一种酸枣仁皂苷抑制间充质干细胞氧化损伤并增强其免疫调节能力的用途
1.本发明涉及干细胞领域,具体涉及一种可以抑制间充质干细胞氧化损伤和增强其免疫调节能力的物质及其应用。
背景技术:
2.间充质干细胞(mesenchymal stem cells,mscs)是一类来源于中胚层的成体干细胞,具有较强的自我复制能力,在合适的条件下可以被诱导分化成脂肪细胞、骨细胞和软骨细胞。目前,间充质干细胞已被临床用于治疗多种自身免疫性疾病并取得了一定的疗效。但有证据表明,在病理条件下,mscs仍面临着免疫调节功能受损、存活困难等问题。因此寻找一种解决此类问题的新方法将有助于进一步提升mscs的临床疗效。通过对现有研究及专利的检索发现,尽管现有技术能解决一定的问题,但仍存在一定的弊端。
3.如专利cn111437289a表明,使用甘露糖预处理脐带mscs后,可增强其抑制外周血单个核细胞(peripheral blood mononuclear cells,pbmc)异常增殖的作用,并促进转化生长因子
‑
β、前列腺素e2的分泌。mansoureh togha等人发现,雷帕霉素联合骨髓mscs能更有效的削弱cd8+t淋巴细胞的细胞毒性,抑制干扰素γ的分泌,促进白介素4的合成。shyamkishor sah等人发现,高表达sod3的脐带mscs具有更强的免疫调节能力,能更有效的抑制淋巴细胞的增殖和向皮肤的炎性浸润。尽管上述方案已解决了部分问题,但仍存在一定的缺陷与不足。如:(1)所添加的成分价格昂贵、临床推广难度大。(2)所添加的成分仅具有增强mscs免疫调节能力或抑制mscs氧化损伤能力,未考虑临床使用的实际情况。(3)具体操作过程中涉及外源基因的导入,可能引发未知的风险。(4)所添加的成分属于罕见物质,患者临床使用时依从性差。(5)所选用的培养添加剂未经过毒性试验的验证,是否存在细胞毒性仍未可知。
4.另外,有研究表明,部分中药活性单体也具有促进mscs增殖、抑制其凋亡、增强其免疫调节能力的作用。如周春根等的研究表明,人参皂苷rg1对mscs具有促进增殖、抑制凋亡、影响旁分泌与分化的作用。刘晓丹等发现黄芪甲苷配伍三七总皂苷能在体外促进缺血再灌注模型骨髓mscs的增殖、迁移,抑制其凋亡,并诱导其向神经元、星形胶质细胞定向分化。除此之外,还有报道表明部分中药活性成分如蛇床子素、槲皮素具有增强mscs免疫调节能力。但以上研究仍存在一定的缺陷,如(1)部分中药单体在扩增培养过程中,对mscs 成瘤性的影响不可知,存在一定的未知风险。(2)大部分中药单体主要影响mscs的分化能力,对提升mscs临床疗效的意义不大。(3)蛇床子素、槲皮素对mscs的作用单一,难以同时解决临床应用时所面临的免疫调节功能受损与存活困难的问题。
5.综上所述,针对现有发明或报道中所存在的缺陷,我们致力于提供一种安全有效、价格低廉、便于临床推广的增强mscs免疫调节能力并保护其免受氧化损伤的新方法。有利于进一步提升mscs的临床疗效,故而对促进同种异体间充质干细胞临床应用于自身免疫性疾病和炎性疾病的治疗具有重要意义。
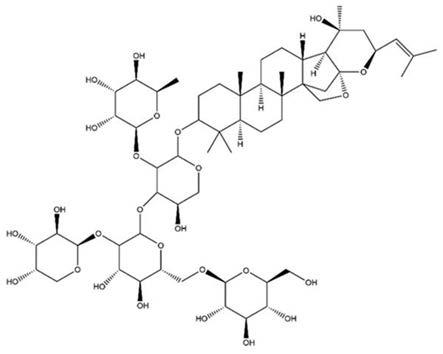
6.酸枣仁皂苷a是中药酸枣仁的主要活性成分,酸枣仁临床主要用于治疗失眠多梦、头晕健忘,已上市的产品有酸枣仁合剂、枣仁安神液等。本发明首次发现了酸枣仁皂苷a还具有抑制间充质干细胞氧化应激损伤并增强其免疫调节能力的作用。
技术实现要素:
7.针对上述问题,本发明的目的在于提供安全有效、价格低廉、临床易推广的一种新方法。具体是提供一种酸枣仁皂苷的新用途,特别是酸枣仁皂苷a在抑制间充质干细胞氧化应激损伤和增强其免疫调节能力两方面的新用途,更加贴合mscs临床使用时的实际情况,有助于临床疗效的发挥。
