实心符号)或运载体(空心符号)处理的大鼠的csf中的平均值±sem绝对的(图11a)或相对的(图11b)游离α-突触核蛋白浓度。图12a-12c:抗α-突触核蛋白抗体aslo0452ngl-3和aslo0452ngl-3-d265a阻断同侧至对侧的α-突触核蛋白扩散。(图12a):注射lv-α-syn进入右侧海马体(黑色箭头)的非tg小鼠用每周给药的抗α-突触核蛋白小鼠igg1抗体(aslo0452ngl-3、aslo0452ngl-3d265a、9e4)或使用nip228同种型对照抗体被动免疫,持续13周,然后通过syn-1免疫细胞化学和自动图像分析测定α-突触核蛋白扩散。(图12b):从a中表示的同侧海马冠状切片的免疫细胞化学分析获得的α-突触核蛋白免疫反应性数据的定量。每个柱形是10个独立抗体治疗(每个抗体治疗组n=10只小鼠)的平均值±sem值。*p<0.05相对于nip228;dunnett's后检验的单向anova。(图12c):从图a中所表示的对侧海马冠状切片的免疫细胞化学分析获得的α-突触核蛋白免疫反应性数据的定量。每个柱形是10个独立抗体治疗(每个抗体治疗组n=10只小鼠)的平均值±sem值。*p<0.05相对于nip228;dunnett's后检验的单向anova。图13a-13c:抗α-突触核蛋白抗体(aslo0452ngl-3和aslo0452ngl-3-d265a)减少慢病毒表达的α-突触核蛋白沿轴突的沉积和传播。(图13a):注射lv-α-syn进入右侧海马体的非tg小鼠用每周给药的抗α-突触核蛋白小鼠igg1抗体(aslo0452ng1-3、aslo0452ng1-3d265a、9e4)或使用nip228同种型对照抗体被动免疫,持续13周,随后通过免疫细胞化学分析沿同侧和对侧经海马轴突(黑色箭头)的α-突触核蛋白沉积物。(图13b):通过syn-1的免疫细胞化学和自动图像分析确定同侧轴突α-突触核蛋白沉积物的定量。每个柱形是10个独立抗体治疗(每个抗体治疗组n=10只小鼠)的平均值±sem值。*p<0.05相对于nip228;dunnett's后检验的单向anova。(图13c):通过用syn-1的免疫细胞化学和自动图像分析确定的对侧轴突α-突触核蛋白沉积物的定量。每个柱形是10个独立抗体治疗(每个抗体治疗组n=10只小鼠)的平均值±sem值。*p<0.05相对于nip228;dunnett's后检验的单向anova。图14a-14c:抗α-突触核蛋白(抗体aslo0452ngl-3和aslo0452ngl-3-d265a)减少ca1海马体神经元和第5层新皮质神经元中α-突触核蛋白沉积。(图14a):注射lv-α-syn进入右侧海马体的非tg小鼠用每周给药的抗α-突触核蛋白小鼠igg1抗体(aslo0452ng1-3、aslo0452ng1-3d265a、9e4)或使用nip228同种型对照抗体被动免疫,持续13周,随后通过免疫细胞化学分析同侧ca1海马体神经元和同侧第5层新皮质神经元(黑色箭头)中α-突触核蛋白沉积物。(图14b):通过syn-1的免疫细胞化学和自动图像分析确定同侧第5层新皮质神经元中α-突触核蛋白沉积物的定量。所示数据表示每0.1sqmm的α-突触核蛋白阳性细胞(神经元)的数量。每个柱形是10个独立抗体治疗(每个抗体治疗组n=10只小鼠)的平均值±sem值。*p<0.05相对于nip228;dunnett's后检验的单向anova。(图14c):通过syn-1的免疫细胞化学和自动图像分析确定同侧ca1海马体神经元中α-突触核蛋白沉积物的定量。所示数据表示每0.1sqmm的α-突触核蛋白阳性细胞(神经元)的数量。每个柱形是10个独立抗体治疗(每个抗体治疗组n=10只小鼠)的平均值±sem值。*p<0.05相对于nip228;dunnett's后检验的单向anova。图15a-15c:aslo0452ngl-3和aslo0452ngl-3-d265a抗体阻断α-突触核蛋白转基因小鼠中α-突触核蛋白扩散。(图15a):注射lv-α-syn进入右侧海马体(黑色箭头)的a-syntg小鼠用每周给药的抗α-突触核蛋白小鼠igg1抗体(aslo0452ng1-3、aslo0452ng1-3d265a、9e4)或使用nip228同种型对照抗体被动免疫,持续13周,随后通过syn-1的免疫细胞化学和自动图像分析确定α-突触核蛋白扩散。(图15b):从组a中表示的同侧海马冠状切片的免疫细胞化学分析获得的α-突触核蛋白免疫反应性数据的定量。每个柱形是10个独立抗体处理的平均值±sem值(每个抗体处理组n=10只小鼠)。*p<0.05相对于nip228;dunnett's后检验的单向anova。(图15c):从组a中表示的对侧海马冠状切片的免疫细胞化学分析获得的α-突触核蛋白免疫反应性数据的定量。每个柱形是10个独立抗体治疗(每个抗体治疗组n=10只小鼠)的平均值±sem值。*p<0.05相对于nip228;dunnett's后检验的单向anova。图16a-16c:aslo0452ngl-3和aslo0452ngl-3-d265a减少转基因小鼠中慢病毒表达的α-突触核蛋白沿轴突的沉积和传播。(图16a):注射lv-α-syn进入右侧海马体(黑色箭头)的a-syntg小鼠用每周给药的抗α-突触核蛋白小鼠igg1抗体(aslo0452ng1-3、aslo0452ng1-3d265a、9e4)或用nip228同种型对照抗体被动免疫,持续13周,随后用过免疫细胞化学分析沿同侧和对侧经海马体轴突(黑色箭头)的α-突触核蛋白沉积物。(图16b):通过syn-1免疫细胞化学和自动图像分析确定同侧轴突α-突触核蛋白的定量。每个柱形是10个独立抗体治疗(每个抗体治疗组n=10只小鼠)的平均值±sem值。*p<0.05相对于nip228;dunnett's后检验的单向anova。(图16c):通过syn-1免疫细胞化学和自动图像分析确定通过对侧α-突触核蛋白的定量。每个柱形是10个独立抗体治疗(每个抗体治疗组n=10只小鼠)的平均值±sem值。*p<0.05相对于nip228;dunnett's后检验的单向anova。图17a-17d:aslo0452ngl-3和aslo0452ngl-3-d265a抗体减少α-突触核蛋白转基因小鼠中ca1海马体神经元和第5层新皮质神经元中的α-突触核蛋白沉积。(图17a):注射lv-α-syn进入右侧海马体的a-syntg小鼠用每周给药的抗α-突触核蛋白小鼠igg1抗体(aslo0452ng1-3、aslo0452ng1-3d265a、9e4)或用nip228同种型对照抗体被动免疫,持续13周,随后通过免疫细胞化学分析同侧ca1海马体神经元和第5层新皮质神经元(黑色箭头)中的α-突触核蛋白沉积物。(图17b):通过syn-1的免疫细胞化学和自动图像分析确定同侧第5层新皮质神经元中的α-突触核蛋白沉积物的定量。所示数据表示每0.1sqmm的α-突触核蛋白阳性细胞(神经元)的数量。每个柱形是10个独立抗体治疗(每个抗体治疗组n=10只小鼠)的平均值±sem值。*p<0.05相对于nip228;dunnett's后检验的单向anova。(图17c):通过syn-1的免疫细胞化学和自动图像分析确定同侧ca1海马体神经元中的α-突触核蛋白沉积物的定量。所示数据表示每0.1sqmm的α-突触核蛋白阳性细胞(神经元)的数量。每个柱形是10个独立抗体治疗(每个抗体治疗组n=10只小鼠)的平均值±sem值。*p<0.05相对于nip228;dunnett's后检验的单向anova。(图17d):通过syn-1的免疫细胞化学和自动图像分析确定对侧ca1海马体神经元中的α-突触核蛋白沉积物的定量。所示数据表示每0.1sqmm的α-突触核蛋白阳性细胞(神经元)的数量。每个柱形是10个独立抗体治疗(每个抗体治疗组n=10只小鼠)的平均值±sem值。*p<0.05相对于nip228;dunnett's后检验的单向anova。图18aslo452-ngl3-higg1tm与bbbt0626gl-scfv-bs2-also0452-ngl-3-higg1tm的表位竞争在htrf测定中证明,bbb部分的掺入不改变aslo0452-ngl3-higg1tm的结合特异性。dylight650标记的抗α-突触核蛋白抗体aslo0452hgl3-higg1tm结合至生物素化的α-突触核蛋白,所述生物素化的α-突触核蛋白又结合至穴状化合物(cyrptate)标记的链霉亲和素。激发所述穴状化合物后,发生能量转移(fret),并且当dylight650标记的aslo0452hgl3-higg1tm存在时,激发dylight650并产生荧光。如果存在竞争物igg,则阻断dylight650标记的aslo0452的结合,并防止dylight650标记的aslo0452的激发,这导致荧光信号的减少。未标记的aslo0452和bbbt0626-bs2-also0452higg1tm二者能够类似地竞争dylight650标记的aslo0452hgl3-higg1tm。图19bbbt0626gl-bs2-aslo452-ngl-3-higg1tm的小鼠脑内皮细胞结合证明,当与aslo452-ngl3-higg1tm偶联时bbb部分的有效靶标结合。已经将fmat(荧光微体积测定技术)或镜球测定技术二者用于确定抗体对脑内皮细胞的特异性结合。此分析测量人igg对小鼠脑内皮细胞(b.end3)的结合。b.end3细胞被bbbt0626higg1tm、bbbt0626glscfv-bs2-aslo0452-higg1tm和bbbt0626glscfv-bs2-nip228higg1tm相似地结合,但不被对照抗体nip228higg1tm结合。用小鼠抗fc单抗(人特异性)检测此结合,所述小鼠抗fc单抗又用alexafluor647标记的山羊抗小鼠fc检测。发明详述本发明基于aslo0452ngl-3和aslo0543的惊人和意外的发现。此发现已经形成了新的一组具有由aslo0452ngl-3和aslo0543共有的特性的抗体,以及各自具有aslo0452ngl-3和aslo0543特性的抗体亚组。在一个实施方案中,本发明的抗体或其抗原结合片段源自包含seqidno:2的氨基酸序列的可变重链区(vh)和seqidno:3的氨基酸序列的可变轻链区(vl)的抗体asyn0087,如本文所公开的。在一个特定的实施方案中,本发明的抗体或其抗原结合片段源自抗体asyn0087,其中所述抗体或抗原结合片段具有小于500nm的kd且结合与抗体asyn0087、aslo0452ngl-3和aslo0543中任一个结合的表位相同的表位,如本文所述。如同asyn0087,所述aslo0452ngl-3和aslo0543抗体结合人α-突触核蛋白的c末端区(残基96-140)。更具体地,aslo0452ngl-3和aslo0543抗体或其抗原结合片段结合人α-突触核蛋白(如seqidno:1)的约氨基酸102和约氨基酸130之间包含的区域。在一些实施方案中,本文公开的任何抗体或抗原结合片段结合由人α-突触核蛋白(如seqidno:1)的约氨基酸120和约130之间组成区域。在一些实施方案中,本文所公开的任何抗体或其抗原结合片段结合与由所述9e4抗体结合的表位不同的表位。aslo0452ngl-3和aslo0543抗体对α-突触核蛋白有选择性。所述抗体或其抗原结合片段不结合至其他突触核蛋白家族成员,如β-突触核蛋白或γ-突触核蛋白。更具体地,所述抗体或其抗原结合片段对人α-突触核蛋白是特异性的。aslo0452ngl-3和aslo0543抗体结合至人、大鼠和食蟹猴α-突触核蛋白。与不结合至人、食蟹猴和大鼠α-突触核蛋白的抗体比较,aslo0452ngl-3和aslo0543的结合至人、食蟹猴和大鼠α-突触核蛋白的能力指示结合至人α-突触核蛋白上的不同表位。因此,aslo0452ngl-3和aslo0543抗体能够用于疾病的食蟹猴和大鼠模型中的体内安全性评价和研究中。aslo0452ngl-3和aslo0543抗体以高亲和力结合至人α-突触核蛋白。aslo0452ngl-3和aslo0543抗体以如例如使用octet分析(参见如实施例9)测量的小于500皮摩尔(pm)、小于400pm、小于300pm、小于150pm、小于120pm、小于115pm、小于110pm或106pm或更小的kd结合至α-突触核蛋白。aslo0452ngl-3和aslo0543抗体以如例如使用kinexa分析(对于参考kinexa分析方案,参见如实施例9)测量的小于300皮摩尔(pm)、小于250pm、小于200pm、小于150pm、小于120pm、小于110pm或108pm、小于100pm、小于80pm或74pm或更小的kd结合至人α-突触核蛋白。aslo0452ngl-3fab片段以高亲和力结合至人α-突触核蛋白。aslo0452ngl-3fab片段以例如使用kinexa分析(参见如实施例9.3)测量的小于300皮摩尔(pm)、小于200pm、小于180pm、174pm或更小的kd结合至人α-突触核蛋白。aslo0452ngl-3和aslo0543抗体结合至天然内源性人α-突触核蛋白。aslo0452ngl-3和aslo0543抗体结合至人α-突触核蛋白的聚集体。特别地,因此,所述抗体结合至聚集不需要的表位。aslo0452ngl-3和aslo0543抗体能够隔离(sequestering)α-突触核蛋白的单体和聚集形式两者。本发明的抗体或其抗原结合片段能够结合α-突触核蛋白的单体和聚集形式。aslo0452ngl-3和aslo0543抗体结合α-突触核蛋白的疾病相关的、病理学形式,如帕金森疾病脑组织中的路易体、路易神经突、路易斑点。在正常(非疾病的)脑中观察到最小染色。aslo0452ngl-3抗体减少脑组织间液中的α-突触核蛋白水平。特别地,aslo0452ngl-3抗体减少脑组织间液中的游离未结合α-突触核蛋白水平。aslo0452ngl-3抗体减少脑脊髓液中α-突触核蛋白水平。特别地,aslo0452ngl-3抗体减少脑脊髓液中游离未结合α-突触核蛋白水平。aslo0452ngl-3抗体减少体内α-突触核蛋白扩散。与不抑制扩散的抗体比较,这种抑制α-突触核蛋白扩散的新功能指示与人α-突触核蛋白上的不同表位结合。在一些实施方案中,本文公开的任何抗体或其抗原结合片段具有aslo0452ngl-3的功能特性(如本文所列举的任何aslo0452ngl-3功能特性)中的任一个或多个。在一些实施方案中,本文公开的任何抗体或其抗原结合片段具有aslo0543的功能特性(如本文所列举的任何aslo0543功能特性)中的任一个或多个。在一个实施方案中,本发明的抗体或其抗原结合片段与抗体aslo0452ngl-3和/或aslo0543竞争结合至人α-突触核蛋白。在另一个实施方案中,本发明的抗体或其抗原结合片段结合至与抗体aslo0452ngl-3和/或aslo0543所结合的人α-突触核蛋白上的相同的表位。可容易地确定抗体或其抗原结合片段是否结合至如上述定义的参考抗体或其抗原结合片段的表位。这些方法是本领域中常规手段。例如,可通过生物化学竞争测定将抗体与另一个比较,由此两个抗体(一个出于检测目的标记且一个未标记)与指定抗原同时温育。如果该标记的抗体实现结合信号,则认为所述两个抗体识别感兴趣的蛋白质上的独特的非重叠表位。如果获得未结合信号,则相反地,它们的特征在于在蛋白质序列上具有重叠的表位,因为一种抗体的结合在空间上阻碍了第二抗体的结合。进一步地,也可使用修饰的蛋白质(例如截短物、源自抗原一级氨基酸序列的线性肽序列、物种直向同源物)以及通过蛋白水解消化和结合至指定蛋白的抗体的质谱分析来鉴定指定表位的氨基酸位点。这些方法学用于产生抗体和抗原之间相互作用的区域。可进行另外的常规实验(例如肽突变和结合分析)以确认任何观察到的结合缺失是否实际上是由于结合本发明的表位或者某些其他现象(例如空间位阻)是否有责任。可使用elisa、ria、biacore、流式细胞术或其他已知的抗体结合测定进行此类实验。例如,为了精确定位特定表位,表位:互补位界面的数学模型可由通过使用高分辨率成像方法(如与x射线衍射共结晶)解析抗原:抗体复合物的结构所产生的数据而得到。为了确认所得数学模型的相关性,在鉴定定义表位的关键接触残基方面,必须随后进行抗原的点诱变,并建立由这种突变引起的对抗原和抗体之间结合强度的影响的分析。使用这种组合方法,可建立包含表位的关键接触残基的精确图谱。结合至本发明的抗体或其抗原结合片段的表位的抗体或其抗原结合片段可通过产生本发明的抗体或其抗原结合片段的变体来生成。此类变体抗体或其抗原结合片段可具有与本发明的抗体或其抗原结合片段的cdr具有高水平同一性的cdr。例如,在一些实施方案中,本文公开的任何抗体或其抗原结合片段的任何cdr与本文提及的任一种或多种特定cdr序列(如具有seqidno:5、15、16、20、10和21的cdr中的任一种或多种)相比,可相差1或2个氨基酸残基。此外,这些抗体可在框架区具有一个或多个变化(如保守氨基酸取代)。在一个实施方案中,本发明的抗体或其抗原结合片段具有cdr氨基酸序列中的变化,所述cdr氨基酸序列与抗体aslo0452ngl-3的cdr维持至少80%、至少85%、至少90%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%和直至99%序列同一性。特别地,涵盖保守氨基酸取代。保守取代是在具有相关侧链的氨基酸家族内发生的那些。遗传学上编码的氨基酸通常分为下列家族:(1)酸性:天冬氨酸、谷氨酸;(2)碱性:赖氨酸、精氨酸、组氨酸;(3)非极性:丙氨酸、缬氨酸、亮氨酸、异亮氨酸、脯氨酸、苯丙氨酸、甲硫氨酸、色氨酸;和(4)无电荷极性:甘氨酸、天冬酰胺、谷氨酰胺、半胱氨酸、丝氨酸、苏氨酸、酪氨酸。