Tel1在制备降低烟曲霉唑类耐药性的药物中的应用的制作方法
tel1在制备降低烟曲霉唑类耐药性的药物中的应用
技术领域
1.本发明属于生物医药领域,尤其涉及一种丝氨酸/苏氨酸蛋白激酶tel1在制备降低烟曲霉唑类耐药性的药物中的应用。
背景技术:
2.烟曲霉(aspergillusfumigatus)是最常见的曲霉属真菌,其分生孢子是环境空气标本中真菌的主要形态。人体从气道吸入的孢子若不能排出,可引起侵袭性曲霉病、慢性肺曲霉病、过敏性曲霉病和脑曲霉病等。由于细胞毒性药物、糖皮质激素、靶向药物、新冠肺炎流行、造血干细胞移植和器官移植等,侵袭性曲霉病的发病率逐年升高。在严重免疫功能低下的患者中,曲霉病已成为最常见的死亡原因之一,急性白血病和造血干细胞移植者患曲霉病的病死率高达40%~50%。
3.侵袭性曲霉病首选伏立康唑治疗,慢性肺曲霉病和过敏性曲霉病多用伊曲康唑和伏立康唑治疗。但是,近年来全世界烟曲霉对唑类抗真菌药物的敏感性显著下降。例如2004年英国曼彻斯特报道烟曲霉临床分离株唑类耐药率5%,2009年已达20%。目前中国和全球最常见的烟曲霉唑类耐药基因型为tr34/l98h突变型。
4.唑类抗真菌药与14-α-甾醇脱甲基酶(cyp51)结合,抑制烟曲霉细胞膜麦角甾醇的生物合成。cyp51a突变是烟曲霉临床株唑类耐药最常见的原因,我们与其他研究者发现cyp51a突变是通过修饰结合位点来影响唑类亲和力。非cyp51a突变所致唑类耐药的机制包括:(1)外排泵过度表达:afumdr4与伏立康唑耐药相关,cdr1b与伊曲康唑耐药相关;(2)cyp51过度表达:srba、hape和atrr缺失可导致唑类耐药;(3)应对氧化应激:烟曲霉yap1突变导致伏立康唑耐药;(4)线粒体参与:有证据表明线粒体复合体i可以消除唑类导致的活性氧释放对烟曲霉的毒性,从而提高对唑类药物的耐受。
5.生物膜是附着于物体表面,被胞外基质包裹的有组织的微生物群体,具有与游离状态时不同的组织成分。尽管以往的科学研究多以微生物在液态培养基中浮游生长或者固态培养基中菌落生长为基础,现在公认对于绝大多数微生物来说生物膜附着更能反映其自然生长状态。烟曲霉生物膜主要由菌丝骨架和胞外基质组成,形成过程可分为三个阶段:分生孢子萌发和粘附、孢子生成菌丝(10~12小时)、生物膜形成及成熟(48~72小时)。生物膜可以帮助真菌抵抗药物的作用,生物膜导致的耐药机制有:细胞外基质屏障、药物外排泵的过度表达、缺氧耐受、菌体细胞休眠和生物膜-宿主免疫系统相互作用等。
技术实现要素:
6.为克服现有技术的缺点和不足,本发明的目的在于提供一种丝氨酸/苏氨酸蛋白激酶tel1的抑制剂用于增强烟曲霉对唑类药物的敏感性,即做类药物的增效剂。
7.本发明是这样实现的,丝氨酸/苏氨酸蛋白激酶tel1在制备降低烟曲霉唑类耐药性的药物中的应用。
8.优选地,所述药物为有效剂量的抑制丝氨酸/苏氨酸蛋白激酶tel1/atm的抑制剂。
9.优选地,所述药物为tel1/atm抑制剂ku-60019。
10.优选地,所述药物包括药物学上可以接受的载体;所述载体为稀释剂、缓冲剂、混悬剂、乳剂、颗粒剂、包囊剂、赋形剂、填充剂、粘合剂、喷雾剂、透皮吸收剂、湿润剂、崩解剂、吸收促进剂、表面活性剂、着色剂、矫味剂或吸附载体。
11.本发明克服现有技术的不足,公开了tel1在制备降低烟曲霉唑类耐药性的药物中的应用。丝氨酸/苏氨酸蛋白激酶tel1在哺乳动物直系同源物(homolog ous protein)是共济失调毛细血管扩张突变蛋白激酶(ataxia-telangiectasia mutated protein kinases,atm蛋白激酶)。丝氨酸/苏氨酸蛋白激酶tel1/atm属于磷脂酰肌醇-3-激酶相关蛋白激酶(phosphatidylinositol3-kinase-related kinase,pikk)家族。atm基因于1995年在共济失调毛细血管扩张症(ataxia telangiectasia)的研究中首次得到克隆(a single ataxia telangiectasia gene witha product similar to pi-3kinase,science,268(1995)1749-1753.)。2000年tel1在酿酒酵母(saccharomyces cerevisiae)中被发现(j.c.mallory,t.d.petes,protein kinase activity of tel1p and mec1p,two saccharomyces cerevisiaeproteins related to the human atm protein kinase,proc natl acad sci u sa,97(2000)13749-13754.)。真菌tel1与人类atm基因高度同源,基因产物蛋白结构也高度保守和相似。
