大豆转录因子GmMYB521及其编码基因在调控植物油脂代谢中的应用的制作方法
本发明属于生物
技术领域:
,具体涉及大豆转录因子gmmyb521及其编码基因在调控油脂代谢中的应用。
背景技术:
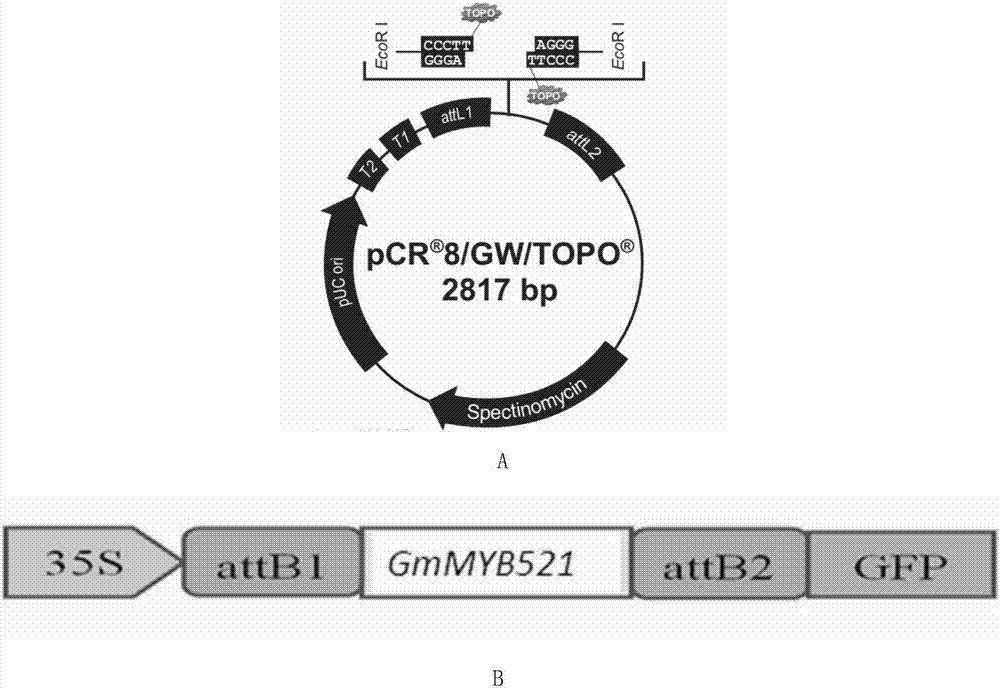
:人类饮食中71%的油脂来自于植物。在世界上的几种主要产油作物中,大豆总产油量约占30%,居世界植物油产量的第一位,棕榈油及油菜子油分别位居第二及第三(如表1所示)。表1、世界上主要的产油作物种类生产量(百万吨)占总产量百分比相对顺序大豆(soybean)15.5029.11棕榈(palm)8.5216.02油菜籽(rapeseed)7.0313.23向日葵(sunflower)7.0013.14棉花籽(cottonseed)3.316.25椰子(coconut)2.715.16花生(peanut)2.695.07橄榄(olive)1.633.18脂肪酸的合成是植物体中最重要的代谢途径之一,它存在于植物体的任何一个细胞中,是生长发育所必须的。它的阻断会导致细胞的死亡,因而至今为止还没有发现一个阻断脂肪酸合成的植物突变体。植物中脂肪酸的合成主要是在质体中进行,而动物及真菌的脂肪酸合成发生于胞质。因此植物需要存在一种不同于动物及真菌的机制——从质体运出脂肪酸到细胞的其它部位。因而在细胞中必定存在脂肪酸生产和运输的控制机制,但是迄今尚不清楚在脂肪酸的合成中质体内外是如何联系的。植物同其它真核生物在参与脂肪酸合成途径的酶上有很大差异。从乙酰coa和丙二酰coa合成16或18个碳原子的脂肪酸至少需要30个不同的酶催化的反应来完成这一过程,而在动物、真菌及一些细菌中,以上反应是由一个存在于胞质中的多酶复合体来完成的。植物中,参加脂肪酸合成的酶分别以可溶的形式独立存在于质体的胞质中。尽管植物中参与脂肪代谢的酶很容易被分离,但问题是这些酶是否在体内也可形成一个多酶复合体。脂肪酸合成途径中最重要的碳源是由accase合成的丙二酰coa,在进入脂肪酸合成途径之前,丙二酰由coa转移到酰基载体蛋白(acp)上,从此脂肪酸合成都需要acp的参与,直到形成16或18个碳原子的脂肪酸,并被用于合成甘油或被运出质体。acp是一个分子量为9kd的酸性蛋白,它具有一个可以通过硫酯化结合乙酰基的基团。当丙二酰由coa转移到acp后,硫酯化的丙二酰由coa进行一系列的聚合反应,接受乙酰acp或乙酰coa的乙酰基团。这一聚合反应是通过释放一个co2分子来形成一个c-c键,co2的释放使这一反应变得不可逆,从而使聚合反应不断进行。大多数植物中,油脂都以三酰甘油(triacylglycerols,tag)的形式储藏,它的含量是一个非常重要的农艺性状,tag的生物合成称之为kennedy途径,如同真核生物中合成膜甘油酯的途径,脂肪酸去除coa后被转移到3-磷酸甘油的1和2位,形成中间产物pa。pa去磷酸化产生dag。在tag合成的最后一步,第三个脂肪酸分子被转移到空的dag3’-oh位置,这一步反应是由二酰甘油乙酰转移酶(diacylglycerolacyltransferase,dgat)催化的,此反应被认为是tag生物合成中唯一的限速步骤。人们已对脂类合成途径有了认知,并且已经克隆参与脂类合成的酶基因。技术实现要素:本发明的一个目的是提供gmmyb521蛋白的新用途。所述gmmyb521蛋白是如下a)或b)或c)或d)的蛋白质:a)氨基酸序列是序列2所示的蛋白质;b)在序列2所示的蛋白质的n端和/或c端连接标签得到的融合蛋白质;c)将序列2所示的氨基酸序列经过一个或几个氨基酸残基的取代和/或缺失和/或添加得到的具有相同功能的蛋白质;d)与序列2所示的氨基酸序列具有70%、具有75%、具有80%、具有85%、具有90%、具有95%、具有96%、具有97%、具有98%或具有99%的同源性且具有相同功能的蛋白质。本发明提供了gmmyb521蛋白在调控植物油脂含量和/或油脂代谢中的应用。本发明还提供了与gmmyb521蛋白相关的生物材料的新用途。