一种预防或治疗骨关节炎的方法和药物与流程
本发明涉及通过使用纤溶酶原预防或治疗骨关节炎的方法和药物。
背景技术:
骨关节炎,又称骨性关节炎、退行性关节炎、退行性关节病、骨关节病,其是最常见的关节炎形式,影响约2.37亿(3.3%)的人口,也是关节疼痛甚至残疾的主导原因之一[1],在60岁以上的人中,约有10%的男性和18%的女性受到影响[2]。骨关节炎是一种多因素引起的,以关节软骨进行性分解为特征的异质性疾病,这些因素包括创伤、异常的机械负荷、营养供应不足和遗传诱因等,还有代谢因素和髌下脂肪垫。过去的研究关注异常的生物力学对软骨下骨、关节软骨完整性和软骨细胞病理生理的影响;最近的证据表明,骨关节炎的临床症状不仅影响关节软骨,而且影响多种关节组织的完整性,包括滑膜、骨、韧带、支持的肌肉和半月板等[3]。
骨关节炎的根本原因并非是骨骼发生了病变,现代医学研究发现,患病的根本原因在于软骨等“关节保护系统”对关节保护能力的丧失。具体表现在:各类骨关节疾病的发生往往始于滑膜病变、软骨受损或是变性;服用某些抗炎类、激素类药物所造成的软骨损伤也是许多骨关节疾病的主要成因之一。由于关节滑膜、软骨的损伤以及关节滑液的缺失,导致关节骨骼缺少必要的保护,以至于人体活动时,关节处的骨骼因缺乏必要的“软骨保护”直接发生剧烈硬性摩擦,而引发患者关节疼痛、肿胀、变形、骨刺增生等多种症状[4]。因此,修复受损关节软骨、滑膜并促进软骨、滑膜的再生能力,催生关节滑液,从而恢复关节器官的“软骨保护层”是治疗骨关节疾病的关键所在。本研究证明纤溶酶原可以改善骨关节的结构完整性,增强关节局部成骨细胞活性和/或降低破骨细胞的活性、促进软骨再生和软骨下骨骨重建和有效缓解疼痛等,为包括骨关节炎在内的关节损伤疾病的治疗开辟了新的途径。
技术实现要素:
发明概述
本发明涉及通过使用纤溶酶原预防或治疗骨关节病的方法和药物。本发明证明纤溶酶原可以改善骨关节的结构完整性,增强关节局部成骨细胞活性和/或降低破骨细胞的活性、促进软骨再生和软骨下骨骨重建,并可以有效缓解疼痛等症状,为骨关节疾病的治疗开辟了新的途径。
具体地,本发明涉及:
1.一种治疗骨关节炎的方法,包括给药受试者有效量的纤溶酶原。
2.项1的方法,所述纤溶酶原增加关节软骨的量和/或促进关节软骨损伤修复。
3.项1或2的方法,所述纤溶酶原改善关节滑膜炎症状况。
4.项1-3任一项的方法,所述纤溶酶原促进关节的软骨下骨骨重建。
5.项1-4任一项的方法,所述纤溶酶原改善关节的炎症状况、疼痛和/或改善关节功能。
6.项1-5任一项的方法,其中所述纤溶酶原减轻关节肿胀和疼痛。
7.一种促进骨关节炎受试者关节软骨再生的方法,包括给药受试者有效量的纤溶酶原。
8.一种促进受试者关节损伤修复的方法,包括给药受试者有效量的纤溶酶原。
9.项8的方法,所述纤溶酶原促进关节软骨再生和/或软骨下骨骨重建。
10.项8或9的方法,其中所述受试者为骨关节炎受试者。
11.项8-10任一项的方法,其中所述纤溶酶原改善关节组织的炎症状况和/或减轻关节疼痛。
12.项1-11任一项的方法,其中所述纤溶酶原与序列2、6、8、10或12具有至少75%、80%、85%、90%、95%、96%、97%、98%或99%的序列同一性,并且仍然具有纤溶酶原活性。
13.项1-11任一项的方法,所述纤溶酶原是包含纤溶酶原活性片段、并且仍然具有纤溶酶原活性的蛋白质。
14.项1-11任一项的方法,所述纤溶酶原选自glu-纤溶酶原、lys-纤溶酶原、小纤溶酶原、微纤溶酶原、delta-纤溶酶原或它们的保留纤溶酶原活性的变体。
15.项1-11任一项的方法,所述纤溶酶原为天然或合成的人纤溶酶原、或其仍然保留纤溶酶原活性的变体或片段。
16.项1-11任一项的方法,所述纤溶酶原为来自灵长类动物或啮齿类动物的人纤溶酶原直向同系物或其仍然保留纤溶酶原活性的变体或片段。
17.项1-11任一项的方法,所述纤溶酶原的氨基酸如序列2、6、8、10或12所示。
18.项1-11任一项的方法,其中所述纤溶酶原是人天然纤溶酶原。
19.项1-18任一项的方法,其中所述受试者是人。
20.项1-19任一项的方法,其中所述受试者缺乏或缺失纤溶酶原。
21.项1-20任一项的方法,其中所述纤溶酶原与一种或多种关节损伤治疗药物或方法、或一种或多种骨关节炎治疗药物或方法联合应用。
22.一种用于项1-21任一项的方法的纤溶酶原。
23.一种药物组合物,其包含药学上可接受的载剂和用于项1-21中任一项所述方法的纤溶酶原。
24.一种预防性或治疗性试剂盒,其包含:(i)用于项1-21中任一项所述方法的纤溶酶原和(ii)用于递送所述纤溶酶原至所述受试者的构件(means)。
25.根据项24所述的试剂盒,其中所述构件为注射器或小瓶。
26.项24或25的试剂盒,其还包含标签或使用说明书,该标签或使用说明书指示将所述纤溶酶原投予所述受试者以实施项1-21中任一项所述方法。
27.一种制品,其包含:
含有标签的容器;和
包含(i)用于项1-21中任一项所述方法的纤溶酶原或包含纤溶酶原的药物组合物,其中所述标签指示将所述纤溶酶原或组合物投予所述受试者以实施项1-21中任一项所述方法。
28.项24-26中任一项的试剂盒或项27的制品,还包含另外的一个或多个构件或容器,该构件或容器中含有一种或多种其它的关节损伤治疗药物或用品、或一种或多种其它的骨关节炎治疗药物或用品。
29.项24-26中任一项的试剂盒或项27的制品,还包括与骨关节炎并发的其它疾病的药物。
30.包含纤溶酶原的用于治疗骨关节炎的药剂。
31.包含纤溶酶原的用于治疗关节损伤的药剂。
32.包含纤溶酶原的用于治疗骨关节炎的药物组合物,试剂盒、制品。
33.包含纤溶酶原的用于治疗关节损伤的药物组合物,试剂盒、制品。
34.纤溶酶原在制备治疗骨关节炎的药物、制品或试剂盒中的用途。
35.项34的用途,所述纤溶酶原增加关节软骨的量和/或促进关节软骨损伤修复。
36.项34或35的用途,所述纤溶酶原改善关节滑膜炎症状况。
37.项34-36任一项的用途,所述纤溶酶原促进关节的软骨下骨骨重建。
38.项34-37任一项的用途,所述纤溶酶原改善关节的炎症状况、疼痛和/或改善关节功能。
39.项34-38任一项的用途,其中所述纤溶酶原减轻关节肿胀和疼痛。
40.纤溶酶原在制备促进骨关节炎受试者关节软骨再生的药物中的用途。
41.纤溶酶原在制备促进受试者关节损伤修复的药物中的用途。
42.项41的用途,所述纤溶酶原促进关节软骨再生和/或软骨下骨骨重建。
43.项41或42的用途,其中所述受试者为骨关节炎受试者。
44.项41-43任一项的用途,其中所述纤溶酶原改善关节组织的炎症状况和/或减轻关节疼痛。
45.项34-44任一项的方法,其中所述纤溶酶原与序列2、6、8、10或12具有至少75%、80%、85%、90%、95%、96%、97%、98%或99%的序列同一性,并且仍然具有纤溶酶原活性。
46.项34-44任一项的用途,所述纤溶酶原是包含纤溶酶原活性片段、并且仍然具有纤溶酶原活性的蛋白质。
47.项34-44任一项的用途,所述纤溶酶原选自glu-纤溶酶原、lys-纤溶酶原、小纤溶酶原、微纤溶酶原、delta-纤溶酶原或它们的保留纤溶酶原活性的变体。
48.项34-44任一项的用途,所述纤溶酶原为天然或合成的人纤溶酶原、或其仍然保留纤溶酶原活性的变体或片段。
49.项34-44任一项的用途,所述纤溶酶原为来自灵长类动物或啮齿类动物的人纤溶酶原直向同系物或其仍然保留纤溶酶原活性的变体或片段。
50.项34-44任一项的用途,所述纤溶酶原的氨基酸如序列2、6、8、10或12所示。
51.项34-44任一项的用途,其中所述纤溶酶原是人天然纤溶酶原。
52.项34-51任一项的用途,其中所述受试者是人。
53.项34-52任一项的用途,其中所述纤溶酶原与一种或多种关节损伤治疗药物或用途、或一种或多种骨关节炎治疗药物或方法联合应用。
发明详述
术语的定义和技术方案的描述
“关节”为骨之间的连接组织,该连接组织中有腔隙,能做不同程度的活动。关节主要的结构有关节囊、关节腔、关节软骨、滑液,以及韧带、关节周围的肌肉和肌腱、滑膜皱襞、滑囊、半月板、软骨下骨等。关节囊是包绕关节相对的两个骨端的结缔组织膜囊。它分为内外两层:外层为纤维层,由致密结缔组织构成,厚而坚韧,主要起着维持关节的牢固性及稳定性作用;内层为滑膜层,薄而柔软,由血管丰富的疏松结缔组织构成,覆盖在纤维层的内面形成环绕滑膜腔的囊。关节腔是由关节囊与关节面围成的组织腔隙。正常情况下,关节腔内会有少许粘稠的液体(即滑液),具有润滑和营养关节的作用。关节软骨是指覆盖在关节表面的那层很薄的软骨。它覆盖于关节面上,具有很好的弹性,从而有减少摩擦、缓冲震荡与冲击力的作用。滑液即关节液,由滑膜所分泌。正常滑液清亮、粘稠。当关节有炎症时,滑液量明显增多,关节腔内压力增高,因而产生局部肿痛。
本发明的一些实施方案涉及纤溶酶原治疗关节损伤的方法和促进关节损伤受试者关节损伤修复的方法,包括给药受试者有效量的纤溶酶原。在一些实施方案中,所述的关节损伤包括炎症、退行性病变、代谢障碍或创伤引起的关节损伤。在一些实施方案中,所述纤溶酶原通过促进关节软骨再生和/或软骨下骨骨重建治疗关节损伤,或促进关节损伤的修复。在一些实施方案中,纤溶酶原改善关节损伤受试者的关节组织的炎症和/或减轻关节疼痛。在一些实施方案中,所述关节组织的炎症包括关节滑膜的炎症。在一些实施方案中,所述纤溶酶原减轻关节损伤受试者关节肿胀和疼痛。
在一些实施方案中,本发明涉及促进关节软骨再生的方法,包括给药受试者有效量的纤溶酶原。在一些实施方案中,本发明涉及纤溶酶原通过促进软骨再生促进关节损伤的修复和治疗关节损伤疾病。
在上述纤溶酶原治疗关节损伤、促进关节损伤修复的一些实施方案中,所述关节损伤为骨关节炎导致的关节损伤。
在上述纤溶酶原治疗关节损伤、促进关节损伤修复的一些实施方案中,所述纤溶酶原通过促进关节软骨再生和/或软骨下骨骨重建,使关节损伤得以修复,由关节损伤发展成骨关节炎的过程得以截断。因此,本发明也涉及纤溶酶原用于预防骨关节炎的方法,包括对骨关节损伤的受试者施用有效量的纤溶酶原。
在上述所有纤溶酶原治疗关节损伤、促进关节损伤修复、促进软骨再生、预防关节损伤发展为骨关节炎的方法中,所述纤溶酶原可以单独施用,也可以与一种或多种其它药物或方法联合施用。所述联合施用包括在一种或多种其它药物或方法施用之前,同时或之后施用纤溶酶原。
“骨关节炎(osteoarthritis,oa)”为一种退行性病变,是由于增龄、肥胖、劳损、创伤、关节先天性异常、关节畸形等诸多因素引起的关节软骨退化损伤、关节边缘和软骨下骨反应性增生,又称骨关节病、退行性关节炎、老年性关节炎等。临床表现为缓慢发展的关节疼痛、压痛、僵硬、关节肿胀、活动受限和关节畸形等。
骨关节炎是骨关节的退行性病变,好发于负重关节及活动量较多的关节,例如,颈椎、腰椎、膝关节、髋关节等。过度负重或使用这些关节,均可促进退行性变化的发生。临床表现为缓慢发展的关节疼痛、压痛、僵硬、关节肿胀、活动受限和关节畸形等。影像学改变包括关节软骨及软骨下骨质的异常改变,关节间隙狭窄(此表明关节软骨已开始变薄),骨质钙化,关节边缘变尖,骨赘形成和/或软骨下骨性囊腔形成。本发明所述的“骨关节炎”涵盖身体各部位发生的骨关节炎,包括颈椎、腰椎、膝关节、髋关节部位的骨关节炎或相关退行性病变。
本发明的一些实施方案涉及纤溶酶原治疗骨关节炎的方法,包括给药受试者有效量的纤溶酶原。在一些实施方案中,所述纤溶酶原促进关节软骨再生和/或软骨下骨骨重建。在一些实施方案中,纤溶酶原改善关节组织的炎症状况和/或减轻关节肿胀和/或疼痛。在一些实施方案中,纤溶酶原改善关节滑膜的炎症和/或减轻关节肿胀和/或疼痛。在一些实施方案中,所述骨关节炎为颈椎、腰椎、膝关节、髋关节部位的骨关节炎或相关退行性病变。
在上述纤溶酶原治疗骨关节炎的实施方案中,所述“治疗”包括减轻、缓解、改善或消除如下的一个或多种症状或体征:关节疼痛、压痛、僵硬、关节肿胀、关节功能障碍或受限、关节软骨或软骨下骨质异常改变、关节间隙狭窄,骨质钙化、关节边缘变尖、骨赘、软骨下骨性囊腔。在所述的纤溶酶原治疗骨关节炎的实施方案中,所述骨关节炎包括颈椎、腰椎、膝关节、髋关节的骨关节炎及其相关退行性病变。
“骨重建”也称“骨重塑”,指成熟的骨组织从骨骼中移除(称为骨吸收的过程)并且形成新的骨组织(称为骨化或新骨形成的过程)。这些过程还控制骨折等损伤后骨骼的再成形或置换,以及在正常活动期间发生的微损伤。重塑也响应机械负载的功能需求。骨骼的结构以及钙的充足供应需要成骨细胞(分泌新骨)和破骨细胞(破骨)二种细胞类型与骨重建部位存在的其它细胞群(例如免疫细胞等)的密切配合,并依赖于复杂的信号传导途径和控制机制,实现适当的生长和分化速率。
在一些实施方案中,本发明涉及纤溶酶原促进关节软骨再生和/或软骨下骨骨重建。在一些实施方案中,本发明涉及纤溶酶原促进(增强)骨关节炎受试者骨关节炎患病部位成骨细胞的活性和/或降低破骨细胞的活性。
在一些实施方案中,本发明涉及纤溶酶原促进受试者关节损伤修复的方法,包括给药受试者有效量的纤溶酶原。在一些实施方案中,所述的关节损伤包括炎症、退行性病变、代谢障碍或创伤引起的关节损伤。在一些实施方案中,所述纤溶酶原通过促进关节软骨再生和/或软骨下骨骨重建治疗关节损伤,或促进关节损伤的修复。在一些实施方案中,所述纤溶酶原促进(增强)关节损伤受试者关节损伤部位成骨细胞的活性和/或降低破骨细胞的活性。在一些实施方案中,所述关节损伤为骨关节炎导致的关节损伤。在一些实施方案中,所述的关节损伤为炎症、代谢障碍或创伤引起的关节损伤。
“改善”关节滑膜炎症(状况)、“改善”关节的炎症(状况)、“改善”关节组织的炎症(状况)或“改善”关节组织的炎症是指炎症或炎症状况经过给药纤溶酶原治疗得到改善,炎症反应最终缓解、消退。
在上述所有纤溶酶原治疗骨关节炎的实施方案中,所述纤溶酶原可以单独施用,也可以与一种或多种其它药物或方法联合施用。所述联合施用包括在一种或多种其它药物或方法施用之前,同时或之后施用纤溶酶原。
纤溶酶是纤溶酶原激活系统(pa系统)的关键组分。它是一种广谱的蛋白酶,能够水解细胞外基质(ecm)的几个组分,包括纤维蛋白、明胶、纤连蛋白、层粘连蛋白和蛋白聚糖[5]。此外,纤溶酶能将一些金属蛋白酶前体(pro-mmps)激活形成具有活性的金属蛋白酶(mmps)。因此纤溶酶被认为是胞外蛋白水解作用的一个重要的上游调节物[6,7]。纤溶酶是由纤溶酶原通过两种生理性的pas:组织型纤溶酶原激活剂(tpa)或尿激酶型纤溶酶原激活剂(upa)蛋白水解形成的。由于纤溶酶原在血浆和其他体液中相对水平较高,传统上认为pa系统的调节主要通过pas的合成和活性水平实现。pa系统组分的合成受不同因素严格调节,如激素、生长因子和细胞因子。此外,还存在纤溶酶和pas的特定生理抑制剂。纤溶酶的主要抑制剂是α2-抗纤溶酶(α2-antiplasmin)。pas的活性同时被upa和tpa的纤溶酶原激活剂抑制剂-1(pai-1)抑制以及主要抑制upa的溶酶原激活剂抑制剂-2(pai-2)调节。某些细胞表面具有直接水解活性的upa特异性细胞表面受体(upar)[8,9]。