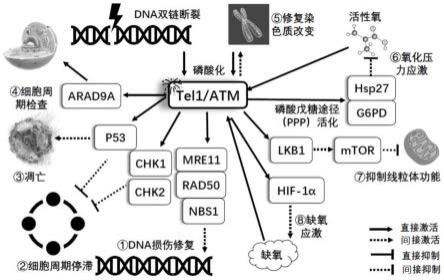
12.如图1所示,tel1/atm具有以下重要功能:(1)dna损伤修复:当dna损伤发生时,atm迅速被激活,与dna损伤部位的mrn(mre11/rad50/nbs1)复合物共信号传导,还激活了数百个传感器和效应器的dna损伤修复信号通路的级联,从而启动dna损伤修复反应;(2)细胞周期停滞:直接磷酸化并稳定p53,诱导细胞g1/s和g2/m周期停滞,防止未修复的dna突变传递到子代细胞中;(3)凋亡:当dna损伤太严重而无法有效修复时,激活的p53可诱导细胞凋亡;(4)细胞周期检查:激活细胞周期调控蛋白rad9a、检查点激酶(checkpoint kinase,chk)1和chk2等使细胞周期进展暂时停止,以实现dna损伤修复;(5)染色质改变修复:将染色质改变信号传导到chk2和p53等底物,继而活化多个通路以维持染色质完整性;(6)氧化压力应激;(7)抑制线粒体功能;和(8)缺氧应激。
13.atm抑制剂可抑制多种肿瘤耐药,因此,若干atm抑制剂被开发和用于肿瘤辅助治疗。比如,多形性胶质母细胞瘤(glioblastoma multiform)是最常见的脑实质肿瘤,患者通常在一年内死于该疾病。atm抑制剂ku-60019与放射治疗联合使用时,可显著诱导该肿瘤消退和提高动物存活率,此抑制剂已进入i期临床试验。
14.为了验证tel1/atm抑制剂ku-60019是否通过抑制真菌的tel1从而增强唑类药物抑菌效果,本发明将tel1/atm抑制剂ku-60019与烟曲霉和伊曲康唑共孵育,进行药敏实验。结果发现在抑制剂ku-60019作用下,伊曲康唑对烟曲霉标准株的最小抑菌浓度值(mic值)降低为无抑制剂时的1/4,耐药株伊曲康唑mic值降低到无抑制剂的1/2~1/8。实验结果证明,tel1/atm抑制剂ku-60019可明显增加烟曲霉对伊曲康唑敏感性,并且印证tel1参与了烟曲霉唑类耐药。
15.唑类药物可以导致烟曲霉线粒体活性氧泄露,靶向烟曲霉细胞膜,从而破坏氧化还原稳态。但是烟曲霉可通过抑制线粒体呼吸复合物i(mitochondrial complex i,nadh:ubiquinone oxidoreductase,泛醌氧化还原酶),消除升高的活性氧。烟曲霉唑类耐药与线粒体复合物i相关,线粒体复合物i能力丧失或抑制会导致烟曲霉对唑类耐药。tel1/atm激
活可抑制细胞线粒体功能,应对氧化应激。这也是人肿瘤atm上调,导致耐药的机制之一。
16.生物膜也是烟曲霉应对唑类的重要方法。烟曲霉生物膜在其成熟过程中,始终保持缺氧微环境的空间梯度。即使丝状真菌生物膜内菌丝之间有充足的氧气,线粒体耗氧还是会自我诱导出缺氧区。由于缺氧等原因,生物膜基部菌丝代谢状态降低。然而唑类(特别是伏立康唑)需要代谢活性才能有效地影响麦角甾醇合成,发挥抗真菌活。这些缺氧区的形成,不仅有助于烟曲霉生物膜生长,还有助于增加抗真菌药物耐药性。而tel1/atm激活有助于细胞应对缺氧,有助于真菌在生物膜中生长。tel1/atm应对缺氧在肿瘤耐药机制中发挥一定作用。
17.综合分析tel1/atm功能、人肿瘤atm上调导致耐药以及烟曲霉唑类耐药的机制后,判定烟曲霉唑类耐药株生物膜蛋白组学分析中tel1显著上调,是为了更好地应对唑类药物压力。具体表现为:
18.(1)烟曲霉唑类耐药株tel1上调可抑制线粒体功能和降低活性氧水平
19.唑类药物引起烟曲霉内源性活性氧增多,烟曲霉唑类耐药株在唑类压力下tel1表达明显上调,从而抑制线粒体和减少释放的活性氧,导致其生物膜抗药性进一步增强。在本发明的蛋白组学结果中,耐药株在伊曲康唑压力下,氧化应激相关蛋白过氧化物酶asp f3表达也都明显上调,也支持了本发明的假设。
20.(2)烟曲霉唑类耐药株tel1上调有助于适应生物膜缺氧微环境
21.烟曲霉生物膜在其成熟过程中会形成缺氧微环境,tel1通路激活有助于克服生物膜基底层菌丝的缺氧问题,从而更好地应对缺氧并发展耐药倾向。
