一种与致病力相关的灰霉病菌基因BcSEP6及其应用的制作方法
本发明属微生物基因工程技术领域,具体涉及植物保护领域中控制真菌侵染结构形成与致病力的基因及其编码蛋白质的应用。
背景技术:
灰霉病菌(Botrytis cinerea)通常又称作灰葡萄孢,属于子囊菌门(Ascomycota)真菌,是灰霉病的病原菌,可侵染400多种植物,包括几乎所有的蔬菜及果树作物。寄主从苗期到挂果期均可发病,而且,植株各部位均可被灰霉病菌侵染,叶部发病的典型症状表现为“V”形病斑,花部主要表现为腐烂和调萎,果实主要表现为腐烂和脱落。病害的发生和蔓延与环境的湿度、温度存在密切的关系,在20℃-23℃,相对湿度90%以上时发生严重。因此,灰霉病属低温高湿型病害,在多雨季节或者保护地生产中极易发生,每年因该病导致的经济损失高达100-1000亿美元。由于寄主范围广泛,生产上危害严重,再加上相关分子研究技术成熟,灰霉菌已成为最重要的模式植物病原真菌之一,受到广泛研究。
灰霉病菌是典型的死体营养型病原真菌,可生成多种致病因子参与致病,主要包括细胞壁降解酶类、角质酶、毒素、植物激素、抵抗寄主反应的酶类、小RNA及小分子物质等,这些因子相互协作使灰霉病菌能够杀死寄主细胞并分解死亡的寄主组织作为营养。自然条件下,灰霉菌多以分生孢子作为侵染寄主的初侵染和再侵染来源。灰霉病菌常以菌丝体、分生孢子或菌核附着在植物病残体上或在土壤中越冬和越夏,成为下一生长季的初侵染来源。当条件适宜时,菌核萌发产生菌丝体和分生孢子梗,并产生大量的分生孢子。成熟的分生孢子能够借助风、雨水、灌溉水和农事操作等进行传播。在低温高湿条件下,分生孢子萌发形成芽管,芽管末端稍稍膨大发育成附着胞或者进一步形成侵染垫等侵染结构,主要从衰败的花器、伤口以及坏死组织侵入。
附着胞、侵染垫等侵染结构的发育,对于灰霉病菌侵染寄主是至关重要的,如果侵染结构发育受到影响,灰霉病菌将难以侵入寄主,其危害程度随之会受到严重削弱。灰霉病菌侵染结构的发育受到营养、界面等外源条件及发育信号的联合调控,相关分子机制尚不清楚,对该领域进行深入研究不仅有助于揭示灰霉病菌等死体营养型病原真菌致病的分子机制,而且对于研发防治包括灰霉病菌在内的植物病原真菌的药剂具有重要应用价值。
从分生孢子萌发到发育出功能完备的侵染结构,是一个精细的调控过程,鉴定该调控过程的重要组分,并验证相关基因的致病功能,有可能从中发现可以作为杀真菌剂作用靶标的蛋白质,为开发防治灰霉病及其它类似病害的高效药剂奠定理论和技术基础。
Sep6是一种GTP结合蛋白质,广泛存在于包括真菌在内的真核生物(植物除外),该蛋白质可通过聚合成异源寡聚体蛋白质复合体或进一步形成纤丝,参与调控细胞多种关键的生命过程。通过对灰霉病菌Sep6编码基因的分析,评价该基因在灰霉病菌致病过程的作用,有利于鉴定潜在的防治靶标,用于筛选新型杀真菌药剂。
技术实现要素:
本发明的目的旨在提供一种控制灰霉病菌侵染结构发育和致病性基因及其编码的蛋白质。
本发明所提供的控制侵染结构发育和致病性基因来源于灰霉病菌,名称为BcSEP6,其DNA序列如SEQ ID No:1所示。该DNA序列为BcSEP6基因开放阅读框,由1345个核苷酸组成,其中包含4个外显子,分别位于SEQ ID No:1的第1位至105位核苷酸之间,第169位至331位核苷酸之间,第397位至635位核苷酸之间和第764位至1345位核苷酸之间,组成的编码区长度合计为1089个核苷酸。
本发明提供了来自灰霉病菌的控制侵染结构发育和致病性基因BcSEP6所编码的蛋白质,其氨基酸序列如SEQ ID No:2所示,该序列由362个氨基酸组成。