纤溶酶原是一个单链糖蛋白,由791个氨基酸组成,分子量约为92kda[10,11]。纤溶酶原主要在肝脏合成,大量存在于胞外液中。血浆中纤溶酶原含量约为2μm。因此纤溶酶原是组织和体液中蛋白质水解活性的一个巨大的潜在来源[12,13]。纤溶酶原存在两种分子形式:谷氨酸-纤溶酶原(glu-plasminogen)和赖氨酸-纤溶酶原(lys-plasminogen)。天然分泌和未裂解形式的纤溶酶原具有一个氨基末端(n-末端)谷氨酸,因此被称为谷氨酸-纤溶酶原。然而,在纤溶酶存在时,谷氨酸-纤溶酶原在lys76-lys77处水解成为赖氨酸-纤溶酶原。与谷氨酸-纤溶酶原相比,赖氨酸-纤溶酶原与纤维蛋白具有更高的亲和力,并可以更高的速率被pas激活。这两种形式的纤溶酶原的arg560-val561肽键可被upa或tpa切割,导致二硫键连接的双链蛋白酶纤溶酶的形成[14]。纤溶酶原的氨基末端部分包含五个同源三环,即所谓的kringles,羧基末端部分包含蛋白酶结构域。一些kringles含有介导纤溶酶原与纤维蛋白及其抑制剂α2-ap特异性相互作用的赖氨酸结合位点。最新发现一个纤溶酶原为38kda的片段,其中包括kringles1-4,是血管生成的有效抑制剂。这个片段被命名为血管抑素,可通过几个蛋白酶水解纤溶酶原产生。
纤溶酶的主要底物是纤维蛋白,纤维蛋白的溶解是预防病理性血栓形成的关键[15]。纤溶酶还具有对ecm几个组分的底物特异性,包括层粘连蛋白、纤连蛋白、蛋白聚糖和明胶,表明纤溶酶在ecm重建中也起着重要作用[11,16,17]。间接地,纤溶酶还可以通过转化某些蛋白酶前体为活性蛋白酶来降解ecm的其他组分,包括mmp-1,mmp-2,mmp-3和mmp-9。因此,有人提出,纤溶酶可能是细胞外蛋白水解的一个重要的上游调节器[18]。此外,纤溶酶具有激活某些潜在形式的生长因子的能力[19-21]。在体外,纤溶酶还能水解补体系统的组分并释放趋化补体片段。
“纤溶酶”是存在于血液中的一种非常重要的酶,能将纤维蛋白凝块水解为纤维蛋白降解产物和d-二聚体。
“纤溶酶原”是纤溶酶的酶原形式,根据swissprot中的序列,按含有信号肽的天然人源纤溶酶原氨基酸序列(序列4)计算由810个氨基酸组成,分子量约为90kd,主要在肝脏中合成并能够在血液中循环的糖蛋白,编码该氨基酸序列的cdna序列如序列3所示。全长的纤溶酶原包含七个结构域:位于c末端的丝氨酸蛋白酶结构域、n末端的panapple(pap)结构域以及5个kringle结构域(kringle1-5)。参照swissprot中的序列,其信号肽包括残基met1-gly19,pap包括残基glu20-val98,kringle1包括残基cys103-cys181,kringle2包括残基glu184-cys262,kringle3包括残基cys275-cys352,kringle4包括残基cys377-cys454,kringle5包括残基cys481-cys560。根据ncbi数据,丝氨酸蛋白酶域包括残基val581-arg804。
glu-纤溶酶原是天然全长的纤溶酶原,由791个氨基酸组成(不含有19个氨基酸的信号肽),编码该序列的cdna序列如序列1所示,其氨基酸序列如序列2所示。在体内,还存在一种是从glu-纤溶酶原的第76-77位氨基酸处水解从而形成的lys-纤溶酶原,如序列6所示,编码该氨基酸序列的cdna序列如序列5所示。delta-纤溶酶原(δ-plasminogen)是全长纤溶酶原缺失了kringle2-kringle5结构的片段,仅含有kringle1和丝氨酸蛋白酶域[22,23],有文献报道了delta-纤溶酶原的氨基酸序列(序列8)[23],编码该氨基酸序列的cdna序列如序列7。小纤溶酶原(mini-plasminogen)由kringle5和丝氨酸蛋白酶域组成,有文献报道其包括残基val443-asn791(以不含有信号肽的glu-纤溶酶原序列的glu残基为起始氨基酸)[24],其氨基酸序列如序列10所示,编码该氨基酸序列的cdna序列如序列9所示。而微纤溶酶原(micro-plasminogen)仅含有丝氨酸蛋白酶结构域,有文献报道其氨基酸序列包括残基ala543-asn791(以不含有信号肽的glu-纤溶酶原序列的glu残基为起始氨基酸)[25],也有专利文献cn102154253a报道其序列包括残基lys531-asn791(以不含有信号肽的glu-纤溶酶原序列的glu残基为起始氨基酸),本专利序列参考专利文献cn102154253a,其氨基酸序列如序列12所示,编码该氨基酸序列的cdna序列如序列11所示。
本发明的“纤溶酶”与“纤维蛋白溶酶”、“纤维蛋白溶解酶”可互换使用,含义相同;“纤溶酶原”与“纤维蛋白溶酶原”、“纤维蛋白溶解酶原”可互换使用,含义相同。
在本申请中,所述纤溶酶原“缺乏”的含义或活性为受试者体内纤溶酶原的含量比正常人低,低至足以影响所述受试者的正常生理功能;所述纤溶酶原“缺失”的含义或活性为受试者体内纤溶酶原的含量显著低于正常人,甚至活性或表达极微,只有通过外源提供才能维持正常生理功能。
本领域技术人员可以理解,本发明纤溶酶原的所有技术方案适用于纤溶酶,因此,本发明描述的技术方案涵盖了纤溶酶原和纤溶酶。
在循环过程中,纤溶酶原采用封闭的非活性构象,但当结合至血栓或细胞表面时,在纤溶酶原激活剂(plasminogenactivator,pa)的介导下,其转变为呈开放性构象的活性纤溶酶。具有活性的纤溶酶可进一步将纤维蛋白凝块水解为纤维蛋白降解产物和d-二聚体,进而溶解血栓。其中纤溶酶原的pap结构域包含维持纤溶酶原处于非活性封闭构象的重要决定簇,而kr结构域则能够与存在于受体和底物上的赖氨酸残基结合。已知多种能够作为纤溶酶原激活剂的酶,包括:组织纤溶酶原激活剂(tpa)、尿激酶纤溶酶原激活剂(upa)、激肽释放酶和凝血因子xii(哈格曼因子)等。
“纤溶酶原活性片段”是指在纤溶酶原蛋白中,能够与底物中的靶序列结合并发挥蛋白水解功能的活性片段。本发明涉及纤溶酶原的技术方案涵盖了用纤溶酶原活性片段代替纤溶酶原的技术方案。本发明所述的纤溶酶原活性片段为包含纤溶酶原的丝氨酸蛋白酶域的蛋白质,优选,本发明所述的纤溶酶原活性片段包含序列14、与序列14具有至少80%、90%、95%、96%、97%、98%、99%同源性的氨基酸序列的蛋白质。因此,本发明所述的纤溶酶原包括含有该纤溶酶原活性片段、并且仍然保持该纤溶酶原活性的蛋白。
目前,对于血液中纤溶酶原及其活性测定方法包括:对组织纤溶酶原激活剂活性的检测(t-paa)、血浆组织纤溶酶原激活剂抗原的检测(t-paag)、对血浆组织纤溶酶原活性的检测(plga)、血浆组织纤溶酶原抗原的检测(plgag)、血浆组织纤溶酶原激活剂抑制物活性的检测、血浆组织纤溶酶原激活剂抑制物抗原的检测、血浆纤维蛋白溶酶-抗纤维蛋白溶酶复合物检测(pap)。其中最常用的检测方法为发色底物法:向受检血浆中加链激酶(sk)和发色底物,受检血浆中的plg在sk的作用下,转变成plm,后者作用于发色底物,随后用分光光度计测定,吸光度增加与纤溶酶原活性成正比。此外也可采用免疫化学法、凝胶电泳、免疫比浊法、放射免疫扩散法等对血液中的纤溶酶原活性进行测定。
“直系同源物或直系同系物(ortholog)”指不同物种之间的同源物,既包括蛋白同源物也包括dna同源物,也称为直向同源物、垂直同源物。其具体指不同物种中由同一祖先基因进化而来的蛋白或基因。本发明的纤溶酶原包括人的天然纤溶酶原,还包括来源于不同物种的、具有纤溶酶原活性的纤溶酶原直系同源物或直系同系物。
“保守取代变体”是指其中一个给定的氨基酸残基改变但不改变蛋白质或酶的整体构象和功能,这包括但不限于以相似特性(如酸性,碱性,疏水性,等)的氨基酸取代亲本蛋白质中氨基酸序列中的氨基酸。具有类似性质的氨基酸是众所周知的。例如,精氨酸、组氨酸和赖氨酸是亲水性的碱性氨基酸并可以互换。同样,异亮氨酸是疏水氨基酸,则可被亮氨酸,蛋氨酸或缬氨酸替换。因此,相似功能的两个蛋白或氨基酸序列的相似性可能会不同。例如,基于megalign算法的70%至99%的相似度(同一性)。“保守取代变体”还包括通过blast或fasta算法确定具有60%以上的氨基酸同一性的多肽或酶,若能达75%以上更好,最好能达85%以上,甚至达90%以上为最佳,并且与天然或亲本蛋白质或酶相比具有相同或基本相似的性质或功能。“分离的”纤溶酶原是指从其天然环境分离和/或回收的纤溶酶原蛋白。在一些实施方案中,所述纤溶酶原会纯化(1)至大于90%、大于95%、或大于98%的纯度(按重量计),如通过lowry法所确定的,例如超过99%(按重量计),(2)至足以通过使用旋转杯序列分析仪获得n端或内部氨基酸序列的至少15个残基的程度,或(3)至同质性,该同质性是通过使用考马斯蓝或银染在还原性或非还原性条件下的十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(sds-page)确定的。分离的纤溶酶原也包括通过生物工程技术从重组细胞制备,并通过至少一个纯化步骤分离的纤溶酶原。
术语“多肽”、“肽”和“蛋白质”在本文中可互换使用,指任何长度的氨基酸的聚合形式,其可以包括遗传编码的和非遗传编码的氨基酸,化学或生物化学修饰的或衍生化的氨基酸,和具有经修饰的肽主链的多肽。该术语包括融合蛋白,包括但不限于具有异源氨基酸序列的融合蛋白,具有异源和同源前导序列(具有或没有n端甲硫氨酸残基)的融合物;等等。
关于参照多肽序列的“氨基酸序列同一性百分数(%)”定义为在必要时引入缺口以实现最大百分比序列同一性后,且不将任何保守替代视为序列同一性的一部分时,候选序列中与参照多肽序列中的氨基酸残基相同的氨基酸残基的百分率。为测定百分比氨基酸序列同一性目的的对比可以以本领域技术范围内的多种方式实现,例如使用公众可得到的计算机软件,诸如blast、blast-2、align或megalign(dnastar)软件。本领域技术人员能决定用于比对序列的适宜参数,包括对所比较序列全长实现最大对比需要的任何算法。然而,为了本发明的目的,氨基酸序列同一性百分数值是使用序列比较计算机程序align-2产生的。
在采用align-2来比较氨基酸序列的情况中,给定氨基酸序列a相对于给定氨基酸序列b的%氨基酸序列同一性(或者可表述为具有或包含相对于、与、或针对给定氨基酸序列b的某一%氨基酸序列同一性的给定氨基酸序列a)如下计算:
分数x/y乘100
其中x是由序列比对程序align-2在该程序的a和b比对中评分为相同匹配的氨基酸残基的数目,且其中y是b中的氨基酸残基的总数。应当领会,在氨基酸序列a的长度与氨基酸序列b的长度不相等的情况下,a相对于b的%氨基酸序列同一性会不等于b相对于a的%氨基酸序列同一性。除非另有明确说明,本文中使用的所有%氨基酸序列同一性值都是依照上一段所述,使用align-2计算机程序获得的。
如本文中使用的,术语“治疗”和“处理”指获得期望的药理和/或生理效果。所述效果可以是完全或部分预防疾病或其症状,和/或部分或完全治愈疾病和/或其症状,并且包括:(a)预防疾病在受试者体内发生,所述受试者可以具有疾病的素因,但是尚未诊断为具有疾病;(b)抑制疾病,即阻滞其形成;和(c)减轻疾病和/或其症状,即引起疾病和/或其症状消退。
术语“个体”、“受试者”和“患者”在本文中可互换使用,指哺乳动物,包括但不限于鼠(大鼠、小鼠)、非人灵长类、人、犬、猫、有蹄动物(例如马、牛、绵羊、猪、山羊)等。
“治疗有效量”或“有效量”指在对哺乳动物或其它受试者施用以治疗疾病时足以实现对疾病的所述预防和/或治疗的纤溶酶原的量。“治疗有效量”会根据所使用的纤溶酶原、要治疗的受试者的疾病和/或其症状的严重程度以及年龄、体重等而变化。
本发明纤溶酶原的制备
纤溶酶原可以从自然界分离并纯化用于进一步的治疗用途,也可以通过标准的化学肽合成技术来合成。当通过化学合成多肽时,可以经液相或固相进行合成。固相多肽合成(spps)(其中将序列的c末端氨基酸附接于不溶性支持物,接着序贯添加序列中剩余的氨基酸)是适合纤溶酶原化学合成的方法。各种形式的spps,诸如fmoc和boc可用于合成纤溶酶原。用于固相合成的技术描述于barany和solid-phasepeptidesynthesis;第3-284页于thepeptides:analysis,synthesis,biology.第2卷:specialmethodsinpeptidesynthesis,parta.,merrifield,等j.am.chem.soc.,85:2149-2156(1963);stewart等,solidphasepeptidesynthesis,2nded.piercechem.co.,rockford,ill.(1984);和ganesana.2006minirev.medchem.6:3-10和camareroja等2005proteinpeptlett.12:723-8中。简言之,用其上构建有肽链的功能性单元处理小的不溶性多孔珠。在偶联/去保护的重复循环后,将附接的固相游离n末端胺与单个受n保护的氨基酸单元偶联。然后,将此单元去保护,露出可以与别的氨基酸附接的新的n末端胺。肽保持固定在固相上,之后将其切掉。
可以使用标准重组方法来生产本发明的纤溶酶原。例如,将编码纤溶酶原的核酸插入表达载体中,使其与表达载体中的调控序列可操作连接。表达调控序列包括但不限于启动子(例如天然关联的或异源的启动子)、信号序列、增强子元件、和转录终止序列。表达调控可以是载体中的真核启动子系统,所述载体能够转化或转染真核宿主细胞(例如cos或cho细胞)。一旦将载体掺入合适的宿主中,在适合于核苷酸序列的高水平表达及纤溶酶原的收集和纯化的条件下维持宿主。
合适的表达载体通常在宿主生物体中作为附加体或作为宿主染色体dna的整合部分复制。通常,表达载体含有选择标志物(例如氨苄青霉素抗性、潮霉素抗性、四环素抗性、卡那霉素抗性或新霉素抗性)以有助于对外源用期望的dna序列转化的那些细胞进行检测。