本发明提供了与gmmyb521蛋白相关的生物材料在调控植物油脂含量和/或油脂代谢中的应用;所述与gmmyb521蛋白相关的生物材料为下述a1)至a12)中的任一种:a1)编码gmmyb521蛋白的核酸分子;a2)含有a1)所述核酸分子的表达盒;a3)含有a1)所述核酸分子的重组载体;a4)含有a2)所述表达盒的重组载体;a5)含有a1)所述核酸分子的重组微生物;a6)含有a2)所述表达盒的重组微生物;a7)含有a3)所述重组载体的重组微生物;a8)含有a4)所述重组载体的重组微生物;a9)含有a1)所述核酸分子的转基因植物细胞系;a10)含有a2)所述表达盒的转基因植物细胞系;a11)含有a3)所述重组载体的转基因植物细胞系;a12)含有a4)所述重组载体的转基因植物细胞系。上述应用中,a1)所述核酸分子为如下1)或2)或3)所示的基因:1)其编码序列是序列1所示的cdna分子或dna分子;2)与1)限定的核苷酸序列至少具有70%、至少具有75%、至少具有80%、至少具有85%、至少具有90%、至少具有95%、至少具有96%、至少具有97%、至少具有98%或至少具有99%同源性且编码gmmyb521蛋白的cdna分子或基因组dna分子;3)在严格条件下与1)或2)限定的核苷酸序列杂交,且编码gmmyb521蛋白的cdna分子或基因组dna分子。所述严格条件可为如下:50℃,在7%十二烷基硫酸钠(sds)、0.5mnapo4和1mmedta的混合溶液中杂交,在50℃,2×ssc,0.1%sds中漂洗;还可为:50℃,在7%sds、0.5mnapo4和1mmedta的混合溶液中杂交,在50℃,1×ssc,0.1%sds中漂洗;还可为:50℃,在7%sds、0.5mnapo4和1mmedta的混合溶液中杂交,在50℃,0.5×ssc,0.1%sds中漂洗;还可为:50℃,在7%sds、0.5mnapo4和1mmedta的混合溶液中杂交,在50℃,0.1×ssc,0.1%sds中漂洗;还可为:50℃,在7%sds、0.5mnapo4和1mmedta的混合溶液中杂交,在65℃,0.1×ssc,0.1%sds中漂洗;也可为:在6×ssc,0.5%sds的溶液中,在65℃下杂交,然后用2×ssc,0.1%sds和1×ssc,0.1%sds各洗膜一次。上述应用中,所述重组载体是将上述基因插入表达载体中,得到表达上述蛋白的重组载体。用于构建所述植物表达载体的出发载体可为任意一种植物表达载体,例如gateway系统载体或双元农杆菌载体等,如pgwb411、pgwb412、pbin438、pcambia1302、pcambia2301、pcambia1301、pcambia1300、pbi121、pcambia1391-xa或pcambia1391-xb(cambia公司)。构建植物表达载体时,在其转录起始核苷酸前可加上任何一种增强型、组成型、组织特异型或诱导型启动子,如花椰菜花叶病毒(camv)35s启动子、泛生素基因ubiquitin启动子(pubi)等,它们可单独使用或与其它的植物启动子结合使用;此外,使用本发明的基因构建植物表达载体时,还可使用增强子,包括翻译增强子或转录增强子,这些增强子区域可以是atg起始密码子或邻接区域起始密码子等,但必需与编码序列的阅读框相同,以保证整个序列的正确翻译。所述翻译控制信号和起始密码子的来源是广泛的,可以是天然的,也可以是合成的。翻译起始区域可以来自转录起始区域或结构基因。为了便于对转基因植物细胞或植物进行鉴定及筛选,可对所用植物表达载体进行加工,如加入可在植物中表达的编码可产生颜色变化的酶或发光化合物的基因(gus基因、萤光素酶基因等)、具有抗性的抗生素标记物(庆大霉素标记物、卡那霉素标记物等)或是抗化学试剂标记基因(如抗除莠剂基因)等。从转基因植物的安全性考虑,可不加任何选择性标记基因,直接以逆境筛选转化植株。携带有本发明gmmyb521的植物表达载体可通过使用ti质粒、ri质粒、植物病毒载体、直接dna转化、微注射、电导、农杆菌介导等常规生物学方法转化植物细胞或组织,并将转化的植物细胞或组织培育成植株。