8.为了实现上述目的,本发明提供了如下技术方案:
9.本发明提供了一种酸枣仁皂苷a在抑制间充质干细胞氧化损伤和增强其免疫调节能力方面的应用。
10.进一步地,所述间充质干细胞包括脐带间充质干细胞、脂肪间充质干细胞、骨髓间充质干细胞、滑膜间充质干细胞。
11.进一步地,所述酸枣仁皂苷a的剂型包括于片剂、散剂、颗粒剂、胶囊剂、混悬剂、注射剂、气雾剂、粉雾剂、洗剂、搽剂、软膏剂。
12.进一步地,通过酸枣仁皂苷a促进间充质干细胞的存活,降低间充质干细胞乳酸脱氢酶和丙二醛的含量,降低间充质干细胞cleaved
‑
caspase3蛋白的表达,上调间充质干细胞 bcl
‑
2/bax的蛋白比值来实现抑制间充质干细胞氧化损伤。
13.进一步地,通过酸枣仁皂苷a增强间充质干细胞对外周血单个核细胞异常激活的抑制,不改变间充质干细胞对t淋巴细胞亚型分化的调节来实现增强间充质干细胞免疫调节能力。
14.与现有技术相比本发明的有益效果。
15.(1)安全性高:酸枣仁皂苷a来源于中药酸枣仁,而酸枣仁则是鼠李科植物酸枣的干燥成熟种子,生活中常见,属于日常食用范畴,并且相关研究也表明酸枣仁醇提物安全无毒,本发明实验结果也证明其不具有细胞毒性,因此酸枣仁皂苷a具有较高的安全性;另外,本发明在具体实施方式中不涉及基因修饰的操作,故而不存在未知因素与不可控风险。
16.(2)依从性好:中药作为中国传统医学的瑰宝流传至今,在国内拥有广泛的拥护者,而酸枣仁更是在日常生活中较为常见,临床上普遍使用,所以在使用基于酸枣仁皂苷a的 mscs细胞疗法时,有利于提高患者的依从性。
17.(3)价格低廉:酸枣仁皂苷a在酸枣仁中含量较高,目前提取工艺已成熟完备,且成品易于储存于运输;与其他培养添加剂或操作方案相比价格实惠,临床上更利于推广应用。
18.(4)效果突出:酸枣仁皂苷a对mscs具有多重生物活性,既可以抑制机体炎症所导致的mscs氧化应激损伤,又可以增强mscs的免疫调节能力,而现有技术对mscs往往只表现出单一的药理作用。
19.(5)与其他中药活性成分相比的有益效果:
①
酸枣仁皂苷a对mscs的增殖能力没有显著影响,不会提升mscs的成瘤风险。
②
酸枣仁皂苷a能显著增强mscs的免疫调节能力,有利于进一步提升临床疗效;另外还可以抑制mscs氧化应激损伤,更有利于解决其临床应用时所面临的困难。
附图说明
20.图1 jua化学结构式
21.图2 jua对huc
‑
mscs增殖能力的影响
22.图3 h2o2造模浓度的选择
23.图4 jua对h2o2诱导的huc
‑
mscs存活率的影响
24.图5jua对h2o2诱导的huc
‑
mscs ldh释放量的影响
25.图6 jua对h2o2诱导的huc
‑
mscs mda含量的影响
26.图7 jua对h2o2诱导的huc
‑
mscs凋亡的影响
27.图8 jua对huc
‑
mscs抑制pbmc异常增殖能力的影响
28.图9 jua对huc
‑
mscs调节t淋巴细胞亚型分化能力的影响
具体实施方式
29.下面结合具体实施例对本发明做详细的说明。为了阐明本发明所述的jua抑制间充质干细胞氧化应激损伤并增强其免疫调节能力的应用,提供以下具体的实施方式并加以说明,但不限于这些实施方式。
30.一、实验材料l
‑
dmem培养基(gibco)、rpmi1640培养基(gibco)、胎牛血清(四季青)、edta
‑
0.25%胰蛋白酶(gibco)青
‑
链双抗(hyclone)、cck
‑
8试剂盒(美仑)、ldh测定试剂盒(万类)、 mda测定试剂盒(万类)、bax、bcl
