能够特异结合HPVl6-E6蛋白的纳米抗体及其编码序列、制备方法和应用与流程
本发明属于免疫学技术领域,涉及一种新的hpv16-e6抗体或其功能性片段,具体为能够特异结合hpvl6-e6蛋白的纳米抗体及其编码序列、制备方法和应用。
背景技术:
流行病学调查证实了hpv感染与宫颈癌的发生有极密切的相关性。hpv16是最常见的与宫颈癌相关的高危型病毒。e6基因属于hpv早期基因,e6蛋白属于小分子碱性蛋白,含锌指结构。e6蛋白可通过锌指结构与宿主细胞的多种蛋白相结合,使细胞发生恶性转化,在hpv的致癌过程中发挥着重要的作用。
1993年比利时的hamerscasterman报道了在骆驼血清中存在缺失轻链的重链抗体,这类抗体的抗原结合位点仅由重链的可变区形成,尽管天然缺失轻链可变区,却仍具有良好和广泛的抗原结合力。而且单独克隆并表达出来的重链可变区具有与原重链抗体相当的结构稳定性以及与抗原的结合活性,是目前已知的可结合目标抗原的最小单位,被称为纳米抗体。纳米抗体具有易表达、水溶性好、稳定性强等优点,使得纳米抗体具有广阔的应用前景。
目前hpvl6感染相关的临床诊断主要依靠以pcr技术为主的分子生物学方法,该方法具有设备条件要求高,检测结果假阳性率高等缺点。
技术实现要素:
解决的技术问题:为了克服现有技术的不足,获得一种检测敏感性高,易于实际应用的纳米抗体,本发明提供了能够特异结合hpvl6-e6蛋白的纳米抗体及其编码序列、制备方法和应用。
技术方案:能够特异结合hpvl6-e6蛋白的纳米抗体,所述抗体vhh链包括框架区和互补决定区;所述框架区包括以下4段氨基酸序列:fr1、seqidno:1,fr2、seqidno:3,fr3、seqidno:5,fr4、seqidno:7;所述互补决定区包括以下3段氨基酸序列:cdr1、seqidno:2,cdr2、seqidno:4,cdr3、seqidno:6。
能够特异结合hpvl6-e6蛋白的纳米抗体,hpvl6-e6蛋白纳米抗体的氨基酸序列为seqidno:8。
编码能够特异结合hpvl6-e6蛋白的纳米抗体的基因,其核苷酸序列为seqidno:9。
重组载体,所述重组载体中包含seqidno:9序列。
重组细胞,所述重组细胞中包含seqidno:9序列。
以上任一所述能够特异结合hpvl6-e6蛋白的纳米抗体的制备方法,包括以下步骤:
(1)构建抗hpvl6-e6蛋白的纳米抗体噬菌体文库;
(2)选择与hpvl6-e6具有特异结合的纳米抗体噬菌体;
(3)培养包含编码抗hpvl6-e6蛋白的纳米抗体核酸的载体的宿主;
(4)从宿主细胞中纯化获得抗hpvl6-e6蛋白的纳米抗体。
优选的,采用巢式pcr方法构建hpvl6-e6蛋白纳米抗体的文库,其中第一轮扩增反应引物序列为seqidno:10-11,第二轮扩增反应引物序列为seqidno:12-13。
优选的,步骤(2)中采用ni-ida亲和层析的方法筛选抗hpvl6-e6蛋白纳米抗体。
所述能够特异结合hpvl6-e6蛋白的纳米抗体在制备检测hpvl6-e6蛋白的试剂盒中的应用。
所述能够特异结合hpvl6-e6蛋白的纳米抗体在制备治疗或预防宫颈癌的药物中的应用。
本发明所述能够特异结合hpvl6-e6蛋白的纳米抗体的作用原理在于:hpv病毒e6的致癌作用主要是基于e6和其他蛋白的相互作用,这种蛋白之间相互作用对于小分子抑制剂很难达到破坏效果。而基于抗体与抗原的特异结合特点,而且纳米抗体可以靶向胞内蛋白,因此可以作为阻碍胞内靶蛋白和其他蛋白相互作用的抑制剂,抑制靶蛋白与其他蛋白的相互作用,具有其他传统抗体无可比拟的治疗优势。
有益效果:(1)本发明所述能够特异结合hpvl6-e6蛋白的纳米抗体具有高度特异性,检测敏感性高;(2)所述抗体能够在大肠杆菌中高效表达,因而为后续的免疫试剂及肿瘤药物组合物的量产化生产提供基础。
附图说明