这些家族可进一步分类:丝氨酸和苏氨酸是脂肪族-羟基家族;天冬酰胺和谷氨酰胺是含有酰胺的家族;丙氨酸、缬氨酸、亮氨酸和异亮氨酸是脂肪族家族;以及苯丙氨酸、色氨酸和酪氨酸是芳香族家族。因此,一般来说,可预期用异亮氨酸或缬氨酸单独替代亮氨酸、用天冬氨酸单独替代谷氨酸、用丝氨酸单独替代苏氨酸或者用结构相关的氨基酸类似地替代氨基酸,其将不会对所得抗体的结合功能或特性具有重要影响,特别是该替代不涉及cdr位点内的氨基酸。在一个实施方案中,根据本发明的抗体或其抗原结合片段包含至少一个cdr,所述cdr选自下列:(i)seqidno:5的h-cdr1,(ii)seqidno:6的h-cdr2,(iii)seqidno:7的h-cdr3,(iv)seqidno:9的l-cdr1,(v)seqidno:10的l-cdr2,(vi)seqidno:11的l-cdr3。在一个实施方案中,本发明的抗体或其抗原结合片段具有至少一个cdr,其选自抗体aslo0452ngl-3的cdr,即至少一个选自seqidno:5、seqidno:15、seqidno:16、seqidno:20、seqidno:10、seqidno:21中任一个的cdr。在另一个实施方案中,本发明的抗体或其抗原结合片段的重链的cdr是抗体aslo0452ngl-3的重链的cdr3;和/或本发明的抗体或其抗原结合片段的轻链的cdr3是抗体aslo0452ngl-3的轻链的cdr3。因此,在一个实施方案中,根据本发明的抗体或其抗原结合片段的重链的cdr3是抗体aslo0452ngl-3的重链的seqidno:16的cdr3;和/或根据本发明的抗体或其抗原结合片段的cdr3是抗体aslo0452ngl-3的轻链的seqidno:21的cdr3。在进一步的实施方案中,本发明的抗体或其抗原结合片段的重链的cdr3是抗体aslo0452ngl-3的重链的cdr3。在一个实施方案中,本发明的抗体或其抗原结合片段的轻链的cdr3是抗体aslo0452ngl-3的轻链的cdr3。在一个实施方案中,本发明的抗体或其抗原结合片段具有抗体aslo0452ngl-3的6个cdr,即具有氨基酸序列seqidno:5、seqidno:15和seqidno:16的三个重链cdr;和具有氨基酸序列seqidno:20、seqidno:10和seqidno:21的三个轻链cdr。本发明提供包含可变重链和可变轻链的本发明的抗体或其抗原结合片段,所述可变重链与由seqidno:13定义的核苷酸序列具有至少80%、85%、90%、95%、96%、97%、98%、99%或100%同一性且所述轻链与由seqidno:18定义的核苷酸序列具有至少80%、85%、90%、95%、96%、97%、98%、99%或100%同一性。本发明还提供包含可变重链和可变轻链的抗体或其抗原结合片段,所述可变重链具有由seqidno:13定义的核苷酸序列且所述轻链具有由seqidno:18定义的核苷酸序列。本发明提供包含可变重链和可变轻链的本发明的抗体或其抗原结合片段,所述可变重链与由seqidno:14定义的氨基酸序列具有至少80%、85%、90%、95%、96%、97%、98%、99%或100%同一性且所述轻链与由seqidno:19定义的氨基酸序列具有至少80%、85%、90%、95%、96%、97%、98%、99%或100%同一性。在一个实施方案中,所述抗体或其抗原结合片段包含(i)可变重链和可变轻链,所述可变重链与seqidno:14定义的氨基酸序列具有至少80%、85%、90%、95%、96%、97%、98%、99%或100%同一性且所述轻链与seqidno:19定义的氨基酸序列具有至少80%、85%、90%、95%、96%、97%、98%、99%或100%同一性,和(ii)抗体aslo0452ngl-3的6个cdr。本发明提供包含可变重链和可变轻链的根据本发明的抗体或其抗原结合片段,所述可变重链具有seqidno:14的氨基酸序列且所述可变轻链具有seqidno:19的氨基酸序列。在一个实施方案中,根据本发明的抗体或其抗原结合片段具有抗体aslo0543的6个cdr。因此,在一个实施方案中,根据本发明的抗体或其抗原结合片段包含:a)具有下列序列的3个重链cdr:(i)seqidno:25的h-cdr1,(ii)seqidno:26的h-cdr2;和(iii)seqidno:27的h-cdr3,和b)具有下列序列的3个轻链cdr:(i)seqidno:31的l-cdr1,(ii)seqidno:32的l-cdr2;和(iii)seqidno:33的l-cdr3。在进一步的实施方案中,根据本发明的抗体或其抗原结合片段包含可变重链和可变轻链,所述可变重链与由seqidno:14定义的序列具有至少80%、85%、90%、95%、96%、97%、98%、99%或100%同一性且所述轻链与由seqidno:19定义的序列具有至少80%、85%、90%、95%、96%、97%、98%、99%或100%同一性,且进一步包含:a)具有下列序列的3个重链cdr:(vii)seqidno:25的h-cdr1,(viii)seqidno:26的h-cdr2;和(ix)seqidno:27的h-cdr3,和b)具有下列序列的3个cdr:(vii)seqidno:31的l-cdr1,(viii)seqidno:32的l-cdr2;和(ix)seqidno:33的l-cdr3。本发明还提供包含可变重链和可变轻链的抗体或其抗原结合片段,所述可变重链具有由seqidno:23定义的核苷酸序列且所述可变轻链具有由seqidno:29定义的核苷酸序列的。本发明还提供包含可变重链和可变轻链的本发明的抗体或其抗原结合片段,所述可变重链具有seqidno:24的氨基酸序列且所述可变轻链具有seqidno:30的氨基酸序列。在进一步的实施方案中,所述抗体或其抗原结合片段包含由seqidno:22定义的氨基酸序列的重链和具有由seqidno:28定义的氨基酸序列的轻链。可精确地定义抗体的框架区和cdr(参见kabat等.sequencesofproteinsofimmunologicalinterest,第五版,u.s.departmentofhealthandhumanservices(1991),91-3242,1991;和chothia等j.moi.biol.(1987),196:901-917,其中二者通过提述并入本文)。预期本发明的抗体或其抗原结合片段的氨基酸序列的微小变化涵盖在本发明中,条件是所述氨基酸序列中的变化与本文任意处所定义的本发明的抗体或其抗原结合片段保持至少75%、更优选至少80%、至少90%、至少95%且最优选至少99%序列同一性。特别地,涵盖保守氨基酸替代。本发明还提供包含如本文任意处所定义的本发明的抗体或其抗原结合片段的轻链的单链氨基酸序列。本发明还提供包含如本文任意处所定义的本发明的抗体或其抗原结合片段的重链的单链氨基酸序列。用于比较的序列最佳比对可(例如)通过以下进行:通过smith和waterman的局部同源性比对算法(smith和waterman,adv.appl.math.2(1981),484;其通过提述并入本文),通过needleman和wunsch的算法(needleman和wunsch,j.mol.biol.(1970),48:443;其通过提述并入本文),通过pearson和lipman的相似性方法的检索(pearson和lipman,procnatlacadsciusa(1988),85:2444;其通过提述并入本文),通过这些算法的计算机执行(gap、bestfit、fasta和tfasta-遗传学计算机组的序列分析软件包,universityofwisconsinbiotechnologycenter,1710universityavenue,madison,wis.53705)或者通过目测(参见currentprotocolsinmolecularbiology,f.m.ausbel等编,currentprotocols,ajointventurebetweengreenepublishingassociates,in.andjohnwiley&sons,inc.(1995增刊)ausbubel;其通过提述并入本文)。适用于确定序列相似性或同一性百分比的算法的实例是blast和blast2.0算法(参见altschul等j.mol.biol.(1990),215(3):403-410;和“http://www.ncbi.nlm.nih.gov/”ofthenationalcenterforbiotechnologyinformation;其中二者通过提述并入本文)。在一个实施方案中,本发明的抗体或其抗原结合片段是分离的。在另一个实施方案中,本发明的抗体或其抗原结合片段是纯化的。在一个实施方案中,本发明的抗体或其抗原结合片段是单克隆抗体。在另一个实施方案中,本发明的抗体或其抗原结合片段是人源化抗体。在又一个实施方案中,本发明的抗体或其抗原结合片段是人抗体。在一个实施方案中,本发明的抗体或其抗原结合片段是iga、igd、ige、igm或igg,如igg1、igg2、igg3和igg4抗体或其抗原结合片段。在另一个实施方案中,本发明的抗体或其抗原结合片段已降低了对igg1fc受体的结合亲和力。因此,本发明的抗体或抗原结合片段具有低的免疫原性作用。在一个实施方案中,所述抗体或其抗原结合片段是igg1tm抗体或其抗原结合片段。igg1tm是igg1三突变体,其包含fc域中的3个点突变(l234f/l235e/p331s),所述突变可降低所述抗体或其抗原结合片段与fc-γ受体(fcγr)的结合亲和力(oganesyan等actacrystallogrdbiolcrystallogr,(2008)64:700-704;其通过提述并入本文)。在一些实施方案中,因此通过本发明的抗体对抗体介导的α-突触核蛋白扩散的阻止不需要fc相关效应器功能作为关键作用机制。抗原结合片段包括fab、fv、scfv、dab、fd、fab’、f(ab’)2或具有足够的结合框架的分离的互补决定区(cdr)。fab片段可为由vl、vh、cl和ch1域组成的单价片段。f(ab')2片段可为包含在铰链区通过二硫桥连接的两个fab片段的双价片段。fc片段可由ch2和ch3域组成。fv片段可由抗体的单臂的vl和vh域组成。dab片段(ward等nature(1989),341:544-546;其通过提述并入本文)可以由vh域组成。具有足够结合框架的分离的互补决定区(cdr)可为可变区的抗原结合部分。轻链可变区的抗原结合部分和重链可变区的抗原结合部分(如fv片段的两个域、vl和vh)可使用重组的方法通过使它们能够制成单个蛋白链,其中vl和vh区配对以形成单价分子的合成的接头连接(称之为单链fv(scfv);参见如bird等science(1988),242(4877):423-426;和huston等procnatlacadsciusa(1988),85:5879-5883;其中二者通过提述并入本文)。这些是使用本领域技术人员已知的常规技术获得的,并且以与完整抗体相同的方式筛选部分的功用。本发明的抗体或其抗原结合片段可具有如上所定义的任何或全部有利特性或其组合。特别地,本发明的抗体或其抗原结合片段对α-突触核蛋白有选择性,并且能够减缓或阻止体内α-突触核蛋白的细胞至细胞的传播和扩散。使用实施例中本文所述的技术通过测定其比活性可容易地确定本发明得到的抗体或其抗原结合片段的功能且特别是(i)其结合α-突触核蛋白的表位的能力;和(ii)其减慢或阻止体内α-突触核蛋白的细胞至细胞传播和扩散的能力。本公开提供递送根据本发明的抗体或其抗原结合片段穿过血脑屏障(bbb)的组合物,其使用可穿过脑内皮细胞同时与所述抗体或片段关联(例如同时与所述抗体或片段融合或缀合)的转运蛋白分子。本文提供bbb序列。如本文所用,术语“有效负载(payload)”用作本文所述的抗体或其抗原结合片段的简写,其可通过本文提供的转运蛋白分子促进转运穿过bbb。在特定的实施方案中,所述“有效负载”涵盖本发明抗体的重链可变区,更特别地是also0452ngl-3或aslo0543的重链可变区。有效负载是转运蛋白分子的一部分(如作为融合多肽)或通过二硫键或其他共价键与所述多肽连接。可替换地,有效负载可以任何允许转运蛋白分子促进其穿过bbb的转运的方式与转运蛋白分子相关联,如下文进一步所述的。在某些方面,有效负载在bbb转运后仍然是转运蛋白分子的一部分,并以此形式保留中枢神经系统(cns)活性。可替换地,有效负载可在bbb转运期间与转运蛋白分子相关联,但是以允许其在bbb转运后与转运蛋白分子解离的方式。本公开进一步提供用于治疗或诊断cns的疾病或病症(特别是α-突触核蛋白病)的方法,其包含使用这些与根据本发明的抗体或其抗原结合片段相关联的转运蛋白分子。在某些方面,本公开提供包含免疫球蛋白衍生的多肽的分离的转运蛋白分子。在某些方面,所述多肽是骆驼抗体fc5的模拟物或非模拟物,使用荧光微量体积测定技术(fmat)鉴定和分离所述多肽以检测与脑微血管内皮细胞(bmvec),例如小鼠b.end3细胞的结合。在某些方面,所述免疫球蛋白衍生的多肽是抗体或其抗原结合片段,其中“活性的”意指转运蛋白分子可(例如)与一个或多个物种的bmvec(如小鼠bmvec、大鼠bmvenc、食蟹猴bmvec或人bmvec)结合,内化到一个或多个物种的bmvec中和/或单独或与有效负载相关联穿过血脑屏障。在一些实施方案中,所述bbb转运蛋白分子是美国临时专利申请号62/094,503(其通过提述以其整体并入本文)中所述的bbb转运蛋白分子。在某些方面,所述转运蛋白分子包含如美国临时专利申请号62/094,503(其通过提述以其整体并入本文)中所述的bbbt0241、bbbt0626、bbbt0626gl、bbbt0632、bbbt0632glbbbt0654、bbbt0726、bbbt0727、bbbt0732、bbbt0754、bbbt0674、bbbt0755、bbbt0643、bbbt0579或bbbt0671中的一个或多个。在特定的实施方案总,所述bbb转运蛋白分子是bbbt0626或bbbt0632。在某些实施方案中,所述bbb转运蛋白分子是种系的,如bbbt0626gl是bbbt062的种系形式,称之为“bbbt0626gl”。在进一步特定的实施方案中,所述bbb转运蛋白分子是bbbt0632gl或bbbt0626gl。在某些方面,转运蛋白分子不与bmvec结合但是仍能够如体外转胞吞作用测定中所示的转运穿过bbb。关于bbb转运蛋白分子,所述vhcdr对应于经典kabat编号位点,即kabath-cdr1位于位点31-35,h-cdr2是位点50-65,且h-cdr3位于位点95-102。l-cdr2和l-cdr3也对应于经典kabat编号位点,即各自为位点50-56和89-97。如本文所用,术语“l-cdr1”或“轻链cdr1”对应于位于vl中的kabat位点23-34的序列(与此相对,根据kabat编号模式的经典的l-cdr1位点对应于位点24-34)。在某些方面,所述免疫球蛋白衍生的多肽包含免疫球蛋白重链互补决定区(cdr)。例如,所述免疫球蛋白衍生的多肽可包括免疫球蛋白重链互补决定区-1(h-cdr1)、免疫球蛋白重链互补决定区-2(vh-cdr2)、免疫球蛋白重链互补决定区-3(vh-cdr3)。在某些方面,所述免疫球蛋白衍生的多肽可进一步包含或可替换地包含免疫球蛋白轻链cdr。例如,所述免疫球蛋白衍生的多肽可包括免疫球蛋白轻链互补决定区-1(l-cdr1)、免疫球蛋白轻链互补决定区-2(l-cdr2)和免疫球蛋白轻链互补决定区-3(l-cdr3)。在某些方面,所述免疫球蛋白衍生的多肽含有各自具有下列氨基酸序列的h-cdr1、h-cdr2、h-cdr3、l-cdr1、l-cdr2和l-cdr3:(a)seqidno:40作为h-cdr1、seqidno:41作为h-cdr2、seqidno:42作为h-cdr3、seqidno:36作为l-cdr1、seqidno:37作为l-cdr2和seqidno:38作为l-cdr3,其中所述cdr与bbbt0626和bbbt0626gl的cdr相似。(b)seqidno:40作为h-cdr1、seqidno:41作为h-cdr2、seqidno:42作为h-cdr3、seqidno:44作为l-cdr1、seqidno:45作为l-cdr2和seqidno:46作为l-cdr3,其中所述cdr与bbbt0626和bbbt0626gl的cdr相同。在某些方面,所述免疫球蛋白衍生的多肽含有各自具有下列氨基酸序列的h-cdr1、h-cdr2、h-cdr3、l-cdr1、l-cdr2和l-cdr3:(a)seqidno:49作为h-cdr1、seqidno:50作为h-cdr2、seqidno:51作为h-cdr3、seqidno:53作为l-cdr1、seqidno:54作为l-cdr2和seqidno:55作为l-cdr3,其中所述cdr与bbbt0632gl的cdr相似。(b)seqidno:49作为h-cdr1、seqidno:50作为h-cdr2、seqidno:51作为h-cdr3、seqidno:53作为l-cdr1、seqidno:54作为l-cdr2和seqidno:55作为l-cdr3,其中所述cdr与bbbt0632gl的cdr相同。在某些可替换的实施方案中,如上所述的一个或多个cdr与所列举的cdr相同,除了例如1、2、3、4或5个单个氨基酸的缺失、取代或插入。在某些实施方案中,上述提供的转运蛋白分子可穿过血脑屏障。在某些方面,所述h-cdr1、h-cdr2、h-cdr3、l-cdr1、l-cdr2和l-cdr3可位于免疫球蛋白框架区以产生抗体vh和抗体vl。