一种降低黄酒酵母尿素积累的方法及应用与流程
本发明涉及一种降低黄酒酵母尿素积累的方法及应用,属于遗传工程领域。
背景技术:
小分子介导的蛋白二聚化是广泛应用于调控蛋白亚细胞定位的有效手段之一。雷帕霉素可以使fk506结合蛋白——fkbp12与mtor的fbr亚基结合,形成异源二聚体。以此为基础,通过将目的蛋白与fkbp12蛋白融合表达、将已知亚细胞定位的锚定蛋白与fbr亚基融合,在雷帕霉素的介导下,可以形成目的蛋白-fkbp12-雷帕霉素-fbr-锚定蛋白复合体。目的蛋白在锚定蛋白的作用下,定位到目标亚细胞器中。
dal80是酿酒酵母氮代谢阻遏效应全局调控因子之一,主要在细胞核内通过与相关基因启动子区域的结合,阻遏基因的转录表达。黄酒生成是酿酒酵母的发酵过程,受氮代谢阻遏效应的影响,尿素等非偏好型氮源在发酵过程中利用滞后,最终在发酵体系中积累。积累的尿素与乙醇自发反应会生成2a类潜在致癌物质氨基甲酸乙酯,严重影响黄酒产品的安全性。因此阻止转录阻遏因子dal80进入细胞核,消除dal80对尿素利用相关基因表达的阻遏作用,是降低黄酒发酵过程中氨基甲酸乙酯前体物质——尿素积累的途径之一。
技术实现要素:
本发明的第一个目的是提供一种降低黄酒酵母尿素积累的方法,通过添加雷帕霉素调控dal80定位于细胞质,降低黄酒酵母发酵过程中尿素的积累。
在本发明的一种实施方式中,所述方法在黄酒酵母基因组上,融合表达dal80、fkbp12,同时在质粒载体上融合表达rps1b、fbr亚基,在雷帕霉素作用下,形成dal80-fkbp12-雷帕霉素-rps1b-fbr蛋白复合体,使dal80在rps1b蛋白的作用下定位于细胞质中,无法进入细胞核,降低黄酒发酵过程中尿素的积累。
在本发明的一种实施方式中,所述方法在黄酒酵母基因组上融合表达dal80、fkbp12、在质粒上融合表达rps1b、frb,通过添加雷帕霉素,使fkbp12和frb结合,并使dal80在细胞质定位蛋白rps1b的作用下,转移到细胞质。
本发明的第二个目的是提供一种构建黄法酒酵母中尿素含量降低的黄酒酵母,主要包括如下步骤:
(1)构建dal80、fkbp12重组框;
(2)通过crispr-cas9系统将dal80、fkbp12重组框整合到重组黄酒酵母jnz01(mata,δura3,δtrp1,tor1s1972r,δfpr1)基因组dal80基因之后,得到重组黄酒酵母jnz8(mata,δura3,δtrp1,tor1s1972r,δfpr1,dal80::fkbp12);
(3)消除crispr-cas9系统质粒;
(4)构建rps1b、frb融合表达框;
(5)将rps1b、frb融合表达框插入游离高拷贝表达质粒prs426-tef-ura3中,得到重组质粒prs426-tef-rps1b-frb-ura3,转化重组黄酒酵母jnz8得到jnz9(mata,δura3,δtrp1,tor1s1972r,δfpr1,dal80::fkbp12,prs426-tef-rps1b-frb-ura3);
(6)在重组黄酒酵母的培养过程中,向培养基中添加雷帕霉素,诱导dal80定位于细胞质。
在本发明的一种实施方式中,所述方法具体为:
(1)通过融合pcr构建dal80、fkbp12重组框;
(2)通过crispr-cas9系统将dal80、fkbp12重组框整合到重组黄酒酵母jnz01(mata,δura3,δtrp1,tor1s1972r,δfpr1)基因组dal80基因之后,得到重组黄酒酵母jnz8(mata,δura3,δtrp1,tor1s1972r,δfpr1,dal80::fkbp12);