大肠杆菌(escherichiacoli)是可以用于克隆主题抗体编码多核苷酸的原核宿主细胞的例子。适合于使用的其它微生物宿主包括杆菌,诸如枯草芽孢杆菌(bacillussubtilis)和其他肠杆菌科(enterobacteriaceae),诸如沙门氏菌属(salmonella)、沙雷氏菌属(serratia)、和各种假单胞菌属(pseudomonas)物种。在这些原核宿主中,也可以生成表达载体,其通常会含有与宿主细胞相容的表达控制序列(例如复制起点)。另外,会存在许多公知的启动子,诸如乳糖启动子系统,色氨酸(trp)启动子系统,beta-内酰胺酶启动子系统,或来自噬菌体λ的启动子系统。启动子通常会控制表达,任选在操纵基因序列的情况中,并且具有核糖体结合位点序列等,以启动并完成转录和翻译。
其他微生物,诸如酵母也可用于表达。酵母(例如酿酒酵母(s.cerevisiae))和毕赤酵母(pichia)是合适的酵母宿主细胞的例子,其中合适的载体根据需要具有表达控制序列(例如启动子)、复制起点、终止序列等。典型的启动子包含3-磷酸甘油酸激酶和其它糖分解酶。诱导型酵母启动于特别包括来自醇脱氢酶、异细胞色素c、和负责麦芽糖和半乳糖利用的酶的启动子。
在微生物外,哺乳动物细胞(例如在体外细胞培养物中培养的哺乳动物细胞)也可以用于表达并生成本发明的抗-tau抗体(例如编码主题抗-tau抗体的多核苷酸)。参见winnacker,fromgenestoclones,vchpublishers,n.y.,n.y.(1987)。合适的哺乳动物宿主细胞包括cho细胞系、各种cos细胞系、hela细胞、骨髓瘤细胞系、和经转化的b细胞或杂交瘤。用于这些细胞的表达载体可以包含表达控制序列,如复制起点,启动子和增强子(queen等,immunol.rev.89:49(1986)),以及必需的加工信息位点,诸如核糖体结合位点,rna剪接位点,多聚腺苷酸化位点,和转录终止子序列。合适的表达控制序列的例子是白免疫球蛋白基因、sv40、腺病毒、牛乳头瘤病毒、巨细胞病毒等衍生的启动子。参见co等,j.immunol.148:1149(1992)。
一旦合成(化学或重组方式),可以依照本领域的标准规程,包括硫酸铵沉淀,亲和柱,柱层析,高效液相层析(hplc),凝胶电泳等来纯化本发明所述的纤溶酶原。该纤溶酶原是基本上纯的,例如至少约80%至85%纯的,至少约85%至90%纯的,至少约90%至95%纯的,或98%至99%纯的或更纯的,例如不含污染物,所述污染物如细胞碎片,除主题抗体以外的大分子,等等。
药物配制剂
可以通过将具有所需纯度的纤溶酶原与可选的药用载体,赋形剂,或稳定剂(remington'spharmaceuticalsciences,16版,osol,a.ed.(1980))混合形成冻干制剂或水溶液制备治疗配制剂。可接受的载体、赋形剂、稳定剂在所用剂量及浓度下对受者无毒性,并包括缓冲剂例如磷酸盐,柠檬酸盐及其它有机酸;抗氧化剂包括抗坏血酸和蛋氨酸;防腐剂(例如十八烷基二甲基苄基氯化铵;氯化己烷双胺;氯化苄烷铵(benzalkoniumchloride),苯索氯铵;酚、丁醇或苯甲醇;烷基对羟基苯甲酸酯如甲基或丙基对羟基苯甲酸酯;邻苯二酚;间苯二酚;环己醇;3-戊醇;间甲酚);低分子量多肽(少于约10个残基);蛋白质如血清白蛋白,明胶或免疫球蛋白;亲水聚合物如聚乙烯吡咯烷酮;氨基酸如甘氨酸,谷氨酰胺、天冬酰胺、组氨酸、精氨酸或赖氨酸;单糖,二糖及其它碳水化合物包括葡萄糖、甘露糖、或糊精;螯合剂如edta;糖类如蔗糖、甘露醇、岩藻糖或山梨醇;成盐反离子如钠;金属复合物(例如锌-蛋白复合物);和/或非离子表面活性剂,例如tweentm,pluronicstm或聚乙二醇(peg)。优选冻干的抗-vegf抗体配制剂在wo97/04801中描述,其包含在本文中作为参考。
本发明的配制剂也可含有需治疗的具体病症所需的一种以上的活性化合物,优选活性互补并且相互之间没有副作用的那些。例如,抗高血压的药物,抗心律失常的药物,治疗糖尿病的药物等。
本发明的纤溶酶原可包裹在通过诸如凝聚技术或界面聚合而制备的微胶囊中,例如,可置入在胶质药物传送系统(例如,脂质体,白蛋白微球,微乳剂,纳米颗粒和纳米胶囊)中或置入粗滴乳状液中的羟甲基纤维素或凝胶-微胶囊和聚-(甲基丙烯酸甲酯)微胶囊中。这些技术公开于remington'spharmaceuticalsciences16thedition,osol,a.ed.(1980)。
用于体内给药的本发明的纤溶酶原必需是无菌的。这可以通过在冷冻干燥和重新配制之前或之后通过除菌滤膜过滤而轻易实现。
本发明的纤溶酶原可制备缓释制剂。缓释制剂的适当实例包括具有一定形状且含有糖蛋白的固体疏水聚合物半通透基质,例如膜或微胶囊。缓释基质实例包括聚酯、水凝胶(如聚(2-羟基乙基-异丁烯酸酯)(langer等,j.biomed.mater.res.,15:167-277(1981);langer,chem.tech.,12:98-105(1982))或聚(乙烯醇),聚交酯(美国专利3773919,ep58,481),l-谷氨酸与γ乙基-l-谷氨酸的共聚物(sidman,等,biopolymers22:547(1983)),不可降解的乙烯-乙烯乙酸酯(ethylene-vinylacetate)(langer,等,出处同上),或可降解的乳酸-羟基乙酸共聚物如luprondepottm(由乳酸-羟基乙酸共聚物和亮氨酰脯氨酸(leuprolide)乙酸酯组成的可注射的微球体),以及聚d-(-)-3-羟丁酸。聚合物如乙烯-乙酸乙烯酯和乳酸-羟基乙酸能持续释放分子100天以上,而一些水凝胶释放蛋白的时间却较短。可以根据相关机理来设计使蛋白稳定的合理策略。例如,如果发现凝聚的机理是通过硫代二硫键互换而形成分子间s-s键,则可通过修饰巯基残基、从酸性溶液中冻干、控制湿度、采用合适的添加剂、和开发特定的聚合物基质组合物来实现稳定。
给药和剂量
可以通过不同方式,例如通过静脉内,腹膜内,皮下,颅内,鞘内,动脉内(例如经由颈动脉),肌内来实现本发明药物组合物的施用。
用于胃肠外施用的制备物包括无菌水性或非水性溶液、悬浮液和乳剂。非水性溶剂的例子是丙二醇、聚乙二醇、植物油如橄榄油,和可注射有机酯,如油酸乙酯。水性载体包括水、醇性/水性溶液、乳剂或悬浮液,包括盐水和缓冲介质。胃肠外媒介物包含氯化钠溶液、林格氏右旋糖、右旋糖和氯化钠、或固定油。静脉内媒介物包含液体和营养补充物、电解质补充物,等等。也可以存在防腐剂和其他添加剂,诸如例如,抗微生物剂、抗氧化剂、螯合剂、和惰性气体,等等。
医务人员会基于各种临床因素确定剂量方案。如医学领域中公知的,任一患者的剂量取决于多种因素,包括患者的体型、体表面积、年龄、要施用的具体化合物、性别、施用次数和路径、总体健康、和同时施用的其它药物。本发明包含纤溶酶原的药物组合物的剂量范围可以例如为例如每天约0.0001至2000mg/kg,或约0.001至500mg/kg(例如0.02mg/kg,0.25mg/kg,0.5mg/kg,0.75mg/kg,10mg/kg,50mg/kg等等)受试者体重。例如,剂量可以是1mg/kg体重或50mg/kg体重或在1-50mg/kg的范围,或至少1mg/kg。高于或低于此例示性范围的剂量也涵盖在内,特别是考虑到上述的因素。上述范围中的中间剂量也包含在本发明的范围内。受试者可以每天、隔天、每周或根据通过经验分析确定的任何其它日程表施用此类剂量。例示性的剂量日程表包括连续几天1-10mg/kg。在本发明的药物施用过程中需要实时评估治疗效果和安全性。
制品或药盒
本发明的一个实施方案涉及一种制品或药盒,其包含可用于治疗骨关节炎的本发明纤溶酶原或纤溶酶。所述制品优选包括一个容器,标签或包装插页。适当的容器有瓶子,小瓶,注射器等。容器可由各种材料如玻璃或塑料制成。所述容器含有组合物,所述组合物可有效治疗本发明的疾病或病症并具有无菌入口(例如所述容器可为静脉内溶液包或小瓶,其含有可被皮下注射针穿透的塞子的)。所述组合物中至少一种活性剂为纤溶酶原/纤溶酶。所述容器上或所附的标签说明所述组合物用于治疗本发明所述骨关节炎。所述制品可进一步包含含有可药用缓冲液的第二容器,诸如磷酸盐缓冲的盐水,林格氏溶液以及葡萄糖溶液。其可进一步包含从商业和使用者角度来看所需的其它物质,包括其它缓冲液,稀释剂,过滤物,针和注射器。此外,所述制品包含带有使用说明的包装插页,包括例如指示所述组合物的使用者将纤溶酶原组合物以及治疗伴随的疾病的其它药物给药患者。
附图说明
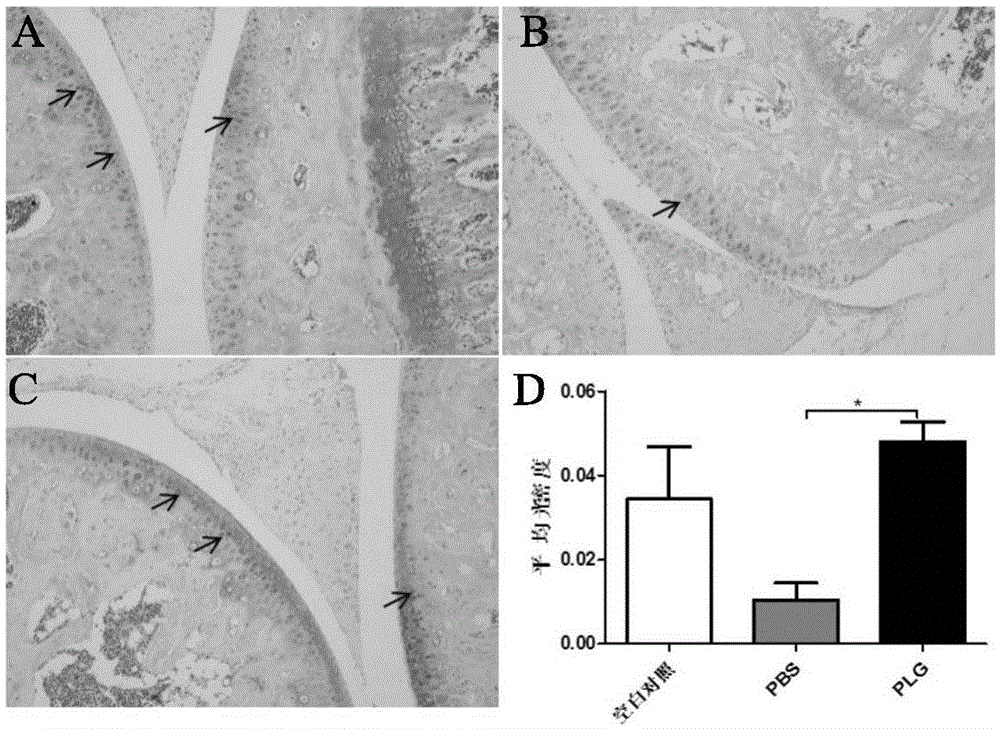
图1a-d显示0.5μg/kg维生素d衰老模型小鼠膝关节safranino染色结果。a为空白对照组,b为给溶媒pbs对照组,c为给纤溶酶原组,d为定量分析结果。结果显示,给纤溶酶原组膝关节软骨(箭头标识)明显多于给溶媒pbs对照组,统计差异显著(*表示p<0.05),且与给溶媒pbs对照组相比给纤溶酶原组膝关节软骨量更加接近空白对照小鼠。说明纤溶酶原能够显著减少维生素d衰老模型小鼠膝关节软骨丢失。
图2a-e给予纤溶酶原14天二型胶原酶诱导的骨关节炎模型小鼠膝关节safranino染色评分结果。a、c为给溶媒pbs对照组,b、d为给纤溶酶原组,e为定量分析结果。结果显示,给纤溶酶原组膝关节病理评分明显低于给溶媒pbs对照组,且统计差异极其显著(**表示p<0.01)。说明纤溶酶原能够减轻二型胶原酶诱导的骨关节炎模型小鼠膝关节的损伤。
图3a-d给予纤溶酶原14天二型胶原酶诱导的骨关节炎模型plg-/-小鼠膝关节safranino染色代表性图片。a、c为给溶媒pbs对照组,b、d为给纤溶酶原组。结果显示,给溶媒pbs对照组软骨组织(细箭头标识)结构排列紊乱,细胞数目明显减少,safranino着色明显减少,骨小梁(粗箭头标识)变细、断裂;给纤溶酶原组相对于给溶媒pbs对照组,软骨组织结构相对整齐,软骨处细胞数目相对较多,safranino着色范围相对较广。说明纤溶酶原能够减少二型胶原酶诱导的骨关节炎plg-/-小鼠膝关节的损伤。
图4a-b给予纤溶酶原14天韧带切断诱导的骨关节炎模型小鼠膝关节safranino染色代表性图片。a为给溶媒pbs对照组,b为给纤溶酶原组。结果显示,给溶媒pbs对照组软骨(三角形标识)丢失严重,骨小梁(箭头标识)变细,断裂,出现较大面积的无骨小梁骨髓腔;给纤溶酶原组,较之于pbs对照组,骨小梁连续性较好,没有较严重的断裂,没有较大面积的无骨小梁区域,软骨组织相对较多。说明纤溶酶原能够改善韧带切断诱导的骨关节炎模型小鼠膝关节组织结构状况。
图5a-e给予纤溶酶原14天韧带切断诱导的骨关节炎模型小鼠膝关节碱性磷酸酶染色果。a、c为给溶媒pbs对照组,b、d为给纤溶酶原组,e为定量分析结果。结果显示,给纤溶酶原组小鼠膝关节软骨表面(细箭头标识)和生长板(粗箭头标识)碱性磷酸酶着色均多于给溶媒pbs对照组,且统计差异显著(*表示p<0.05)。说明纤溶酶原能够显著促进韧带切断诱导的骨关节炎模型小鼠膝关节碱性磷酸酶活性的增加,即纤溶酶原促使膝关节成骨细胞活性明显增加。
图6a-d给予纤溶酶原28天mia骨关节炎模型小鼠膝关节ii型胶原免疫组化染色结果。a为假手术组,b为给溶媒对照组,c为给纤溶酶原组,d为定量分析结果。结果显示,假手术组小鼠膝关节有一定量的ii型胶原(箭头标识),给溶媒对照组膝关节ii型胶原量与假手术组无明显差别,而给纤溶酶原组膝关节ii型胶原量明显多于给溶媒对照组和假手术组,且平均光密度定量分析结果统计差异显著(*表示p<0.05)。结果表明纤溶酶原能够促进骨关节炎模型小鼠膝关节软骨再生。
图7a-d给予纤溶酶原28天mia骨关节炎模型小鼠膝关节safranino染色代表性图片。a为假手术组,b为给溶媒对照组,c为给纤溶酶原组,d为病理评分结果。结果显示,假手术组小鼠膝关节有一定量的软骨存在(箭头标识);给溶媒对照组膝关节软骨量明显减少并且病理学评分明显增加,说明mia成功诱导骨关节炎;给纤溶酶原组膝关节软骨量明显多于给溶媒对照组,病理学评分也明显低于给溶媒对照组,且统计学差异显著(*表示p<0.05)。该结果表明纤溶酶原能够促进软骨再生,改善骨关节炎损伤。
图8a-d给予纤溶酶原28天骨关节炎模型小鼠左侧膝关节股骨表面safranino染色结果。a为假手术组,b为给溶媒对照组,c为给纤溶酶原组,d为平均光密度定量分析结果。结果显示,假手术组小鼠膝关节处股骨表面有少量的软骨(箭头标识),给溶媒对照组股骨表面软骨量与假手术组无明显差别,而给纤溶酶原组股骨表面软骨量明显多于给溶媒对照组和假手术组,且平均光密度定量分析结果统计差异显著(*表示p<0.05)。结果表明纤溶酶原能够促进骨关节炎模型小鼠股骨表面软骨再生。
图9给予纤溶酶原7天mia骨关节炎模型大鼠疼痛检测结果。结果显示,假手术组大鼠具有较高的疼痛阈值;给溶媒对照组大鼠左右腿疼痛阈值均明显降低,明显低于假手术组;给纤溶酶原组大鼠左右腿疼痛阈值明显升高,与给溶媒对照组相比,左腿统计差异接近显著(p=0.08),右腿统计差异显著(*表示p<0.05)。结果表明纤溶酶原能够明显减轻骨性关节炎疼痛。
图10给予纤溶酶原28天骨关节炎模型小鼠疼痛检测结果。结果显示,假手术组小鼠疼痛阈值相对较高;给溶媒对照组小鼠疼痛阈值明显降低,明显低于假手术组小鼠;给纤溶酶原组小鼠疼痛阈值明显升高,明显高于给溶媒对照组小鼠,且统计差异接近显著(p=0.09)。该结果表明纤溶酶原能够减轻骨关节炎疼痛。
图11a-f给予纤溶酶原28天骨关节炎模型小鼠左侧膝关节safranino染色结果。结果显示,假手术组左侧膝关节股骨和胫骨表面存在一定量的关节软骨(箭头标识),给溶媒对照组胫骨和股骨表面软骨量与假手术组差别不明显,给纤溶酶原组软骨量明显多于给溶媒对照组和假手术组。说明纤溶酶原可促进骨关节炎模型小鼠关节软骨再生。
图12a-d显示纤溶酶原抑制骨关节炎模型小鼠骨吸收。结果显示,假手术组(图12a)膝关节存在一定量的酸性磷酸酶(箭头标识);给溶媒对照组(图12b)膝关节酸性磷酸酶增多,明显多于假手术组;给纤溶酶原组(图12c)膝关节酸性磷酸酶明显少于给溶媒对照组,且统计差异显著(*表示p<0.05)(图12d)。该结果表明纤溶酶原可减少骨关节炎膝关节酸性磷酸酶,降低破骨细胞活性,抑制骨吸收。
图13a-f显示纤溶酶原促进mia骨关节炎膝关节胫骨端sox9阳性干细胞数量增加。结果显示,假手术组(图13a,d)膝关节胫骨存在一定量的sox9阳性干细胞(箭头标识),给溶媒对照组(图13b,e)膝关节胫骨sox9阳性干细胞数量明显减少,给纤溶酶原组(图13c,f)膝关节胫骨sox9阳性干细胞数量明显多于给溶媒对照组。该结果表明纤溶酶原可促进骨关节炎膝关节胫骨端sox9阳性干细胞数量增加,修复骨关节炎对膝关节的损伤。