在某些方面,框架区可以是人来源的框架区。在某些方面,例如通过柔性肽接头使所述抗体vh和抗体vl融合在一起以形成scfv分子。在某些方面,所述vh和vl进一步包含一个或多个免疫球蛋白恒定域,如ch1域、铰链区、ch2域、ch3域、cl-κ域和/或clλ域。在某些方面,所述一个或多个免疫球蛋白恒定域源自人免疫球蛋白,如人igg1免疫球蛋白。在某些方面,所述vh和vl和/或恒定域可包含突变以促进例如更长或更短的半衰期,增加的或减少的效应器功能或经由肽融合、二硫键或化学缀合附接有效负载的能力。在本发明的某些方面,提供与可穿过脑内皮细胞的转运蛋白分子相关的根据本发明的抗体或其抗原结合片段,如本文所述。在特定的方面,本公开提供与包含免疫球蛋白衍生的多肽的转运蛋白分子相关的根据本发明的抗体或其抗原结合片段,其中所述多肽包含免疫球蛋白重链可变区(vh)区域和免疫球蛋白轻链可变区(vl)区域。在某些方面,所述免疫球蛋白衍生的多肽包含本文提供的序列,其包括:(a)vh氨基酸序列和vl氨基酸序列,所述vh氨基酸序列与seqidno:39至少80%、84%、85%、90%、95%、96%、97%、98%或99%相同,并且所述vl氨基酸序列与seqidno:43至少80%、84%、85%、90%、95%、96%、97%、98%或99%相同,其中seqidno:39和seqidno:43编码bbbt0626gl的vh和vl区。(b)vh氨基酸序列和vl氨基酸序列,所述vh氨基酸序列与seqidno:47至少80%、84%、85%、90%、95%、96%、97%、98%或99%相同,并且所述vl氨基酸序列与seqidno:43至少80%、84%、85%、90%、95%、96%、97%、98%或99%相同,其中seqidno:47和seqidno:43编码bbbt0626的vh和vl区。(c)vh氨基酸序列和vl氨基酸序列,所述vh氨基酸序列与seqidno:48至少80%、84%、85%、90%、95%、96%、97%、98%或99%相同,并且所述vl氨基酸序列与seqidno:43至少80%、84%、85%、90%、95%、96%、97%、98%或99%相同,其中seqidno:48和seqidno:43编码bbbt0632的vh和vl区。在某些方面,本公开提供与包含免疫球蛋白衍生的多肽的转运蛋白分子相关的根据本发明的抗体或其抗原结合片段,其中所述免疫球蛋白衍生的多肽包含vh区和vl区,其中:(a)所述vh区包含seqidno:34,且所述vl区包含seqidno:35;或(b)所述vh区包含seqidno:39,且所述vl区包含seqidno:43;或(c)所述vh区包含seqidno:39,且所述vl区包含seqidno:35;或(d)所述vh区包含seqidno:47,且所述vl区包含seqidno:43;或(e)所述vh区包含seqidno:47,且所述vl区包含seqidno:35;或(f)所述vh区包含seqidno:48,且所述vl区包含seqidno:52;或在进一步的实施方案中,共价连接如上文所述的转运蛋白分子的vh和vl区以形成单链片段(scfv)。在某些方面,本文提供的转运蛋白分子具有转运蛋白活性,例如它可与来自一个或多个物种的bmvec(如小鼠、大鼠、食蟹猴或人bmvec)结合,它可以内化成一个或多个物种的bmvec,或者它可穿过血脑屏障。在某些方面,如本文所提供的转运蛋白分子包含免疫球蛋白衍生的多肽,其中所述免疫球蛋白衍生的多肽包含抗体或其bbb可穿透的片段。如本文所述的“bbb可穿透的片段”是可特异性结合至一个或多个物种的bmvec且穿过体外或体内bmvec从外周血管进入cns血管的转运蛋白分子的片段。可通过本领域普通技术人员已知的各种体外或体内测定来检测给定片段是否是bbb可穿透的片段。例如,如us62/094,503中所述,可在体外转胞吞作用测定中、在体内测定(如利尿测定)中检测转运蛋白分子。可用于测定穿过bbb的有效负载的体内递送的其他测定包括但不限于慢性压迫性损伤(cci);保留性神经损伤模型(sni)或脊神经结扎(snl),其中所有这些都可经由pawflick或hargreaves方法测定(hargreavesk,等,pain;1988;32;77-88)。在某些方面,如本文所提供的转运蛋白分子可与来自一个或多个物种的bmvec(如人、食蟹猴、鼠类、大鼠或牛bmvec)结合。可以本领域普通技术人员已知的各种方式(例如如us62/094,503中所述的fmat测定)证明结合。在某些方面,所述bmvec是脑血管内皮细胞(bcec)。在某些方面,如本文所提供的转运蛋白分子可在体外转胞吞作用测定中穿过单层bcec。在某些方面,可通过cns中转运蛋白蛋白分子的可视化来证明转运蛋白分子活性。例如,可将氚标记的转运蛋白分子外周地(例如静脉内地)递送至受试者(例如小鼠),然后通过定量全身放射线照相术在cns中可视化。在某些方面,所述转运蛋白分子位于cns的特定区域,如小脑皮质、大脑灰质、脊髓灰质、脑桥或其组合。在某些方面,如本文所述的转运蛋白分子包含包含两个或多个亚基(如重链或其片段和轻链或其片段)或由其组成的抗体或其bbb可穿透片段,其中所述重链或轻链是相关联的,如作为单个融合蛋白(如scfv)或作为通过一个或多个二硫键保持在一起的两个亚基。在一些方面,所述重链包含vh域或区,且所述轻链包含vl域或区。在一个实施方案中,本发明提供与血脑屏障转运蛋白分子相关联的根据本发明的抗体或其抗原结合片段,如本文所述。在特定的实施方案中,根据本发明的抗体或其抗原结合片段与血脑屏障转运蛋白分子相关联,其中所述转运蛋白分子是单链片段(scfv),其包含:i.seqidno:39的bbbt0626gl的重链可变区(vh)和seqidno:43的bbbt0626gl的轻链可变区(vl),或ii.seqidno:47的bbbt0626的重链可变区(vh)和seqidno:43的bbbt0626的轻链可变区(vl),iii.seqidno:48的bbbt0632gl的重链可变区(vh)和seqidno:52的bbbt0632gl的轻链可变区(vl)在某些方面,所述重链进一步包含重链恒定域(如ch1域、铰链、ch2域和/或ch3域)或其片段。在某些方面,所述重链恒定域是igg恒定域或其片段,如人igg恒定域,如人igg1、igg2、igg3或igg4恒定域。在某些方面,所述igg恒定域或其片段包含相对于野生型igg恒定域的改变了的糖基化和/或一个或多个氨基酸取代,其中该修饰的igg具有特定的性质,例如相较于具有野生型igg恒定域的igg的半衰期的增加的或减少的半衰期,相对于野生型igg恒定域的增加或减少的效应器功能,或经由肽键、二硫键或化学缀合连接异源部分的能力。在某些方面,所述igg恒定域或其片段具有相对于野生型igg恒定域的改变了的糖基化,其中该修饰的igg具有特定的性质,例如例如相较于具有野生型igg恒定域的igg的半衰期的增加的或减少的半衰期,相对于野生型igg恒定域的增加或减少的效应器功能。在一些实施方案中,如本文所定义的,根据本发明的抗体或其抗原结合片段与bbbt0626或bbbt0626gl相关联,其形成双特异性抗体分子。在其他实施方案中,根据本发明的所述双特异性抗体包含与根据本发明的抗α-突触核蛋白抗体的单链片段(scfv)相关联的人igg1tm骨架(即igg1tm的重链ch1、ch2、ch3区),所述单链片段包含移接至重链的n末端(“bis2形式”)或c末端(“bis3形式”)或vl的n末端(“bis1形式”)的bbbt0626或bbbt0626gl的vh和vl区。bis形式参考公开于dimasi等.jmolbiol.2009年10月30日;393(3):672-92中。在一些实施方案中,根据本发明的所述双特异性抗体进一步包含包含与根据本发明的抗α-突触核蛋白抗体的vl相关联的κ或λcl区的轻链。在特定的实施方案中,根据本发明的双特异性抗体包含人igg1tm骨架,其与下列相关联:(i)bbbt0626gl的单链片段,其包含seqidno:39的重链可变区(vh)和seqidno:43的轻链可变区(vl);或(ii)bbbt0626的单链片段,其包含seqidno:47的重链可变区(vh)和seqidno:43的轻链可变区(vl),其中将所述scfv移接至seqidno:12的aslo0452ngl-3的重链的n末端(bis2形式)或c末端(bis3形式)或seqidno.17的轻链的n末端(“bis1形式”)。在更进一步的特定实施方案中,根据本发明的双特异性抗原包含与bbbt0626gl的单链片段(scfv)相关联的人igg1tm骨架,所述单链片段包含(i)seqidno:39的重链可变区(vh)和(ii)seqidno:43的轻链可变区(vl);其中将所述scfv移接至seqidno:12的aslo0452ngl-3的重链的n末端(bis2形式)或c末端(bis3形式)或seqidno.17的轻链的n末端(“bis1形式”)。在其他特定的实施方案中,根据本发明的双特异性抗体包含人igg1tm骨架,其与下列相关联:(i)bbbt0626gl的单链片段(scfv),其包含seqidno:39的重链可变区(vh)或seqidno:43的轻链可变区(vl);或(ii)bbbt0626的单链片段,其包含seqidno:47的重链可变区(vh)和seqidno:43的轻链可变区(vl),其中将所述scfv移接至seqidno:22的aslo0543的重链的n末端(bis2形式)或c末端(bis3形式)或seqidno.28的轻链的n末端(“bis1形式”)。本发明还提供该发明的抗体或其抗原结合片段,其供用作药物。本发明还提供该发明的抗体或其抗原结合片段,其用于预防或治疗中枢神经系统的疾病,特别是α-突触核蛋白病。在一个实施方案中,所述α-突触核蛋白病选自帕金森氏病(pd)、路易体痴呆(dlb)和多系统萎缩症(msa)。在优选的实施方案中,所述α-突触核蛋白病是帕金森氏病(pd)。本发明还提供该发明的抗体或其抗原结合片段的用于制造用于预防或治疗中枢神经系统的疾病,特别是α-突触核蛋白病的药物的用途。在一个实施方案中,所述α-突触核蛋白病选自帕金森氏病(pd)、路易体痴呆(dlb)和多系统萎缩症(msa)。在优选的实施方案中,所述α-突触核蛋白病是帕金森氏病(pd)。本发明该提供治疗或预防患者中的疾病的方法,该方法包含向所述患者施用本发明的抗体或其抗原结合片段。在一个实施方案中,所述α-突触核蛋白病选自帕金森氏病(pd)、路易体痴呆(dlb)和多系统萎缩症(msa)。在优选的实施方案中,所述α-突触核蛋白病是帕金森氏病(pd)。在使用中,本发明的抗体或其抗原结合片段能够通过抑制体内α-突触核蛋白的传播和扩散来治疗或防止疾病进展。因此,本发明的抗体或其抗原结合片段提供优于其他疗法的明显优势。本发明还提供在有需要的受试者中减缓或防止疾病进展的方法,其包括向所述患者施用本发明的抗体或其抗原结合片段。在一个实施方案中,所述治疗疾病的方法包括施用治疗有效量的本发明的抗体或其抗原结合片段。在另一个实施方案中,所述预防疾病或减缓或防止疾病进展的方法包括施用预防有效量的本发明的抗体或其抗原结合片段。施用本发明的抗体或其抗原结合片段的剂量范围是产生所需治疗效果的剂量范围。应当理解,所需的剂量范围取决于所述抗体或其抗原结合片段或组合物的明确的性质、施用途径、制剂的性质、患者的年龄、本质、患者病况的程度或严重性、禁忌症(如果有的话)和主治医师的判断。可使用标准经验方式调整这些剂量水平中的变化以进行优化。适当的剂量是每千克体重的1至50mg的范围。它们可为5至30mg/kg、10至25mg/kg或15至20mg/kg的范围。单位剂量可以每日或更少的频率(例如每周或每月)施用。可通过长时间的重复施用本发明的抗体或其抗原结合片段来实现施用。施用可为并发的或连续的,并且可以任何顺序实现施用。本文所定义的预防或治疗可作为单独疗法应用,或者除了本发明的抗体或其抗原结合片段,可包含施用通常用于治疗α-突触核蛋白并的其他药剂或既定疗法(例如l-3,4-二羟基苯丙氨酸(l-dopa)、多巴胺(受体)激动剂、儿茶酚-o-甲基转移酶(cmot)抑制剂和/或单胺氧化酶b型(mao-b)抑制剂)。其他药剂或既定疗法的施用可与本发明的抗体或抗原结合片段组合,或作为其附属物或与其结合,并且可通过同时、顺序或单独地给药治疗的单个组分。组合治疗可以本领域技术人员认为必需的或方便的任何方式进行,并且出于此说明书的目的,预期对于组合中所用的化合物的顺序、数量、重复或相对量没有限制。治疗有效量是指抗体或其抗原结合片段的量,当单独施用或组合施用至患者用于治疗疾病或至少一种疾病的临床症状时,其足以影响疾病或症状的这种治疗。所述治疗有效量可以根据(例如)抗体和/或疾病症状、待治疗患者的年龄、体重和/或健康以及处方医师的判断而变化。在任何给定的情况下,适当的治疗有效量可由本领域技术人员确定或能够通过常规实验确定。治疗有效量也是其中所述抗体或抗体或其抗原结合片段的任何毒性或有害作用超过有益效果的量。“预防有效量”是当单独或组合施用至患者时,抑制或延迟疾病或至少一种疾病的临床症状的发作或复发的抗体或抗体或其抗原结合片段的任何量。在一些实施方案中,所述预防有效量完全阻止所述疾病的发作或复发。“抑制”发作意指减少疾病发作的可能性或完全预防疾病的发作。本发明还提供了包含本发明的抗体或其抗原结合片段的药物组合物。因此,本发明提供了药物组合物,其包含本发明的抗体或其抗原结合片段,以及药学上可接受的赋形剂。适当的药学上可接受的赋形剂可促进活性化合物加工成适用于药物施用的制剂。可配制本发明的药物组合物以用于但不限于肠胃外递送,例如肌肉内、皮下或静脉内。适用于肌肉内、皮下或静脉内注射的组合物包括无菌水溶液。所述药物组合物可采取水溶液的形式,并且可包括生理上相容的缓冲液,例如hank溶液、ringer溶液或生理缓冲盐水。所述药物组合物可另外地或可替换地含有增加悬浮液粘度的物质,如羧甲基纤维素钠、山梨糖醇或葡聚糖。所述药物组合物可制备成适当的油性注射悬浮液。适当的亲脂性溶剂或运载体包括脂肪油(如芝麻油)或合成脂肪酸酯(如油酸乙酯或甘油三酯)或脂质体。任选地,所述药物组合物可含有合适的稳定剂或增加化合物溶解度的药剂,以允许制备高浓度的溶液。本发明提供编码本发明的抗体或其抗原结合片段的分离的核酸分子。本发明还提供包含本发明的分离的核酸分子的载体。本发明进一步提供包含本发明的载体的宿主细胞。本发明的抗体或其抗原结合片段不限于特定的生成(generation)或生产(production)方法。因此,本发明提供由分泌抗体的杂交瘤制备的抗体,以及已经用编码所述抗体的核酸转化或转染的重组生产的细胞产生的抗体。这种杂交瘤、重组生产的细胞和核酸形成本发明的一部分。实施例实施例1:抗体产生使用一系列对重组人α-syn(“huα-syn”)的选择循环从噬菌体展示文库中分离抗α-syn特异性抗体,均被动地固定在微量滴定孔上并游离于溶液中。克隆到基于丝状噬菌体m13的噬菌粒载体中的幼稚人单链fv(scfv)噬菌体展示文库用于选择(lloyd等,proteinengdessel.(2009),22(3):159-68;和vaughan等,natbiotechnol.(1996),14(3);309-14;其中二者通过提述并入本文)。在上述两轮或三轮选择后,最初在均相fret(荧光共振能量转移)(均相时间分辨荧光,cisbiointernational)测定中筛选来自选择输出的代表性数目的单个克隆与可溶性人α-突触核蛋白的结合作为周质大肠杆菌提取物中的可溶性scfv片段(kipriyanov等jimmunolmethods(1997)200:69-77;其通过提述并入本文)。测定(图1)是均相测定技术,其利用非常接近的供体和受体荧光团之间的荧光共振能量转移(mathisgclinchem(1995)41:1391-1397.;其通过提述并入本文)。此测定可用于通过直接或间接地将感兴趣的分子之一偶联至供体荧光团(如铕(eu3+)穴状化合物)以及将其他感兴趣的分子偶联至受体荧光团xl665(稳定交联的别藻蓝蛋白)来测定大分子相互作用。所述穴状化合物分子的激发(在337nm处)导致620nm处的荧光发射。来自此发射的能量被转移到非常接近所述穴状化合物的xl665,导致来自xl665的特定长寿命荧光(665nm处)的发射。测定所述供体(620nm处)和受体(665nm处)的特定信号,其允许计算665/620nm比率以补偿测定中有色化合物的存在。测试未纯化的抗α-synscfv样品与生物素化的α-syn的结合。将5微升的含有40nm的与0.8nm链霉亲和素铽(cisbiointernational,610satlb)组合的生物素化的人α-syn溶液加入到384孔低体积测定板(costar,3676)中。接下来,将10微升的每种稀释的抗体测试样品加入板中。最后,将5微升含有dc抗myc(cisbiointernational,661mycdab)的溶液加入到测定板中。所有稀释均在于dulbeccopbs(invitrogen,14190185)中含有0.8m氟化钾(bdh103444t)和0.1%牛血清白蛋白(bsa,sigmaa9576)的测定缓冲液中进行。将测定板在室温温育3小时,随后在4℃温育16小时,然后使用envision读板仪(perkinelmer)在620nm和665nm发射波长处读取时间分辨荧光。通过计算665/620nm比率,然后计算每个样品的%δf值来分析数据。665/620nm比率用于校正样品干扰,其使用以下等式:使用以下等式计算每个样品的%δf:通过用非标记的人或大鼠α-syn替代测试样品来定义阴性对照(非特异性结合)。随后,%δf值用于计算%特异性结合,如以下等式中所述:使用graphpadprism软件通过使用四参数逻辑斯谛方程的曲线拟合确定ic50值:y=底部+(顶部-底部)/(1+10^((logec50-x)*hillslope))x是浓度的对数。