‑
2、caspase3一抗(万类)、山羊抗兔igg二抗(博奥森)、 cd4
‑
apc(bd)、ifn
‑
γ
‑
pe(southernbiotech)、cd4
‑
percp(bd)、il
‑
17a
‑
pe(bd)、cd4
‑
fitc (bd)cd25
‑
apc(bd)。酸枣仁皂苷a(cas号:55466
‑
04
‑
1),纯度≥98%,购自四川省维克奇生物科技有限公司。
31.二、实验方法
32.cck
‑
8法测定jua干预对huc
‑
mscs体外活性的影响取第3代huc
‑
mscs,使用edta
‑
0.25%胰酶消化并调整细胞密度为5
×
104cell/ml,接种于 96孔板。实验分为对照组、实验组,每组均设置6个平行复孔。实验组分别添加终浓度为6.25、 12.5、25、50、100μmol/l的jua进行干预。继续培养48h后,每孔加入10μl的cck
‑
8溶液,培养2h后用酶标仪检测其在450mm处的吸光度,并按照公式增殖率=(a实验
‑
a空白) /(a对照
‑
a空白)
×
100%计算各组huc
‑
mscs的增殖率。
33.h2o2损伤huc
‑
mscs浓度的筛选取第5代huc
‑
mscs,使用edta
‑
0.25%胰酶消化并调整细胞密度为5
×
104cell/ml,接种于 96孔板。实验分为对照组、模型组、实验组,每组均设置6个平行复孔。实验组分别添加终浓度为62.5、125、250、500、1000μmol/l的30%h2o2溶液进行干预。继续培养12h后,每孔加入10μl的cck
‑
8溶液,培养2h后用酶标仪检测其在450mm处的吸光度,并计算各组huc
‑
mscs的存活率。
34.cck
‑
8法检测jua对h2o2损伤的huc
‑
mscs存活率的影响取第5代huc
‑
mscs,使用edta
‑
0.25%胰酶消化并调整细胞密度为5
×
104cell/ml,接种于 96孔板,每孔100μl。实验分为对照组、模型组、实验组均设置6个平行复孔。实验组分别添加终浓度为12.5、25、50μmol/l的jua溶液进行干预。继续培养48h后,每孔加入终浓
度为250μmol/l 30%h2o2溶液,培养12h后,再每孔加入10μl的cck
‑
8溶液,2h后使用用酶标仪检测其在450mm处的吸光度,并计算各组huc
‑
mscs的存活率。
35.微量酶标法检测jua对h2o2损伤的huc
‑
mscs ldh释放量的影响取第5代huc
‑
mscs,使用edta
‑
0.25%胰酶消化并调整细胞密度为5
×
104cell/ml,接种于96孔板,每孔100μl。实验分为对照组、模型组和实验组,每组均设置6个平行复孔。实验组分别添加终浓度为12.5、25、50μmol/l的jua溶液进行干预。继续培养48h后,每孔加入终浓度为250μmol/l 30%h2o2溶液,培养12h后,取上清液,过滤,并按照试剂盒说明书进行操作。
36.tba法检测jua对h2o2损伤的huc
‑
mscs mda含量的影响取第5代huc
‑
mscs,使用edta
‑
0.25%胰酶消化并调整细胞密度为5
×
104cell/ml,接种于6 孔板,每孔1ml。实验分为对照组、模型组和实验组,每组均设置3个平行复孔。实验组分别添加终浓度为12.5、25、50μmol/l的jua溶液进行干预。继续培养48h后,每孔加入终浓度为250μmol/l 30%h2o2溶液,培养12h后,细胞刮板刮取细胞,并按照试剂盒说明书进行操作。
37.westernblot法检测jua对h2o2损伤的huc
‑
mscs凋亡相关蛋白表达的影响取第5代huc