被转化的植物宿主既可以是双子叶植物,如油菜、大豆、苜蓿、向日葵、拟南芥或棉花等,也可以是单子叶植物,如水稻、小麦、玉米等。上述应用中,所述调控为提高,体现在提高植物组织油脂含量,具体体现在提高植物种子中的油脂含量。本发明还提供了gmmyb521蛋白或与gmmyb521蛋白相关的生物材料的新用途。本发明提供了gmmyb521蛋白或与gmmyb521蛋白相关的生物材料在培育高油脂的转基因植物中的应用。本发明还提供了gmmyb521蛋白或与gmmyb521蛋白相关的生物材料在植物育种中的应用。本发明还有一个目的是提供一种培育高油脂的转基因植物的方法。本发明提供的培育高油脂的转基因植物的方法包括提高受体植物中gmmyb521蛋白的含量和/或活性,得到转基因植物的步骤;所述转基因植物的油脂含量高于受体植物。上述方法中,所述提高受体植物中gmmyb521蛋白的表达量和/或活性的方法为在受体植物中过表达gmmyb521蛋白;所述过表达的方法为将gmmyb521蛋白的编码基因导入受体植物。在本发明的具体实施例中,所述gmmyb521蛋白的编码基因是通过pgwb411-gmmyb521重组载体导入受体植物。所述pgwb411-gmmyb521重组载体为将gmmyb521蛋白的编码基因同源重组到pgwb411载体的重组位点attl1和attl2之间,且保持pgwb411载体的其他序列不变后得到的载体。上述方法中,所述gmmyb521蛋白的编码基因的核苷酸序列是序列1所示的dna分子。上述应用或上述方法中,所述油脂含量为植物组织油脂含量;所述植物组织具体为植物种子。上述方法中,所述植物为单子叶植物或双子叶植物,具体为大豆、百脉根、苜蓿、水黄皮、油菜、向日葵、拟南芥或玉米。在本发明的具体实施例中采用双子叶植物拟南芥,具体为哥伦比亚生态型拟南芥(col-0)。本发明提供了一种与植物组织油脂含量相关的大豆转录因子gmmyb521及其编码基因gmmyb521,并将该编码基因gmmyb521转入拟南芥中,得到转gmmyb521拟南芥,该转gmmyb521拟南芥与野生型拟南芥相比,种子中的油脂含量显著提高。说明大豆转录因子gmmyb521及其编码基因可以调控植物种子中油脂含量。本发明的大豆转录因子gmmyb521及其编码基因gmmyb521对提高和改良作物油脂成份,特别是对于提高大豆等油料植物籽粒中油脂成份,培育高油脂品种具有重要的理论和现实意义。附图说明图1为克隆载体和植物表达载体pgwb411-gmmyb521示意图。图2为转gmmyb521拟南芥纯合株系的分子鉴定。图3为转gmmyb521拟南芥纯合株系和对照种子的总油脂含量比较。具体实施方式下述实施例中所使用的实验方法如无特殊说明,均为常规方法。下述实施例中所用的材料、试剂等,如无特殊说明,均可从商业途径得到。下述实施例中的定量试验,均设置三次重复实验,结果取平均值。下述实施例中的大豆材料:大豆黑农44(hn44)记载在如下文献中:满为群等,大豆新品种黑农44的选育及不同种植方式对其产量和品种的影响,黑龙江农业科学2004年5期,1-5。公众可从中国科学院遗传与发育生物学研究所处获得,该生物材料只为重复本发明的相关实验所用,不可作为其它用途使用。下述实施例中的表达载体pgwb411记载在如下文献中:departmentofmolecularandfunctionalgenomics,shimaneuniversity,aatsue,shimane690-8504,japan,e.mail:tnakagaw@life.shimane-u.ac.jpisuyoshinakagawa,etal.,gatwayvectorsforplanttransformation,plantbiotechnology,2009,26,275-284。由tsuyoshinakagawa博士提供,公众得到tsuyoshinakagawa博士同意后可从中科院遗传与发育生物学研究所获得。下述实施例中的农杆菌gv3101记载在如下文献中:leecw等,agrobacteriumtumefacienspromotestumorinductionbymodulatingpathogendefenseinarabidopsisthaliana,plantcell,2009,21(9),2948-62。