y是特异性结合。y起始于底部,以s形状进入顶部。将作为未纯化的周质提取物与人α-syn结合的单链fv克隆进行dna测序(osbourn等,immunotechnology(1996),2:181-196;和vaughan等,natbiotechnol.(1996),14(3);309-14;其中二者通过提述并入本文)。在细菌中再次表达和通过亲和层析纯化独特的scfv(如wo01/66754中所述;其通过提述并入本文)。如上所述,通过滴定htrf测定中结合生物素化的人α-syn的纯化的制品来确定这些样品的效价。选择表现出最强α-syn相互作用的纯化scfv制品用于转化为igg形式。通过滴定htrf测定中的抗体,针对克隆结合至α-syn的强度进行排序。进一步在octetred生物传感器上分析最佳的α-syn结合物的结合至作为igg的α-syn的两种动力学(koff)(参见实施例9中所述的方法),以及对突触核蛋白家族成员(人α-syn、β-syn、γ-syn)选择性(参见实施例4中所述的方法)和与鼠α-syn的交叉反应性(参见实施例4中所述的方法)。实施例2:aslo0452ngl-3的衍生物通过在delfia测定(参见实施例4中所述的方法)中筛选与人α-syn的结合来鉴定c末端反应性α-syn特异性克隆(asyn0087)。asyn0087特异性结合至人、食蟹猴和啮齿类α-syn(参见实施例5中所述的方法)。asyn0087回复到最接近合理的人种系序列(tomlinsonvbase.mrccentreofproteinengineering,cambridge,uk.1997;其通过提述并入本文),其在优化之前按照标准突变技术而不影响效价。种系化后,再评估所述克隆与α-syn的结合。没有观察到有害作用。使用如clarkson和lowman(2004)(phagedisplay:apracticalapproach.oxford:oxforduniversitypress;通过提述并入本文)中所述的标准分子生物学技术,通过寡核苷酸定向突变可变重(vh)互补决定区(cdr)2和3以及轻(vl)链cdr1和3来产生源自先导克隆的大的scfv-噬菌体文库。对可溶性生物素化的人α-syn进行基于亲和力的噬菌体展示选择,以选择对人α-syn具有更高亲和力的变体。除了每轮选择进行的降低可溶性生物素化的人α-syn浓度之外,基本上进行如上文先前所述的选择。最初在关于它们与亲本的α-syn结合克隆asyn0087竞争结合可溶性α-syn的能力的htrf测定中筛选来自每次选择输出的代表性克隆作为周质大肠杆菌提取物中的可溶性scfv片段。在这些群体筛选中的每个文库的表现用于报告哪些cdr突变文库在基因方面上加在一起,或“重组”以产生新文库,并且这些重组文库在可溶性生物素化的人α-syn上进行进一轮的亲和力驱动的选择。对于单个和重组突变文库衍生的克隆,在阳性结合物的序列分析后,将所述克隆表达并纯化为scfv片段和igg二者,并通过表位竞争htrf测定再次证实其与可溶性α-syn的结合。通过滴定htrf表位竞争测定中的抗体,与亲本iggasyn0087相比,通过其与α-syn结合的相对改善的ic50值对克隆排序。通过直接结合htrf测定或表位竞争htrf测定进一步分析了最佳的α-syn结合物对突触核蛋白家族成员选择性(人α-syn、β-syn、γ-syn)和与食蟹猴和大鼠α-syn的交叉反应性。从这些迭代轮次的文库重组和筛选中鉴定出两种有效的α-syn特异性的食蟹猴和大鼠α-syn交叉反应性克隆(aslo0452ngl-1和aslo0467)。对aslo0452ngl-1进行针对每个cdr的单个点突变,其中已经观察到ic50效能的正向改善。通过所有20个可能的氨基酸残基单独突变所选cdr中的每个位点,并再次通过表位竞争htrf测定筛选与aslo0452ngl-1相比改善的ic50。确定了四个cdr(h2,h3,l1和l3)上的多个残基并在aslo0452ng1-1和aslo0467上组合,并且再次通过表位竞争htrf测定评估与aslo0452ng1-1相比改善的ic50。从这些试验中,将两种最有改善的结合物鉴定为aslo0452ngl-3和aslo0543。图2比较asyn0087、aslo0452ngl-3和aslo0543的vh和vl区的氨基酸序列。实施例2.1:scfv重构为igg1tm将具有所需的α-syn结合特性的单链fv克隆转化为效应器功能基本上无效的具有以下修饰的全免疫球蛋白g1tm(igg1tm)(oganesyan等actacrystallogrdbiolcrystallogr.(2008),64(pt6):700-4;其通过提述并入本文)抗体形式,如persic等(persic等gene(1997)187:9-18;其通过提述并入本文)所描述的。orip片段包括在表达载体中以促进与cho瞬时细胞一起使用并允许游离型复制。将可变重(vh)域克隆至含有人重链恒定域和调控因子的载体(peu1.4)中以在哺乳动物细胞中表达的完整igg1tm重链。相似地,将可变轻(vh)域克隆至用于表达人轻链(λ)恒定域和调控因子的载体(peu4.4)中以在哺乳动物细胞中表达的完整igg轻链。为了获得igg,将表达重和轻链igg的载体转染至cho瞬时哺乳动物细胞(daramola等,biotechnolprog(2014)30:132-141;通过提述并入本文)中。igg表达并分泌至所述培养基中。在纯化之前过滤收获物,然后使用蛋白a层析法纯化igg。将培养上清液加样至适当大小的陶瓷蛋白a柱(biosepra)上,用50mmtris-hclph8.0、250mmnacl洗涤。用0.1m柠檬酸钠(ph3.0)从该柱上洗脱结合的igg,并通过加入tris-hcl(ph9.0)中和。使用nap10柱(amersham,#17-0854-02)将洗脱的材料缓冲交换到pbs中,并使用基于igg的氨基酸序列的消光系数采用分光光度法测定igg的浓度(mach等,analbiochem(1992)200:74-80;其通过提述并入本文)。使用sec-hplc且通过sds-page分析纯化的igg的聚集和降解纯度。使用igg1tm作为候选药物形式的理由是最小化由于免疫细胞和补体激活的旁邻物杀伤(即可能引起炎症的c3a过量产生)。旁邻细胞杀伤可能是由候选药物和细胞外α-突触核蛋白形成的免疫复合物的潜在积累触发的,其中已证明所述细胞外α-突触核蛋白与脂质膜相互作用(bartels等,biophys.j.(2010),99:2116-2124;其通过提述并入本文)。选择igg1tm形式,因为已经证明其与fcγ受体的结合可忽略不计(fcγr)并且减少免疫复合物的c1q介导的补体激活(oganesyan等actacrystallogrdbiolcrystallogr.(2008),64(pt6):700-4;其通过提述并入本文)。为了使免疫原性的任何潜在风险最小化,aslo0452ngl-3的框架尽可能接近人种系氨基酸序列而不影响效价。这意味着aslo0452ngl-3中的一些氨基酸(包括vernier残基)(foote和winter,jmolbiol.(1992),224(2):487-99;其通过提述并入本文)未突变为最接近的人种系序列。对于aslo0452ngl-3的vh域,v区中存在未突变为人种系igvh3-23和igjh6序列的一个vernier残基(图3c)。对于aslo0452ng1-3的vl域,所有框架残基都与人种系iglv5-45和igjl2或igjl3序列匹配(图3d)。实施例3:亲和力优化的抗α-synigg表位确认从rpeptidellc获得重组人α-、β-和γ-突触核蛋白,人α-突触核蛋白的重组截短形式(aa1-60、aa1-95、aa61-140、96-140、δnac和ncap)和小鼠α-突触核蛋白。使用市售的α-syn截短物进行粗表位作图。简言之,将每毫升1微克的每个截短物在4℃在微量滴定孔中包覆过夜。以pbs冲洗孔后,加入1μg/ml稀释的每种抗α-syn抗体。温育1小时并洗涤后,通过加入与hrp或eu3+偶联的抗人igg检测结合的抗体。随后温育和洗涤,加入适当的检测底物(分别为tmb或delfiaenhancer溶液),并在微量滴定板读数器上读板。这些表位结合研究表明,先导分离物asyn0087识别位于氨基酸102和130之间的α-syn蛋白的c末端区域的表位(图4a)。aslo0452ngl-3和aslo0543二者保持它们对位于氨基酸102和130之间的α-syn蛋白的c末端区域中的与它们亲本先导分离物asyn0087相同的表位的识别(图4b)。实施例4:使用表位竞争htrf测定,相对于突触核蛋白家族成员,aslo0452ngl-3和aslo0543对α-syn的特异性对于旨在用于治疗应用的抗体而言,重要的是对人α-syn具有特异性,以使与脱靶相互作用于其他突触核蛋白(β-突触核蛋白和γ-突触核蛋白)的任何潜在安全性风险最小化。使用测量生物素化的人α-syn与溶液中抗体的结合的htrf表位竞争测定确定aslo0452ngl-3和aslo0543对α-syn而非其他突触核蛋白家族成员(β-syn和γ-syn)的特异性。将α-syn、β-syn和γ-syn滴定到测定中,通过测量生物素化的人α-syn与aslo0452ngl-3/aslo0543的结合的抑制程度来评估aslo0452ng1-3igg和aslo0543igg的选择性。使用软件(graphpad)通过将数据曲线拟合到四参数逻辑斯谛方程来确定ic50值。测量igg与人α-syn、β-syn和γ-syn的直接结合的更灵敏的htrf测定也用于确认α-syn特异性(数据未显示)。对于阴性对照,用同种型对照抗体或仅缓冲液替代抗体测试样品。在aslo0452ng1-3和aslo0543htrf表位竞争测定中用α-syn、β-syn和γ-syn蛋白获得的代表性ic50值在图5中显示。在测试的浓度(高达5μm)下未观察到与β-syn和γ-syn的结合,其证明aslo0452ng1-3和aslo0543对α-syn具有选择性。实施例5:使用htrf表位竞争测定法,aslo0452ng1-3对人、食蟹猴和大鼠α-syn的的特异性鉴于治疗应用,重要的是所述抗体与食蟹猴α-突触核蛋白交叉反应并且期望它与大鼠α-突触核蛋白交叉反应,达到对人α-syn所观察到的10倍内。这是为了能够在食蟹猴和大鼠物种中进行安全性研究。使用测量生物素化的人α-syn与溶液中的as0452ng1-3的结合的htrf表位竞争测定法确定aslo0452ng1-3和aslo0543对于人、食蟹猴和大鼠α-syn的特异性。将人、食蟹猴和大鼠α-syn滴定到所述测定中,并通过测量生物素化的人α-syn与所述抗体结合的抑制程度来评估所述抗体的选择性。使用软件(graphpad)通过将数据曲线拟合到四参数逻辑斯谛方程来确定ic50值。使用直接结合htrf测定形式(未显示)也证实了aslo0452ng1-3和aslo0543的物种交叉反应性。将aslo0452ng1-3(或aslo0543)滴定至所述测定中以通过htrf测定与aslo0452ng1-3(或aslo0543)竞争结合至人或食蟹猴或大鼠α-syn。对于阴性对照,用同种型对照抗体或仅缓冲液替代抗体测试样品。在htrf表位竞争测定中获得的关于人、食蟹猴和大鼠α-syn蛋白的代表性ic50值在图6中显示。aslo0452ngl-3分别以5.7nm和6.8nm的ic50值与人和食蟹猴α-syn结合,以19.6nm的ic50值与大鼠α-syn结合,在4倍以内。aslo0543分别以2.0nm和2.1nm的ic50值与人和食蟹猴α-syn结合,以3.8nm的ic50值与大鼠α-syn结合,在2倍以内。aslo0452ngl-3与人、食蟹猴和大鼠α-syn结合的能力表明:相较于不与人和食蟹猴和大鼠α-syn结合的抗体,与人α-突触核蛋白上的不同表位结合。实施例6:通过流式细胞术测量的、亲和力优化的克隆对天然α-syn的特异性使用α-syn阳性和阴性细胞系通过流式细胞术测定亲和力优化的抗α-synigg(aslo0452ng1-3和aslo0543)与天然内源性人α-syn结合的特异性。简言之,将shsy5y成神经细胞瘤细胞(α-syn阳性)和bt-20乳腺癌细胞(α-syn阴性)固定在0.01%甲醛中,然后用0.5%(v/v)吐温20透化,然后与抗α-syn抗体、阳性对照或同种型对照抗体温育。在大量洗涤后,通过与抗人或抗小鼠igg-fitc第二抗体温育来检测结合的抗体。进一步洗涤后,用facscantoii装置(bectondickinson,franklinlakes,nj)分析所述细胞,并使用flowjo软件(treestar,ashland,or)进行数据分析。将数据绘制为柱状图,其显示单独染色的细胞与第一抗体染色的细胞之间的差异。图7a中的结果显示,与同种型对照和单独的第二抗体(组b)相比,在α-syn阳性sh-sy5y细胞中存在asyn0087(图d)时荧光信号的迁移,这表明内源表达的α-syn的识别。asyn0087不与α-syn阴性人乳腺癌细胞系bt-20结合(组c)。图7b中的结果显示与阳性对照抗体(α-syn阳性sh-sy5y细胞上的4d6)相比(图f),在存在aslo0452ng1-3或aslo0543(图h)的情况下荧光信号的强烈迁移,并且在α-syn阴性bt-20细胞(组g)上没有迁移。这证明aslo0452ng1-3和aslo0543都与天然内源性表达的细胞内人α-syn结合。实施例7:通过delfiaelisa,优化的抗α-synigg对聚集的人α-syn的特异性如emadi等所述(emadi等,biochemistry(2004),43:2871-2878.;其通过提述并入本文)的产生人α-syn的纤维状制品或聚集体。简言之,将200μl的50μm重组α-syn等分到1.8mlsarstedt管中,并在37℃振荡培养箱中以280rpm放置3天。通过掺入添加至最终浓度为10μm的硫磺素t,在室温在黑暗中温育1小时并在450nm的激发波长和485nm的发射波长下在envision酶标仪上读取荧光来确定聚集的α-syn的存在。使用抗体捕获测定法测定亲和力优化的抗α-synigg(aslo0452ng1-3和aslo0543)和先导抗体asyn0087对聚集的人α-syn的特异性。该测定法在成对elisa中通过aslo0452ng1-3,aslo0543或asyn0087测量聚集的人α-syn的捕获。简言之,将抗α-syn抗体的小鼠igg1形式固定在96孔微量滴定板(nunc)的孔上。封闭后,在所述孔中温育聚集的或单体的人α-syn。洗涤后,通过加入相同抗α-syn抗体的人igg1tm形式且随后加入抗人igg-europium缀合物(perkinelmer)或抗人igg-hrp缀合物来检测捕获的人α-syn。温育和洗涤后,加入适当的检测底物(分别为tmb或delfiaenhancer溶液),并在微量滴定板读数器上读板。在该测定中,仅捕获并检测聚集的人α-syn,因为其在单个聚集体上呈现相同表位的多个拷贝。对于单体α-syn,仅存在一个表位拷贝,因此检测第二抗体在捕获抗体存在下不能结合。数据总结在图8中。先导分离物asyn0087能够与聚集的重组人α-syn结合。因此,asyn0087结合的表位本身不参与α-syn的聚集。aslo0452ngl-3和aslo0543二者保留着它们与聚集的重组人α-syn结合的能力。实施例8:通过免疫组织化学,优化的抗α-synigg在疾病相关组织中的特异性通过帕金森病脑组织的免疫组织化学染色确定亲和力优化的抗α-synigg(aslo0452ng1-3和aslo0543)以及先导抗体asyn0087对人α-syn的疾病相关形式的特异性。结果如图9所示,并证明与asyn0087一样,aslo0452ngl-3和aslo0543均可识别帕金森病脑组织切片中人α-syn的疾病相关病理形式(包括路易体、路易神经突、神经元聚集体、lewy斑点和背景脑组织)。在正常或健康的脑组织中未观察到非特异性染色。实施例9:抗α-syn抗体亲和力测定使用两种平台技术测定针对人α-syn的抗α-synigg的平衡解离常数(kd):octetred(fortebio)和kinexa(sapidyneinstruments)。两种测定系统都表现出良好的一致性,其表明aslo0452ngl-3亲和力在亚纳摩尔范围内。表1显示了在整个先导分离和先导优化过程中产生的关键抗α-syn克隆的亲和力测量结果。实施例9.1:通过octet测定aslo0452ngl-3的亲和力使用octetred仪器估计aslo0452ng1-3igg对重组细菌表达的单体人avi-tagα-syn-flag-his的亲和力。将aslo0452ngl-3与不同浓度的每种配体预混合直至达到平衡。然后使用octet通过使用固定在链霉抗生物素蛋白包覆的传感器上的生物素化的α-syn捕获游离的aslo0452ng1-3来测量游离抗体的量。将每个α-syn浓度下检测到的游离抗体的量相对于配体浓度作图,并使用kinexa软件计算平衡解离常数(kd)。表1中显示的结果表明,aslo0452ngl-3igg以106pm的亲和力结合人α-syn。实施例9.2:通过kinexa测定aslo0452ngl-3的亲和力此外,使用kinexa仪器(sapidyneinstruments)测定aslo0452ngl-3igg对细菌重组表达的单体人生物素化α-syn的溶液相亲和力(kd)。将aslo0452ngl-3与不同浓度的每种配体预混合直至达到平衡。