图14a-c显示纤溶酶原改善mia骨关节炎模型小鼠膝关节滑膜炎症。结果显示,假手术组(图14a)膝关节滑膜未见明显的炎症细胞浸润,给溶媒对照组(图14b)膝关节滑膜可见明显的炎症细胞浸润(箭头标识),给纤溶酶原组(图14c)膝关节滑膜炎症细胞浸润明显少于给溶媒对照组。该结果表明纤溶酶原可改善骨关节炎膝关节滑膜炎症。
实施例
实施例1纤溶酶原减少维生素d诱导的骨质疏松小鼠膝关节软骨丢失
取5-6周龄的雄性c57小鼠15只,称重后随机分为3组,空白对照组、给纤溶酶原组和给溶媒pbs对照组,每组各5只。空白对照组小鼠每天腹腔注射50μl玉米油;给纤溶酶原组和给溶媒pbs对照组小鼠按照0.5μg/kg体重每天腹腔注射维生素d(sigmaaldrich),诱导骨质疏松模型[26,27]。与此同时小鼠开始给药,给纤溶酶原组小鼠尾静脉注射人源纤溶酶原1mg/0.1ml/只/天,给溶媒pbs对照组尾静脉注射同体积的pbs,空白对照组小鼠不做给药处理,连续造模给药28天。给药期间所有小鼠饲喂低钙饲料。开始造模给药定为第1天,第29天处死后取材膝关节于4%多聚甲醛固定24小时,然后10%edta中脱钙三周,梯度蔗糖溶液洗涤,以上操作需在4℃条件下进行。然后石蜡包埋,8μm切片行safranino染色。切片在100倍光学显微镜下观察。
软骨染色液(番红o法)染色原理在于嗜碱性的软骨与碱性染料番红o结合呈现红色,番红o是一种结合多阴离子的阳离子染料,其显示软骨是基于阳离子染料与多糖中阴离子基团(硫酸软骨素或硫酸角质素)结合。番红o着色与阴离子的浓度近似成正比关系,间接反映基质中蛋白多糖的含量和分布。
safranino染色即番红染色,主要显示软骨组织中的酸性蛋白聚糖成分,能够显示软骨形成[28]。
结果显示,给纤溶酶原组(图1c)膝关节软骨(箭头标识)明显多于给溶媒pbs对照组(图1b),统计差异显著(*表示p<0.05)(图1d),且与给溶媒pbs对照组相比给纤溶酶原组膝关节软骨更加接近空白对照小鼠(图1a)。说明纤溶酶原能够显著减少维生素d骨质疏松模型小鼠膝关节软骨丢失。
实施例2纤溶酶原减少二型胶原酶诱导的骨关节炎模型小鼠膝关节损伤
10周龄的c57雄性小鼠12只,按照50mg/kg体重腹腔注射戊巴比妥钠,麻醉小鼠,然后肌肉注射tolfedine(0.1ml/kg)。在小鼠右侧膝关节处开个缺口,膝关节弯曲90度,小鼠按照5μg/6μl/只关节腔内注射二型胶原酶(c6885,sigma);左侧膝关节与右侧膝关节操作相同,但仅注射相同体积的生理盐水[29,30]。注射7天后小鼠分别按照体重随机分为两组,给溶媒pbs对照组和给纤溶酶原组,每组各6只,并开始给药。给纤溶酶原组按照1mg/0.1ml/只/天尾静脉注射给予人纤溶酶原,给溶媒pbs对照组尾静脉注射相同体积的pbs,连续给药14天。开始给药定为第1天,第15天处死后取材膝关节于4%多聚甲醛固定24小时,然后10%edta中脱钙三周,梯度蔗糖溶液洗涤,以上操作需在4℃条件下进行。然后石蜡包埋,8μm切片行safranino染色。切片在40(a、b)和100(c、d)倍光学显微镜下观察,并按表1对膝关节进行病理评分[31]。
表1
结果显示,给纤溶酶原组膝关节(图2b、d)病理评分明显低于给溶媒pbs对照组(图2a、c),且统计差异极其显著(**表示p<0.01)(图2e)。说明纤溶酶原能够显著减轻二型胶原酶诱导的骨关节炎对膝关节的损伤,减少关节软骨的丢失。
实施例3纤溶酶原改善二型胶原酶诱导的骨关节炎模型plg-/-小鼠膝关节组织结构状态
10周龄的plg-/-雄性小鼠10只,按照50mg/kg体重腹腔注射戊巴比妥钠,麻醉小鼠,然后肌肉注射tolfedine(0.1ml/kg)。在小鼠右侧膝关节处开个缺口,膝关节弯曲90度,小鼠按照5μg/6μl/只关节腔内注射二型胶原酶(c6885,sigma);左侧膝关节与右侧膝关节操作相同,但仅注射相同体积的生理盐水[29,30]。注射7天后小鼠分别按照体重随机分为两组,给溶媒pbs对照组和给纤溶酶原组,每组各5只,并开始给药。给纤溶酶原组按照1mg/0.1ml/只/天尾静脉注射给予人纤溶酶原,给溶媒pbs对照组尾静脉注射相同体积的pbs,连续给药14天。开始给药定为第1天,第15天处死后取材膝关节于4%多聚甲醛固定24小时,然后10%edta中脱钙三周,梯度蔗糖溶液洗涤,以上操作需在4℃条件下进行。然后石蜡包埋,4μm切片行safranino染色。切片在40(a、b)和100(c、d)倍光学显微镜下观察。
结果显示,给溶媒pbs对照组(图3a、c)软骨组织(细箭头标识)结构排列紊乱,细胞数目明显减少,safranino着色明显减少,骨小梁(粗箭头标识)变细、断裂;给纤溶酶原组(图3b、d)相对于给溶媒pbs对照组,软骨组织结构相对整齐,软骨处细胞数目相对较多,safranino着色范围相对较广。说明纤溶酶原能够减少二型胶原酶诱导的骨关节炎plg-/-小鼠膝关节的损伤。
实施例4纤溶酶原改善韧带切断诱导的骨关节炎模型小鼠膝关节组织结构状态
9-10周龄的c57雄性小鼠10只。按照50mg/kg体重腹腔注射戊巴比妥钠,麻醉小鼠。脱去膝关节周围毛发,临近手术前小鼠肌肉注射tolfedine(0.1ml/kg)镇痛剂。小鼠放置在解剖显微镜下,在右侧膝盖骨远端和胫骨坪开口,分离髌骨韧带,暴露关节腔;钝性分离股骨髁间的脂肪垫,使得前十字交叉韧带能够被观察到;用显微刀片挑断前十字交叉韧带和内侧副韧带,缝合关节囊与皮肤,建立骨关节炎模型[32,33]。左侧膝关节仅暴露关节腔,不进行切断操作。术中应注意避免损伤关节软骨,术后不固定肢体,笼内自由活动。手术后第一天,每隔12小时小鼠肌肉注射tolfedine(0.1mg/kg)及青霉素钾(4万单位/kg)。两周后小鼠根据体重随机分为两组,给溶媒pbs对照组和给纤溶酶原组,每组各5只,并开始给药。给纤溶酶原组按照1mg/0.1ml/只/天尾静脉注射给予人纤溶酶原,给溶媒pbs对照组尾静脉注射相同体积的pbs,连续给药14天。开始给药定为第1天,第15天处死后取材膝关节于4%多聚甲醛固定24小时,然后10%edta中脱钙三周,梯度蔗糖溶液洗涤,以上操作需在4℃条件下进行。然后石蜡包埋,4μm切片行safranino染色。切片在40倍光学显微镜下观察。
结果显示,给溶媒pbs对照组(图4a)软骨(三角形标识)丢失严重,骨小梁(箭头标识)变细,断裂,出现较大面积的无骨小梁骨髓腔;给纤溶酶原组(图4b),较之于pbs对照组,骨小梁连续性较好,没有较严重的断裂,没有较大面积的无骨小梁区域,软骨组织相对较多。说明纤溶酶原能够改善韧带切断诱导的骨关节炎模型小鼠膝关节组织结构状况。
实施例5纤溶酶原增加韧带切断诱导的骨关节炎模型小鼠膝关节碱性磷酸酶活性
9-10周龄的c57雄性小鼠10只。按照50mg/kg体重腹腔注射戊巴比妥钠,麻醉小鼠。脱去膝关节周围毛发,临近手术前小鼠肌肉注射tolfedine(0.1ml/kg)镇痛剂。小鼠放置在解剖显微镜下,在右侧膝盖骨远端和胫骨坪开口,分离髌骨韧带,暴露关节腔;钝性分离股骨髁间的脂肪垫,使得前十字交叉韧带能够被观察到;用显微刀片挑断前十字交叉韧带,缝合关节囊与皮肤,建立骨关节炎模型[32,33]。左侧膝关节仅暴露关节腔,不进行切断操作。术中应注意避免损伤关节软骨,术后不固定肢体,笼内自由活动。手术后第一天,每隔12小时小鼠肌肉注射tolfedine(0.1mg/kg)及青霉素钾(4万单位/kg)。两周后小鼠根据体重随机分为两组,给溶媒pbs对照组和给纤溶酶原组,每组各5只,并开始给药。给纤溶酶原组按照1mg/0.1ml/只/天尾静脉注射给予人纤溶酶原,给溶媒pbs对照组尾静脉注射相同体积的pbs,连续给药14天。开始给药定为第1天,第15天处死后取材膝关节于固定液中固定。固定液配方:2%多聚甲醛,0.075mol/l赖氨酸,0.01mol/l过碘酸钠。固定后4℃pbs洗液梯度洗涤各12小时,然后置于4℃脱钙液中脱钙2周,每5天换一次脱钙液。脱钙完成后4℃pbs洗液梯度洗涤12小时,膝关节经酒精梯度脱水和二甲苯透明后进行石蜡包埋。切片5um,脱蜡复水,氯化镁缓冲液4℃孵育过夜。碱性磷酸酶底物溶液室温孵育1小时,苏木素复染2分钟。流水冲洗5分钟,60℃烤30分钟,中性树胶封片,切片在200倍光学显微镜下观察。
碱性磷酸酶(alkalinephosphatase,alp)是成骨细胞早期分化的标志[32]。
结果显示,给纤溶酶原组小鼠(图5b、d)膝关节软骨表面和生长板碱性磷酸酶着色(箭头标识)均多于给溶媒pbs对照组(图5a、c),且统计差异显著(*表示p<0.05)(图5e)。说明纤溶酶原能够显著促进韧带切断诱导的骨关节炎模型小鼠膝关节碱性磷酸酶活性的增加,即纤溶酶原促使膝关节关节软骨的成骨细胞活性明显增加。
实施例6纤溶酶原促进mia骨关节炎模型小鼠膝关节软骨再生
取8-10周c57雄性小鼠25只,称重,按体重随机分为两组,假手术组5只,模型组20只。所有小鼠按照50mg/kg体重腹腔注射3%戊巴比妥钠麻醉,麻醉后模型组小鼠脱去左侧膝盖处毛发,70%的酒精以及碘酊消毒,左侧膝关节弯曲90度,沿着膝盖水平运行注射器的针(以免刺穿皮肤),直到发现髌骨下方的间隙。用轻微的压力标记该区域,然后垂直提起针头和注射器,将针头插入标记区域,穿过垂直于胫骨的髌腱,关节腔内注射mia(碘乙酸,monoiodoaceticacid,mia)生理盐水溶液0.1mg/10μl;假手术组左侧关节腔注射10μl生理盐水,注射后,按摩膝盖,以确保均匀分布[34]。右侧膝关节不处理。关节腔内mia注射3天后,模型组小鼠进行疼痛测试,根据测试结果小鼠随机分为两组,给溶媒对照组和给纤溶酶原组,每组各10只,并开始给药,记为给药第1天,给纤溶酶原组小鼠按照1mg/0.1ml/只/天尾静脉注射人纤溶酶原,给溶媒对照组小鼠每天尾静脉注射同体积溶媒pbs缓冲液,连续注射28天。假手术组小鼠不做给药处理。mia溶液配制:mia粉末(sigma,57858-5g)溶解于生理盐水中,浓度为10mg/ml,然后0.22μm滤膜过滤,现用现配。第29天处死小鼠取材左侧膝关节于plp固定液中固定,然后10%edta中脱钙三周,梯度蔗糖溶液洗涤,然后石蜡包埋。组织切片厚度为5μm,切片脱蜡复水后水洗1次。pap笔圈出组织,以3%双氧水孵育15分钟,0.01mpbs洗2次,每次5分钟。5%的正常羊血清液(vectorlaboratories,inc.,usa)封闭30分钟;时间到后,弃除羊血清液,滴加兔源抗ii型胶原抗体(abcam,ab34712)4℃孵育过夜,0.01mpbs洗2次,每次5分钟。山羊抗兔igg(hrp)抗体(abcam)(二抗)室温孵育1小时,0.01mpbs洗2次,每次5分钟。按dab试剂盒(vectorlaboratories,inc.,usa)显色,水洗3次后苏木素复染30秒,流水冲洗5分钟。梯度酒精脱水,二甲苯透明并中性树胶封片,切片在200倍光学显微镜下观察拍照,并用image-pro软件处理收集数据。
碘乙酸(monoiodoaceticacid,mia)有破坏关节软骨和周围滑膜韧带的作用,将其注射入关节腔内,能改变关节腔原有的稳态,还可引发关节内炎症反应,从而改变软骨和软骨下骨的新陈代谢,破坏关节内环境的稳定,使软骨细胞凋亡,软骨磨损。研究表明,碘乙酸能够抑制关节软骨细胞代谢,引起软骨细胞死亡,进而导致软骨基质降解,诱导出关节炎软骨改变和骨关节炎模型。ii型胶原是软骨基质的主要成分之一,在软骨机械特性维持上发挥重要作用[35]。
结果显示,假手术组(图6a)小鼠膝关节有一定量的ii型胶原(箭头标识),给溶媒对照组(图6b)膝关节ii型胶原量与假手术组无明显差别,而给纤溶酶原组(图6c)膝关节ii型胶原量明显多于给溶媒对照组和假手术组,且平均光密度定量分析结果统计差异显著(*表示p<0.05)(图6d)。结果表明纤溶酶原能够促进骨关节炎模型小鼠膝关节软骨再生。
实施例7纤溶酶原对mia诱导的骨关节炎改善作用
取8-10周c57雄性小鼠25只,称重,按体重随机分为两组,假手术组5只,模型组20只。所有小鼠按照50mg/kg体重腹腔注射3%戊巴比妥钠麻醉,麻醉后模型组小鼠脱去左侧膝盖处毛发,70%的酒精以及碘酊消毒,左侧膝关节弯曲90度,沿着膝盖水平运行注射器的针(以免刺穿皮肤),直到发现髌骨下方的间隙。用轻微的压力标记该区域,然后垂直提起针头和注射器,将针头插入标记区域,穿过垂直于胫骨的髌腱,关节腔内注射mia生理盐水溶液0.1mg/10μl;假手术组左侧关节腔注射10μl生理盐水,注射后,按摩膝盖,以确保均匀分布[34]。右侧膝关节不处理。关节腔内mia注射3天后,模型组小鼠进行疼痛测试,根据测试结果小鼠随机分为两组,给溶媒对照组和给纤溶酶原组,每组各10只,并开始给药,记为给药第1天,给纤溶酶原组小鼠按照1mg/0.1ml/只/天尾静脉注射人纤溶酶原,给溶媒对照组小鼠每天尾静脉注射同体积溶媒pbs缓冲液,连续注射28天。假手术组小鼠不做给药处理。mia溶液配制:mia粉末(sigma,57858-5g)溶解于生理盐水中,浓度为10mg/ml,然后0.22μm滤膜过滤,现用现配。第29天处死小鼠取材左侧膝关节于plp固定液中固定,然后10%edta中脱钙三周,梯度蔗糖溶液洗涤,然后石蜡包埋。5μm切片行safranino染色。切片在40倍光学显微镜下观察拍照,按照表1进行病理学评分。
结果显示,假手术组(图7a)小鼠膝关节有一定量的软骨存在(箭头标识);给溶媒对照组(图7b)膝关节软骨量明显减少并且病理学评分明显增加,说明mia成功诱导骨关节炎;给纤溶酶原组(图7c)膝关节软骨量明显多于给溶媒对照组,病理学评分也明显低于给溶媒对照组,且统计学差异显著(*表示p<0.05)(图7d)。该结果表明纤溶酶原能够促进软骨再生,修复骨关节炎对关节的损伤。
实施例8纤溶酶原促进骨关节炎模型小鼠膝关节软骨再生