(3)通过在ypd培养基中传代,丢失crispr-cas9系统质粒,在5-foa和5-faa平板上进行筛选;
(4)通过融合pcr构建rps1b、frb融合表达框;
(5)通过酶切连接将rps1b、frb融合表达框插入游离高拷贝表达质粒prs426-tef-ura3中,得到重组质粒prs426-tef-rps1b-frb-ura3,转化重组黄酒酵母jnz8得到jnz9(mata,δura3,δtrp1,tor1s1972r,δfpr1,dal80::fkbp12,prs426-tef-rps1b-frb-ura3)。
在本发明的一种实施方式中,所述重组黄酒酵母jnz01的构建方法为:以黄酒酵母xz-11为出发菌株,敲除其fpr1基因,并突变tor1基因编码的蛋白第1972位丝氨酸残基为精氨酸。
本发明的第三个目的是提供应用所述方法构建的黄酒酵母。
本发明的第四个目的是提供所述黄酒酵母在食品领域的应用。
本发明还要求保护所述黄酒酵母在黄酒生产领域的应用。
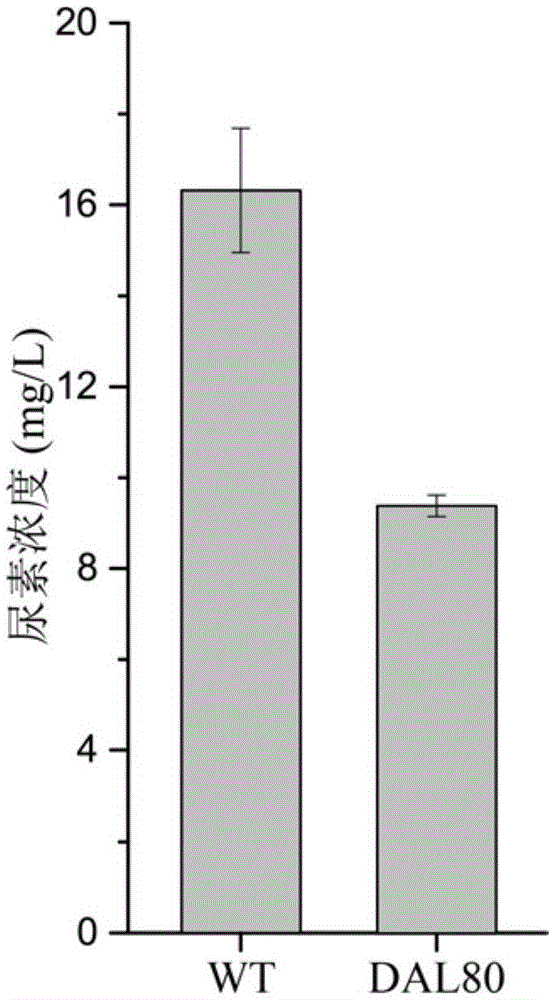
有益效果:本发明提供的通过雷帕霉素介导dal80与rps1b的二聚化降低黄酒酵母尿素积累的方法,可使构建的重组黄酒酵母发酵过程中尿素的积累降低42.43%,降低到9.39mg/l。
附图说明
图1雷帕霉素介导dal80与rps1b的二聚化对菌株尿素积累的影响;
图2为调控gzf3定位于细胞质对菌株尿素代谢相关基因表达的影响。
具体实施方式
实施例1适用于雷帕霉素介导调控蛋白亚细胞定位的黄酒酵母的构建
(1)拆分用于黄酒生产的二倍体酿酒酵母菌株xz-11获得单倍体菌株
按论文“wudh,lixm,shenc,etal.isolationofahaploidfromanindustrialchinesericewineyeastformetabolicengineeringmanipulation[j].journaloftheinstituteofbrewing,2013,119(4):288-293.”中所述方法及步骤获得不含抗性基因的单倍体xz-11a菌株。
(2)构建营养缺陷型单倍体黄酒酵母jnz01
以黄酒酵母xz-11菌株基因组为模板,分别扩增ura3基因上、下游300bp序列,通过融合pcr将上述两个扩增片段融合得到ura3基因敲除框。将ura3敲除框转化xz-11a菌株,在5-foa平板(ynb1.7g/l,硫酸铵5g/l,葡萄糖20g/l,尿嘧啶25mg/l,5-foa1g/l,琼脂粉20g/l)上筛选,获得尿嘧啶缺陷型菌株xz-11a-ura3。以黄酒酵母xz-11菌株基因组为模板,分别扩增trp1基因上、下游300bp序列,通过融合pcr将上述两个扩增片段融合得到trp1基因敲除框。将trp1敲除框转化xz-11a-ura3菌株,在5-faa平板(ynb1.7g/l,硫酸铵5g/l,葡萄糖20g/l,尿嘧啶25mg/l,色氨酸25mg/l,5-faa1g/l,琼脂粉20g/l)上筛选,获得尿嘧啶和色氨酸二重营养缺陷性菌株jnz01(mataδura3,δtrp1)。
(3)构建prs426-tor1sgrna-fpr1sgrna-ura3质粒
首先根据yeastriction(http://yeastriction.tnw.tudelft.nl/#!/)设计运用crispr-cas9系统编辑基因组上fpr1和tor1位点所需的20nt序列(fpr1:ttggttaccattcattacac;tor1:tgttatgttcaacgaaaaat)。以质粒p426-snr52p-grna.can1.y-sup4t(addgene编号43803)为模板,通过融合pcr分别获得fpr1sgrna和tor1sgrna编码框,并将原定位到can1位点的20nt序列分别替换为定位到fpr1和tor1的20nt序列。通过酶切连接,分别将fpr1sgrna和tor1sgrna克隆到质粒prs426-ura3的kpni和xhoi、spei和saci位点,得到质粒prs426-fpr1sgrna-tor1sgrna-ura3。