然后使用kinexa通过使用α-syn包覆的珠子捕获游离的aslo0452ng1-3,洗去未结合的物质并使用荧光标记的物种特异性抗体检测结合的抗体来测定游离抗体的量。将每个α-syn浓度下检测到的游离抗体的量相对于配体浓度作图,并使用kinexa软件计算平衡解离常数(kd)。表1中显示的结果表明aslo0452ngl-3igg以74pm的亲和力结合至α-syn,其显示出与上述octet溶液相亲和力测定的良好一致性。实施例9.3:通过kinexa测定aslo0452ngl-3fab片段的亲和力aslo0452ng1-3fab片段以高亲和力结合α-突触核蛋白。通过kinexa分析(如上文实施例中针对完整抗体所述的)测量的aslo0452ng1-3fab片段对α-突触核蛋白的kd值为174pm(95%ci:15-177pm)。实施例10:aslo0452ngl-3对雄性spraguedawley大鼠前额叶皮质组织间液(isf)游离未结合的α-突触核蛋白水平的影响将成年雄性spraguedawley大鼠(293-417g;harlan,荷兰)麻醉并将导向器植入前额皮质中。在实验前一天,使用立体定位框架将推拉探针(1-3mda聚乙烯膜4mm)植入前额叶皮质中(探针的坐标:ap=-3.4mm(相对前囟),侧向+0.8mm(相对中线),腹侧-5.0mm(相对硬脑膜)),将切牙条设定为-3.3mm(所有坐标根据paxinos和watson,theratbraininstereotaxiccoordinates,academicpress,newyork,第6版2008)。用不锈钢螺钉和牙科粘固剂将探针附接到头骨上。在实验当天,将推拉微透析探针用柔性peek管(westernanalyticalproductsinc.usa;pk005-020)连接至微灌注泵(harvard),并用含有147mmnacl、3.0mmkcl、1.2mmcacl2和1.2mmmgcl2+0.2%bsa的人工csf(灌注液)以0.5μl/min的流速灌注。将探头的出口连接到柔性fep管。在至少2小时的预稳定化后,在pbs中配制的aslo0452ngl-3(或仅pbs(运载体))分别以30或0mg/kg给药。以2ml/kg静脉内施用所述化合物。以120分钟的间隔收集微透析样品。将样品收集到小瓶中(microbiotech/seab,sweden;4001029)。所有样品均储存在-80℃。为了测定大鼠isf中的游离α-突触核蛋白浓度,首先对所述微透析样品进行免疫沉淀以除去aslo0452ngl-3。免疫沉淀共沉淀与治疗性抗体结合的α-突触核蛋白,而未结合的“游离”α-突触核蛋白保留在上清液中。将蛋白a珠子(蛋白a)的溶液加入到96孔非裙边板(聚丙烯0.2ml)中,并使用磁体(dynamagtm96侧)用tbst(50mmtbs加0.1%吐温20)洗涤两次,以将珠子与溶液分离。将解冻的大鼠isf微透析样品(10或20μl)加入到每个孔中,通过上下吸移与珠子混合,并在4℃以倾斜旋转温育10分钟。然后使用磁铁将所述珠子制粒两次以确保完全除去所述珠子。将免疫沉淀的isf样品从抗α-突触核蛋白elisa试剂盒(sensolytetm定量elisa试剂盒,人/小鼠/大鼠,anaspec,us,as-55550)转移到含已经添加至总体积为100μl的样品稀释缓冲液的96孔板中。将每孔100μl的校准样品一式两份加入板中,并向每个孔中加入50μl检测抗体工作溶液。将该板在+4-8℃温育过夜,同时摇动并避光,然后用350μl洗涤缓冲液洗涤六次。最后,向每个孔中加入100μltmb颜色底物,并将该板在室温在黑暗中温育10-15分钟。为了终止该反应,向每个孔中加入50μl终止溶液,并在2小时内在450nm的吸光度下读板。通过绘制作为线性标度上的吸光度单位对对数标度的浓度的标准曲线的响应来进行定量。四参数函数用于曲线拟合。在单次静脉内施用30mg/kg的aslo0452ngl-3后,证实了isf中游离α-突触核蛋白的时间依赖性降低(图10)。实施例11:aslo0452ngl-3对雄性spraguedawley大鼠脑脊髓液游离未结合α-突触核蛋白水平的影响将成年雄性spraguedawley大鼠麻醉并将导管置于小脑延髓池中以供应csf取样。将0.8cm留置套管插入小脑延髓池,并通过头骨顶部的切口进行外露。用牙科丙烯酸粘合剂将csf导管的末端固定在适当位置,并用三个不锈钢螺钉将其固定在头骨上。在使用所述化合物之前使动物至少恢复2天。将aslo0452ng-3配制在缓冲液中,以3、10、30或100mg/kg给药。仅以2ml/kg静脉内施用所述化合物或仅运载体。在收集在至少两天内取出的至少四个洁净csf样品后,施用所述化合物。所有动物在第0天给药aslo0452ng-3或运载体。在每个指定的时间点收集csf样品。将所有样品储存在-80℃直至装运。为了测定csf中的游离α-突触核蛋白,在分析之前通过免疫沉淀(ip)除去与aslo0452ng1-3结合的α-突触核蛋白。ip将共同沉淀与治疗性抗体结合的α-突触核蛋白,而未结合的“游离”α-突触核蛋白保留在上清液中。使用从anaspec获得的商业elisa试剂盒测定上清液中α-突触核蛋白的游离水平。如isf结果所述进行该分析(如实施例10中所述)。在3-100mg/kg的剂量范围内单次静脉内施用aslo0452ngl-3后,证实了csf中游离α-突触核蛋白的剂量和时间依赖性降低(图11)。实施例12:通过减少α-突触核蛋白病的慢病毒体内模型中的α-突触核蛋白扩散的aslo0452ngl-3的功能表征使用α-突触核蛋白病的慢病毒体内小鼠模型研究了高亲和力抗α-突触核蛋白抗体aslo0452ng1-3阻断α-突触核蛋白扩散的能力。为此,将表达α-突触核蛋白(lv-α-syn)的慢病毒载体注射到非转基因野生型小鼠(非tg)和过表达α-突触核蛋白的转基因小鼠(α-syntg)的右侧海马体中,然后用抗-α突触核蛋白小鼠igg1抗体(包括aslo0452ngl-3)和同种型对照小鼠igg1nip228每周被动免疫持续13周。在免疫期结束时,对所述小鼠实施安乐死并将其脑在4%pfa中固定,然后进行冠状切片并通过具有自动图像分析的免疫细胞化学来分析lv-α-syn注射位点的同侧和对侧的α-突触核蛋白免疫反应性水平。手术和被动免疫三到四个月大的非转基因野生型小鼠(非tg;n=40)和α-突触核蛋白转基因小鼠(α-syntg;n=40)接受单次单侧注射表达α-突触核蛋白(lv-α-syn)的慢病毒载体到右侧海马(前囟的-2.0、1.5、1.3)。在lv-α-syn注射手术后两周,小鼠接受每周剂量的抗α-突触核蛋白小鼠igg1抗体:aslo0452ngl-3(非tgn=10;α-syntgn=10)、aslo0452ngl-3d265a(非tgn=10;α-syntgn=10)、9e4(非tgn=10;α-syntgn=10),或者给药它们nip228同种型对照小鼠igg1(非tgn=10;α-syntgn=10)。所有小鼠igg通过腹膜内(ip)途径以20mg/kg给药13周。将动物分组饲养,每笼最多或4只。将动物保持在12/12光/暗循环,随意获取食物和水。笼子每周更换一次并每天监测。报告任何不良事件。所有动物都耐受手术治疗以及免疫接种。在抗体治疗期结束时,按照动物的人道处理指南对小鼠实施安乐死,并将它们的大脑在冠状轴上连续切片,并通过免疫细胞化学方法评估α-突触核蛋白扩散的神经病理学分析。α-突触核蛋白免疫细胞化学取出脑,在4%多聚甲醛中固定,并使用振动切片机在冠状轴上以40μm间隔切割切片,并在-30℃储存在冷冻保护介质(30%甘油、30%乙二醇、40%pbs)中。在pbs洗涤和封闭缓冲液步骤后,将切片在4℃与第一抗体(以1:500稀释的来自bd的抗-α-突触核蛋白mabsyn-1)温育过夜,在pbs中洗涤并在室温与第二抗体温育1小时(以1:100稀释的来自vectorlaboratories的生物素化的抗小鼠igg)。在最后的pbs洗涤步骤后,使用抗生物素蛋白/生物素-过氧化物酶复合物检测系统(eliteabc,vectorlaboratories)定位染色的α-突触核蛋白。然后用自动图像分析分析切片的慢病毒载体(lv-α-syn)注射位点的同侧和对侧的α-突触核蛋白水平。统计使用graphpadprism软件(sandiegocalifornia,usa)分析从非tg和α-syntg处理组的α-突触核蛋白水平的自动图像采集产生的数据。使用dunnett's多重比较后检验进行单向anova。附图中所示的数据表示为平均值±标准误(sem)。当p<0.05时认为组间的差异具有统计学意义。所有分析均对评估者不知情。所述抗体治疗组也对评估者不知情。结果在非tg小鼠和α-syntg小鼠中,注射lv-α-syn的同侧位点上的α-突触核蛋白免疫反应性在神经纤维网中强烈,且覆盖在海马体的大部分表面(图12a、图15a;nip228-同侧)。非tg小鼠和α-syntg小鼠的对侧非注射的海马体也表现出高水平的α-突触核蛋白免疫反应性,其表明慢病毒表达的α-突触核蛋白已经从注射的右侧海马体扩散到左侧海马体(图12a、图15a;nip228-对侧)。在这种慢病毒α-突触核蛋白注射小鼠模型中,先前的实验已经表明,只有表达的α-突触核蛋白扩散到对侧位点,没有慢病毒本身转移的证据,如通过pcr分析所确定的(数据未显示)。用9e4抗体(9e4–prx002(prothena)的小鼠形式)被动免疫的非tg小鼠中慢病毒表达的α-突触核蛋白的同侧和对侧海马体水平与用nip228同种型对照小鼠igg1被动免疫的非tg小鼠中慢病毒表达的α-突触核蛋白的同侧和对侧海马体水平几乎相同(图12a、b、c;9e4与nip228比较),其指示当9e4经由ip途径以每周20mg/kg给药13周时,9e4不会阻断这种α-突触核蛋白扩散模型中的α-突触核蛋白的传播。相反地,用aslo0452ngl-3抗体或aslo0452ngl-3(aslo0452-ngl-3-d265a)的效应器无效突变体形式(其中在小鼠igg1中的位点265处的天冬氨酸由丙氨酸取代(d265a)导致在这种同种型和在小神经胶质细胞中发现的低亲和力的iggfc受体(fcγriib和fcγriii)之间相互作用的缺失)被动免疫的非tg小鼠中慢病毒表达的α-突触核蛋白的同侧和对侧海马体水平显著低于用nip228同种型对照小鼠igg1被动免疫的非tg小鼠中慢病毒表达的α-突触核蛋白的同侧和对侧海马体水平(图12a、b、c;aslo0452-ngl-3和aslo0452-ngl-3-d265a与nip228比较)。这指示用aslo0452-ngl-3或aslo0452-ngl-3的效应器无效的d265a突变体形式被动免疫的小鼠强有效地阻断这种α-突触核蛋白病的小鼠模型中α-突触核蛋白的扩散。当将lv-α-syn载体注射到α-syntg小鼠的右侧海马体中时,获得了相似的结果;当与nip228处理的α-syntg小鼠比较时,aslo0452-ngl-3或aslo0452-ngl-3-d265a而非9e4的被动免疫导致了海马体中α-突触核蛋白免疫反应性的同侧和对侧水平的稳固的且统计学上显著的减少(图15a、b、c)。在更高的放大倍数下,沿注射位点的同侧轴突和非tg小鼠的非注射位点的对侧轴突可观察到慢病毒表达的α-突触核蛋白免疫反应性沉积物(图13a;黑色箭头显示跨海马体的轴突),其表明α-突触核蛋白扩散至对侧海马体可以大部分沿着轴突发生(跨轴突扩散)。用9e4抗体或nip228同种型对照小鼠igg1被动免疫的非tg小鼠中的轴突α-突触核蛋白沉积物的同侧和对侧水平不是显著差异的(图13a、b、c;9e4与nip228比较),这指示在我们的实验条件下,9e4不影响轴突α-突触核蛋白沉积物的水平以及不减少这种慢病毒α-突触核蛋白病扩散模型中α-突触核蛋白沿轴突的传播。相反地,用aslo0452ngl-3抗体或aslo0452-ngl-3的效应器无效突变体形式(aslo0452-ngl-3-d265a)被动免疫的非tg小鼠中的轴突α-突触核蛋白沉积物的同侧和对侧二者水平显著低于用nip228同种型对照小鼠igg1处理的非tg小鼠中的轴突α-突触核蛋白沉积物的水平(图13a、b、c;aslo0452-ngl-3和aslo0452-ngl-3-d265a与nip228比较),这指示用aslo0452ngl-3抗体或aslo0452-ngl-3的效应器无效突变体形式被动免疫小鼠清除了轴突α-突触核蛋白沉积物并稳固地阻断了这种慢病毒α-突触核蛋白病小鼠模型中α-突触核蛋白沿轴突的同侧至对侧的转移。当将lv-α-syn载体注射到α-syntg小鼠的右侧海马体中时,获得了非常相似的结果;当与nip228处理的α-syntg小鼠比较时,aslo0452-ngl-3或aslo0452-ngl-3-d265a而非9e4的被动免疫导致沿轴突的α-突触核蛋白免疫反应性的同侧和对侧水平的稳固的且统计学上显著的降低(图16a、b、c)。在lv-α-syn注射的非tg小鼠中,在更高的放大倍数下,在同侧海马的神经纤维网中检测到强烈的α-突触核蛋白免疫反应性,并且在同侧新皮质的神经纤维中检测到较小程度的α-突触核蛋白免疫反应性(图14a)。此外,在同侧海马体的ca1区域的许多可识别的神经元细胞体(体细胞)中检测到α-突触核蛋白的严重沉积,并且在同侧新皮层的第5层神经元中检测到较弱的α-突触核蛋白免疫反应性(图14;黑色箭头)。虽然当与用nip228同种型对照小鼠igg免疫的非tg小鼠相比,用9e4抗体的处理不显著改变含有α-突触核蛋白的同侧ca1海马体神经元或同侧第5层新皮质神经元的数量(图14a、b、c;9e4与nip228比较),但与用nip228同种型对照小鼠igg免疫的非tg小鼠相比,用aslo0452ngl-3抗体或aslo0452-ngl-3的效应器无效突变体形式(aslo0452-ngl-3-d265a)处理的非tg小鼠已显著减少了含有α-突触核蛋白沉积物的同侧ca1神经元和同侧第5层神经元的数量(图14a、b、c;aslo0452-ngl-3和aslo0452-ngl-3-d265a与nip228比较)。另外,当与nip228同种型对照小鼠igg处理的非tg小鼠比较时,在aslo0452-ngl-3处理和aslo0452-ngl-3-d265a处理的非tg中,神经纤维网和海马体的同侧ca1区域的神经元和新皮质的同侧第5层区域的α-突触核蛋白免疫反应性强度是显著更低的(图14a;aslo0452-ngl-3和aslo0452-ngl-3-d265a与nip228比较)。当将lv-α-syn载体注射到α-syntg小鼠的右侧海马体中时,获得了相似的结果。当与nip228处理的α-syntg小鼠相比,用aslo0452-ngl-3或aslo0452-ngl-3-d265a而非9e4的处理导致了同侧ca1海马体和第5层新皮质区域中以及对侧ca1海马体区域中的含有强α-突触核蛋白免疫反应性的神经元数量的显著降低(图17a、b、c、d)。已经证明了用高亲和力抗α-突触核蛋白小鼠igg1抗体aslo0452-ngl-3被动免疫均已立体定向注射慢病毒载体(促使海马体一侧的α-突触核蛋白的表达)的非tg野生型小鼠或α-syntg小鼠稳固地减少了在α-突触核蛋白繁殖的这种小鼠模型中观察到的慢病毒表达的α-突触核蛋白的同侧至对侧的跨轴突的扩散。相较于不抑制测试模型中扩散的抗体(例如9e4抗体),这种新公开的抗α-突触核蛋白抗体的抑制体内α-突触核蛋白扩散的特性指示与人α-突触核蛋白的不同表位结合。此外,显示aslo0452-ngl-3的效应器无效d265a突变体形式与aslo0452-ngl-3在减少模型中α-突触核蛋白扩散方面同样有效的数据指示α-突触核蛋白扩散的抗体介导的阻止不需要fc相关效应器功能作为关键作用机制,且特别地,它指示在抗体介导的α-突触核蛋白扩散阻断中,存在于小胶质神经细胞上的fc受体(fcγriib和fcγriii)似乎不需要或不作用。总之,靶向α-突触核蛋白的本发明的抗体及其抗原结合片段通过阻断或减缓α-突触核蛋白病理摄取至受体细胞中并防止解剖学上连接的脑区域之间的α-突触核蛋白病理的播种(seeding)和传播在pd或dlb或msa中具有疾病改善的潜力。用此方法,靶向α-突触核蛋白的抗体可治疗或防止疾病进展并对患有突触核蛋白病的患者具有治疗益处。实施例13:0452ngl-3-bbbt0626gl双特异性抗体的生成如下文所述生成移接到aslo0452ngl-3的重链的n末端或c末端的包含与bbbt0626gl的单链片段相关联的人igg1tm骨架的根据本发明的示例性双特异性抗体。通过合成产生编码含有bsshii和bsteii侧翼核酸内切酶限制性位点、各自的bbbt0626glscfv或bbbt0626wt的上游和aslo0452ngl-3vh的下游的bbbt0626glscfv-(g4s)x2-aslo0452ngl-3vh或bbbt0626wt-(g4s)x2-aslo452ngl-3vh的dna片段生成bis2形式的根据本发明的双特异性抗体。然后,将消化的dna片段直接克隆至higg1tm载体骨架中。通过pcr扩增,随后使用限制性核酸内切酶位点(sfii和xbai)通过直接克隆产生bis3形式的根据本发明的双特异性抗体。生成下列两个pcr片段:(1)扩增从sfii限制性位点至ch3域的c末端的higg1tm-ch3域和(2)重叠pcr片段,其具有掺入ch3-(g4s)x3接头的c-末端(seqidno:57)和bbbt0626g1scfv的n-末端的正向pcr寡聚物连同扩增bbbt0626glscfv的c-末端的寡聚物和经过xbai限制性位点的紧邻下游载体序列。两个pcr片段在拉通式(pull-through)pcr反应中连接在一起,并随后经由sfii和xbai限制性核酸内切酶位点定向克隆入peu1_4(人igg1tm载体)。