使用FAD依赖性葡糖脱氢酶的连续葡萄糖监测的制作方法
本发明涉及使用fad依赖性葡糖脱氢酶的连续葡萄糖监测。
背景技术:
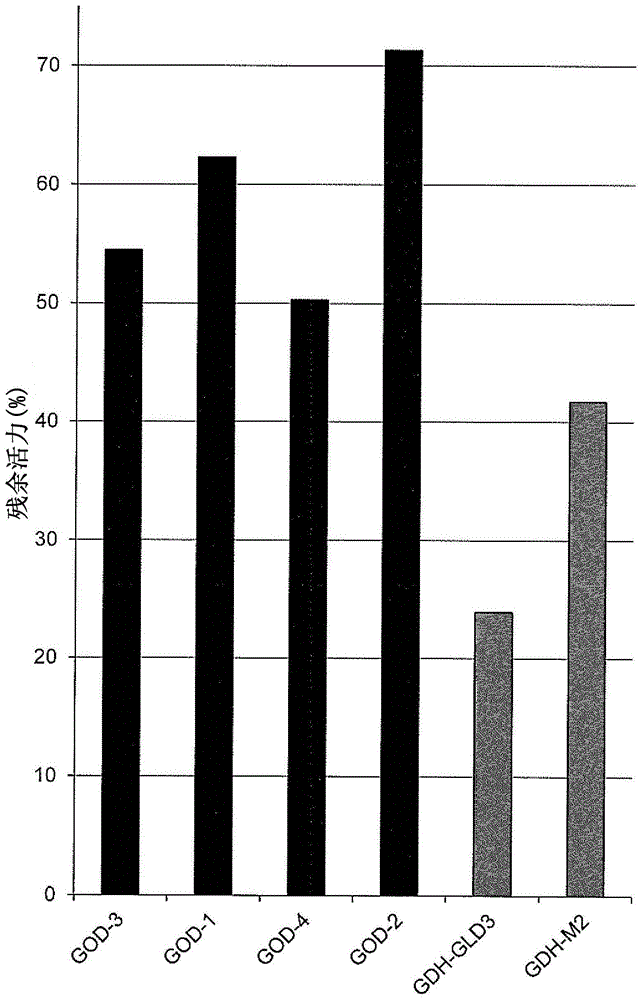
:连续葡萄糖监测(cgm,也称为快速葡萄糖监测,fgm)系统对于糖尿病的诊断和控制是重要的。cgm系统能够在几天至约1-2周的一段时间内测量血糖水平。cgm系统包括葡萄糖传感器。可以用葡萄糖氧化酶(god)进行葡萄糖水平的测量。当氧化葡萄糖时,葡萄糖氧化酶使用氧作为电子受体。这可能受测量样品中存在的溶解氧的影响。目前,就我们所知,可商购的cgm装置全部基于葡萄糖氧化酶(god)。某些god,如来自属于曲霉属(aspergillussp.)的微生物,如黑曲霉(aspergillusniger)的god已知即使置于严苛的热状况下时,仍保持酶活力(非专利文献1:thejournalofbiologicalchemistryvol.278,no.27,issueofjuly4,pp.24324-24333,2003)。为此,一般说来,cgm系统使用热稳定的god。专利文献文档1(wo2015/099112,us20160319246)描述了来自属于毛霉属(mucor)的微生物的gdh。非专利文献2(satake等人,jbioscibioeng.2015nov;120(5):498-503)描述了来自普雷恩毛霉(mucorprainii)的fad-gdh。该文档说明“基于其高热稳定性和高葡萄糖选择性,葡萄糖氧化酶(god)(ec1.1.3.4)已广泛用作介导的电流测量的葡萄糖传感器”。专利文献文档2(wo2002/036779,us2004/0023330)描述了来自洋葱伯克霍尔德菌(burkholderiacepacia)的gdh。所公开的gdh是膜结合蛋白并且包括细胞色素c域。洋葱伯克霍尔德菌(b.cepacia)gdh的氨基酸序列与来自属于毛霉属(mucor)的微生物的fad依赖性gdh(在本文中也称为fad-gdh)的氨基酸序列不类似。专利文献文档3(wo2013/051682)描述了来自季也蒙毛霉(mucorguilliermondii)的gdh(mggdh)。专利文献文档4(jp专利申请公开(kokai)no.2013-116102)描述了来自冻土毛霉(mucorhiemalis)的gdh(mhgdh)。专利文献文档5(wo2013/022074,us20140154777)描述了来自单卷霉(circinellasimplex)的csgdh和来自卷霉属(circinella)种的crgdh。专利文献文档6(jp专利申请公开(kokai)no.2013-176363)描述了来自毛霉属(mucor)rd056860的mdrgdh。专利文献文档7(jp专利申请公开(kokai)no.2013-176364)描述了来自细孢毛霉(mucorsubtilissimus)的msgdh。专利文献文档8(wo2012/169512,us20140287445)描述了来自普雷恩毛霉(m.prainii)的gdh。专利文献文档9(wo2015/129475,ep3112461)描述了来自普雷恩毛霉(m.prainii)的gdh。专利文献文档10(jp专利申请公开(kokai)no.2014-207911)描述了pqq依赖性gdh。据报道所公开的pqq依赖性gdh对麦芽糖具有活力。已报道了实现基于pqq依赖性gdh的葡萄糖传感器中的困难。一个原因是相对低的底物特异性,即已知除葡萄糖之外,pqq依赖性gdh对麦芽糖起作用。参见非专利文献3。