(4)构建fpr1基因敲除框和tor1基因点突变重组框
以基因组为模板,分别扩增fpr1基因上下游各200bp的片段,通过融合pcr得到fpr1基因的敲除框。以基因组为模板,分别扩增tor1基因5801~5923bp和5911~6000bp间序列,在引物中将5914~5919bp区域的序列由原始的agccgc突变为cgtcga,将1972位的丝氨酸突变为精氨酸,同时在1973位上引入同义突变产生一个sali酶切位点。通过融合pcr,将两段片段融合为tor1基因点突变重组框。
(5)crispr-cas9系统一步敲除fpr1基因和点突变tor1基因
将质粒p414-tef1p-cas9-cyc1t(addgene编号43802)转化到出发菌株黄酒酵母xz-11中,在sc-trp1平板(ynb1.7g/l,硫酸铵5g/l,葡萄糖20g/l,尿嘧啶25mg/l,琼脂粉20g/l)筛选阳性转化子。将质粒p426-fpr1sgrna-tor1sgrna-ura3、fpr1基因敲除框和tor1基因点突变重组框一起转化带有p414-tef1p-cas9-cyc1t质粒的出发菌,在sc平板(ynb1.7g/l,硫酸铵5g/l,葡萄糖20g/l,琼脂粉20g/l)上筛选。通过菌落pcr筛选同时敲除fpr1基因和点突变tor1基因的重组菌,得到阳性转化子jnz0(mata,δura3,δtrp1,tor1s1972r,δfpr1,p426-fpr1sgrna-tor1sgrna-ura3,p414-tef1p-cas9-cyc1t)。
(6)消除crispr-cas9系统质粒
将筛选得到的阳性转化子于ypd培养基(酵母膏10g/l,蛋白胨20g/l,葡萄糖20g/l)中传代培养,每培养24h后以10%接种量转接到新鲜的ypd培养基中。每次转接同时在含有5-foa和5-faa的平板(ynb1.7g/l,硫酸铵5g/l,葡萄糖20g/l,尿嘧啶25mg/l,色氨酸25mg/l,5-foa1g/l,5-faa1g/l,琼脂粉20g/l)上划线,直至在含有5-foa和5-faa的平板上获得转化子。通过菌落pcr验证转化子中p426-fpr1sgrna-tor1sgrna-ura3和p414-tef1p-cas9-cyc1t已消除,得到重组黄酒酵母jnz01(mata,δura3,δtrp1,tor1s1972r,δfpr1)。
实施例2融合pcr构建融合表达dal80、fkbp12重组框
首先通过全质粒pcr将(ggggs)3linker引入到高拷贝质粒载体prs426-tef-ura3上bamhi位点之后,得到prs426-tef-gs-ura3。通过基因合成获得fkbp12,通过酶切连接克隆到载体prs426-tef-gs-ura3的ecori和xhoi位点,得到prs426-tef-gs-fkbp12-ura3。从基因组上扩增得到dal80末尾不含终止密码子的500bp序列,通过酶切连接克隆到载体prs426-tef-gs-fkbp12-ura3的spei和bamhi位点,得到prs426-tef-dal80d-gs-fkbp12-ura3。然后以得到的质粒为模板,进行pcr扩增,通过在上游引物中引入dal80下游50bp序列,得到融合表达dal80、fkbp12重组框。