来自灰霉病菌的控制侵染结构发育和致病性基因BcSEP6可应用于植物抗灰霉病基因工程领域。
来自灰霉病菌的控制侵染结构发育和致病性基因BcSEP6所编码的蛋白质可作为靶标应用于抗灰霉病菌药剂的设计与筛选。
本发明证明了BcSEP6基因的缺失或突变,导致灰霉病菌侵染结构形成率及致病力显著降低,说明BcSEP6基因是灰霉病菌引起农作物灰霉病所必需的基因。因此,筛选能够阻止该基因表达与其蛋白质的表达、修饰及定位的化合物,可以有效控制灰霉病的发生,从而有助于开发新型杀菌剂。即本发明所提供的BcSEP6基因的一个重要用途是:该基因的表达与其编码的蛋白质产物,可以作为重要候选靶标位点,用于抗灰霉病菌药剂的设计与筛选。
本发明所使用的灰霉病菌菌株B05.10,购买自美国真菌遗传材料保藏中心(Fungal Genetics Stock Center,FGSC),其他人员如需要该菌株,可从该保藏中心购买获得,相关信息如下:保藏中心地址:Fungal Genetics Stock Center,Department ofPlant Pathology,Kansas State University,4024Throckmorton Plant Sciences Center,Manhattan,KS 66506USA;网址:http://www.fgsc.net/scripts/StrainSearchReturnPage.asp?OrgID=23812;菌株编号:FGSC 10317。
附图说明
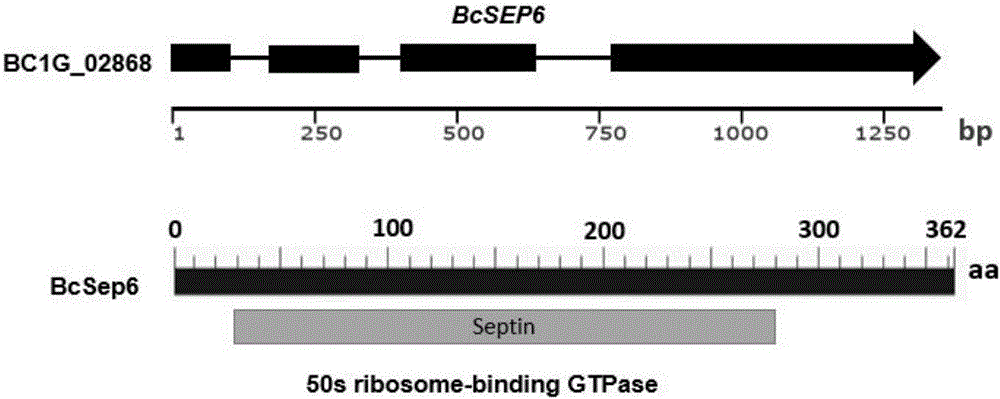
图1为BcSEP6在灰霉病菌基因组中的基因结构图谱
其中BcSEP6基因编号为BC1G_02868,预测该基因具有一个GTPase结构功能域
图2为灰霉病菌BcSEP6基因的敲除策略(通过同源重组进行基因替换)示意图
其中:B05.10为野生型菌株,pSEP6为敲除载体,ΔBcsep6为BcSEP6基因缺失突变体。a、b、c、d、e、f、HpTa、HpTb为验证引物及其方向。
图3为BcSEP6基因缺失突变体的PCR验证电泳图
其中:a、b、c、d、e、f、HpTa、HpTb为验证引物,相应位置见图2,a+b引物验证基因上游是否发生同源交换,c+d引物验证基因下游是否发生同源交换;e+f引物验证目的基因是否被敲除。HpTa+HpTb引物验证抗性筛选基因是否发生插入。ΔBcsep6-1和ΔBcsep6-2为两株独立获得的BcSEP6基因缺失突变体。RT为随机插入转化子
图4为敲除突变体ΔBcsep6与野生型菌株B05.10的培养特征对比照片
其中:所用基本培养基为PDA,B05.10为野生型,ΔBcsep6-1和ΔBcsep6-2为两株独立获得的BcSEP6缺失突变体,RT-1和RT-2为随机插入转化子。
图5为BcSEP6基因的缺失突变体与野生型菌株的产孢量统计分析
其中:B05.10为野生型,ΔBcsep6-1和ΔBcsep6-2为两株独立获得的BcSEP6缺失突变体,RT-1为随机插入转化子,***代表具有显著性差异。
图6为BcSEP6基因的缺失突变体与野生型菌株的分生孢子形态差异比较
其中:B05.10为野生型,ΔBcsep6-1和ΔBcsep6-2为两株独立获得的BcSEP6缺失突变体,RT-1为随机插入转化子,标尺为25μm。
图7为BcSEP6基因的缺失突变体与野生型菌株的侵染垫发育对比分析