这些双特异性抗体在基于cho的表达系统中表达,并且经由蛋白a柱纯化所得的抗体。测试所有bbbt0626衍生的双特异性抗体与小鼠脑内皮细胞系(b.end3)的体外结合,以证实bbb转运蛋白部分的结合活性,并且还在基于htrf表位竞争测定中测试了结合also0452ngl-3的竞争,以证实与aslo0452ngl-3表位的结合。表1克隆id方法亲和力-kd95%ciaslo0087octet﹥1.0μmn/aaslo0452ngl-1octet1.1μm0.3-2.3nmaslo0452ngl-3kinexa74pm15-177nmaslo0452ngl-3octet106pm10-292nmaslo0543kinexa108pm34-223nmaslo0543octet113pm5-333nm序列表<110>麦迪穆有限责任公司(medimmunelimited)<120>针对α-突触核蛋白的抗体及其用途<130>1848081-0002-091-wo1<140><141><150>62/344,746<151>2016-06-02<160>61<170>patentin3.5版<210>1<211>140<212>prt<213>人(homosapiens)<400>1metaspvalphemetlysglyleuserlysalalysgluglyvalval151015alaalaalaglulysthrlysglnglyvalalaglualaalaglylys202530thrlysgluglyvalleutyrvalglyserlysthrlysgluglyval354045valhisglyvalalathrvalalaglulysthrlysgluglnvalthr505560asnvalglyglyalavalvalthrglyvalthralavalalaglnlys65707580thrvalgluglyalaglyserilealaalaalathrglyphevallys859095lysaspglnleuglylysasnglugluglyalaproglngluglyile100105110leugluaspmetprovalaspproaspasnglualatyrglumetpro115120125serglugluglytyrglnasptyrgluprogluala130135140<210>2<211>122<212>prt<213>人(homosapiens)<400>2gluvalglnleuleugluserglyglyglyleuvalglnproglygly151015serleuargleusercysalaalaserglyphethrphesersertyr202530alametsertrpvalargglnalaproglylysglyleuglutrpval354045seralaileserglyserglyglyserthrtyrtyralaaspserval505560lysglyargphethrileserargaspasnserlysasnthrleutyr65707580leuglnmetasnserleuargalagluaspthralavaltyrtyrcys859095alaglyglyalavalasnvaltyrtyrtyrtyrglymetaspvaltrp100105110glyglnglythrmetvalthrvalserser115120<210>3<211>115<212>prt<213>人(homosapiens)<400>3glnalavalleuthrglnproserserleuseralaserproglyala151015seralaserleuthrcysthrleuargserglyasnasnvalglyasn202530tyrargiletyrtrptyrglnglnlysserglyserproproglntyr354045leuleuargtyrlysseraspalaasplyshisglnglyserglyval505560proserargpheserglyserlysaspalaseralaasnalaglyile65707580leupheileserglyleuglnsergluaspglualaasptyrtyrcys859095metvaltrphisserglyalatrpvalpheglyglyglythrlysleu100105110thrvalleu115<210>4<211>122<212>prt<213>人(homosapiens)<220><221>变体<222>(50)..(50)<223>/取代="ala"<220><221>变体<222>(53)..(53)<223>/取代="gly"<220><221>变体<222>(54)..(54)<223>/取代="ser"<220><221>变体<222>(101)..(101)<223>/取代="arg"<220><221>变体<222>(102)..(102)<223>/取代="arg"<220><221>变体<222>(103)..(103)<223>/取代="gly"<220><221>变体<222>(104)..(104)<223>/取代="arg"<220><221>变体<222>(105)..(105)<223>/取代="ile"<220><221>misc_特征<222>(1)..(122)<223>/注释="序列中给出的变体残基对于变体位点的注释中的变体残基没有偏好"<400>4gluvalglnleuleugluserglyglyglyleuvalglnproglygly151015serleuargleusercysalaalaserglyphethrphesersertyr202530alametsertrpvalargglnalaproglylysglyleuglutrpval354045serserileserhisleuglyglyserthrtyrtyralaaspserval505560lysglyargphethrileserargaspasnserlysasnthrleutyr65707580leuglnmetasnserleuargalagluaspthralavaltyrtyrcys859095alaglyglyalaasnhisvallystyrtyrtyrglymetaspvaltrp100105110glyglnglythrmetvalthrvalserser115120<210>5<211>5<212>prt<213>人(homosapiens)<400>5sertyralametser15<210>6<211>17<212>prt<213>人(homosapiens)<220><221>变体<222>(1)..(1)<223>/取代="ala"<220><221>变体<222>(4)..(4)<223>/取代="gly"<220><221>变体<222>(5)..(5)<223>/取代="ser"<220><221>misc_特征<222>(1)..(17)<223>/注释="序列中给出的变体残基对于变体位点的注释中的变体残基没有偏好"<400>6serileserhisleuglyglyserthrtyrtyralaaspservallys151015gly<210>7<211>13<212>prt<213>人(homosapiens)<220><221>变体<222>(3)..(3)<223>/取代="arg"<220><221>变体<222>(4)..(4)<223>/取代="arg"<220><221>变体<222>(5)..(5)<223>/取代="gly"<220><221>变体<222>(6)..(6)<223>/取代="arg"<220><221>变体<222>(7)..(7)<223>/取代="ile"<220><221>misc_特征<222>(1)..(13)<223>/注释="序列中给出的变体残基对于变体位点的注释中的变体残基没有偏好"<400>7glyalaasnhisvallystyrtyrtyrglymetaspval1510<210>8<211>114<212>prt<213>人(homosapiens)<220><221>变体<222>(27)..(27)<223>/取代="ser"<220><221>变体<222>(28)..(28)<223>/取代="gly"<220><221>变体<222>(29)..(29)<223>/取代="asp"<220><221>变体<222>(30)..(30)<223>/取代="phe"<220><221>变体<222>(31)..(31)<223>/取代="ser"<220><221>变体<222>(32)..(32)<223>/取代="arg"<220><221>变体<222>(42)..(42)<223>/取代="pro"<220><221>变体<222>(82)..(82)<223>/取代="leu"<220><221>变体<222>(99)..(99)<223>/取代="ser"<220><221>变体<222>(100)..(100)<223>/取代="ser"<220><221>变体<222>(102)..(102)<223>/取代="ala"<220><221>变体<222>(104)..(104)<223>/取代="tyr"<220><221>misc_特征<222>(1)..(114)<223>/注释="序列中给出的变体残基对于变体位点的注释中的变体残基没有偏好"<400>8glnalavalleuthrglnproserserleuseralaserproglyala151015seralaserleuthrcysthrleuargserglyalaproleuprolys202530tyrargiletyrtrptyrglnglnlysserglyserproproglntyr354045leuleuargtyrlysseraspalaasplyshisglnglyserglyval505560proserargpheserglyserlysaspalaseralaasnalaglyile65707580leupheserglyleuglnsergluaspglualaasptyrtyrcysmet859095valtrpasphisglyvaltrpvalpheglyglyglythrlysleuthr100105110valleu<210>9<211>14<212>prt<213>人(homosapiens)<220><221>变体<222>(5)..(5)<223>/取代="ser"<220><221>变体<222>(6)..(6)<223>/取代="gly"<220><221>变体<222>(7)..(7)<223>/取代="asp"<220><221>变体<222>(8)..(8)<223>/取代="phe"<220><221>变体<222>(9)..(9)<223>/取代="ser"<220><221>变体<222>(10)..(10)<223>/取代="arg"<220><221>misc_特征<222>(1)..(14)<223>/注释="序列中给出的变体残基对于变体位点的注释中的变体残基没有偏好"<400>9thrleuargserglyalaproleuprolystyrargiletyr1510<210>10<211>11<212>prt<213>人(homosapiens)<400>10tyrlysseraspalaasplyshisglnglyser1510<210>11<211>9<212>prt<213>人(homosapiens)<220><221>变体<222>(4)..(4)<223>/取代="ser"<220><221>变体<222>(5)..(5)<223>/取代="ser"<220><221>变体<222>(7)..(7)<223>/取代="ala"<220><221>变体<222>(9)..(9)<223>/取代="tyr"<220><221>misc_特征<222>(1)..(9)<223>/注释="序列中给出的变体残基对于变体位点的注释中的变体残基没有偏好"<400>11metvaltrpasphisglyvaltrpval15<210>12<211>452<212>prt<213>人(homosapiens)<400>12gluvalglnleuleugluserglyglyglyleuvalglnproglygly151015serleuargleusercysalaalaserglyphethrphesersertyr202530alametsertrpvalargglnalaproglylysglyleuglutrpval354045serserileserhisleuglyglyserthrtyrtyralaaspserval505560lysglyargphethrileserargaspasnserlysasnthrleutyr65707580leuglnmetasnserleuargalagluaspthralavaltyrtyrcys859095alaglyglyalaasnhisglylystyrtyrtyrglymetasplystrp100105110glyglnglythrthrvalthrvalserseralaserthrlysglypro115120125servalpheproleualaproserserlysserthrserglyglythr130135140alaalaleuglycysleuvallysasptyrpheprogluprovalthr145150155160valsertrpasnserglyalaleuthrserglyvalhisthrphepro165170175alavalleuglnserserglyleutyrserleuserservalvalthr180185190valproserserserleuglythrglnthrtyrilecysasnvalasn195200205hislysproserasnthrlysvalasplysargvalgluprolysser210215220cysasplysthrhisthrcysproprocysproalaproglupheglu225230235240glyglyproservalpheleupheproprolysprolysaspthrleu245250255metileserargthrprogluvalthrcysvalvalvalaspvalser260265270hisgluaspprogluvallyspheasntrptyrvalaspglyvalglu275280285valhisasnalalysthrlysproargglugluglntyrasnserthr290295300tyrargvalvalservalleuthrvalleuhisglnasptrpleuasn305310315320glylysglutyrlyscyslysvalserasnlysalaleuproalaser325330335ileglulysthrileserlysalalysglyglnproarggluprogln340345350valtyrthrleuproproserarggluglumetthrlysasnglnval355360365serleuthrcysleuvallysglyphetyrproseraspilealaval370375380glutrpgluserasnglyglnprogluasnasntyrlysthrthrpro385390395400provalleuaspseraspglyserphepheleutyrserlysleuthr405410415valasplysserargtrpglnglnglyasnvalphesercysserval420425430methisglualaleuhisasnhistyrthrglnlysserleuserleu435440445serproglylys450<210>13<211>366<212>dna<213>人(homosapiens)<220><221>cds<222>(1)..