实施例3通过crispr-cas9系统在重组黄酒酵母jnz01基因组上整合融合表达dal80、fkbp12重组框
将质粒p414-tef1p-cas9-cyc1t(addgene编号43802)转化到重组黄酒酵母jnz01中,在sc-trp1平板(ynb1.7g/l,硫酸铵5g/l,葡萄糖20g/l,尿嘧啶25mg/l,琼脂粉20g/l)筛选阳性转化子,得到菌株jnz01-cas9。根据yeastriction(http://yeastriction.tnw.tudelft.nl/#!/)设计运用crispr-cas9系统编辑基因组上dal80位点所需的20nt序列(tacatcctttcttaatgttg)。以质粒p426-snr52p-grna.can1.y-sup4t(addgene编号43803)为模板,通过全质粒pcr将20nt序列替换掉原质粒上定位到can1位点的20nt序列,得到p426-dal80sgrna。将质粒p426-dal80sgrna和融合表达dal80、fkbp12重组框一起转化菌株jnz01-cas9,在sc培养基(ynb1.7g/l,硫酸铵5g/l,葡萄糖20g/l,琼脂粉20g/l)上筛选。通过菌落pcr筛选融合表达dal80、fkbp12重组框整合到基因组上的阳性转化子。
实施例4消除crispr-cas9系统质粒
将筛选得到的阳性转化子于ypd培养基(酵母膏10g/l,蛋白胨20g/l,葡萄糖20g/l)中传代培养,每培养24h后以10%接种量转接到新鲜的ypd培养基中。每次同时转接在5-foa和5-faa平板(ynb1.7g/l,硫酸铵5g/l,葡萄糖20g/l,尿嘧啶25mg/l,色氨酸25mg/l,5-foa1g/l,5-faa1g/l,琼脂粉20g/l)上划线,直至5-foa和5-faa平板上克隆同时能在两种平板上生长。通过菌落pcr验证转化子中p426-dal80sgrna和p414-tef1p-cas9-cyc1t已消除,得到重组黄酒酵母jnz8。
实施例5融合表达rps1b、frb
通过基因合成获得frb,通过酶切连接克隆到载体prs426-tef-gs-ura3的ecori和xhoi位点,得到prs426-tef-gs-frb-ura3。从重组黄酒酵母jnz01(mata,δura3,δtrp1,tor1s1972r,δfpr1)基因组上扩增得到不含终止密码子的rps1b基因序列,通过酶切连接克隆到载体prs426-tef-gs-frb-ura3的spei和bamhi位点,得到prs426-tef-rps1b-gs-frb-ura3。将质粒prs426-tef-rps1b-gs-frb-ura3转化重组黄酒酵母jnz8,在sc-ura3平板(ynb1.7g/l,硫酸铵5g/l,葡萄糖20g/l,色氨酸25mg/l,琼脂粉20g/l)筛选阳性转化子,得到菌株jnz9。
实施例6应用jnz9菌株进行发酵降低发酵液中尿素积累量
在发酵培养基(ynb1.7g/l,硫酸铵5g/l,葡萄糖20g/l,20种必需氨基酸各5g/l,尿素5g/l,雷帕霉素1μg/ml)中用jnz9和jnz01进行发酵。30℃220rpm培养48h后,利用高效液相色谱检测发酵液中尿素含量(检测方法同论文“zhangw,chengy,liy,etal.adaptiveevolutionrelievesnitrogencataboliterepressionanddecreasesureaaccumulationinculturesofthechinesericewineyeaststrainsaccharomycescerevisiaexz-11[j].jagricfoodchem,2018,66(34):9061-9069.”中的方法)。最终调控dal80定位于细胞质后,发酵液中尿素含量降低为9.39mg/l,比不调控的情况降低42.43%。