取11-15周龄db/db雌性小鼠18只,称重,按体重随机分为两组,假手术组3只,模型组15只。模型组小鼠按照50mg/kg体重腹腔注射3%戊巴比妥钠麻醉,脱去背部两侧毛发以70%的酒精以及碘酊消毒,切开皮肤、背部肌肉和腹膜,以小镊子轻轻将白色发亮脂肪团拉出切口外,分离脂肪团,便可见卵巢。先将卵巢下端输卵管用丝线结扎,然后摘除卵巢。假手术组小鼠麻醉后只切开皮肤、背部肌肉和腹膜,不摘除卵巢。所有手术小鼠缝合消毒后每只肌肉注射抗生素(5000u/只),皮下注射止痛药(2mg/kg),连续注射3天。卵巢切除65天后,所有小鼠腹腔注射戊巴比妥钠麻醉,脱去右侧膝盖处毛发,70%的酒精以及碘酊消毒,右侧膝关节弯曲90度,找到精确的注射部位,沿着膝盖水平运行注射器的针(以免刺穿皮肤),直到发现髌骨下方的间隙。用轻微的压力标记该区域,然后垂直提起针头和注射器,将针头插入标记区域,穿过垂直于胫骨的髌腱,应该感觉不到任何阻力,模型组小鼠右侧关节腔内注射5μg/6μlii型胶原酶生理盐水溶液,假手术组小鼠右侧关节腔内注射6μl生理盐水[36,37]。注射后,按摩膝盖,以确保均匀分布。左侧膝关节不做注射处理。胶原酶注射7天后,所有小鼠称重,模型组小鼠按照体重随机分为两组,给溶媒对照组7只和给纤溶酶原组8只,并开始给药,记为给药第1天,给纤溶酶原组小鼠按照1mg/0.1ml/只/天尾静脉注射人纤溶酶原,给溶媒对照组小鼠每天尾静脉注射同体积溶媒pbs缓冲液,连续注射28天。假手术组小鼠不做给药处理。第29天处死小鼠取材左侧膝关节于plp固定液中固定,然后10%edta中脱钙三周,梯度蔗糖溶液洗涤。然后石蜡包埋,5μm切片行safranino染色。切片在100倍光学显微镜下观察拍照,并用image-pro软件处理收集数据。
结果显示,假手术组(图8a)小鼠膝关节处股骨表面有少量的软骨(箭头标识),给溶媒对照组(图8b)股骨表面软骨量与假手术组无明显差别,而给纤溶酶原组(图8c)股骨表面软骨量明显多于给溶媒对照组和假手术组,且平均光密度定量分析结果统计差异显著(*表示p<0.05)(图8d)。结果表明纤溶酶原能够促进骨关节炎模型小鼠股骨表面软骨再生。
实施例9纤溶酶原减轻mia骨关节炎模型大鼠关节炎疼痛
取sd大鼠21只,体重约200g~250g,称重,按体重随机分为两组,假手术组5只,模型组16只。所有大鼠腹腔注射3%戊巴比妥钠(50mg/kg)麻醉,模型组大鼠脱去左、右两侧膝关节处毛发,70%的酒精以及碘酊消毒,膝关节弯曲90度,找到精确的注射部位,沿着膝盖水平运行注射器的针(以免刺穿皮肤),直到发现髌骨下方的间隙。用轻微的压力标记该区域,然后垂直提起针头和注射器,将针头插入标记区域,穿过垂直于胫骨的髌腱,应该感觉不到任何阻力,双侧关节腔内分别注射mia生理盐水溶液2mg/50μl;假手术组左、右两侧关节腔内注射50μl生理盐水。注射后,按摩膝盖,以确保均匀分布[38]。mia溶液配制:mia粉末(sigma,57858-5g)溶解于生理盐水中,浓度40mg/ml,0.22μm滤膜过滤,现用现配。mia注射3天后,模型组大鼠进行疼痛测试,根据测试结果随机分为两组,给溶媒对照组和给纤溶酶原组,每组各8只,并开始给药,记为第1天。给溶媒对照组每只大鼠每天尾静脉注射0.7ml溶媒(10mm柠檬酸钠缓冲液,2%盐酸精氨酸,3%甘露醇,ph7.4),给纤溶酶原组大鼠按照7mg/0.7ml/只/天尾静脉注射人纤溶酶原,连续给药7天,给药期间注意观察动物状态。第8天左右腿进行疼痛测试[38]。
结果显示,假手术组大鼠疼痛阈值相对较高;给溶媒对照组大鼠左右腿疼痛阈值均明显降低,明显低于假手术组;给纤溶酶原组大鼠左右腿疼痛阈值明显升高,与给溶媒对照组相比,左腿统计差异接近显著(p=0.08),右腿统计差异显著(*表示p<0.05)(图9)。结果表明纤溶酶原能够明显减轻骨性关节炎疼痛。
实施例10纤溶酶原减轻骨关节炎模型小鼠关节炎疼痛
取7周龄c57雌性小鼠19只,称重,按体重随机分为两组,假手术组3只,模型组16只。模型组小鼠按照50mg/kg体重腹腔注射3%戊巴比妥钠麻醉,脱去背部两侧毛发以70%的酒精以及碘酊消毒,切开皮肤、背部肌肉和腹膜,以小镊子轻轻将白色发亮脂肪团拉出切口外,分离脂肪团,便可见卵巢。先将卵巢下端输卵管用丝线结扎,然后摘除卵巢。假手术组小鼠麻醉后只切开皮肤、背部肌肉和腹膜,不摘除卵巢。所有手术小鼠缝合消毒后肌肉注射抗生素(5000u/只),皮下注射止痛药(2mg/kg),连续注射3天。卵巢切除65天后,所有小鼠腹腔注射戊巴比妥钠麻醉,脱去右侧膝盖处毛发,70%的酒精以及碘酊消毒,右侧膝关节弯曲90度,找到精确的注射部位,沿着膝盖水平运行注射器的针(以免刺穿皮肤),直到发现髌骨下方的间隙。用轻微的压力标记该区域,然后垂直提起针头和注射器,将针头插入标记区域,穿过垂直于胫骨的髌腱,应该感觉不到任何阻力,模型组小鼠右侧关节腔内注射5μg/6μlii型胶原酶生理盐水溶液,假手术组小鼠右侧关节腔内注射6μl生理盐水[36,37]。注射后,按摩膝盖,以确保均匀分布。左侧膝关节不做注射处理。胶原酶注射7天后,所有小鼠称重,模型组小鼠按照体重随机分为两组,给溶媒对照组和给纤溶酶原组,每组各8只,并开始给药,记为给药第1天,给纤溶酶原组小鼠按照1mg/0.1ml/只/天尾静脉注射人纤溶酶原,给溶媒对照组小鼠每天尾静脉注射同体积溶媒pbs缓冲液,连续注射28天。假手术组小鼠不做给药处理。第29天右腿进行疼痛测试[34]。
结果显示,假手术组小鼠疼痛阈值相对较高;给溶媒对照组小鼠疼痛阈值明显降低,明显低于假手术组小鼠;给纤溶酶原组小鼠疼痛阈值明显升高,明显高于给溶媒对照组小鼠,且统计差异接近显著(p=0.09)(图10)。该结果表明纤溶酶原能够减轻骨关节炎疼痛。
实施例11纤溶酶原促进骨关节炎模型小鼠软骨再生
取7周龄c57雌性小鼠19只,称重,按体重随机分为两组,假手术组3只,模型组16只。模型组小鼠按照50mg/kg体重腹腔注射3%戊巴比妥钠麻醉,脱去背部两侧毛发以70%的酒精以及碘酊消毒,切开皮肤、背部肌肉和腹膜,以小镊子轻轻将白色发亮脂肪团拉出切口外,分离脂肪团,便可见卵巢。先将卵巢下端输卵管用丝线结扎,然后摘除卵巢。假手术组小鼠麻醉后只切开皮肤、背部肌肉和腹膜,不摘除卵巢。所有手术小鼠缝合消毒后肌肉注射抗生素(5000u/只),皮下注射止痛药(2mg/kg),连续注射3天。卵巢切除65天后,所有小鼠腹腔注射戊巴比妥钠麻醉,脱去右侧膝盖处毛发,70%的酒精以及碘酊消毒,右侧膝关节弯曲90度,找到精确的注射部位,沿着膝盖水平运行注射器的针(以免刺穿皮肤),直到发现髌骨下方的间隙。用轻微的压力标记该区域,然后垂直提起针头和注射器,将针头插入标记区域,穿过垂直于胫骨的髌腱,应该感觉不到任何阻力,模型组小鼠右侧关节腔内注射5μg/6μlii型胶原酶生理盐水溶液,假手术组小鼠右侧关节腔内注射6μl生理盐水[36,37]。注射后,按摩膝盖,以确保均匀分布。左侧膝关节不做注射处理。胶原酶注射7天后,所有小鼠称重,模型组小鼠按照体重随机分为两组,给溶媒对照组和给纤溶酶原组,每组各8只,并开始给药,记为给药第1天,给纤溶酶原组小鼠按照1mg/0.1ml/只/天尾静脉注射人纤溶酶原,给溶媒对照组小鼠每天尾静脉注射同体积溶媒pbs缓冲液,连续注射28天。假手术组小鼠不做给药处理。第29天处死小鼠取材两侧膝关节于plp固定液中固定,然后10%edta中脱钙三周,梯度蔗糖溶液洗涤。然后石蜡包埋,5μm切片行safranino染色。切片在100倍光学显微镜下观察拍照,并用image-pro软件处理收集数据。
结果显示,假手术组(图11a,d)左侧膝关节股骨和胫骨表面存在一定量的关节软骨(箭头标识),给溶媒对照组(图11b,e)胫骨和股骨表面软骨量与假手术组差别不明显,给纤溶酶原组(图11c,f)软骨量明显多于给溶媒对照组和假手术组。说明纤溶酶原可促进骨关节炎模型小鼠关节软骨再生。
实施例12纤溶酶原抑制骨关节炎模型小鼠骨吸收
取7周龄c57雌性小鼠19只,称重,按体重随机分为两组,假手术组3只,模型组16只。模型组小鼠按照50mg/kg体重腹腔注射3%戊巴比妥钠麻醉,脱去背部两侧毛发以70%的酒精以及碘酊消毒,切开皮肤、背部肌肉和腹膜,以小镊子轻轻将白色发亮脂肪团拉出切口外,分离脂肪团,便可见卵巢。先将卵巢下端输卵管用丝线结扎,然后摘除卵巢。假手术组小鼠麻醉后只切开皮肤、背部肌肉和腹膜,不摘除卵巢。所有手术小鼠缝合消毒后肌肉注射抗生素(5000u/只),皮下注射止痛药(2mg/kg),连续注射3天。卵巢切除65天后,所有小鼠腹腔注射戊巴比妥钠麻醉,脱去右侧膝盖处毛发,70%的酒精以及碘酊消毒,右侧膝关节弯曲90度,找到精确的注射部位,沿着膝盖水平运行注射器的针(以免刺穿皮肤),直到发现髌骨下方的间隙。用轻微的压力标记该区域,然后垂直提起针头和注射器,将针头插入标记区域,穿过垂直于胫骨的髌腱,应该感觉不到任何阻力,模型组小鼠右侧关节腔内注射5μg/6μlii型胶原酶生理盐水溶液,假手术组小鼠右侧关节腔内注射6μl生理盐水[36,37]。注射后,按摩膝盖,以确保均匀分布。左侧膝关节不做注射处理。胶原酶注射7天后,所有小鼠称重,模型组小鼠按照体重随机分为两组,给溶媒对照组和给纤溶酶原组,每组各8只,并开始给药,记为给药第1天,给纤溶酶原组小鼠按照1mg/0.1ml/只/天尾静脉注射人纤溶酶原,给溶媒对照组小鼠每天尾静脉注射同体积溶媒pbs缓冲液,连续注射28天。假手术组小鼠不做给药处理。第29天处死小鼠取材左侧膝关节于plp固定液中固定,然后10%edta中脱钙三周,梯度蔗糖溶液洗涤,切片5μm,脱蜡复水,进行酸性磷酸酶(trap)染色。
酸性磷酸酶(trap)为破骨细胞的特异性标志酶,破骨细胞是骨吸收的主要功能细胞。结果显示,假手术组(图12a)膝关节存在一定量的酸性磷酸酶(箭头标识);给溶媒对照组(图12b)膝关节酸性磷酸酶增多,明显多于假手术组;给纤溶酶原组(图12c)膝关节酸性磷酸酶明显少于给溶媒对照组,且统计差异显著(*表示p<0.05)(图12d)。该结果表明纤溶酶原可减少骨关节炎膝关节酸性磷酸酶,降低破骨细胞活性,抑制骨吸收。
实施例13纤溶酶原促进mia骨关节炎膝关节胫骨端sox9阳性干细胞数量增加
取8-10周c57雄性小鼠25只,称重,按体重随机分为两组,假手术组5只,模型组20只。所有小鼠按照50mg/kg体重腹腔注射3%戊巴比妥钠麻醉,麻醉后模型组小鼠脱去左侧膝盖处毛发,70%的酒精以及碘酊消毒,左侧膝关节弯曲90度,沿着膝盖水平运行注射器的针(以免刺穿皮肤),直到发现髌骨下方的间隙。用轻微的压力标记该区域,然后垂直提起针头和注射器,将针头插入标记区域,穿过垂直于胫骨的髌腱,关节腔内注射mia生理盐水溶液0.1mg/10μl;假手术组左侧关节腔注射10μl生理盐水,注射后,按摩膝盖,以确保均匀分布[34]。右侧膝关节不处理。关节腔内mia注射3天后,模型组小鼠进行疼痛测试,根据测试结果小鼠随机分为两组,给溶媒对照组和给纤溶酶原组,每组各10只,并开始给药,记为给药第1天,给纤溶酶原组小鼠按照1mg/0.1ml/只/天尾静脉注射人纤溶酶原,给溶媒对照组小鼠每天尾静脉注射同体积溶媒pbs缓冲液,连续注射28天。假手术组小鼠不做给药处理。mia溶液配制:mia粉末(sigma,57858-5g)溶解于生理盐水中,浓度为10mg/ml,然后0.22μm滤膜过滤,现用现配。第29天处死小鼠取材左侧膝关节于plp固定液中固定,然后10%edta中脱钙三周,梯度蔗糖溶液洗涤,然后石蜡包埋。组织切片厚度为5μm,切片脱蜡复水后水洗1次。pap笔圈出组织,以3%双氧水孵育15分钟,0.01mpbs洗2次,每次5分钟。5%的正常羊血清液(vectorlaboratories,inc.,usa)封闭30分钟;时间到后,弃除羊血清液,滴加兔源抗sox9抗体(abcam,ab185966)4℃孵育过夜,0.01mpbs洗2次,每次5分钟。山羊抗兔igg(hrp)抗体(abcam)(二抗)室温孵育1小时,0.01mpbs洗2次,每次5分钟。按dab试剂盒(vectorlaboratories,inc.,usa)显色,水洗3次后苏木素复染30秒,流水冲洗5分钟。梯度酒精脱水,二甲苯透明并中性树胶封片,切片在100(a-c)和400(d-f)倍光学显微镜下观察拍照,并用image-pro软件处理收集数据。
sox9是软骨发育形成过程中的关键转录因子,决定间充质干细胞的聚集和向软骨细胞分化,对软骨的发育成熟及损伤修复等过程起着重要的调节作用[39]。
结果显示,假手术组(图13a,d)膝关节胫骨存在一定量的sox9阳性干细胞(箭头标识),给溶媒对照组(图13b,e)膝关节胫骨sox9阳性干细胞数量明显减少,给纤溶酶原组(图13c,f)膝关节胫骨sox9阳性干细胞数量明显多于给溶媒对照组。该结果表明纤溶酶原可促进骨关节炎膝关节胫骨端sox9阳性干细胞数量增加,修复骨关节炎对膝关节的损伤。
实施例14纤溶酶原改善mia骨关节炎模型小鼠膝关节滑膜炎症
取8-10周c57雄性小鼠25只,称重,按体重随机分为两组,假手术组5只,模型组20只。所有小鼠按照50mg/kg体重腹腔注射3%戊巴比妥钠麻醉,麻醉后模型组小鼠脱去左侧膝盖处毛发,70%的酒精以及碘酊消毒,左侧膝关节弯曲90度,沿着膝盖水平运行注射器的针(以免刺穿皮肤),直到发现髌骨下方的间隙。用轻微的压力标记该区域,然后垂直提起针头和注射器,将针头插入标记区域,穿过垂直于胫骨的髌腱,关节腔内注射mia生理盐水溶液0.1mg/10μl;假手术组左侧关节腔注射10μl生理盐水,注射后,按摩膝盖,以确保均匀分布[34]。右侧膝关节不处理。关节腔内mia注射3天后,模型组小鼠进行疼痛测试,根据测试结果小鼠随机分为两组,给溶媒对照组和给纤溶酶原组,每组各10只,并开始给药,记为给药第1天,给纤溶酶原组小鼠按照1mg/0.1ml/只/天尾静脉注射人纤溶酶原,给溶媒对照组小鼠每天尾静脉注射同体积溶媒pbs缓冲液,连续注射28天。假手术组小鼠不做给药处理。mia溶液配制:mia粉末(sigma,57858-5g)溶解于生理盐水中,浓度为10mg/ml,然后0.22μm滤膜过滤,现用现配。第29天处死小鼠取材左侧膝关节于plp固定液中固定,然后10%edta中脱钙三周,梯度蔗糖溶液洗涤,然后石蜡包埋(保留膝关节周边的肌肉)。组织切片厚度为5μm,切片脱蜡复水并用苏木素和伊红染色(h&e染色),1%盐酸酒精分化,氨水返蓝,并酒精梯度脱水,二甲苯透明,中性树胶封片,切片在400倍光学显微镜下观察。
结果显示,假手术组(图14a)膝关节滑膜未见明显的炎症细胞浸润,给溶媒对照组(图14b)膝关节滑膜可见明显的炎症细胞浸润(箭头标识),给纤溶酶原组(图14c)膝关节滑膜炎症细胞浸润明显少于给溶媒对照组。该结果表明纤溶酶原可改善骨关节炎膝关节滑膜炎症。
以上所有实施例中使用的(人)纤溶酶原来自人捐赠者血浆,基于文献[40-42]所描述的方法并进行工艺优化,从人血浆中纯化所得。纤溶酶原单体的纯度>95%。
参考文献:
[1]nationalinstituteofarthritisandmusculoskeletalandskindiseases.april2015.archivedfromtheoriginalon18may2015.retrieved13may2015.