(366)<400>13gaggtgcagctgttggagtctgggggaggcttggtacagcctgggggg48gluvalglnleuleugluserglyglyglyleuvalglnproglygly151015tccctgagactctcctgtgcagcctctggattcacctttagcagctat96serleuargleusercysalaalaserglyphethrphesersertyr202530gccatgagctgggtccgccaggctccagggaaggggctggagtgggtc144alametsertrpvalargglnalaproglylysglyleuglutrpval354045tcatccatttcccaccttggtggtagcacatactacgcagactccgtg192serserileserhisleuglyglyserthrtyrtyralaaspserval505560aagggccggttcaccatctccagagacaattccaagaacacgctgtat240lysglyargphethrileserargaspasnserlysasnthrleutyr65707580ctgcaaatgaacagcctgagagccgaggacacggccgtgtattactgt288leuglnmetasnserleuargalagluaspthralavaltyrtyrcys859095gcgggaggggcaaaccacgggaagtactactacggaatggacaagtgg336alaglyglyalaasnhisglylystyrtyrtyrglymetasplystrp100105110ggccaagggaccacggtcaccgtctcctca366glyglnglythrthrvalthrvalserser115120<210>14<211>122<212>prt<213>人(homosapiens)<400>14gluvalglnleuleugluserglyglyglyleuvalglnproglygly151015serleuargleusercysalaalaserglyphethrphesersertyr202530alametsertrpvalargglnalaproglylysglyleuglutrpval354045serserileserhisleuglyglyserthrtyrtyralaaspserval505560lysglyargphethrileserargaspasnserlysasnthrleutyr65707580leuglnmetasnserleuargalagluaspthralavaltyrtyrcys859095alaglyglyalaasnhisglylystyrtyrtyrglymetasplystrp100105110glyglnglythrthrvalthrvalserser115120<210>15<211>17<212>prt<213>人(homosapiens)<400>15serileserhisleuglyglyserthrtyrtyralaaspservallys151015gly<210>16<211>13<212>prt<213>人(homosapiens)<400>16glyalaasnhisglylystyrtyrtyrglymetasplys1510<210>17<211>221<212>prt<213>人(homosapiens)<400>17glnalavalleuthrglnproalaserleuseralaserproglyala151015seralaserleuthrcysthrleuargserglyalaproleuprolys202530tyrargiletyrtrptyrglnglnlysproglyserproproglntyr354045leuleuargtyrlysseraspalaasplyshisglnglyserglyval505560proserargpheserglyserlysaspalaseralaasnalaglyile65707580leuleuileserglyleuglnsergluaspglualaasptyrtyrcys859095metvaltrpasphisglyvaltrptyrpheglyglyglythrlysleu100105110thrvalleuglyglnprolysalaalaproservalthrleuphepro115120125proserserglugluleuglnalaasnlysalathrleuvalcysleu130135140ileseraspphetyrproglyalavalthrvalalatrplysalaasp145150155160serserprovallysalaglyvalgluthrthrthrproserlysgln165170175serasnasnlystyralaalasersertyrleuserleuthrproglu180185190glntrplysserhisargsertyrsercysglnvalthrhisglugly195200205serthrvalglulysthrvalalaprothrglucysser210215220<210>18<211>345<212>dna<213>人(homosapiens)<400>18caggctgtgctgactcagccggcttccctctctgcgtctcctggagcatcagccagtctc60acctgcaccttgcgcagtggggcgcccctgccgaagtataggatatactggtatcagcag120aagccagggagtcctccccagtatctcctgaggtacaaatcagacgcagataaacaccag180ggctctggagtccccagccgcttttctggatccaaagatgcttcggccaatgcagggatt240ttactcatctctgggctccagtctgaggatgaggctgactattattgtatggtttgggac300cacggcgtctggtatttcggcggagggaccaagctgaccgtccta345<210>19<211>115<212>prt<213>人(homosapiens)<400>19glnalavalleuthrglnproalaserleuseralaserproglyala151015seralaserleuthrcysthrleuargserglyalaproleuprolys202530tyrargiletyrtrptyrglnglnlysproglyserproproglntyr354045leuleuargtyrlysseraspalaasplyshisglnglyserglyval505560proserargpheserglyserlysaspalaseralaasnalaglyile65707580leuleuileserglyleuglnsergluaspglualaasptyrtyrcys859095metvaltrpasphisglyvaltrptyrpheglyglyglythrlysleu100105110thrvalleu115<210>20<211>14<212>prt<213>人(homosapiens)<400>20thrleuargserglyalaproleuprolystyrargiletyr1510<210>21<211>9<212>prt<213>人(homosapiens)<400>21metvaltrpasphisglyvaltrptyr15<210>22<211>452<212>prt<213>人(homosapiens)<400>22gluvalglnleuleugluserglyglyglyleuvalglnproglygly151015serleuargleusercysalaalaserglyphethrphesersertyr202530alametsertrpvalargglnalaproglylysglyleuglutrpval354045seralaileserglyserglyglyserthrtyrtyralaaspserval505560lysglyargphethrileserargaspasnserlysasnthrleutyr65707580leuglnmetasnserleuargalagluaspthralavaltyrtyrcys859095alaglyglyalaargargglyargiletyrtyrglymetasplystrp100105110glyglnglythrthrvalthrvalserseralaserthrlysglypro115120125servalpheproleualaproserserlysserthrserglyglythr130135140alaalaleuglycysleuvallysasptyrpheprogluprovalthr145150155160valsertrpasnserglyalaleuthrserglyvalhisthrphepro165170175alavalleuglnserserglyleutyrserleuserservalvalthr180185190valproserserserleuglythrglnthrtyrilecysasnvalasn195200205hislysproserasnthrlysvalasplysargvalgluprolysser210215220cysasplysthrhisthrcysproprocysproalaproglupheglu225230235240glyglyproservalpheleupheproprolysprolysaspthrleu245250255metileserargthrprogluvalthrcysvalvalvalaspvalser260265270hisgluaspprogluvallyspheasntrptyrvalaspglyvalglu275280285valhisasnalalysthrlysproargglugluglntyrasnserthr290295300tyrargvalvalservalleuthrvalleuhisglnasptrpleuasn305310315320glylysglutyrlyscyslysvalserasnlysalaleuproalaser325330335ileglulysthrileserlysalalysglyglnproarggluprogln340345350valtyrthrleuproproserarggluglumetthrlysasnglnval355360365serleuthrcysleuvallysglyphetyrproseraspilealaval370375380glutrpgluserasnglyglnprogluasnasntyrlysthrthrpro385390395400provalleuaspseraspglyserphepheleutyrserlysleuthr405410415valasplysserargtrpglnglnglyasnvalphesercysserval420425430methisglualaleuhisasnhistyrthrglnlysserleuserleu435440445serproglylys450<210>23<211>366<212>dna<213>人(homosapiens)<400>23gaggtgcagctgttggagtctgggggaggcttggtacagcctggggggtccctgagactc60tcctgtgcagcctctggattcacctttagcagctatgccatgagctgggtccgccaggct120ccagggaaggggctggagtgggtctcagctattagtggtagtggtggtagcacatactac180gcagactccgtgaagggccggttcaccatctccagagacaattccaagaacacgctgtat240ctgcaaatgaacagcctgagagccgaggacacggccgtgtattactgtgcgggaggggca300cggcgcgggcgcatctactacggaatggacaaatggggccaagggacaacggtcaccgtc360tcctca366<210>24<211>122<212>prt<213>人(homosapiens)<400>24gluvalglnleuleugluserglyglyglyleuvalglnproglygly151015serleuargleusercysalaalaserglyphethrphesersertyr202530alametsertrpvalargglnalaproglylysglyleuglutrpval354045seralaileserglyserglyglyserthrtyrtyralaaspserval505560lysglyargphethrileserargaspasnserlysasnthrleutyr65707580leuglnmetasnserleuargalagluaspthralavaltyrtyrcys859095alaglyglyalaargargglyargiletyrtyrglymetasplystrp100105110glyglnglythrthrvalthrvalserser115120<210>25<211>5<212>prt<213>人(homosapiens)<400>25sertyralametser15<210>26<211>17<212>prt<213>人(homosapiens)<400>26alaileserglyserglyglyserthrtyrtyralaaspservallys151015gly<210>27<211>13<212>prt<213>人(homosapiens)<400>27glyalaargargglyargiletyrtyrglymetasplys1510<210>28<211>221<212>prt<213>人(homosapiens)<400>28glnalavalleuthrglnproalaserleuseralaserproglyala151015seralaserleuthrcysthrleuargserserglyasppheserarg202530tyrargiletyrtrptyrglnglnlysproglyserproproglntyr354045leuleuargtyrlysseraspalaasplyshisglnglyserglyval505560proserargpheserglyserlysaspalaseralaasnalaglyile65707580leuleuileserglyleuglnsergluaspglualaasptyrtyrcys859095metvaltrpserserglyalatrptyrpheglyglyglythrlysleu100105110thrvalleuglyglnprolysalaalaproservalthrleuphepro115120125proserserglugluleuglnalaasnlysalathrleuvalcysleu130135140ileseraspphetyrproglyalavalthrvalalatrplysalaasp145150155160serserprovallysalaglyvalgluthrthrthrproserlysgln165170175serasnasnlystyralaalasersertyrleuserleuthrproglu180185190glntrplysserhisargsertyrsercysglnvalthrhisglugly195200205serthrvalglulysthrvalalaprothrglucysser210215220<210>29<211>345<212>dna<213>人(homosapiens)<400>29caggctgtgctgactcagccggcttccctctctgcgtctcctggagcatcagccagtctc60acctgcaccttgcgcagttccggggacttctcccggtataggatatactggtatcagcag120aagccagggagtcctccccagtatctcctgaggtacaaatcagacgcagataaacaccag180ggctctggagtccccagccgcttttctggatccaaagatgcttcggccaatgcagggatt240ttactcatctctgggctccagtctgaggatgaggctgactattattgtatggtttggtcc300agcggcgcttggtacttcggcggagggaccaagctgaccgtccta345<210>30<211>115<212>prt<213>人(homosapiens)<400>30glnalavalleuthrglnproalaserleuseralaserproglyala151015seralaserleuthrcysthrleuargserserglyasppheserarg202530tyrargiletyrtrptyrglnglnlysproglyserproproglntyr354045leuleuargtyrlysseraspalaasplyshisglnglyserglyval505560proserargpheserglyserlysaspalaseralaasnalaglyile65707580leuleuileserglyleuglnsergluaspglualaasptyrtyrcys859095metvaltrpserserglyalatrptyrpheglyglyglythrlysleu100105110thrvalleu115<210>31<211>14<212>prt<213>人(homosapiens)<400>31thrleuargserserglyasppheserargtyrargiletyr1510<210>32<211>11<212>prt<213>人(homosapiens)<400>32tyrlysseraspalaasplyshisglnglyser1510<210>33<211>9<212>prt<213>人(homosapiens)<400>33metvaltrpserserglyalatrptyr15<210>34<211>123<212>prt<213>人(homosapiens)<220><221>变体<222>(1)..