公众可从中国科学院遗传与发育生物学研究所处获得,该生物材料只为重复本发明的相关实验所用,不可作为其它用途使用。下述实施例中的哥伦比亚生态型拟南芥(col-0)的种子购自arabidopsisbiologicalresourcecenter(abrc)。实施例1、与油脂代谢调控相关的大豆转录因子gmmyb521的筛选与克隆一、与油脂代谢调控相关的大豆转录因子gmmyb521的筛选以两个油脂含量有差异的大豆品种黑农44(hn44)和zyd7构建重组自交系,定位种子油脂含量相关的qtl,其中,hn44籽粒中油脂含量为23%,而zyd7籽粒的油脂含量为12%。在所定位的qtl区间,检测到williams82参考基因组新版本中编号为glyma.20g047600的基因与种子油脂含量相关,鉴于数据库中没有命名该基因,暂时将其命名为gmmyb521。二、与油脂代谢调控相关的大豆转录因子gmmyb521的克隆1、cdna的制备提取黑农44(hn44)幼苗的总rna,用逆转录酶将rna反转录合成cdna。2、引物的设计根据在plantgdb的大豆基因组序列中gmmyb521(glyma.20g047600)全长cdna序列的信息设计引物,引物序列如下:gmmyb521-f:ggggacaagtttgtacaaaaaagcaggcttcatgagtagtgtgactagtgggggt;gmmyb521-r:ggggaccactttgtacaagaaagctgggttgcgctgacaatcttcaccaagaag。3、pcr扩增以步骤1获得的cdna为模板,以gmmyb521-f和gmmyb521-r为引物进行pcr扩增,得到约1.5kb的pcr产物并对其进行测序。经过测序,该pcr产物大小为1566bp,具有序列表中序列1所示的核苷酸,该核苷酸所示的基因为gmmyb521,该基因编码的蛋白命名为gmmyb521,该蛋白包含321个氨基酸,其氨基酸序列为序列表中的序列2。实施例2、转gmmyb521拟南芥的获得及其功能分析一、转gmmyb521拟南芥的获得及鉴定1、植物表达载体构建(1)使用invitrogen公司提供的gateway系统将实施例1中得到的大小为1566bp的pcr产物(gmmyb521)运用ta克隆的原理克隆与载体/gw/topo(/gw/tacloningkit,catalognumber:k2500-20,invitogencorporation,carlsbad,ca,usa)连接,得到中间载体/gw/-gmmyb521,其示意图如图1a所示。(2)将中间载体/gw/-gmmyb521和过表达载体pgwb411在重组酶的作用下进行lr重组反应,由于/gw/topo载体和过表达载体pgwb411上均带有重组位点attl1和attl2,因此连接目的基因的中间载体/gw/-gmmyb521可以与过表达载体pgwb411发生同源重组,最终使目的基因gmmyb521成功构建到过表达载体pgwb411上,得到重组载体pgwb411-gmmyb521。重组反应体系如下:1ul/gw/-gmmyb521,1ulpgwb411,1ullrbuffer,1ullrenzymemix,1ultebufferph8.0。25℃反应6h,加0.5ul蛋白酶k后,37℃反应10min,得到重组载体pgwb411-gmmyb521。重组载体pgwb411-gmmyb521经过测序表明:该载体为将序列表中序列1所示的dna分子同源重组到pgwb411载体的重组位点attl1和attl2之间,且保持pgwb411载体的其他序列不变后得到的载体,其部分结构示意图如图1b所示。2、重组农杆菌的获得将含有gmmyb521的重组载体pgwb411-gmmyb521用电击法导入农杆菌gv3101,得到重组农杆菌,并将其命名为gv3101/gmmyb521。