[2]glyn-joness,palmeraj,agricolar,priceaj,vincenttl,weinansh,carraj(july2015)."osteoarthritis".lancet.386(9991):376–87.
[3]berenbaumf(january2013)."osteoarthritisasaninflammatorydisease(osteoarthritisisnotosteoarthrosis!)".osteoarthritisandcartilage.21(1):16–21.
[4]lig,yinj,gaoj,chengts,pavlosnj,zhangc,zhengmh(2013)."subchondralboneinosteoarthritis:insightintoriskfactorsandmicrostructuralchanges".
[5]alexandercmandwerb,z.(1991).extracellularmatrixdegradation.incellbiologyofextracellularmatrix,hayed,ed.(newyork:plenumpress),pp.255-302.
[6]werb,z.,mainardi,c.l.,vater,c.a.,andharris,e.d.,jr.(1977).endogenousactiviationoflatentcollagenasebyrheumatoidsynovialcells.evidenceforaroleofplasminogenactivator.n.engl.j.med.296,1017-1023.
[7]he,c.s.,wilhelm,s.m.,pentland,a.p.,marmer,b.l.,grant,g.a.,eisen,a.z.,andgoldberg,g.i.(1989).tissuecooperationinaproteolyticcascadeactivatinghumaninterstitialcollagenase.proc.natl.acad.sci.u.s.a86,2632-2636.
[8]stoppelli,m.p.,corti,a.,soffientini,a.,cassani,g.,blasi,f.,andassoian,r.k.(1985).differentiation-enhancedbindingoftheamino-terminalfragmentofhumanurokinaseplasminogenactivatortoaspecificreceptoronu937monocytes.proc.natl.acad.sci.u.s.a82,4939-4943.
[9]vassalli,j.d.,baccino,d.,andbelin,d.(1985).acellularbindingsiteforthemr55,000formofthehumanplasminogenactivator,urokinase.j.cellbiol.100,86-92.
[10]wiman,b.andwallen,p.(1975).structuralrelationshipbetween"glutamicacid"and"lysine"formsofhumanplasminogenandtheirinteractionwiththenh2-terminalactivationpeptideasstudiedbyaffinitychromatography.eur.j.biochem.50,489-494.
[11]saksela,o.andrifkin,d.b.(1988).cell-associatedplasminogenactivation:regulationandphysiologicalfunctions.annu.rev.cellbiol.4,93-126.
[12]raum,d.,marcus,d.,alper,c.a.,levey,r.,taylor,p.d.,andstarzl,t.e.(1980).synthesisofhumanplasminogenbytheliver.science208,1036-1037.
[13]davidm.waisman.plasminogen:structure,activation,andregulation.springerus,2003.
[14]sottrup-jensen,l.,zajdel,m.,claeys,h.,petersen,t.e.,andmagnusson,s.(1975).amino-acidsequenceofactivationcleavagesiteinplasminogen:homologywith"pro"partofprothrombin.proc.natl.acad.sci.u.s.a72,2577-2581.
[15]collen,d.andlijnen,h.r.(1991).basicandclinicalaspectsoffibrinolysisandthrombolysis.blood78,3114-3124.
[16]alexander,c.m.andwerb,z.(1989).proteinasesandextracellularmatrixremodeling.curr.opin.cellbiol.1,974-982.
[17]mignatti,p.andrifkin,d.b.(1993).biologyandbiochemistryofproteinasesintumorinvasion.physiolrev.73,161-195.
[18]collen,d.(2001).ham-wassermanlecture:roleoftheplasminogensysteminfibrin-homeostasisandtissueremodeling.hematology.(am.soc.hematol.educ.program.)1-9.
[19]rifkin,d.b.,moscatelli,d.,bizik,j.,quarto,n.,blei,f.,dennis,p.,flaumenhaft,r.,andmignatti,p.(1990).growthfactorcontrolofextracellularproteolysis.celldiffer.dev.32,313-318.
[20]andreasen,p.a.,kjoller,l.,christensen,l.,andduffy,m.j.(1997).theurokinase-typeplasminogenactivatorsystemincancermetastasis:areview.int.j.cancer72,1-22.
[21]rifkin,d.b.,mazzieri,r.,munger,j.s.,noguera,i.,andsung,j.(1999).proteolyticcontrolofgrowthfactoravailability.apmis107,80-85.
[22]mardervj,novokhatnyv.directfibrinolyticagents:biochemicalattributes,preclinicalfoundationandclinicalpotential[j].journalofthrombosisandhaemostasis,2010,8(3):433-444.
[23]huntja,pettewayjrsr,scuderip,etal.simplifiedrecombinantplasmin:productionandfunctionalcomparisonofanovelthrombolyticmoleculewithplasma-derivedplasmin[j].thrombhaemost,2008,100(3):413-419.
[24]medynskid,tuanm,liuw,etal.refolding,purification,andactivationofminiplasminogenandmicroplasminogenisolatedfrome.coliinclusionbodies.[j].proteinexpression&purification,2007,52(2):395-402.
[25]nagain,demarsine,vanhoefb,etal.recombinanthumanmicroplasmin:productionandpotentialtherapeuticproperties[j].journalofthrombosisandhaemostasis,2003,1(2):307-313.
[26].e.daci,a.verstuyf,k.moermansetal.boneresorptioninducedby1a,25dihydroxyvitamind3invivoisnotalteredbyinactivationoftheplasminogenactivatorinhibitor1.bonevol.27,no.1july2000:97–102.
[27].mohammeds.razzaque,despinasitara,takashitaguchietal.prematureaging-likephenotypeinfibroblastgrowthfactor23nullmiceisavitamind-mediatedprocess.fasebj.2006apr;20(6):720–722.
[28]mackay,a.m.,beck,s.c.,murphy,j.m.,barry,f.p.,chichester,c.o.,&pittenger,m.f.(1998).chondrogenicdifferentiationofculturedhumanmesenchymalstemcellsfrommarrow.tissueengineer.4:415-428.
[29].peterl.e.m.vanlent,1arjenb.blom,1rikf.p.schelbergenetal.activeinvolvementofalarminss100a8ands100a9intheregulationofsynovialactivationandjointdestructionduringmouseandhumanosteoarthritis.arthritisrheum.2012may;64(5):1466-76.doi:10.1002/art.34315.
[30].peterm.vanderkraan,ellyl.vitters,henkm.vanbeuningenetal.degenerativekneejointlesionsinmiceafterasingleintra-articularcollagenaseinjection.anewmodelofosteoarthritis.jexppathol(oxford).1990feb;71(1):19-31.
[31].ostergaardk,andersencb,petersenjetal.validityofhistopathologicalgradingofarticularcartilagefromosteoarthritickneejoints.annrheumdis.1999apr;58(4):208-13.
[32].weinrebm,shinard,rodang.differentpatternofalkalinephosphatase,osteopontin,andosteocalcinexpressionindevelopingratbonevisualizedbyinsituhybridizationj.jboneminerres,1990,5(8):831-842.
[33]s.kamekuram.d.y,k.hoshim.d.,ph.detal.osteoarthritisdevelopmentinnovelexperimentalmousemodelsinducedbykneejointinstability.osteoarthritiscartilage.2005jul;13(7):632-41.
[34]ogbonnaac,clarkaketal.pain-likebehaviourandspinalchangesinthemonosodiumiodoacetatemodelofosteoarthritisinc57bl/6mice.eurjpain.2013apr;17(4):514-26.
[35]keigosato,hisashimera,shigeyukiwakitanietal.effectofepigallocatechin-3-gallateontheincreaseintypeiicollagenaccumulationincartilage-likemscsheets.journaloftheagriculturalchemicalsocietyofjapan,2017,81(6):5.
[36]gustavoduque,daochaohuang,nataliedionetal.interferon-gplaysaroleinboneformationinvivoandrescuesosteoporosisinovariectomizedmice.journalofboneandmineralresearch,vol.26,no.7,july2011,pp1472–1483.
[37]peterm.vanderkraan,ellyl.vitters,henkm.vanbeuningenetal.degenerativekneejointlesionsinmiceafterasingleintra-articularcollagenaseinjection.anewmodelofosteoarthritis.jexppathol(oxford).1990feb;71(1):19-31.
[38]comber,bramwells,fieldmjetal.themonosodiumiodoacetatemodelofosteoarthritis:amodelofchronicnociceptivepaininrats?neuroscilett.2004nov11;370(2-3):236-40.
[39]hardinghamte,oldershawra,tewsr.cartilage,sox9andnotchsignalsinchondrogenesis[j].journalofanatomy,2010,209(4):469-480.
[40]kennethcrobbins,louissummaria,davidelwynetal.furtherstudiesonthepurificationandcharacterizationofhumanplasminogenandplasmin.journalofbiologicalchemistry,1965,240(1):541-550.
[41]summarial,spitzf,arzadonletal.isolationandcharacterizationoftheaffinitychromatographyformsofhumanglu-andlys-plasminogensandplasmins.jbiolchem.1976jun25;251(12):3693-9.
[42]haganjj,ablondifb,derenzoec.purificationandbiochemicalpropertiesofhumanplasminogen.jbiolchem.1960apr;235:1005-10.