其中:B05.10为野生型,ΔBcsep6-1和ΔBcsep6-2为两株独立获得的BcSEP6缺失突变体,RT-1为随机插入转化子,标尺为50μm。
图8为BcSEP6基因的缺失突变体与野生型菌株致病力对比分析
其中:所选择寄主为菜豆叶片,采用离体叶片接种方法,左侧为接种菌丝块,右侧为接种孢子悬浮液,3天后进行评价。B05.10为野生型,ΔBcsep6-1和ΔBcsep6-2和ΔBcsep6-3为三株独立获得的BcSEP6缺失突变体,RT-1,RT-2为随机插入转化子;dpi表示接种后天数。
具体实施方式
为了更好地描述本发明,下面通过具体的实施例予以进一步的说明,下述实施例中的方法,如无特别说明,均为常规方法。
实施例1 BcSEP6基因的相关性分析
灰霉病菌BcSEP6基因的开放阅读框由1345个核苷酸组成,包含4个外显子,编码区cDNA全长为1089个核苷酸,编码的蛋白质产物由362个氨基酸组成。对BcSEP6基因在灰霉病菌基因组网站(https://www.broadinstitute.org/fungal-genome-initiative/botrytis-cinerea-genome-project)中进行预测分析,BcSEP6基因编号为BC1G_02868,该基因具有一个保守GTPase结构功能域。
实施例2 BcSEP6基因的敲除与遗传互补
1)敲除载体的构建
利用Primer 5.0设计引物,由苏州金唯智生物技术有限公司进行引物合成,采用引物Sep6-UP-F(5'-GATCTTCACTAGTGGGAATTCTCGATGTGCGAATGGAAGTG-3')与Sep6-UP-R(5'-TTGGGTACCGAGCTCGAATTCGCTGTCGGTGGGTTTGTGGT-3'),以灰霉病菌菌株B05.10的基因组DNA为模板扩增BcSEP6基因上游600bp片段;采用引物Sep6-DN-F(5'-TGGGGATCCTCTAGAGTCGACCCACGATGTAATTCATGCTCCT-3')与Sep6-DN-R(5'-CTTGCATGCCTGCAGGTCGACATCGGTGAACTACTACGAAACTCC-3')扩增灰霉病菌BcSEP6基因下游809bp片段,反应体系(25μL)为:10mmol/L dNTP Mixture,0.5μL;10×PCR buffer,2.5μL;上下游引物各1μL(10μmol/mL);模板DNA,1μL;Ex-Taq,0.2μL(5U);ddH2O,18.8μL;扩增程序为:94℃预变性3分钟,然后(1)94℃,变性30秒;(2)58℃,退火30秒;(3)72℃,延伸60秒;(4)将1-3步循环30次;(5)72℃延伸10分钟。将上述DNA扩增产物进行纯化回收定量后,利用NEB生物公司的ClonExpress试剂盒,将上游片段和下游片段分别定向克隆至pXEH载体的EcoR I位点与Sal I位点,构建成敲除载体pSEP6(如图2所示),并进行测序验证。
2)灰霉病菌的转化
a.根癌农杆菌的培养
挑取含有双元载体pSEP6的根癌农杆菌菌株Agl-1单菌落,接种至含50μg/ml卡那霉素、10μg/ml利福平的MM液体培养基(磷酸氢二钾0.205%,磷酸二氢钾0.145%,氯化钠0.015%,七水硫酸镁0.05%,六水氯化钙0.01%,七水硫酸亚铁0.00025%,硫酸铵0.05%,葡萄糖0.2%)中,250rpm、28℃振荡培养48h;4000rpm,离心5分钟,弃上清,IM液体培养基(磷酸氢二钾0.205%,磷酸二氢钾0.145%,氯化钠0.015%,七水硫酸镁0.05%,六水氯化钙0.01%,七水硫酸亚铁0.00025%,硫酸铵0.05%,葡萄糖0.2%,200μMAS,MES 0.854%,甘油0.5%)重悬,4000rpm离心5分钟,弃上清;IM培养基重悬,28℃,250rpm振荡培养6h,进行预诱导。