引用列表专利文献ptl1:wo2015/099112(us20160319246)ptl2:wo2002/036779(us2004/0023330)ptl3:wo2013/051682ptl4:jp专利申请公开(kokai)no.2013-116102(美国专利no.9,260,699)ptl5:wo2013/022074(us20140154777)ptl6:jp专利申请公开(kokai)no.2013-176363ptl7:jp专利申请公开(kokai)no.2013-176364ptl8:wo2012/169512(us20140287445)ptl9:wo2015/129475(ep3112461)ptl10:jp专利申请公开(kokai)no.2014-207911非专利文献npl1:thejournalofbiologicalchemistryvol.278,no.27,issueofjuly4,pp.24324-24333,2003npl2:satake,etal,jbioscibioeng.2015nov;120(5):498-503npl3:reviewofadverseeventsassociatedwithfalseglucosereadingsmeasuredbygdh-pqq-basedglucoseteststripsinthepresenceofinterferingsugars",diabetescare.2010apr;33(4):728-729技术实现要素:技术问题本发明的目标是提供改善的cgm方法。问题的解决方法在理论上,可以通过包含葡糖脱氢酶(gdh)或者葡萄糖氧化酶(god)的葡萄糖传感器来进行连续葡萄糖监测。当使葡萄糖脱氢时,葡糖脱氢酶使用电子受体。这可能是有利的,其中测量可以不受溶解氧影响。然而,就本发明人的知识所及,当前可用的商品化cgm装置全部基于葡萄糖氧化酶(god)。据信这是由于god的热稳定性。某些god,如来自属于曲霉属(aspergillussp.)的微生物,如黑曲霉(aspergillusniger)的god普遍已知即使当在严苛的热状况下放置较短的一段时间,仍保持酶活力(参见非专利文献1)。已知当处于严苛的热状况下时,god保持了高于gdh的活力,特别是就适合于葡萄糖传感器的酶来说。这表示god比gdh更稳定。然而,本发明人意外地发现与处于相同条件下的可商购的god相比,某些fad依赖性葡糖脱氢酶在环境温度下,在较长的一段时间内保持了更高的酶活力。鉴于以下事实这是特别意外的:当置于高温下,例如,处于包括热处理的加速试验下时,与所述fad依赖性葡糖脱氢酶相比,可商购的god保持更高的酶活力。考虑到这些发现,本文提供了包括使用fad-gdh的连续葡萄糖监测(cgm)方法。fad-gdh能够在特定的一段时间内保持初始活力。本文还提供了筛选适合在cgm中使用的fad-gdh的方法以及包括fad-gdh的cgm装置。提供了以下示例性实施方式:实施方式1.连续葡萄糖监测方法,包括使用具有以下能力的gdh(a)当保持在约37-40℃时,在1周内保持初始活力的约60%或以上,(b)当保持在约37-40℃时,在2周内保持初始活力的约40%或以上,或者(c)当保持在约37-40℃时,在3周内保持初始活力的约30%或以上,其中所述gdh是fad依赖性gdh并且其中所述gdh不是膜结合蛋白。实施方式2.根据实施方式1所述的方法,其中当保持在ph7.4时,gdh能够保持实施方式1的(a)至(c)中所指明的活力。实施方式3.根据实施方式1或2所述的方法,其中可以在不进行再校准的情况下进行连续葡萄糖监测。实施方式4.筛选在连续葡萄糖监测中使用的gdh的方法,所述方法包括以下步骤:(i)制备候选gdh,(ii)将所述候选gdh在约30-40℃保持预定时间段,(iii)在步骤(ii)之后确定所述gdh的残余活力,(iv)将步骤(iii)中所确定的残余活力与所述候选gdh的初始活力相比较,其中,当所述残余gdh活力(a)当在约30-40℃保持1周时,与初始gdh活力相比,为约60%或以上,(b)当在约30-40℃保持2周时,与初始gdh活力相比,为约40%或以上,或者(c)当在约30-40℃保持3周时,与初始gdh活力相比,为约30%或以上,则将所述候选gdh选作具有在连续葡萄糖监测中使用1至3周的潜力的gdh。