序列表
<110>深圳瑞健生命科学研究院有限公司
<120>一种预防或治疗骨关节炎的方法和药物
<130>pb00391
<150>201711350845.3
<151>2017-12-15
<160>14
<170>patentinversion3.5
<210>1
<211>2376
<212>dna
<213>不含有信号肽的天然纤溶酶原(glu-plg,glu-纤维蛋白溶酶原)核酸序列
<400>1
gagcctctggatgactatgtgaatacccagggggcttcactgttcagtgtcactaagaag60
cagctgggagcaggaagtatagaagaatgtgcagcaaaatgtgaggaggacgaagaattc120
acctgcagggcattccaatatcacagtaaagagcaacaatgtgtgataatggctgaaaac180
aggaagtcctccataatcattaggatgagagatgtagttttatttgaaaagaaagtgtat240
ctctcagagtgcaagactgggaatggaaagaactacagagggacgatgtccaaaacaaaa300
aatggcatcacctgtcaaaaatggagttccacttctccccacagacctagattctcacct360
gctacacacccctcagagggactggaggagaactactgcaggaatccagacaacgatccg420
caggggccctggtgctatactactgatccagaaaagagatatgactactgcgacattctt480
gagtgtgaagaggaatgtatgcattgcagtggagaaaactatgacggcaaaatttccaag540
accatgtctggactggaatgccaggcctgggactctcagagcccacacgctcatggatac600
attccttccaaatttccaaacaagaacctgaagaagaattactgtcgtaaccccgatagg660
gagctgcggccttggtgtttcaccaccgaccccaacaagcgctgggaactttgtgacatc720
ccccgctgcacaacacctccaccatcttctggtcccacctaccagtgtctgaagggaaca780
ggtgaaaactatcgcgggaatgtggctgttaccgtgtccgggcacacctgtcagcactgg840
agtgcacagacccctcacacacataacaggacaccagaaaacttcccctgcaaaaatttg900
gatgaaaactactgccgcaatcctgacggaaaaagggccccatggtgccatacaaccaac960
agccaagtgcggtgggagtactgtaagataccgtcctgtgactcctccccagtatccacg1020
gaacaattggctcccacagcaccacctgagctaacccctgtggtccaggactgctaccat1080
ggtgatggacagagctaccgaggcacatcctccaccaccaccacaggaaagaagtgtcag1140
tcttggtcatctatgacaccacaccggcaccagaagaccccagaaaactacccaaatgct1200
ggcctgacaatgaactactgcaggaatccagatgccgataaaggcccctggtgttttacc1260
acagaccccagcgtcaggtgggagtactgcaacctgaaaaaatgctcaggaacagaagcg1320
agtgttgtagcacctccgcctgttgtcctgcttccagatgtagagactccttccgaagaa1380
gactgtatgtttgggaatgggaaaggataccgaggcaagagggcgaccactgttactggg1440
acgccatgccaggactgggctgcccaggagccccatagacacagcattttcactccagag1500
acaaatccacgggcgggtctggaaaaaaattactgccgtaaccctgatggtgatgtaggt1560
ggtccctggtgctacacgacaaatccaagaaaactttacgactactgtgatgtccctcag1620
tgtgcggccccttcatttgattgtgggaagcctcaagtggagccgaagaaatgtcctgga1680
agggttgtaggggggtgtgtggcccacccacattcctggccctggcaagtcagtcttaga1740
acaaggtttggaatgcacttctgtggaggcaccttgatatccccagagtgggtgttgact1800
gctgcccactgcttggagaagtccccaaggccttcatcctacaaggtcatcctgggtgca1860
caccaagaagtgaatctcgaaccgcatgttcaggaaatagaagtgtctaggctgttcttg1920
gagcccacacgaaaagatattgccttgctaaagctaagcagtcctgccgtcatcactgac1980
aaagtaatcccagcttgtctgccatccccaaattatgtggtcgctgaccggaccgaatgt2040
ttcatcactggctggggagaaacccaaggtacttttggagctggccttctcaaggaagcc2100
cagctccctgtgattgagaataaagtgtgcaatcgctatgagtttctgaatggaagagtc2160
caatccaccgaactctgtgctgggcatttggccggaggcactgacagttgccagggtgac2220
agtggaggtcctctggtttgcttcgagaaggacaaatacattttacaaggagtcacttct2280
tggggtcttggctgtgcacgccccaataagcctggtgtctatgttcgtgtttcaaggttt2340
gttacttggattgagggagtgatgagaaataattaa2376
<210>2
<211>791
<212>prt
<213>不含有信号肽的天然纤溶酶原(glu-plg,glu-纤维蛋白溶酶原)氨基酸序列
<400>2
gluproleuaspasptyrvalasnthrglnglyalaserleupheser
151015
valthrlyslysglnleuglyalaglyserilegluglucysalaala
202530
lyscysglugluaspglugluphethrcysargalapheglntyrhis
354045
serlysgluglnglncysvalilemetalagluasnarglysserser
505560
ileileileargmetargaspvalvalleupheglulyslysvaltyr
65707580
leuserglucyslysthrglyasnglylysasntyrargglythrmet
859095
serlysthrlysasnglyilethrcysglnlystrpserserthrser
100105110
prohisargproargpheserproalathrhisprosergluglyleu
115120125
glugluasntyrcysargasnproaspasnaspproglnglyprotrp
130135140
cystyrthrthraspproglulysargtyrasptyrcysaspileleu
145150155160
glucysglugluglucysmethiscysserglygluasntyraspgly
165170175
lysileserlysthrmetserglyleuglucysglnalatrpaspser
180185190
glnserprohisalahisglytyrileproserlyspheproasnlys
195200205
asnleulyslysasntyrcysargasnproasparggluleuargpro
210215220
trpcysphethrthraspproasnlysargtrpgluleucysaspile
225230235240
proargcysthrthrproproproserserglyprothrtyrglncys
245250255
leulysglythrglygluasntyrargglyasnvalalavalthrval
260265270
serglyhisthrcysglnhistrpseralaglnthrprohisthrhis
275280285
asnargthrprogluasnpheprocyslysasnleuaspgluasntyr
290295300
cysargasnproaspglylysargalaprotrpcyshisthrthrasn
305310315320
serglnvalargtrpglutyrcyslysileprosercysaspserser
325330335
provalserthrgluglnleualaprothralaproprogluleuthr
340345350
provalvalglnaspcystyrhisglyaspglyglnsertyrarggly
355360365
thrserserthrthrthrthrglylyslyscysglnsertrpserser
370375380
metthrprohisarghisglnlysthrprogluasntyrproasnala
385390395400
glyleuthrmetasntyrcysargasnproaspalaasplysglypro
405410415
trpcysphethrthraspproservalargtrpglutyrcysasnleu
420425430
lyslyscysserglythrglualaservalvalalaproproproval
435440445
valleuleuproaspvalgluthrproserglugluaspcysmetphe
450455460
glyasnglylysglytyrargglylysargalathrthrvalthrgly
465470475480
thrprocysglnasptrpalaalaglngluprohisarghisserile
485490495
phethrprogluthrasnproargalaglyleuglulysasntyrcys
500505510
argasnproaspglyaspvalglyglyprotrpcystyrthrthrasn
515520525
proarglysleutyrasptyrcysaspvalproglncysalaalapro
530535540
serpheaspcysglylysproglnvalgluprolyslyscysprogly
545550555560
argvalvalglyglycysvalalahisprohissertrpprotrpgln
565570575
valserleuargthrargpheglymethisphecysglyglythrleu
580585590
ileserproglutrpvalleuthralaalahiscysleuglulysser
595600605
proargprosersertyrlysvalileleuglyalahisglngluval
610615620
asnleugluprohisvalglngluilegluvalserargleupheleu
625630635640
gluprothrarglysaspilealaleuleulysleuserserproala
645650655
valilethrasplysvalileproalacysleuproserproasntyr
660665670
valvalalaaspargthrglucyspheilethrglytrpglygluthr
675680685
glnglythrpheglyalaglyleuleulysglualaglnleuproval
690695700
ilegluasnlysvalcysasnargtyrglupheleuasnglyargval
705710715720
glnserthrgluleucysalaglyhisleualaglyglythraspser
725730735
cysglnglyaspserglyglyproleuvalcyspheglulysasplys
740745750
tyrileleuglnglyvalthrsertrpglyleuglycysalaargpro
755760765
asnlysproglyvaltyrvalargvalserargphevalthrtrpile
770775780
gluglyvalmetargasnasn
785790
<210>3
<211>2433
<212>dna
<213>含有信号肽的天然纤溶酶原(来源于swissprot)的核酸序列
<400>3
atggaacataaggaagtggttcttctacttcttttatttctgaaatcaggtcaaggagag60
cctctggatgactatgtgaatacccagggggcttcactgttcagtgtcactaagaagcag120
ctgggagcaggaagtatagaagaatgtgcagcaaaatgtgaggaggacgaagaattcacc180
tgcagggcattccaatatcacagtaaagagcaacaatgtgtgataatggctgaaaacagg240
aagtcctccataatcattaggatgagagatgtagttttatttgaaaagaaagtgtatctc300
tcagagtgcaagactgggaatggaaagaactacagagggacgatgtccaaaacaaaaaat360
ggcatcacctgtcaaaaatggagttccacttctccccacagacctagattctcacctgct420
acacacccctcagagggactggaggagaactactgcaggaatccagacaacgatccgcag480
gggccctggtgctatactactgatccagaaaagagatatgactactgcgacattcttgag540
tgtgaagaggaatgtatgcattgcagtggagaaaactatgacggcaaaatttccaagacc600
atgtctggactggaatgccaggcctgggactctcagagcccacacgctcatggatacatt660
ccttccaaatttccaaacaagaacctgaagaagaattactgtcgtaaccccgatagggag720
ctgcggccttggtgtttcaccaccgaccccaacaagcgctgggaactttgtgacatcccc780
cgctgcacaacacctccaccatcttctggtcccacctaccagtgtctgaagggaacaggt840
gaaaactatcgcgggaatgtggctgttaccgtgtccgggcacacctgtcagcactggagt900
gcacagacccctcacacacataacaggacaccagaaaacttcccctgcaaaaatttggat960
gaaaactactgccgcaatcctgacggaaaaagggccccatggtgccatacaaccaacagc1020
caagtgcggtgggagtactgtaagataccgtcctgtgactcctccccagtatccacggaa1080
caattggctcccacagcaccacctgagctaacccctgtggtccaggactgctaccatggt1140
gatggacagagctaccgaggcacatcctccaccaccaccacaggaaagaagtgtcagtct1200
tggtcatctatgacaccacaccggcaccagaagaccccagaaaactacccaaatgctggc1260
ctgacaatgaactactgcaggaatccagatgccgataaaggcccctggtgttttaccaca1320
gaccccagcgtcaggtgggagtactgcaacctgaaaaaatgctcaggaacagaagcgagt1380
gttgtagcacctccgcctgttgtcctgcttccagatgtagagactccttccgaagaagac1440
tgtatgtttgggaatgggaaaggataccgaggcaagagggcgaccactgttactgggacg1500
ccatgccaggactgggctgcccaggagccccatagacacagcattttcactccagagaca1560
aatccacgggcgggtctggaaaaaaattactgccgtaaccctgatggtgatgtaggtggt1620
ccctggtgctacacgacaaatccaagaaaactttacgactactgtgatgtccctcagtgt1680
gcggccccttcatttgattgtgggaagcctcaagtggagccgaagaaatgtcctggaagg1740
gttgtaggggggtgtgtggcccacccacattcctggccctggcaagtcagtcttagaaca1800
aggtttggaatgcacttctgtggaggcaccttgatatccccagagtgggtgttgactgct1860
gcccactgcttggagaagtccccaaggccttcatcctacaaggtcatcctgggtgcacac1920
caagaagtgaatctcgaaccgcatgttcaggaaatagaagtgtctaggctgttcttggag1980
cccacacgaaaagatattgccttgctaaagctaagcagtcctgccgtcatcactgacaaa2040
gtaatcccagcttgtctgccatccccaaattatgtggtcgctgaccggaccgaatgtttc2100
atcactggctggggagaaacccaaggtacttttggagctggccttctcaaggaagcccag2160
ctccctgtgattgagaataaagtgtgcaatcgctatgagtttctgaatggaagagtccaa2220
tccaccgaactctgtgctgggcatttggccggaggcactgacagttgccagggtgacagt2280
ggaggtcctctggtttgcttcgagaaggacaaatacattttacaaggagtcacttcttgg2340
ggtcttggctgtgcacgccccaataagcctggtgtctatgttcgtgtttcaaggtttgtt2400
acttggattgagggagtgatgagaaataattaa2433
<210>4
<211>810
<212>prt
<213>含有信号肽的天然纤溶酶原(来源于swissprot)的氨基酸序列
<400>4
metgluhislysgluvalvalleuleuleuleuleupheleulysser
151015
glyglnglygluproleuaspasptyrvalasnthrglnglyalaser
202530
leupheservalthrlyslysglnleuglyalaglyserilegluglu
354045
cysalaalalyscysglugluaspglugluphethrcysargalaphe
505560
glntyrhisserlysgluglnglncysvalilemetalagluasnarg
65707580
lysserserileileileargmetargaspvalvalleupheglulys
859095
lysvaltyrleuserglucyslysthrglyasnglylysasntyrarg
100105110
glythrmetserlysthrlysasnglyilethrcysglnlystrpser
115120125
serthrserprohisargproargpheserproalathrhisproser
130135140
gluglyleuglugluasntyrcysargasnproaspasnaspprogln
145150155160
glyprotrpcystyrthrthraspproglulysargtyrasptyrcys
165170175
aspileleuglucysglugluglucysmethiscysserglygluasn
180185190
tyraspglylysileserlysthrmetserglyleuglucysglnala
195200205
trpaspserglnserprohisalahisglytyrileproserlysphe
210215220
proasnlysasnleulyslysasntyrcysargasnproaspargglu
225230235240
leuargprotrpcysphethrthraspproasnlysargtrpgluleu
245250255
cysaspileproargcysthrthrproproproserserglyprothr
260265270
tyrglncysleulysglythrglygluasntyrargglyasnvalala
275280285
valthrvalserglyhisthrcysglnhistrpseralaglnthrpro
290295300
histhrhisasnargthrprogluasnpheprocyslysasnleuasp
305310315320
gluasntyrcysargasnproaspglylysargalaprotrpcyshis
325330335
thrthrasnserglnvalargtrpglutyrcyslysileprosercys
340345350
aspserserprovalserthrgluglnleualaprothralapropro
355360365
gluleuthrprovalvalglnaspcystyrhisglyaspglyglnser
370375380
tyrargglythrserserthrthrthrthrglylyslyscysglnser
385390395400
trpsersermetthrprohisarghisglnlysthrprogluasntyr
405410415
proasnalaglyleuthrmetasntyrcysargasnproaspalaasp
420425430
lysglyprotrpcysphethrthraspproservalargtrpglutyr
435440445
cysasnleulyslyscysserglythrglualaservalvalalapro
450455460
proprovalvalleuleuproaspvalgluthrproserglugluasp
465470475480
cysmetpheglyasnglylysglytyrargglylysargalathrthr
485490495
valthrglythrprocysglnasptrpalaalaglngluprohisarg
500505510
hisserilephethrprogluthrasnproargalaglyleuglulys
515520525
asntyrcysargasnproaspglyaspvalglyglyprotrpcystyr
530535540
thrthrasnproarglysleutyrasptyrcysaspvalproglncys
545550555560
alaalaproserpheaspcysglylysproglnvalgluprolyslys
565570575
cysproglyargvalvalglyglycysvalalahisprohissertrp
580585590
protrpglnvalserleuargthrargpheglymethisphecysgly
595600605
glythrleuileserproglutrpvalleuthralaalahiscysleu
610615620
glulysserproargprosersertyrlysvalileleuglyalahis
625630635640
glngluvalasnleugluprohisvalglngluilegluvalserarg
645650655
leupheleugluprothrarglysaspilealaleuleulysleuser
660665670
serproalavalilethrasplysvalileproalacysleuproser
675680685
proasntyrvalvalalaaspargthrglucyspheilethrglytrp
690695700
glygluthrglnglythrpheglyalaglyleuleulysglualagln
705710715720
leuprovalilegluasnlysvalcysasnargtyrglupheleuasn
725730735
glyargvalglnserthrgluleucysalaglyhisleualaglygly
740745750
thraspsercysglnglyaspserglyglyproleuvalcyspheglu
755760765
lysasplystyrileleuglnglyvalthrsertrpglyleuglycys
770775780
alaargproasnlysproglyvaltyrvalargvalserargpheval
785790795800
thrtrpilegluglyvalmetargasnasn
805810
<210>5
<211>2145
<212>dna
<213>lys77-plg(lys-纤溶酶原)核酸序列
<400>5
aaagtgtatctctcagagtgcaagactgggaatggaaagaactacagagggacgatgtcc60
aaaacaaaaaatggcatcacctgtcaaaaatggagttccacttctccccacagacctaga120
ttctcacctgctacacacccctcagagggactggaggagaactactgcaggaatccagac180
aacgatccgcaggggccctggtgctatactactgatccagaaaagagatatgactactgc240
gacattcttgagtgtgaagaggaatgtatgcattgcagtggagaaaactatgacggcaaa300
atttccaagaccatgtctggactggaatgccaggcctgggactctcagagcccacacgct360
catggatacattccttccaaatttccaaacaagaacctgaagaagaattactgtcgtaac420
cccgatagggagctgcggccttggtgtttcaccaccgaccccaacaagcgctgggaactt480
tgtgacatcccccgctgcacaacacctccaccatcttctggtcccacctaccagtgtctg540
aagggaacaggtgaaaactatcgcgggaatgtggctgttaccgtgtccgggcacacctgt600
cagcactggagtgcacagacccctcacacacataacaggacaccagaaaacttcccctgc660
aaaaatttggatgaaaactactgccgcaatcctgacggaaaaagggccccatggtgccat720
acaaccaacagccaagtgcggtgggagtactgtaagataccgtcctgtgactcctcccca780
gtatccacggaacaattggctcccacagcaccacctgagctaacccctgtggtccaggac840
tgctaccatggtgatggacagagctaccgaggcacatcctccaccaccaccacaggaaag900
aagtgtcagtcttggtcatctatgacaccacaccggcaccagaagaccccagaaaactac960
ccaaatgctggcctgacaatgaactactgcaggaatccagatgccgataaaggcccctgg1020
tgttttaccacagaccccagcgtcaggtgggagtactgcaacctgaaaaaatgctcagga1080
acagaagcgagtgttgtagcacctccgcctgttgtcctgcttccagatgtagagactcct1140
tccgaagaagactgtatgtttgggaatgggaaaggataccgaggcaagagggcgaccact1200