b.灰霉病菌的产孢培养
选用B05.10菌株,取少量孢子涂布于PDA培养基(马铃薯20%煮烂过滤,葡萄糖2%,琼脂1.5%),置28℃培养8h令孢子快速萌发,然后转移至20℃培养3-5天,待菌体表面被灰色孢子覆盖之后,用IM液体培养基刮取、收集孢子,显微镜观察,利用血球计数器调节孢子浓度为1×106/mL。
c.根癌农杆菌与灰霉病菌分生孢子共培养及转化子筛选
将在IM液体培养基中预先诱导6h的农杆菌菌液和灰霉病菌孢子液等体积混合,加入AS,使终浓度达到500μM,混匀,然后按250~350μL/皿,均匀涂抹至铺有玻璃纸的IM培养基上,22℃黑暗培养48h;共培养完毕后,将玻璃纸转移至含有100μg/mL潮霉素的PDA培养基上,相同条件下继续培养。4~7天后挑取扩展的菌落到含有同样抗生素的筛选培养基上。
3)缺失突变体的验证
选用四对引物通过PCR扩增对转化子进行筛选。扩增结果符合如下结果的确定为BcSEP6基因缺失突变体:潮霉素抗性基因内部引物HpTa(5'-TGCGCCCAAGCTGCATCAT-3')和HpTb(5'-TGAACTCACCGCGACGTCTGT-3')可以扩增到800bp片段(野生型菌株无扩增条带);上游同源臂之外基因组上的引物a(5'-TCATTCGTCCAACTCCCACAG-3')与潮霉素抗性基因的引物b(5'-ACAGACGTCGCGGTGAGTTCA-3')配对可以扩增到预期(1.2kb)的重组片段;同时下游同源臂之外基因组上的引物d(5'-CAACAAACTCAGACTCCGCTG-3')与潮霉素抗性基因的引物c(5'-ATGATGCAGCTTGGGCGCA-3')配对可以扩增到预期(1.2kb)的重组片段;而编码区引物e(5'-GAATCTGGACTTGGAAAGAC-3')与f(5'-CTTCCTAGGTCTAGCTTCAC-3')无扩增条带(野生型菌株可扩增到0.94kb片段)。结果,从转化子中筛选到2株独立的BcSEP6基因缺失突变体:ΔBcsep6-1和ΔBcsep6-2(如图3所示),用于后续功能分析。
实施例3 BcSEP6基因在灰霉病菌生长发育过程中的作用
采用平板培养法,评价BcSEP6突变体的生长发育等相关表型的变异情况。将待测菌株打取菌饼分别接种到含有100mg/L潮霉素的PDA培养基及不含有潮霉素的PDA培养基上,20℃黑暗培养3至5天后拍照观察。结果发现,BcSEP6突变体的生长菌落直径与野生型B05.10及随机插入转化子RT-1,RT-2相比严重滞后,而且BcSEP6突变体菌落(菌丝)形态发生明显变化(见图4),这说明BcSEP6基因在灰霉病菌正常的营养生长阶段发挥重要功能,调控灰霉病菌的生长发育。
实施例4 BcSEP6基因在灰霉病菌产孢及分生孢子形态方面的作用
采用平板培养法,评价BcSEP6突变体的产孢能力的变异情况。将待测菌株接种到不含有潮霉素的PDA培养基上,20℃黑暗培养7至10天后,待菌丝长满平板后开始产孢,用一定量的无菌水刮洗平板收集孢子悬浮液,并进行定量分析(见图5),发现BcSEP6突变体的产孢量与野生型相比显著下降;同时取10ul左右孢子悬浮液滴到载玻片上,制成临时玻片,显微观察BcSEP6突变体的分生孢子形态(见图6),发现BcSEP6突变体的分生孢子形态与野生型相比也发生了变异,孢子体形变小,由椭圆形变得更趋圆形;综上所述,BcSEP6基因在灰霉病菌产孢及分生孢子形态建成方面也发挥重要作用。