实施方式5.根据实施方式4所述的方法,其中步骤(ii)未替换为包括在约50-60℃的温度下热处理约5-30分钟的加速热处理步骤。实施方式6.连续葡萄糖监测装置,其包括具有以下能力的gdh(a)当保持在约37-40℃时,在1周内保持初始活力的约60%,(b)当保持在约37-40℃时,在2周内保持初始活力的约40%,或者(c)当保持在约37-40℃时,在3周内保持初始活力的约30%,其中所述gdh是fad依赖性gdh并且其中所述gdh不是膜结合蛋白。实施方式7.根据实施方式6所述的装置,其中当保持在ph7.4时,所述gdh能够保持实施方式6的(a)至(c)中所指明的活力。实施方式8.根据实施方式1-3中任一项所述的方法,其中所述gdh具有以下特征:(1)如实施方式1的(a)至(c)中任一项所定义的活力,(2)底物特异性:相对于对d-葡萄糖的反应性(100%),对麦芽糖的反应性为2%或以下;(3)最适活力ph:6.5至7.5;和(4)当通过sds-page测量时,约65至约81kda的分子量。实施方式9.根据实施方式1-5和8中任一项所述的方法,其中所述gdh来自属于以下各项的微生物:毛霉属(mucor)、犁头霉属(absidia)、放射毛霉属(actinotnucor)、卷霉属(circinella)、parasitella属或根霉属(rhizopus)。实施方式10.根据实施方式9所述的方法,其中所述gdh是来自以下各项的gdh:普雷恩毛霉(mucorprainii)、卷枝毛霉(mucorcircinelloides)、卷枝毛霉f.cirinelloide变型(mucorcircinelioidesf.cirinelloides)、不明毛霉(mucorambiguus)、冻土毛霉(mucorhiemalis)、冻土毛霉林生变型(mucorhiemalisf.silvaticus)、细孢毛霉(mucorsubtilissimus)、季也蒙毛霉(mucorguilliermondii)、爪哇毛霉(mucorjavanicus)、两型孢毛霉(mucordimorphosporus)、毛霉属(mucor)rd056860、细孢毛霉(mucorsubtilissimus)、柱孢犁头霉(absidiacylindrospora)、透孢犁头霉(absidiahyalospora)、雅致放射毛霉(actinomucorelegans)、小卷霉(circinellaminor)、毛霉状卷霉(circinellamucoroides)、蝇卷霉(circinellamuscae)、硬卷霉(circinellarigida)、单卷霉(circinellasimplex)、伞形卷霉(circinellaumbellata)、parasitellaparasitica或者小孢根霉(rhizopusmicrosporus)。实施方式11.根据实施方式6或7所述的装置,其中所述gdh具有以下特性:(1)如实施方式6的(a)至(c)中任一项所定义的活力,(2)底物特异性:相对于对d-葡萄糖的反应性(100%),对麦芽糖的反应性为2%或以下;(3)最适活力ph:6.5至7.5;和(4)当通过sds-page测量时,约65至约81kda的分子量。实施方式12.根据实施方式6、7或11所述的装置,其中所述gdh来自属于毛霉属(mucor)、犁头霉属(absidia)、放射毛霉属(actinotnucor)、卷霉属(circinella)、parasitella属或根霉属(rhizopus)的微生物。实施方式13.