gttactgggacgccatgccaggactgggctgcccaggagccccatagacacagcattttc1260
actccagagacaaatccacgggcgggtctggaaaaaaattactgccgtaaccctgatggt1320
gatgtaggtggtccctggtgctacacgacaaatccaagaaaactttacgactactgtgat1380
gtccctcagtgtgcggccccttcatttgattgtgggaagcctcaagtggagccgaagaaa1440
tgtcctggaagggttgtaggggggtgtgtggcccacccacattcctggccctggcaagtc1500
agtcttagaacaaggtttggaatgcacttctgtggaggcaccttgatatccccagagtgg1560
gtgttgactgctgcccactgcttggagaagtccccaaggccttcatcctacaaggtcatc1620
ctgggtgcacaccaagaagtgaatctcgaaccgcatgttcaggaaatagaagtgtctagg1680
ctgttcttggagcccacacgaaaagatattgccttgctaaagctaagcagtcctgccgtc1740
atcactgacaaagtaatcccagcttgtctgccatccccaaattatgtggtcgctgaccgg1800
accgaatgtttcatcactggctggggagaaacccaaggtacttttggagctggccttctc1860
aaggaagcccagctccctgtgattgagaataaagtgtgcaatcgctatgagtttctgaat1920
ggaagagtccaatccaccgaactctgtgctgggcatttggccggaggcactgacagttgc1980
cagggtgacagtggaggtcctctggtttgcttcgagaaggacaaatacattttacaagga2040
gtcacttcttggggtcttggctgtgcacgccccaataagcctggtgtctatgttcgtgtt2100
tcaaggtttgttacttggattgagggagtgatgagaaataattaa2145
<210>6
<211>714
<212>prt
<213>lys77-plg(lys-纤溶酶原)氨基酸序列
<400>6
lysvaltyrleuserglucyslysthrglyasnglylysasntyrarg
151015
glythrmetserlysthrlysasnglyilethrcysglnlystrpser
202530
serthrserprohisargproargpheserproalathrhisproser
354045
gluglyleuglugluasntyrcysargasnproaspasnaspprogln
505560
glyprotrpcystyrthrthraspproglulysargtyrasptyrcys
65707580
aspileleuglucysglugluglucysmethiscysserglygluasn
859095
tyraspglylysileserlysthrmetserglyleuglucysglnala
100105110
trpaspserglnserprohisalahisglytyrileproserlysphe
115120125
proasnlysasnleulyslysasntyrcysargasnproaspargglu
130135140
leuargprotrpcysphethrthraspproasnlysargtrpgluleu
145150155160
cysaspileproargcysthrthrproproproserserglyprothr
165170175
tyrglncysleulysglythrglygluasntyrargglyasnvalala
180185190
valthrvalserglyhisthrcysglnhistrpseralaglnthrpro
195200205
histhrhisasnargthrprogluasnpheprocyslysasnleuasp
210215220
gluasntyrcysargasnproaspglylysargalaprotrpcyshis
225230235240
thrthrasnserglnvalargtrpglutyrcyslysileprosercys
245250255
aspserserprovalserthrgluglnleualaprothralapropro
260265270
gluleuthrprovalvalglnaspcystyrhisglyaspglyglnser
275280285
tyrargglythrserserthrthrthrthrglylyslyscysglnser
290295300
trpsersermetthrprohisarghisglnlysthrprogluasntyr
305310315320
proasnalaglyleuthrmetasntyrcysargasnproaspalaasp
325330335
lysglyprotrpcysphethrthraspproservalargtrpglutyr
340345350
cysasnleulyslyscysserglythrglualaservalvalalapro
355360365
proprovalvalleuleuproaspvalgluthrproserglugluasp
370375380
cysmetpheglyasnglylysglytyrargglylysargalathrthr
385390395400
valthrglythrprocysglnasptrpalaalaglngluprohisarg
405410415
hisserilephethrprogluthrasnproargalaglyleuglulys
420425430
asntyrcysargasnproaspglyaspvalglyglyprotrpcystyr
435440445
thrthrasnproarglysleutyrasptyrcysaspvalproglncys
450455460
alaalaproserpheaspcysglylysproglnvalgluprolyslys
465470475480
cysproglyargvalvalglyglycysvalalahisprohissertrp
485490495
protrpglnvalserleuargthrargpheglymethisphecysgly
500505510
glythrleuileserproglutrpvalleuthralaalahiscysleu
515520525
glulysserproargprosersertyrlysvalileleuglyalahis
530535540
glngluvalasnleugluprohisvalglngluilegluvalserarg
545550555560
leupheleugluprothrarglysaspilealaleuleulysleuser
565570575
serproalavalilethrasplysvalileproalacysleuproser
580585590
proasntyrvalvalalaaspargthrglucyspheilethrglytrp
595600605
glygluthrglnglythrpheglyalaglyleuleulysglualagln
610615620
leuprovalilegluasnlysvalcysasnargtyrglupheleuasn
625630635640
glyargvalglnserthrgluleucysalaglyhisleualaglygly
645650655
thraspsercysglnglyaspserglyglyproleuvalcyspheglu
660665670
lysasplystyrileleuglnglyvalthrsertrpglyleuglycys
675680685
alaargproasnlysproglyvaltyrvalargvalserargpheval
690695700
thrtrpilegluglyvalmetargasnasn
705710
<210>7
<211>1245
<212>dna
<213>delta-plg(delta-纤溶酶原)核酸序列
<400>7
gagcctctggatgactatgtgaatacccagggggcttcactgttcagtgtcactaagaag60
cagctgggagcaggaagtatagaagaatgtgcagcaaaatgtgaggaggacgaagaattc120
acctgcagggcattccaatatcacagtaaagagcaacaatgtgtgataatggctgaaaac180
aggaagtcctccataatcattaggatgagagatgtagttttatttgaaaagaaagtgtat240
ctctcagagtgcaagactgggaatggaaagaactacagagggacgatgtccaaaacaaaa300
aatggcatcacctgtcaaaaatggagttccacttctccccacagacctagattctcacct360
gctacacacccctcagagggactggaggagaactactgcaggaatccagacaacgatccg420
caggggccctggtgctatactactgatccagaaaagagatatgactactgcgacattctt480
gagtgtgaagaggcggccccttcatttgattgtgggaagcctcaagtggagccgaagaaa540
tgtcctggaagggttgtaggggggtgtgtggcccacccacattcctggccctggcaagtc600
agtcttagaacaaggtttggaatgcacttctgtggaggcaccttgatatccccagagtgg660
gtgttgactgctgcccactgcttggagaagtccccaaggccttcatcctacaaggtcatc720
ctgggtgcacaccaagaagtgaatctcgaaccgcatgttcaggaaatagaagtgtctagg780
ctgttcttggagcccacacgaaaagatattgccttgctaaagctaagcagtcctgccgtc840
atcactgacaaagtaatcccagcttgtctgccatccccaaattatgtggtcgctgaccgg900
accgaatgtttcatcactggctggggagaaacccaaggtacttttggagctggccttctc960
aaggaagcccagctccctgtgattgagaataaagtgtgcaatcgctatgagtttctgaat1020
ggaagagtccaatccaccgaactctgtgctgggcatttggccggaggcactgacagttgc1080
cagggtgacagtggaggtcctctggtttgcttcgagaaggacaaatacattttacaagga1140
gtcacttcttggggtcttggctgtgcacgccccaataagcctggtgtctatgttcgtgtt1200
tcaaggtttgttacttggattgagggagtgatgagaaataattaa1245
<210>8
<211>414
<212>prt
<213>delta-plg(delta-纤溶酶原)氨基酸序列
<400>8
gluproleuaspasptyrvalasnthrglnglyalaserleupheser
151015
valthrlyslysglnleuglyalaglyserilegluglucysalaala
202530
lyscysglugluaspglugluphethrcysargalapheglntyrhis
354045
serlysgluglnglncysvalilemetalagluasnarglysserser
505560
ileileileargmetargaspvalvalleupheglulyslysvaltyr
65707580
leuserglucyslysthrglyasnglylysasntyrargglythrmet
859095
serlysthrlysasnglyilethrcysglnlystrpserserthrser
100105110
prohisargproargpheserproalathrhisprosergluglyleu
115120125
glugluasntyrcysargasnproaspasnaspproglnglyprotrp
130135140
cystyrthrthraspproglulysargtyrasptyrcysaspileleu
145150155160
glucysgluglualaalaproserpheaspcysglylysproglnval
165170175
gluprolyslyscysproglyargvalvalglyglycysvalalahis
180185190
prohissertrpprotrpglnvalserleuargthrargpheglymet
195200205
hisphecysglyglythrleuileserproglutrpvalleuthrala
210215220
alahiscysleuglulysserproargprosersertyrlysvalile
225230235240
leuglyalahisglngluvalasnleugluprohisvalglngluile
245250255
gluvalserargleupheleugluprothrarglysaspilealaleu
260265270
leulysleuserserproalavalilethrasplysvalileproala
275280285
cysleuproserproasntyrvalvalalaaspargthrglucysphe
290295300
ilethrglytrpglygluthrglnglythrpheglyalaglyleuleu
305310315320
lysglualaglnleuprovalilegluasnlysvalcysasnargtyr
325330335
glupheleuasnglyargvalglnserthrgluleucysalaglyhis
340345350
leualaglyglythraspsercysglnglyaspserglyglyproleu
355360365
valcyspheglulysasplystyrileleuglnglyvalthrsertrp
370375380
glyleuglycysalaargproasnlysproglyvaltyrvalargval
385390395400
serargphevalthrtrpilegluglyvalmetargasnasn
405410
<210>9
<211>1104
<212>dna
<213>mini-plg(小纤维蛋白溶酶原)核酸序列
<400>9
gtcaggtgggagtactgcaacctgaaaaaatgctcaggaacagaagcgagtgttgtagca60
cctccgcctgttgtcctgcttccagatgtagagactccttccgaagaagactgtatgttt120
gggaatgggaaaggataccgaggcaagagggcgaccactgttactgggacgccatgccag180
gactgggctgcccaggagccccatagacacagcattttcactccagagacaaatccacgg240
gcgggtctggaaaaaaattactgccgtaaccctgatggtgatgtaggtggtccctggtgc300
tacacgacaaatccaagaaaactttacgactactgtgatgtccctcagtgtgcggcccct360
tcatttgattgtgggaagcctcaagtggagccgaagaaatgtcctggaagggttgtaggg420
gggtgtgtggcccacccacattcctggccctggcaagtcagtcttagaacaaggtttgga480
atgcacttctgtggaggcaccttgatatccccagagtgggtgttgactgctgcccactgc540
ttggagaagtccccaaggccttcatcctacaaggtcatcctgggtgcacaccaagaagtg600
aatctcgaaccgcatgttcaggaaatagaagtgtctaggctgttcttggagcccacacga660
aaagatattgccttgctaaagctaagcagtcctgccgtcatcactgacaaagtaatccca720
gcttgtctgccatccccaaattatgtggtcgctgaccggaccgaatgtttcatcactggc780
tggggagaaacccaaggtacttttggagctggccttctcaaggaagcccagctccctgtg840
attgagaataaagtgtgcaatcgctatgagtttctgaatggaagagtccaatccaccgaa900
ctctgtgctgggcatttggccggaggcactgacagttgccagggtgacagtggaggtcct960
ctggtttgcttcgagaaggacaaatacattttacaaggagtcacttcttggggtcttggc1020
tgtgcacgccccaataagcctggtgtctatgttcgtgtttcaaggtttgttacttggatt1080
gagggagtgatgagaaataattaa1104
<210>10
<211>367
<212>prt
<213>mini-plg(小纤维蛋白溶酶原)氨基酸序列
<400>10
valargtrpglutyrcysasnleulyslyscysserglythrgluala
151015
servalvalalaproproprovalvalleuleuproaspvalgluthr
202530
proserglugluaspcysmetpheglyasnglylysglytyrarggly
354045
lysargalathrthrvalthrglythrprocysglnasptrpalaala
505560
glngluprohisarghisserilephethrprogluthrasnproarg
65707580
alaglyleuglulysasntyrcysargasnproaspglyaspvalgly
859095
glyprotrpcystyrthrthrasnproarglysleutyrasptyrcys
100105110
aspvalproglncysalaalaproserpheaspcysglylysprogln
115120125
valgluprolyslyscysproglyargvalvalglyglycysvalala
130135140
hisprohissertrpprotrpglnvalserleuargthrargphegly
145150155160
methisphecysglyglythrleuileserproglutrpvalleuthr
165170175
alaalahiscysleuglulysserproargprosersertyrlysval
180185190
ileleuglyalahisglngluvalasnleugluprohisvalglnglu
195200205
ilegluvalserargleupheleugluprothrarglysaspileala
210215220
leuleulysleuserserproalavalilethrasplysvalilepro
225230235240
alacysleuproserproasntyrvalvalalaaspargthrglucys
245250255
pheilethrglytrpglygluthrglnglythrpheglyalaglyleu
260265270
leulysglualaglnleuprovalilegluasnlysvalcysasnarg
275280285
tyrglupheleuasnglyargvalglnserthrgluleucysalagly
290295300
hisleualaglyglythraspsercysglnglyaspserglyglypro
305310315320
leuvalcyspheglulysasplystyrileleuglnglyvalthrser
325330335
trpglyleuglycysalaargproasnlysproglyvaltyrvalarg
340345350
valserargphevalthrtrpilegluglyvalmetargasnasn
355360365
<210>11
<211>750
<212>dna
<213>micro-plg(微纤维蛋白溶酶原)核酸序列
<400>11
gccccttcatttgattgtgggaagcctcaagtggagccgaagaaatgtcctggaagggtt60
gtaggggggtgtgtggcccacccacattcctggccctggcaagtcagtcttagaacaagg120
tttggaatgcacttctgtggaggcaccttgatatccccagagtgggtgttgactgctgcc180
cactgcttggagaagtccccaaggccttcatcctacaaggtcatcctgggtgcacaccaa240
gaagtgaatctcgaaccgcatgttcaggaaatagaagtgtctaggctgttcttggagccc300
acacgaaaagatattgccttgctaaagctaagcagtcctgccgtcatcactgacaaagta360
atcccagcttgtctgccatccccaaattatgtggtcgctgaccggaccgaatgtttcatc420
actggctggggagaaacccaaggtacttttggagctggccttctcaaggaagcccagctc480
cctgtgattgagaataaagtgtgcaatcgctatgagtttctgaatggaagagtccaatcc540
accgaactctgtgctgggcatttggccggaggcactgacagttgccagggtgacagtgga600
ggtcctctggtttgcttcgagaaggacaaatacattttacaaggagtcacttcttggggt660
cttggctgtgcacgccccaataagcctggtgtctatgttcgtgtttcaaggtttgttact720
tggattgagggagtgatgagaaataattaa750
<210>12
<211>249
<212>prt
<213>micro-plg(微纤维蛋白溶酶原)氨基酸序列
<400>12
alaproserpheaspcysglylysproglnvalgluprolyslyscys
151015
proglyargvalvalglyglycysvalalahisprohissertrppro
202530
trpglnvalserleuargthrargpheglymethisphecysglygly
354045
thrleuileserproglutrpvalleuthralaalahiscysleuglu
505560
lysserproargprosersertyrlysvalileleuglyalahisgln
65707580
gluvalasnleugluprohisvalglngluilegluvalserargleu
859095
pheleugluprothrarglysaspilealaleuleulysleuserser
100105110
proalavalilethrasplysvalileproalacysleuproserpro
115120125
asntyrvalvalalaaspargthrglucyspheilethrglytrpgly
130135140
gluthrglnglythrpheglyalaglyleuleulysglualaglnleu
145150155160
provalilegluasnlysvalcysasnargtyrglupheleuasngly
165170175
argvalglnserthrgluleucysalaglyhisleualaglyglythr
180185190
aspsercysglnglyaspserglyglyproleuvalcyspheglulys
195200205
asplystyrileleuglnglyvalthrsertrpglyleuglycysala
210215220
argproasnlysproglyvaltyrvalargvalserargphevalthr
225230235240
trpilegluglyvalmetargasnasn
245
<210>13
<211>684
<212>dna
<213>丝氨酸蛋白酶(结构)域的核酸序列
<400>13
gttgtaggggggtgtgtggcccacccacattcctggccctggcaagtcagtcttagaaca60
aggtttggaatgcacttctgtggaggcaccttgatatccccagagtgggtgttgactgct120
gcccactgcttggagaagtccccaaggccttcatcctacaaggtcatcctgggtgcacac180
caagaagtgaatctcgaaccgcatgttcaggaaatagaagtgtctaggctgttcttggag240
cccacacgaaaagatattgccttgctaaagctaagcagtcctgccgtcatcactgacaaa300
gtaatcccagcttgtctgccatccccaaattatgtggtcgctgaccggaccgaatgtttc360
atcactggctggggagaaacccaaggtacttttggagctggccttctcaaggaagcccag420
ctccctgtgattgagaataaagtgtgcaatcgctatgagtttctgaatggaagagtccaa480
tccaccgaactctgtgctgggcatttggccggaggcactgacagttgccagggtgacagt540
ggaggtcctctggtttgcttcgagaaggacaaatacattttacaaggagtcacttcttgg600
ggtcttggctgtgcacgccccaataagcctggtgtctatgttcgtgtttcaaggtttgtt660
acttggattgagggagtgatgaga684
<210>14
<211>228
<212>prt
<213>丝氨酸蛋白酶(结构)域的氨基酸序列
<400>14
valvalglyglycysvalalahisprohissertrpprotrpglnval
151015
serleuargthrargpheglymethisphecysglyglythrleuile
202530
serproglutrpvalleuthralaalahiscysleuglulysserpro
354045
argprosersertyrlysvalileleuglyalahisglngluvalasn
505560
leugluprohisvalglngluilegluvalserargleupheleuglu
65707580
prothrarglysaspilealaleuleulysleuserserproalaval
859095
ilethrasplysvalileproalacysleuproserproasntyrval
100105110
valalaaspargthrglucyspheilethrglytrpglygluthrgln
115120125
glythrpheglyalaglyleuleulysglualaglnleuprovalile
130135140
gluasnlysvalcysasnargtyrglupheleuasnglyargvalgln
145150155160
serthrgluleucysalaglyhisleualaglyglythraspsercys
165170175
glnglyaspserglyglyproleuvalcyspheglulysasplystyr
180185190
ileleuglnglyvalthrsertrpglyleuglycysalaargproasn
195200205
lysproglyvaltyrvalargvalserargphevalthrtrpileglu
210215220
glyvalmetarg
225
- 还没有人留言评论。精彩留言会获得点赞!