‑
mscs,使用edta
‑
0.25%胰酶消化并调整细胞密度为5
×
104cell/ml,接种于6 孔板,每孔1ml。实验分为对照组、模型组和实验组,每组均设置3个平行复孔。实验组分别添加终浓度为12.5、25、50μmol/l的jua溶液进行干预。继续培养48h后,每孔加入终浓度为250μmol/l 30%h2o2溶液,培养12h后,提取细胞总蛋白,经bca法测定蛋白浓度后制备上样缓冲液后进行sds
‑
page凝胶电泳。转膜后利用脱脂牛奶封闭2h,孵育一抗,4℃过夜,洗膜后孵育二抗,并使用凝胶成像系统显影,imagej用来计算相对灰度值。
38.cck
‑
8法检测jua对huc
‑
mscs抑制pbmc异常增殖能力的影响取第5代huc
‑
mscs,使用edta
‑
0.25%胰酶消化并调整细胞密度为1
×
105cell/ml,接种于 96孔板。培养过夜贴壁,以huc
‑
mscs与pbmcs为1∶10的比例加入pbmcs(1
×
104cells) 混合共培养,以植物血凝素为刺激剂,终浓度为10μg/ml,继续培养3d。实验分为pbmcs 组、pbmcs+huc
‑
mscs、pbmcs+huc
‑
mscs+jua组、pbmc+jua组。随后每孔加入10μl 的cck
‑
8溶液,2h后使用用酶标仪检测其在450mm处的吸光度,计算细胞存活率。
39.流式细胞术检测jua对huc
‑
mscs调节t淋巴细胞亚群分化能力的影响取第5代huc
‑
mscs,使用edta
‑
0.25%胰酶消化并调整细胞密度为1
×
105cell/ml,接种于 24孔板,每孔1ml。培养过夜贴壁,以huc
‑
mscs与pbmcs为1∶10的比例加入pbmcs (1
×
106cells)混合共培养,以植物血凝素为刺激剂,终浓度为10μg/ml,继续培养3d。实验分为pbmcs单独培养组、pbmcs+huc
‑
mscs、pbmcs+huc
‑
mscs+jua共培养组。收集 pbmcs,分别加入cd4
‑
apc,ifn
‑
γ
‑
pe抗体各5μl、cd4
‑
percp,il
‑
17a
‑
pe抗体各5μl 及cd4
‑
fitc,cd25
‑
apc抗体各5μl,避光孵育10min,经1500rpm/min离心3min,pbs 洗涤2次,加入500μl pbs悬浮细胞,流式细胞仪上机检测,th1水平以cd4+ifn
‑
γ+表示, th17水平以cd4+il
‑
17a+表示,treg水平以cd4+cd25+表示。
40.统计学分析使用spss 22.0软件统计实验结果,并对实验结果进行t检验和单因素方差分析,以表示。 p<0.05差异有统计学意义。用graph pad prism 5绘制统计图。
41.三、实验结果。
42.jua不影响huc
‑
mscs体外活性为了探讨jua对huc
‑
mscs体外活性的影响,我们选用cck
‑
8法进行检测,结果如表1与附图2所示,各实验组与对照组相比细胞活率均无明显变化,不存在统计学差异(p>0.05),说明终浓度为6.25~100μmol/l的jua均对huc
‑
mscs的体外活性无影响。以上结果证明, 6.25~100μmol/l的jua对huc
‑
mscs无毒性作用。
43.表1细胞活率统计表表1细胞活率统计表
44.成功建立huc
‑
mscs氧化损伤模型病理条件下,机体内炎性细胞因子会激发炎症细胞释放大量的活性氧自由基,从而导致 huc
‑
mscs的氧化应激损伤。为了更好的模拟这一现象,我们决定使用h2o2在体外构建 huc
‑