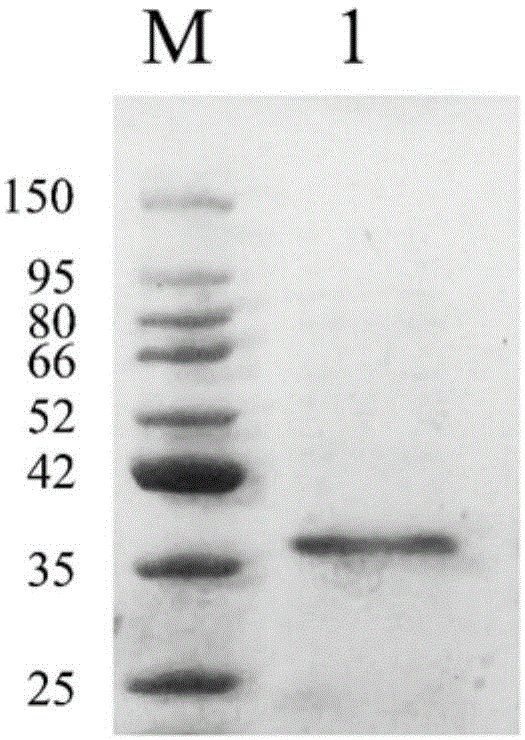
图1为psumo-hpvl6e6蛋白镍柱亲和层析纯化后的蛋白sds-page电泳图,其中m为蛋白分子量标准,1为目标蛋白;
图2为nest-pcr11.5%琼脂糖凝胶电泳图,其中m为marker,1、2为nest-pcr1产物;
图3为nest-pcr21.5%琼脂糖凝胶电泳图,其中m为marker,1为nest-pcr2扩增所得的vhh片段;
图4为抗hpvl6e6纳米抗体文库重组阳性率检测图,其中m为marker,1-9是pcr产物;
图5为抗hpvl6e6纳米抗体sds-page电泳图,其中,m为蛋白分子量标准,1为纯化的抗体蛋白;
图6a为抗hpvl6e6纳米抗体免疫印迹亲和性分析图,psumo空质粒表达蛋白sumo作为对照;
图6b为抗hpv16e6纳米抗体特异识别真核细胞内的hpv16e6的western图。
具体实施方式
以下实施例进一步说明本发明的内容,但不应理解为对本发明的限制。在不背离本发明精神和实质的情况下,对本发明方法、步骤或条件所作的修改和替换,均属于本发明的范围。若未特别指明,实施例中所用的技术手段为本领域技术人员所熟知的常规手段。
实施例1
1hpvl6e6蛋白的表达及纯化
(1)hpvl6e6蛋白的表达
将构建好的psumo-hpv16e6载体转化到bl21(de3)中,挑单克隆培养过夜;然后扩大培养,iptg诱导表达。
(2)hpvl6e6蛋白的纯化
通过ni-ida亲和层析的方法获得纯化的hpvl6e6蛋白。图1中sds-page电泳检测结果显示,经镍柱纯化的hpvl6e6蛋白,蛋白纯度高达90%以上。
2hpvl6e6蛋白纳米抗体文库的构建
(1)hpvl6e6蛋白免疫新疆双峰骆驼
hpvl6e6蛋白常规免疫新疆双峰骆驼,共免疫注射7次;免疫结束后,取骆驼外周血200ml。
(2)外周血淋巴细胞的分离及其总rna的提取
采用percoll密度梯度离心分离纯化骆驼外周血淋巴细胞。然后用trizol法提取总rna,溶解于depc水。
(3)逆转录pcr
i.采用takara公司的rt-pcr试剂盒,将外周血淋巴细胞总rna逆转录成cdna。
ii.通过nest-pcr扩增骆驼重链抗体的vhh片段
a.以上述所得的cdna为模板,进行第一轮扩增反应。引物如下:
nest-pcr1up(seqidno:10):5'>gtcctggctgctcttctacaagg<3';
nest-pcr1down(seqidno:11):5'>ggtacgtgctgttgaactgttcc<3'
pcr反应结束后,琼脂糖凝胶电泳检测,然后切胶回收目的条带。图2凝胶电泳结果显示,扩增基因片段在700bp处有特异性条带。
b.第二轮pcr反应
以nestpcr1dna回收产物为模板,进行nest-pcr2扩增反应。引物如下:
nest-pcr2up(seqidno:12):5'>ccggaattctcaggtgcagctggtggagtctgg<3';
nest-pcr2down(seqidno:13):5'>gcccaagctttgaggagacggtgacctgggt<3'
pcr反应结束后,琼脂糖凝胶电泳检测pcr产物,切胶回收目的条带,即为vhh片段。图3凝胶电泳结果显示,扩增基因片段在500bp处有特异性条带。
(4)vhh片段的双酶切与回收
将第二次pcr产物进行ecorⅰ和hindⅲ双酶切反应。
(5)t7噬菌体抗体库的构建
使用novagen公司t7select10试剂盒构建纳米抗体库。将酶切回收的vhh片段和t7载体连接,进行t7噬菌体包装及抗体库滴度测定,滴度大约为1×107pfu/ml。
(6)t7噬菌体抗体库的扩增
选用液体裂解法对t7噬菌体抗体库进行扩增。扩增后噬菌斑滴度为1.2×1010pfu/ml。