3、转gmmyb521拟南芥的获得将重组农杆菌gv3101/gmmyb321培养至对数期,然后用抽真空法将其转化哥伦比亚生态型拟南芥(col-0)中。经培育后收获种子,将种子播于含卡那霉素(50mg/l)的ms筛选培养基上,收集t1代转化植株的新鲜叶片,提取rna,反转录得到cdna,并以cdna作为模板,采用gmmyb521-f和gmmyb521-f引物进行realtime-pcr鉴定t1代阳性植株。以野生型拟南芥(col-0)为对照,以拟南芥atactin2基因为内标,所用引物为primer-tf:5’-atgcccagaagtcttgttcc-3’和primer-tr:5’-tgctcatacggtcagcgata-3’。取20棵t1代阳性植株,待长至4-6叶时移到蛭石上生长,收获t1代单株,各单株种子分别播种,用相同的ms筛选培养基继续筛选以观察t2代的分离情况,如此重复数代直至获得遗传稳定的t3代转gmmyb521纯合株系。随机选取10个t3代转gmmyb521拟南芥纯系。提取苗期rna,再次用上述realtime-pcr方法鉴定纯系植株中gmmyb521的表达。结果如图2所示。从图中可以看出:t3代转gmmyb521纯合株系oe-8、oe-23、oe-26、oe-21、oe-27、oe-19、oe-18、oe-28、oe-9和oe-2中均能检测到gmmyb321的表达,其表达量分别约为0.024、0.017、0.016、0.014、0.013、0.012、0.011、0.010、0.0095和0.0090。而野生型拟南芥(col-0)中未能检测出gmmyb521的表达。说明本发明成功将gmmyb521基因转入拟南芥中,且得到表达。选取t3代转gmmyb521纯合株系oe-8、oe-23、oe-26、oe-21、oe-27、oe-19、oe-18、oe-28、oe-9和oe-2用于下述实验。二、转gmmyb521基因拟南芥种子中的总油脂含量分析分别测定野生型拟南芥(col-0)、t3代转gmmyb521纯合株系oe-8、oe-23、oe-26、oe-21、oe-27、oe-19、oe-18、oe-28、oe-9和oe-2种子中的总油脂含量。种子总油脂含量测定方法如下:将干燥的种子研磨成粉,称取100mg到离心管中,平行称取四份。加入500μl的正己烷,充分混匀,37℃过夜。慢速离心3分钟,将正己烷吸入称量过新管中。剩下的粉末继续加正己烷重复浸泡、然后离心、然后收集正己烷到同一的离心管中。将离心管放入真空泵中,抽真空,使正己烷完全挥发。然后再次称取离心管的重量。离心管前后重量的变化即是提取的脂类重量;总油脂量(%)的计算公式如下:总油脂量(%)=提取的脂类重量/种子总重量×100%。每个株系取30株的种子,实验重复三次,结果取平均值±标准差。结果如图3所示。从图中可以看出:野生型拟南芥种子的总油脂量约为37.3%(即为种子总重量的百分比);t3代转gmmyb521纯合株系oe-8、oe-23、oe-26、oe-21、oe-27、oe-19、oe-18、oe-28、oe-9和oe-2种子的总油脂量分别约为40.5%、43.0%、41.5%、41.7%、44.0%、41.8%、42.5%、42.5%、39.1%和38.7%。结果表明,10个t3代转gmmyb521纯合株系种子中的总油脂含量均极显著或显著高于对照。上述实验表明,大豆myb类转录因子gmmyb521对种子中总油脂的合成呈正调控作用,其编码基因gmmyb321的过量表达,可提高转基因植株种子中的总油脂含量。