mscs氧化损伤模型。实验结果如表2和附图3所示,125~1000μmol/l 30%h2o2均可以造成huc
‑
mscs存活率的明显下降,与对照组相比,差异具有显著性(
*
p<0.05,
**
p<0.01)。其中,125μmol/l 30%h2o2组损伤不足,而250、500μmol/l30%h2o2组存活率均不足40%,损伤过强,故选择250μmol/l 30%h2o2作用12h为后续实验的造模条件。
45.表2细胞存活率统计表表2细胞存活率统计表表2细胞存活率统计表
46.jua促进h2o2损伤的huc
‑
mscs存活为了进一步的探究jua对h2o2损伤的huc
‑
mscs存活的影响,我们使用cck
‑
8法检测了各组细胞的吸光度值,并计算存活率。结果显示,模型组细胞的存活率明显下降,与对照组相比具有统计学差异(
##
p<0.01),证明huc
‑
mscs氧化损伤模型构建成功。而经过不同浓
度的jua治疗后,各组细胞的存活率均有不同程度的上升,与模型组相比差异具有显著性(
*
p <0.05,
**
p<0.01);并且各组细胞的存活率随着jua浓度的增大而提高,呈剂量依赖性。见表3和附图4。以上结果说明,jua能有效抑制h2o2对huc
‑
mscs损伤,提升其细胞存活率。
47.表3细胞存活率统计表表3细胞存活率统计表
48.jua抑制h2o2损伤的huc
‑
mscs ldh的释放细胞的凋亡或坏死会造成细胞膜结构的破坏,从而导致胞浆中的酶释放到培养液中,ldh就是其中较为稳定的一种,通过检测细胞培养液中的ldh含量可以反映细胞膜的完整性。检测结果显示,模型组细胞培养液中ldh的含量明显上升,与对照组相比具有显著性(
##
p<0.01),表明模型组细胞细胞膜结构破坏严重,造模成功。而经过不同浓度的jua治疗后,各组细胞培养液中ldh的含量均有不同浓度的降低,其中jua
‑
m组、jua
‑
h组与模型组存在统计学差异(
*
p<0.05,
**
p<0.01)。见表4和附图5。综上,jua能有效降低h2o2对huc
‑
mscs细胞膜结构的破坏,减少ldh的释放量。
49.表4 ldh释放量统计表量统计表
50.jua减少h2o2损伤的huc
‑
mscs mda含量mda是生物体内脂质氧化的终产物,影响线粒体呼吸链复合物及线粒体内关键酶活性,因此可以检测胞内mda的含量反映机体脂质过氧化的程度。实验结果如表5和附图6所示,模型组细胞mda含量较对照组明显升高,差异具有显著性(
##
p<0.01),表明h2o2能促进 huc
‑
mscs的脂质过氧化,模型构建成功。而经过中、高浓度的jua治疗后,细胞中mda 的含量明显减少,与模型组之间的差异均具有统计学意义(
*
p<0.05,
**
p<0.01)。以上结果说明,jua能明显抑制h2o2诱导的huc
‑
mscs脂质过氧化,减少mda的含量。
51.表5 mda含量统计表
52.jua下调caspase3、bax蛋白的表达,上调bcl
‑
2蛋白的表达caspase家族在介导细胞凋亡的过程中具有重要作用,其中以caspase3为关键的执行分子。caspase3通常以酶原的形式,在凋亡的早期阶段,被激活成cleaved
‑
caspase3裂解parp,最终导致细胞凋亡。bax与bcl
‑
2同属于bcl
‑
2家族;其中,bax是凋亡前体蛋白,可以促进细胞凋亡,而bcl
‑
2则可以抑制多种细胞毒因素引起的细胞死亡。为了探讨jua对h2o2损伤的 huc
‑
mscs凋亡的影响,我们检测了凋亡相关蛋白cleaved
‑
caspase3、bax、bcl
‑
2的表达情况。实验结果如附图7所示,模型组细胞cleaved