(7)抗hpvl6e6纳米抗体文库的重组阳性率测定
随机挑(5)中的噬菌斑提取噬菌体dna;按照试剂盒说明书,进行pcr反应,琼脂糖凝胶电泳检测pcr产物(见图4)。
pcr反应条件及程序为:95℃5分钟;95℃30秒,55℃30秒,72℃30秒,20个循环;72℃7分钟。
图4电泳结果显示,构建的抗hpvl6e6纳米抗体文库的重组阳性率为100%。
3运用镍离子金属螯合亲和层析介质ni-nta淘选抗hpv16e6纳米抗体
(1)灭菌水清洗ni-nta介质
(2)0.5%bsa封闭液封闭ni-nta介质
(3)负筛去除非特异结合的噬菌体:
抗hpvl6e6的噬菌体库用tbst稀释至1ml,加入到封闭后的ni-nta介质中,室温结合30min;离心,上清即为负筛后的t7噬菌体库。
(4)与hpvl6e6特异结合的噬菌体的筛选:
负筛后的抗hpvl6e6噬菌体库加入hpvl6e6蛋白(1ug/ul),室温结合30min。然后将混合物加入到封闭好的ni-nta介质中,室温结合30min;离心,弃上清。tbst洗获得的沉淀,共洗5次;离心,弃上清;加入400ultb培养基混匀,平均分为两份,一份用来测定筛选后的噬菌体滴度,一份用来扩增筛选后的噬菌体。
(5)筛选后的噬菌体的扩增:
将筛选后的噬菌体加入到50mlod600=0.6的blt5403宿主菌中,37℃震荡培养至有白色絮状沉淀出现时,停止培养;离心,上清即为扩增的第一轮筛选后的噬菌体,用于下一轮的筛选;筛选3-4轮。
4elisa鉴定特异性的抗hpvl6e6的噬菌体阳性克隆
最后一轮筛选所得的噬菌体,在tb固体培养基上培养,随机挑选70个单克隆噬菌斑于blt5403宿主菌中进行液体裂解法扩增,离心,上清于4℃保存。
elisa板每孔加入hpvl6e6蛋白包被,4℃过夜,第二天0.5%bsa室温封闭1h;实验组每孔加入扩增的单克隆噬菌体,对照组加入等量的野生型t7噬菌体,室温孵育1h;tbst洗,然后加入兔抗t7噬菌体10a抗体,室温孵育1h;tbst洗,然后加入hrp标记的羊抗兔抗体,室温孵育1h;tbst洗,加入显色液测吸光值,当实验孔与对照孔吸光值比值大于2时,判定其为阳性克隆;提取阳性克隆噬菌体的dna进行测序。elisa鉴定获得30个阳性克隆。经过dna测序,分析其氨基酸序列,获得的阳性单克隆噬菌体的核苷酸序列,氨基酸序列如下:
核苷酸序列(seqidno:9):
gatgtgcagctggtggagtctgggggaggcttggtgcagcctgggggctctctgagactctcctgtgcagcctctggattcaccttcggtgattatggcatgagctgggtccgccaggctccaggaaagggactcgagtgggtctcaaatattcgtagtggtcctgatagcacaaactatgcagactccgtgaagggccgattcaccatttccagagacgacagcaagagtacgctgtatctgcaaatgaacagtttgaaacctgaggacacagccacgtattactgcgccactgaccgtggagggcgtacgcgccgaggccaggggacccaggtcaccgtctcctca
氨基酸序列(seqidno:8):
dvqlvesggglvqpggslrlscaasgftfgdygmswvrqapgkglewvsnirsgpdstnyadsvkgrftisrddskstlylqmnslkpedtatyycatdrggrtrrgqgtqvtvss
框架区(fr1-fr4)和互补决定区(cdr1-cdr3)氨基酸序列:
fr1(seqidno:1):dvqlvesggglvqpggslrlscaas
cdr1(seqidno:2):gftfgdygms
fr2(seqidno:3):wvrqapgkglewv