实施例7应用jnz9菌株在黄酒发酵体系中降低尿素
以jnz01和jnz9进行黄酒模拟发酵。取100g糯米,在室温中浸渍2-3天;经常压蒸煮后,添加17.4g麦曲(生麦曲13.5g,熟麦曲3.4g)、170g水,并将经活化后的jnz01和jnz9菌株按10%(v/v)的接种量接种,加入雷帕霉素至终浓度为1μg/ml。摇瓶加发酵栓后于30℃静置发酵,每日称重,当日失重小于2g时结束前酵。将摇瓶置于15℃继续静置发酵,15天后结束后酵。检测发酵体系中尿素含量,结果显示,调控dal80定位于细胞质后,发酵体系中尿素含量分别降低为9.45mg/l,比出发菌分别降低29.42%。
实施例8调控dal80定位于细胞质强化尿素代谢途径
通过定量pcr技术分别在谷氨酰胺+尿素为氮源和单独尿素为氮源条件下检测尿素途径相关基因和ncr调控因子表达上下调情况。将在平板上活化的酵母菌株接种到ynb-ura培养基中,30℃,220rpm中过夜培养。将活化后的酵母稀释至od600=0.2-0.3,转接到ypd或ynb添加特异氨基酸氮源的培养基中进行培养。培养结束后离心收集菌体,用超纯水洗涤2次,保存于液氮中或-80℃冰箱待用。酵母总rna提取按照酵母总rna提取试剂盒的说明书操作,提取的酵母总rna随即按照takara反转录试剂盒的说明书合成cdna,cdna用于荧光定量pcr,荧光定量pcr所用引物由软件beacondesigner7设计并经过hplc纯化,退火温度设定为55℃,片段长度为120~150bp。利用2×sybrpremixextaq在lightcycler480荧光定量pcr仪上进行扩增和数据采集。act1基因为内参基因。表2定量pcr所用引物。
表2定量pcr引物
结果如图2所示,在氮源偏好型培养基中,在dal80被定位在细胞质的情况下,尿素降解酶编码基因dur1,2表达上调幅度是jnz01的6.91倍;尿素转运蛋白编码基因dur3是jnz01的3.92倍,精氨酸降解酶编码基因car1是jnz01的5.59倍,广谱型氨基酸转运蛋白编码基因gap1是jnz01的3.45倍;在氮源非偏好型培养基中,在gzf3被定位在细胞质的情况下,与尿素代谢直接相关的基因dur1,2、dur3、car1的转录水平分别比jnz01提高4.58倍、2.55倍、3.37倍。
虽然本发明已以较佳实施例公开如上,但其并非用以限定本发明,任何熟悉此技术的人,在不脱离本发明的精神和范围内,都可做各种的改动与修饰,因此本发明的保护范围应该以权利要求书所界定的为准。
sequencelisting
<110>江南大学
<120>一种降低黄酒酵母尿素积累的方法及应用
<160>4
<170>patentinversion3.3
<210>1
<211>20
<212>dna
<213>人工序列
<400>1
ttggttaccattcattacac20
<210>2
<211>20
<212>dna
<213>人工序列
<400>2
tgttatgttcaacgaaaaat20
<210>3
<211>15
<212>prt
<213>人工序列
<400>3
glyglyglyglyserglyglyglyglyserglyglyglyglyser
151015
<210>4
<211>20
<212>dna
<213>人工序列
<400>4
tacatcctttcttaatgttg20
- 还没有人留言评论。精彩留言会获得点赞!