‑
caspase3蛋白表达明显上升,bcl
‑
2/bax比例明显下降,与对照组相比均存在显著差异(
##
p<0.01),证明h2o2损伤后,huc
‑
mscs凋亡明显增多,模型建立成功。而经过中、高浓度的jua治疗后,cleaved
‑
caspase3蛋白表达量明显下降,bcl
‑
2/bax比例显著上调,与模型组相比均具有统计学意义(
*
p<0.05,
**
p<0.01)。综上所述,jua可以有效改善h2o2损伤后huc
‑
mscs的凋亡情况。
53.jua增强huc
‑
mscs对pbmc异常增殖的抑制pbmc是指外周血中具有单个核的细胞,包括淋巴细胞和单核细胞。我们使用植物血凝素 (phytohemagglutinin,pha)来激活pbmc,使其表现为异常活化,用来模拟机体内免疫的过度激活。检测结果显示,huc
‑
mscs组和huc
‑
mscs+jua组的细胞活率明显下降,与pbmc 组相比均存在显著性差异(
##
p<0.01),而pbmc+jua组与pbmc组相比则不存在显著性差异,证明jua对pbmc的异常增殖没有明显的抑制作用。另外,huc
‑
mscs+jua组较 huc
‑
mscs组细胞活率明显下降,差异具有统计学意义(
*
p<0.05),证明jua可以增强 huc
‑
mscs对pbmc异常增殖的抑制作用。见附图8。
54.jua不改变huc
‑
mscs调节t淋巴细胞亚群分化的能力辅助性t细胞1(t helper cell 1,th1)和辅助性t细胞17(t helper cell 1,th17)同属于辅助性t细胞0分化的不同th细胞亚群;th1主要由il
‑
12诱导分化而来,分泌ifn
‑
γ并驱动刺激巨噬细胞对抗细胞内细菌及原虫的免疫反应;th17主要通过il
‑
6和tgf
‑
β诱导分化而来,主要分泌il
‑
17并驱动嗜中性粒细胞对抗细胞外细菌及霉菌的免疫反应。过分的th1、 th17激活会导致机体自身的免疫失衡,表现为自体免疫性疾病。调节性t细胞(regulatory tcells,tregs)是维持机体免疫平衡的重要因素之一,由胸腺产生后输出至外周,并通过主动调节的方式抑制存在于正常机体内潜在的自身反应性t细胞的活化与增殖,从而调节机体的免疫力,防止自身免疫性疾病的发生。
55.为了探讨jua对huc
‑
mscs调节t淋巴细胞亚群分化能力的影响,流式细胞术被用来检测th1、th17、treg的比例。实验结果如图9所所示,huc
‑
mscs组和huc
‑
mscs+jua组的向
th1、th17分化的比率明显下降,与pbmc组相比,均具有显著性差异(
*
p<0.05,
**
p <0.01);向treg分化的比率明显上升,与pbmc组之间的差异存在统计学意义(
*
p<0.05)。另外,huc
‑
mscs组与huc
‑
mscs+jua组在向th1、th17、treg分化的比率上均不存在明显差异,无统计学意义(p>0.05)。以上结果表明,jua不影响huc
‑
mscs对t淋巴细胞亚群分化的调节作用。
56.综上所述,本研究结果表明,jua具有抑制mscs氧化应激损伤并增强其免疫调节能力的作用,这有利于打破mscs临床使用时所面临的病理条件下存活困难和免疫调节能力受损的困境,进一步证明了在自身免疫性疾病和炎性疾病的临床治疗中使用无基因操作的同种异体mscs的可行性,同时也为开发jua的新用途提供实验基础。
57.上述实例用来介绍本发明的实质性内容,但不能以此限定本发明的保护范围。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1