cdr2(seqidno:4):snirsgpdstn
fr3(seqidno:5):yadsvkgrftisrddskstlylqmnslkpedtatyyc
cdr3(seqidno:6):atdrggrtr
fr4(seqidno:7):rgqgtqvtvss
5抗hpvl6e6纳米抗体的表达纯化
(1)hpvl6e6蛋白纳米抗体的诱导表达
将抗hpvl6e6纳米抗体的核苷酸序列用ecorⅰ和hindⅲ内切酶进行双酶切,电泳切胶回收酶切产物,插入到pet28a中,构建抗hpvl6e6纳米抗体的表达载体;转化到bl21(de3)中,iptg诱导表达抗hpvl6e6纳米抗体。
(2)抗hpvl6e6纳米抗体的纯化
通过ni-ida亲和层析方法纯化抗hpvl6e6纳米抗体。层析柱首先用菌体缓冲液清洗,加入ni-ida填充层析柱;将上述抗hpvl6e6纳米抗体表达菌菌体超声上清加入镍柱中,控制流速;用清洗缓冲液(40mmol/l咪唑)洗掉杂蛋白;用洗脱缓冲液(250mmol/l咪唑)洗脱目的蛋白,并收集洗脱液。然后用15%sds-page电泳检测纯化的抗hpvl6e6纳米抗体。(见图5)检测结果显示,纯化的抗hpvl6e6纳米抗体在20kd处有一条带,符合目的蛋白预期分子量。
6抗hpvl6e6纳米抗体的免疫印迹亲和性分析
对psumo-hpvl6e6质粒表达的hpv16e6蛋白及对照空质粒表达sumo蛋白进行westernblot鉴定,结果表明抗hpvl6e6纳米抗体与空载体表达的sumo蛋白不结合,与hpvl6e6蛋白具有良好的结合能力,在大约35kda处有清晰的目标条带(图6a)。
western检测抗hpv16e6纳米抗体对hpv16阳性细胞株的亲和性:鉴定结果显示抗hpv16e6纳米抗体能够特异识别hpv16阳性宫颈癌细胞株siha细胞内的hpv16e6,人永生化表皮细胞hacat为对照,actin为内参(图6b)。
7elisa检测anti-hpvl6e6纳米抗体的亲和性
以hpvl6e6蛋白和psumo-hpvl6e6空质粒表达sumo蛋白包被酶标板,4℃过夜。tbst洗板3次,加入0.5%bsa封闭液,室温封闭一小时。弃封闭液并用tbst洗3次,加入抗hpvl6e6纳米抗体,或对照抗wasabi纳米抗体,室温孵育一小时。tbst洗板3次后,加入1:1000稀释的兔抗ha单克隆抗体,室温孵育一小时。tbst洗板3次,加入1:2000稀释的hrp标记羊抗兔二抗,室温孵育一小时。tbst洗板3次,加入显色液,于酶标仪上测450nm吸光值。检测结果显示:抗hpvl6e6纳米抗体对hpvl6e6的结合活性远远高于抗wasabi纳米抗体对照组及psumo空质粒表达sumo蛋白组。结果如表1所示:
表1elisa检测hpvl6e6纳米抗体的亲和性的450nm吸光值
序列表
<110>东南大学
<120>能够特异结合hpvl6-e6蛋白的纳米抗体及其编码序列、制备方法和应用
<160>13
<170>siposequencelisting1.0
<210>1
<211>25
<212>prt
<213>骆驼(camel)
<400>1
aspvalglnleuvalgluserglyglyglyleuvalglnproglygly
151015
serleuargleusercysalaalaser
2025
<210>2
<211>10
<212>prt
<213>骆驼(camel)
<400>2
glyphethrpheglyasptyrglymetser
1510
<210>3
<211>13
<212>prt
<213>骆驼(camel)
<400>3
trpvalargglnalaproglylysglyleuglutrpval
1510
<210>4
<211>11
<212>prt
<213>骆驼(camel)
<400>4