(1)<223>/取代="val"<220><221>变体<222>(11)..(11)<223>/取代="leu"<220><221>变体<222>(75)..(75)<223>/取代="arg"<220><221>misc_特征<222>(1)..(123)<223>/注释="序列中给出的变体残基对于变体位点的注释中的变体残基没有偏好"<400>34glnvalglnleuvalglnserglyalagluvallyslysproglyser151015servallysvalsercyslysalaserglyglythrphesersertyr202530alailesertrpvalargglnalaproglyglnglyleuglutrpmet354045glyargileileproileleuglythrserasntyralaglnlysphe505560glnglyargvalthrilethralaaspgluserthrserthralatyr65707580metgluleuserserleuargsergluaspthralavaltyrtyrcys859095alaargargserserleualaalaalaaspargglyalapheaspile100105110trpglyglnglythrmetvalthrvalserser115120<210>35<211>108<212>prt<213>人(homosapiens)<220><221>变体<222>(15)..(15)<223>/取代="gly"or"ala"<220><221>变体<222>(28)..(28)<223>/取代="arg"<220><221>变体<222>(29)..(29)<223>/取代="arg"or"thr"<220><221>变体<222>(32)..(32)<223>/取代="thr"<220><221>变体<222>(33)..(33)<223>/取代="ser"<220><221>变体<222>(37)..(37)<223>/取代="gln"<220><221>变体<222>(44)..(44)<223>/取代="ile"<220><221>变体<222>(47)..(47)<223>/取代="ile"<220><221>变体<222>(50)..(50)<223>/取代="glu"<220><221>变体<222>(51)..(51)<223>/取代="asp"<220><221>变体<222>(57)..(57)<223>/取代="ile"<220><221>变体<222>(65)..(65)<223>/取代="arg"<220><221>变体<222>(68)..(68)<223>/取代="thr"<220><221>变体<222>(92)..(92)<223>/取代="asn"<220><221>变体<222>(93)..(93)<223>/取代="thr"<220><221>变体<222>(95)..(95)<223>/取代="lys"or"his"<220><221>变体<222>(96)..(96)<223>/取代="pro"<220><221>变体<222>(97)..(97)<223>/取代="trp"<220><221>misc_特征<222>(1)..(108)<223>/注释="序列中给出的变体残基对于变体位点的注释中的变体残基没有偏好"<400>35sersergluleuthrglnaspproalavalservalalaleuarggln151015thrvalargilethrcysglnglyaspserleuthrsertyrtyrala202530asntrptyrglnhislysproglyglnalaprovalleuvalmettyr354045glylysasnasnargproserglyvalproaspargpheserglyser505560serserglyasnthralaserleuthrilethrglyalaglnalaglu65707580aspglualaasptyrtyrcysasnserargaspserserglyasnhis859095valvalpheglyglyglythrlysleuthrvalleu100105<210>36<211>11<212>prt<213>人(homosapiens)<220><221>变体<222>(6)..(6)<223>/取代="arg"<220><221>变体<222>(7)..(7)<223>/取代="arg"或"thr"<220><221>变体<222>(10)..(10)<223>/取代="thr"<220><221>变体<222>(11)..(11)<223>/取代="ser"<220><221>misc_特征<222>(1)..(11)<223>/注释="序列中给出的变体残基对于变体位点的注释中的变体残基没有偏好"<400>36glnglyaspserleuthrsertyrtyralaasn1510<210>37<211>7<212>prt<213>人(homosapiens)<220><221>变体<222>(2)..(2)<223>/取代="glu"<220><221>变体<222>(3)..(3)<223>/取代="asp"<220><221>misc_特征<222>(1)..(7)<223>/注释="序列中给出的变体残基对于变体位点的注释中的变体残基没有偏好"<400>37glylysasnasnargproser15<210>38<211>11<212>prt<213>人(homosapiens)<220><221>变体<222>(5)..(5)<223>/取代="asn"<220><221>变体<222>(6)..(6)<223>/取代="thr"<220><221>变体<222>(8)..(8)<223>/取代="lys"或"his"<220><221>变体<222>(9)..(9)<223>/取代="pro"<220><221>变体<222>(10)..(10)<223>/取代="trp"<220><221>misc_特征<222>(1)..(11)<223>/注释="序列中给出的变体残基对于变体位点的注释中的变体残基没有偏好"<400>38asnserargaspserserglyasnhisvalval1510<210>39<211>123<212>prt<213>人(homosapiens)<400>39glnvalglnleuvalglnserglyalagluvallyslysproglyser151015servallysvalsercyslysalaserglyglythrphesersertyr202530alailesertrpvalargglnalaproglyglnglyleuglutrpmet354045glyargileileproileleuglythrserasntyralaglnlysphe505560glnglyargvalthrilethralaaspgluserthrserthralatyr65707580metgluleuserserleuargsergluaspthralavaltyrtyrcys859095alaargargserserleualaalaalaaspargglyalapheaspile100105110trpglyglnglythrmetvalthrvalserser115120<210>40<211>5<212>prt<213>人(homosapiens)<400>40sertyralaileser15<210>41<211>17<212>prt<213>人(homosapiens)<400>41argileileproileleuglythrserasntyralaglnlysphegln151015gly<210>42<211>14<212>prt<213>人(homosapiens)<400>42argserserleualaalaalaaspargglyalapheaspile1510<210>43<211>108<212>prt<213>人(homosapiens)<400>43sersergluleuthrglnaspproalavalservalalaleuglygln151015thrvalargilethrcysglnglyaspserleuargsertyrtyrala202530sertrptyrglnglnlysproglyglnalaprovalleuvaliletyr354045glylysasnasnargproserglyileproaspargpheserglyser505560serserglyasnthralaserleuthrilethrglyalaglnalaglu65707580aspglualaasptyrtyrcysasnserargaspserserglyasnhis859095valvalpheglyglyglythrlysleuthrvalleu100105<210>44<211>11<212>prt<213>人(homosapiens)<400>44glnglyaspserleuargsertyrtyralaser1510<210>45<211>7<212>prt<213>人(homosapiens)<400>45glylysasnasnargproser15<210>46<211>11<212>prt<213>人(homosapiens)<400>46asnserargaspserserglyasnhisvalval1510<210>47<211>123<212>prt<213>人(homosapiens)<220><221>来源<223>/注释="bbbt0626vh"<400>47glyvalglnleuvalglnserglyalagluleulyslysproglyser151015servallysvalsercyslysalaserglyglythrphesersertyr202530alailesertrpvalargglnalaproglyglnglyleuglutrpmet354045glyargileileproileleuglythrserasntyralaglnlysphe505560glnglyargvalthrilethralaaspgluargthrserthralatyr65707580metgluleuserserleuargsergluaspthralavaltyrtyrcys859095alaargargserserleualaalaalaaspargglyalapheaspile100105110trpglyglnglythrmetvalthrvalserser115120<210>48<211>122<212>prt<213>人(homosapiens)<400>48glnvalglnleuvalglnserglyalagluvallyslysproglyser151015servallysvalsercyslysalaserglyglythrpheglythrtyr202530serilethrtrpvalargglnalaproglyglnglyleuglutrpmet354045glyaspilevalproilepheglythrproasntyralaglnasnphe505560glnglyargvalthrileseralaaspvalserthrserthrvaltyr65707580metgluleuserserleuargsergluaspthralavaltyrtyrcys859095alalysargglysertyrtyrglyargglyglytrppheaspprotrp100105110glyargglythrleuvalthrvalserser115120<210>49<211>5<212>prt<213>人(homosapiens)<400>49thrtyrserilethr15<210>50<211>17<212>prt<213>人(homosapiens)<400>50aspilevalproilepheglythrproasntyralaglnasnphegln151015gly<210>51<211>13<212>prt<213>人(homosapiens)<400>51argglysertyrtyrglyargglyglytrppheasppro1510<210>52<211>122<212>prt<213>人(homosapiens)<400>52glnvalglnleuvalglnserglyalagluvallyslysproglyser151015servallysvalsercyslysalaserglyglythrpheglythrtyr202530serilethrtrpvalargglnalaproglyglnglyleuglutrpmet354045glyaspilevalproilepheglythrproasntyralaglnasnphe505560glnglyargvalthrileseralaaspvalserthrserthrvaltyr65707580metgluleuserserleuargsergluaspthralavaltyrtyrcys859095alalysargglysertyrtyrglyargglyglytrppheaspprotrp100105110glyargglythrleuvalthrvalserser115120<210>53<211>5<212>prt<213>人(homosapiens)<400>53thrtyrserilethr15<210>54<211>17<212>prt<213>人(homosapiens)<400>54aspilevalproilepheglythrproasntyralaglnasnphegln151015gly<210>55<211>13<212>prt<213>人(homosapiens)<400>55argglysertyrtyrglyargglyglytrppheasppro1510<210>56<211>50<212>prt<213>人工序列<220><221>来源<223>/注释="人工序列的描述:合成的多肽"<220><221>misc_特征<222>(1)..(50)<223>/注释="此序列包含1-10'glyglyglyglyser'重复单元"<220><221>来源<223>/注释="参见对于替换和优选实施方案的详细描述而提交的说明书"<400>56glyglyglyglyserglyglyglyglyserglyglyglyglysergly151015glyglyglyserglyglyglyglyserglyglyglyglyserglygly202530glyglyserglyglyglyglyserglyglyglyglyserglyglygly354045glyser50<210>57<211>15<212>prt<213>人工序列<220><221>来源<223>/注释="人工序列的描述:合成的肽"<400>57glyglyglyglyserglyglyglyglyserglyglyglyglyser151015<210>58<211>98<212>prt<213>人(homosapiens)<400>58gluvalglnleuleugluserglyglyglyleuvalglnproglygly151015serleuargleusercysalaalaserglyphethrphesersertyr202530alametsertrpvalargglnalaproglylysglyleuglutrpval354045seralaileserglyserglyglyserthrtyrtyralaaspserval505560lysglyargphethrileserargaspasnserlysasnthrleutyr65707580leuglnmetasnserleuargalagluaspthralavaltyrtyrcys859095alalys<210>59<211>11<212>prt<213>人(homosapiens)<400>59trpglyglnglythrthrvalthrvalserser1510<210>60<211>102<212>prt<213>人(homosapiens)<400>60glnalavalleuthrglnproalaserleuseralaserproglyala151015seralaserleuthrcysthrleuargserglyileasnvalglythr202530tyrargiletyrtrptyrglnglnlysproglyserproproglntyr354045leuleuargtyrlysseraspserasplysglnglnglyserglyval505560proserargpheserglyserlysaspalaseralaasnalaglyile65707580leuleuileserglyleuglnsergluaspglualaasptyrtyrcys859095metiletrphistrpval100<210>61<211>10<212>prt<213>人(homosapiens)<400>61pheglyglyglythrlysleuthrvalleu1510当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1