22.相比于现有技术的缺点和不足,本发明具有以下有益效果:
23.(1)本发明首次发现了丝氨酸/苏氨酸蛋白激酶tel1与烟曲霉唑类耐药性存在特殊相关性,作为烟曲霉唑类耐药性的药物靶标,可应用于避免或者降低烟曲霉唑类耐药株耐药性的曲霉病治疗药物的制备和筛选;
24.(2)本发明进一步提供了一种抑制丝氨酸/苏氨酸蛋白激酶tel1/atm的抑制剂ku-60019,可明显增加烟曲霉对曲霉病药物伊曲康唑的敏感性。
附图说明
25.图1是是丝氨酸/苏氨酸蛋白激酶tel1/atm的主要信号通路和功能示意图。
具体实施方式
26.为了使本发明的目的、技术方案及优点更加清楚明白,以下结合附图及实施例,对本发明进行进一步详细说明。应当理解,此处所描述的具体实施例仅仅用以解释本发明,并不用于限定本发明。
27.1、伊曲康唑作用下烟曲霉生物膜中丝氨酸/苏氨酸蛋白激酶tel1的变化
28.1.1、抗真菌剂、菌株准备
29.本发明实施例中烟曲霉菌株和抗真菌剂的制备涉及两株烟曲霉(菌种来源详见:liu m,zeng r,zhang l,li d,lv g,shen y,zheng h,zhang q,zhao j,zheng n,liu w.multiple cyp51a-based mechanisms identified in azole-resistant isolates ofaspergillus fumigatus from china.antimicrob agents chemother.2015jul;59(7):
4321-5.doi:10.1128/aac.00003-15.epub 2015apr 20.pmid:25896700;pmcid:pmc4468671.),包括af293(野生型菌株)和shjt42b(具有tr34/l98h基因型cyp51a的唑类临床分离株)。a.fumigatus菌株在马铃薯葡萄糖琼脂(pda)上维持在4℃,随后在pda斜坡培养基上以37℃培养72h。对于抗真菌剂,伊曲康唑以粉末形式购自美国密苏里州圣路易斯的sigma-aldrich。
30.1.2、生物膜生长
31.生物膜生长按照reichhardt,c等人的描述进行,稍作修改(参考文献:c.reichhardt,j.a.ferreira,l.m.joubert,k.v.clemons,d.a.stevens,l.cegelski,analysis ofthe aspergillus fumigatus biofilm extracellularmatrix by solid-state nuclearmagnetic resonance spectroscopy,eukaryot cell,14(2015)1064-1072.)。现有研究表明,烟曲霉的生物膜活力发生明显变化,当af293在0.25μg/ml浓度的伊曲康唑下生长,而shjt42b在256μg/ml浓度的伊曲康唑下生长(参考文献:m.liu,h.zheng,r.zeng,g.liang,n.zheng,w.liu,effects of itraconazole and micafungin on aspergillus fumigatus biofilms,mycopathologia,(2021))。
32.将在100ml rpmi 1640培养基中含有1
×
105分生孢子/ml的培养物悬浮液接种在每个500ml聚苯乙烯组织培养瓶中;将烧瓶在37℃下静置培养96小时,加入或不加入0.25~256mg/ml的伊曲康唑;收获的生物膜用100ml磷酸盐缓冲盐水(pbs)原位洗涤两次,以去除浮游细胞和松散粘附的细胞;在每个烧瓶中用40ml 10mmtris(ph 7.4)淹没固着细胞,并用涡旋混合器温和混合60秒,添加蛋白酶抑制剂(体积比为1000:1)(solarbio,a8260);将每个样品转移至50ml锥形管中,并在4℃下储存直至处理。
33.1.3、蛋白质提取和消化
34.(1)对于ecm提取,将含有烟曲霉生物膜的管进行超声处理10分钟,然后在涡旋混合器上高速混合30秒。
35.(2)将试管在4~c下以3,600g离心15分钟。将上清液倒入新试管中并再次离心以除去额外的细胞。将上清液转移至新的超滤离心管(merck millipore;amicon ultra-50,ultracel-3k),4℃,5000g 90min,离心浓缩回收至新离心管中,使用bca试剂盒测定蛋白浓度,将上清液在液氮中快速冷冻并冻干。