serasnileargserglyproaspserthrasn
1510
<210>5
<211>37
<212>prt
<213>骆驼(camel)
<400>5
tyralaaspservallysglyargphethrileserargaspaspser
151015
lysserthrleutyrleuglnmetasnserleulysprogluaspthr
202530
alathrtyrtyrcys
35
<210>6
<211>9
<212>prt
<213>骆驼(camel)
<400>6
alathraspargglyglyargthrarg
15
<210>7
<211>11
<212>prt
<213>骆驼(camel)
<400>7
argglyglnglythrglnvalthrvalserser
1510
<210>8
<211>116
<212>prt
<213>骆驼(camel)
<400>8
aspvalglnleuvalgluserglyglyglyleuvalglnproglygly
151015
serleuargleusercysalaalaserglyphethrpheglyasptyr
202530
glymetsertrpvalargglnalaproglylysglyleuglutrpval
354045
serasnileargserglyproaspserthrasntyralaaspserval
505560
lysglyargphethrileserargaspaspserlysserthrleutyr
65707580
leuglnmetasnserleulysprogluaspthralathrtyrtyrcys
859095
alathraspargglyglyargthrargargglyglnglythrglnval
100105110
thrvalserser
115
<210>9
<211>348
<212>dna
<213>骆驼(camel)
<400>9
gatgtgcagctggtggagtctgggggaggcttggtgcagcctgggggctctctgagactc60
tcctgtgcagcctctggattcaccttcggtgattatggcatgagctgggtccgccaggct120
ccaggaaagggactcgagtgggtctcaaatattcgtagtggtcctgatagcacaaactat180
gcagactccgtgaagggccgattcaccatttccagagacgacagcaagagtacgctgtat240
ctgcaaatgaacagtttgaaacctgaggacacagccacgtattactgcgccactgaccgt300
ggagggcgtacgcgccgaggccaggggacccaggtcaccgtctcctca348
<210>10
<211>23
<212>dna
<213>人工序列(artificialsequence)
<400>10
gtcctggctgctcttctacaagg23
<210>11
<211>23
<212>dna
<213>人工序列(artificialsequence)
<400>11
ggtacgtgctgttgaactgttcc23
<210>12
<211>33
<212>dna
<213>人工序列(artificialsequence)
<400>12
ccggaattctcaggtgcagctggtggagtctgg33
<210>13
<211>31
<212>dna
<213>人工序列(artificialsequence)
<400>13
gcccaagctttgaggagacggtgacctgggt31
- 还没有人留言评论。精彩留言会获得点赞!