实施例5 BcSEP6基因在灰霉病菌侵染结构——侵染垫发育过程中的作用
采用玻片培养法,观察BcSEP6突变体发育成侵染垫的能力。将PDB孢子悬浮液(5×104mL-1)滴在载玻片上,室温保湿培养36h。显微观察发现,野生型菌株B05.10及随机插入转化子RT的菌丝体可以形成大量多分枝垫状结构——侵染垫,而突变体则几乎丧失了发育成这种复杂侵染结构的能力;仅仅能形成一些零星的分枝较少的畸形侵染垫(见图7)。该研究结果表明,BcSEP6基因在侵染垫发育过程中发挥举足轻重的作用,该基因是灰霉病菌侵染结构发育所必需的。
实施例6 BcSEP6基因在灰霉病菌致病性方面的作用
采用离体叶片接种法,评价BcSEP6突变体的致病力变化情况。从温室培养的寄主植株上采集具有一定叶龄的叶片,水平放至保湿容器中,使用打孔器打取待测菌株菌饼,正面朝下扣至叶片上,20℃保湿黑暗培养,3天后评价待测菌株的致病力(见图8左)。另外,将待测菌株进行产孢培养后,用PDB缓冲溶液收集分生孢子,并将孢子浓度调至2×106/mL,取5ul孢子悬浮液接种到叶片上,20℃保湿黑暗培养,3天后评价待测菌株的致病力(见图8右)。实验结果表明,不管是菌丝块接种还是孢子悬浮液接种,BcSEP6突变体致病力均显著下降,随机插入转化子致病力与野生型相似,综上研究表明,BcSEP6是一个关键的致病基因,在灰霉病菌侵染寄主过程中发挥重要作用。BcSEP6通过调控灰霉病菌侵染结构的发育,进而影响其致病功能,如果该基因或其编码蛋白质丧失活性,灰霉病菌侵染寄主引发病害的能力将严重下降。
序 列 表
SEQ ID No:1的序列
(i)序列特征:(A)长度:1345 bp;(B)类型:核苷酸;(C)链性:单链
(ii)分子类型:DNA
(iii)序列描述:SEQ ID No:1
1 ATGGCACCAC CCACCGCCGA AAGCGCTTCC CCCATCGGAA TTGCGAATTT
51 GCCCAACCAA AGACACAAGA TTGTCGCAAA GAGAGGTGCT GCCTTCACTA
101 TTATGGTATG TGGTTATCTA TGCTGCTTTC AAATCTCTCA GCAACTTTAT
151 ACTAATCACC TCTTGTAGGT CGCTGGCGAA TCTGGACTTG GAAAGACAAC
201 CTTCATTAAT ACCCTCTTCT CTACCACAAT CAAAAACTAT GCCGATCACA
251 AACGCAGACA CGCAAAGCAA GTGGATAAAA CTGTTGAGAT TGAAATCACA
301 AAGGCTGAAT TAGAAGAGAA ATTCTTCAAG GGTATGGAAT CCAACACATT
351 CAGTTCACTT GAGATAATTA ATTGGCCATT GTTGACACTG TCTTAGTTCG
401 TTTGACTGTT ATCGATACAC CTGGTTTTGG AGATTATGTC AACAATCGCG
451 ACTCCTGGAT GCCAATCATC GAGTTTTTGG ATGATCAACA TGAGTCCTAT
501 ATGTTGCAAG AGCAACAGCC TCGCCGTGTT GATAAAATCG ATTTACGTGT