36.为了消化,蛋白质溶液在56℃下用5mm二硫苏糖醇还原30分钟,然后在室温下在黑暗中用11mm碘乙酰胺烷基化15分钟。然后通过添加100mm teab到尿素浓度小于2m来稀释蛋白质样品。最后在1处加入胰蛋白酶:第一次消化过夜时胰蛋白酶与蛋白质的质量比为50,第二次消化4小时时胰蛋白酶与蛋白质的质量比为1:100。
37.1.4、tmt标记和hplc分级
38.将胰蛋白酶肽用0.5m teab溶解并根据tmt试剂盒操作说明进行标记。标记后,用8%至32%乙腈(ph 9.0)梯度在60分钟内将肽分离成60个级分。
39.将它们合并成18个级分并通过真空离心干燥。使用了十二种培养物(两种菌株的每种处理和未处理样品的三个生物学重复)。
40.1.5、lc-ms/ms分析
41.将多肽溶解在0.1%甲酸(溶剂a)中,直接上样到自制的反相分析柱(15cm长,75μm i.d.)上。梯度包括从6%增加到22%的溶剂b(0.1%甲酸在98%乙腈中)42分钟内,22%到
al.improved atm kinase inhibitor ku-60019radiosensitizes glioma cells,compromises insulin,akt and erk prosurvival signaling,and inhibits migration and invasion.mol cancer ther.2009;8(10):2894-2902.)是一种改进的ku-55933类似物,在无细胞试验中作用于atm,ic50为6.3nm,作用于atm比作用于dna-pk和atr的选择性分别高270和1600倍,并且是高度有效的放射增敏剂。
57.ku-60019是一种改进的ku-55933类似物,分子式:c
30h32
n3o5s,化学名称:(2r,6s)-2,6-二甲基-n-[5-[6-(4-吗啉基)-4-氧代-4h-吡喃-2-基]-9h-噻吨-2-基]-4-吗啉乙酰胺。ku-60019、ku-55933的结构式分别为:
[0058][0059]
将atm抑制剂溶于水,终浓度为3μm;储存条件:-20℃,粉末,3年,或者,-80℃,溶于溶剂,2年。
[0060]
使用方法:使用ku-60019处理细胞1、3和5天。通过alamarblue测定细胞生长:将alamar blue(10%溶液)加到培养基中,达到终浓度1%,实验板在37℃下温育1小时,在fluoro-count酶标仪上(激发光530nm,发射光590nm)测定荧光值,测定结果作为衡量细胞生长的一个指标,通过台酚蓝/荧光激活的细胞分选(facs)实验测定细胞存活。
[0061]
3、tel1与烟曲霉唑类耐药的关系
[0062]
为了验证tel1是否参与烟曲霉唑类耐药,本发明将tel1/atm抑制剂ku-60019与烟曲霉和伊曲康唑共孵育,进行药敏实验。实验过程参照美国临床实验室标准化研究所clsi m-38a2推荐的丝状真菌药敏指南,使用微量液基稀释法并稍作改进,将唑类抗真菌药物进行梯度稀释后与烟曲霉菌悬液加入96孔细胞培养板中,再加入ku-60019,37℃培养46~50h后读数,肉眼观察到无菌生长的最低药物浓度即为mic。如下表2所示:
[0063]
表2有/无tel1抑制剂作用下的烟曲霉伊曲康唑的mic值
[0064][0065]
由表2可知,ku-60019终浓度为5mm时,标准株(af293)伊曲康唑mic值降低到无抑制剂时的1/4,轻度耐药株(shhs18)mic值降低到1/8。ku-60019终浓度为50mm时,高度耐药株(shjt42b)降低为原1/2。该结果说明tel1/atm抑制剂可明显增加烟曲霉对伊曲康唑的敏感性,从侧面印证tel1参与了烟曲霉唑类耐药。
[0066]
在ku-60019作用下,无论烟曲霉标准株af293还是耐药株shjt42b,shjt40或者shhs18对做类药物伊曲康唑itz都有显著增效作用,即相应mic下降。对另外两种唑类药物伏立康唑vrc和泊沙康唑pos,以及费唑类药物两性霉素b(amb),则增效不明显。
[0067]
以上所述仅为本发明的较佳实施例而已,并不用以限制本发明,凡在本发明的精神和原则之内所作的任何修改、等同替换和改进等,均应包含在本发明的保护范围之内。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1