序列表<110>中国科学院遗传与发育生物学研究所<120>大豆转录因子gmmyb521及其编码基因在调控油脂代谢中的应用<160>2<210>1<211>1566bp<212>prt<213>人工序列<220><223><400>1atgagtagtgtgactagtgggggtgataatcgaaaaatatccaaaggtcgtcggtcatca60tcattagaggaagaagctggagttggaggtaacataagaggagaaagtcctctgaagaaa120ggcccatggactgcagcagaggatgcaattttggtagaatatgccaaaaaacacggacaa180ggcaattggaatgcagtacataaatattcaggacttgcccgttgtgggaaaagctgccgt240ctacgatgggcaaatcatttgagaccagatctaaaaaaaggtgaatttactgcagaagaa300gagaatcgaatccttgaactacatgctaagatgggaaacaaatgggctcgaatggctgca360gagttgcctggacgtacggataatgagatcaagaactactggaacacaagaatcaaaaga420atgcaacgagcaggcttaccaatctaccctgaagaattgtgccaacggatacttaattgc480aatcaagaaagtcaaaatattagcatcctgtcaaatgaagcctcccagcatggtgatctg540tcacaaacagactttgatataccagacgtggaattcaaaattttcaaatttcgtcatggt600ctatcacatggacagtcaatatttgatatgcctgaaagtagcttgtttgatcaaagttca660gactcatcccatagttataacttgttcccaactatgcgtcctacaaaacgccctcgggag720tcagaaatgctatatgatagttttgaaagttgtacaattaatgctgccccactatttgat780cagtatgacaattatacttctgaaaaaatttctgaccatcctaggttgtctcttccacgt840gatcccgttcttaacaccaatgatcagtttaatggtgataaccttaccggcagccatgcc900gcattaaatggcaatgcctcttcttctgtgcccatgtttagggccatgaagttggagctc960ccttcactccaatatcctgaaactcaacatggtagctggggcacgcctacgtccccgctt1020ccttcactcgagtctgttgatacattgattcagtctcctgttgttgagcctattctatta1080gatcccatttctccgcagagcagtggtttgctggaagcgatagtccacaactcgaaaagc1140ttgaaaggctcaaacaatgatttattattacaagaaacaattggtacaaccaatgaagta1200gctaagagttcaaccttgaatcattcctttcagacaaaatggtatgaactaggggagcca1260aattccccctttggtcaatctgctgcttcagttctaattgagtatactcctgtaagcatg1320tgctcagtggatggcccccaatcaattgaaaccactcaggatcatgatgataagcatgaa1380gcactcaccacccaatttccagacagttccaggaagaaaaaaaatatattgaaacagatg1440gattatacacagccagatgctttacttgacttaggttggtttgggaatagcaccgagtat1500ggaagtgatcaatctgttctgcaagatgctttaagtgcacttcttggtgaagattgtcag1560cgctag1566<210>2<211>521<212>prt<213>人工序列<220><223><400>2metserservalthrserglyglyaspasnarglysileserlysgly151015argargserserserleuglugluglualaglyvalglyglyasnile202530argglygluserproleulyslysglyprotrpthralaalagluasp354045alaileleuvalglutyralalyslyshisglyglnglyasntrpasn505560alavalhislystyrserglyleualaargcysglylyssercysarg65707580leuargtrpalaasnhisleuargproaspleulyslysglygluphe859095thralagluglugluasnargileleugluleuhisalalysmetgly100105110asnlystrpalaargmetalaalagluleuproglyargthraspasn115120125gluilelysasntyrtrpasnthrargilelysargmetglnargala130135140glyleuproiletyrproglugluleucysglnargileleuasncys145150155160asnglngluserglnasnileserileleuserasnglualasergln165170175hisglyaspleuserglnthrasppheaspileproaspvalgluphe180185190lysilephelysphearghisglyleuserhisglyglnserilephe195200205aspmetprogluserserleupheaspglnserseraspserserhis210215220sertyrasnleupheprothrmetargprothrlysargproargglu225230235240serglumetleutyraspserpheglusercysthrileasnalaala245250255proleupheaspglntyraspasntyrthrserglulysileserasp260265270hisproargleuserleuproargaspprovalleuasnthrasnasp275280285glnpheasnglyaspasnleuthrglyserhisalaalaleuasngly290295300asnalaserserservalprometpheargalametlysleugluleu305310315320proserleuglntyrprogluthrglnhisglysertrpglythrpro325330335thrserproleuproserleugluservalaspthrleuileglnser340345350provalvalgluproileleuleuaspproileserproglnserser355360365glyleuleuglualailevalhisasnserlysserleulysglyser370375380asnasnaspleuleuleuglngluthrileglythrthrasngluval385390395400alalysserserthrleuasnhisserpheglnthrlystrptyrglu405410415leuglygluproasnserpropheglyglnseralaalaservalleu420425430ileglutyrthrprovalsermetcysservalaspglyproglnser435440445ilegluthrthrglnasphisaspasplyshisglualaleuthrthr450455460glnpheproaspserserarglyslyslysasnileleulysglnmet465470475480asptyrthrglnproaspalaleuleuaspleuglytrppheglyasn485490495serthrglutyrglyseraspglnservalleuglnaspalaleuser500505510alaleuleuglygluaspcysglnarg515520当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1