551 ACACGCTTGC TTGTACTTCA TCCGACCTAC TGGCCACACA CTGAAACCCC
601 TCGATATTGA GGTTATGAAG AGACTCAGTT CAAGGGTAAA CTTGATTCCA
651 GTAGTTGCTA AGGCTGATAC CTTAAGCCCG GCTGATTTGG CCCGTTATAA
701 GCAAAAGGTA TAGTGTATTT GAGTAAACCA TTATCGTTGT CGAAGTTAAC
751 GTATTTTCTA CAGATCCAAG CTGTCATTGA AGCACAAGGT ATCAAGATCT
801 ACACACCACC TATCGAGGAA GATGACGAGG CTGCTGCTCA ACATGCCCGA
851 AGTTTGATGG CTGCTATGCC ATTTGCAGTC ATTGGCTCAG AGAAGGATGT
901 CAAGACTAGT GATGGACGTA TCGTTAAGGG TCGTCAATAC GCATGGGGTG
951 TTGCTGAAGT CGAGAACGAA GACCACTGCG ATTTCAAGAA GTTGAGATCC
1001 ATTCTTATCC GAACACACAT GCTCGATCTC ATTCACACTA CCGAAGAAGC
1051 ACACTACGAG GCTTACCGCG CCCAACAAAT GGAGACTCGC AAATTCGGTG
1101 AAGCTAGACC TAGGAAGCTT GATAACCCTA AATTCAAGGA AGAGGAAGAG
1151 AGCTTAAGAA AGCGTTTCAC CGAGCAGGTT AAGATTGAGG AGCACCGTTT
1201 CAGACAGTGG GAGCAGAAGC TCATCAGTGA GCGTGATCGT TTGAACAAGG
1251 ATTTGGAGAG TACACATGCT GCCATCAAGT CCTTAGAACA AGAACTTGAG
1301 CAAATGCAAG GCGGTGCCGC AACTCGCAGT CACGGCCGTC GTTAA
SEQ ID No:2的序列
(i)序列特征:(A)长度:362个氨基酸;(B)类型:氨基酸;(C)链性:单链。
(ii)分子类型:多肽
(iii)序列描述:SEQ ID No:2
1 MAPPTAESAS PIGIANLPNQ RHKIVAKRGA AFTIMVAGES GLGKTTFINT
51 LFSTTIKNYA DHKRRHAKQV DKTVEIEITK AELEEKFFKV RLTVIDTPGF
101 GDYVNNRDSW MPIIEFLDDQ HESYMLQEQQ PRRVDKIDLR VHACLYFIRP
151 TGHTLKPLDI EVMKRLSSRI QAVIEAQGIK IYTPPIEEDD EAAAQHARSL
201 MAAMPFAVIG SEKDVKTSDG RIVKGRQYAW GVAEVENEDH CDFKKLRSIL
251 IRTHMLDLIH TTEEAHYEAY RAQQMETRKF GEARPRKLDN PKFKEEEESL
301 RKRFTEQVKI EEHRFRQWEQ KLISERDRLN KDLESTHAAI KSLEQELEQM
351 QGGAATRSHG RR
- 还没有人留言评论。精彩留言会获得点赞!