根据实施方式12所述的装置,其中所述gdh是来自普雷恩毛霉(mucorprainii)、卷枝毛霉(mucorcircinelloides)、卷枝毛霉f.cirinelloide变型(mucorcircinelioidesf.cirinelloides)、不明毛霉(mucorambiguus)、冻土毛霉(mucorhiemalis)、冻土毛霉林生变型(mucorhiemalisf.silvaticus)、细孢毛霉(mucorsubtilissimus)、季也蒙毛霉(mucorguilliermondii)、爪哇毛霉(mucorjavanicus)、两型孢毛霉(mucordimorphosporus)、毛霉属(mucor)rd056860、细孢毛霉(mucorsubtilissimus)、柱孢犁头霉(absidiacylindrospora)、透孢犁头霉(absidiahyalospora)、雅致放射毛霉(actinomucorelegans)、小卷霉(circinellaminor)、毛霉状卷霉(circinellamucoroides)、蝇卷霉(circinellamuscae)、硬卷霉(circinellarigida)、单卷霉(circinellasimplex)、伞形卷霉(circinellaumbellata)、parasitellaparasitica或者小孢根霉(rhizopusmicrosporus)的gdh。发明的有利作用本发明相对于使用god的常规装置的优势在于由于其长期稳定性,在传感器的寿命期间,可以通过使用但较不经常使用或不通过使用手指针刺校准来使用gdh。基于god的大部分系统在传感器寿命期间需要每特定时间段校准1次,如每12小时1次。对此的原因之一在于在作为使用cgm装置的温度的环境温度(30-40℃)下培育期间,god活力快速降低。作为对比,本发明人意外地发现在环境温度(30-40℃)下保持较长的一段时间,如1至3周,gdh保持了更高的稳定性(酶活力),并且可以在无需再校准或者虽然再校准但不太频繁的情况下使用。附图说明[图1]图1显示了在57℃热处理15分钟之后,不同类型的gdh和god的残余酶活力。[图2]图2显示了在40℃保持长达21天之后,不同类型的gdh和god的残余酶活力。[图3]图3显示了在40℃保持长达7天之后,不同类型的gdh和god的残余酶活力。[图4]图4显示了在40℃保持长达14天之后,不同类型的gdh的残余酶活力。[图5]图5显示了使用pei和pegde的gdh-m2或god-3固定化的电极在0至4小时的残余酶活力。[图6]图6显示了使用pa和pegde的gdh-m2或god-3固定化的电极在0至4小时的残余酶活力。[图7]图7显示了gdh-m2的最适ph。[图8]图8显示了gld3的最适ph。[图9]图9显示了使用gdh-m2电极的循环伏安法。[图10]图10显示了响应电流的葡萄糖浓度依赖性。记录了0.4v的氧化电流值。[图11]图11显示了gdh-m2和gld3的sds-page。具体实施方式1-1.葡糖脱氢酶活力和储存稳定性在一个实施方式中,本发明提供了包括使用gdh的连续葡萄糖监测(cgm)方法。在所述方法中,可以使用具有以下能力的gdh(a)当保持在约37-40℃时,在1周内保持初始活力的约60%或以上,例如,约65%或以上,约70%或以上,约75%或以上,约80%或以上,例如,约60-100%,约60-99%,约70-100%,75-100%,或者80-100%,(b)当保持在约37-40℃时,在2周内保持初始活力的约40%或以上,例如,约45%或以上,约50%或以上,约55%或以上,约60%或以上,约65%或以上,例如,约40-100%,约40-99%,约45-100%,约50-100%,约55-100%,约60-100%,或者约65-100%,或者(c)当保持在约37-40℃时,在3周内保持初始活力的约30%或以上,例如,约35%或以上,约40%或以上,约45%或以上,约50%或以上,约55%或以上,约60%或以上,例如,约30-100%,约30-99%,约40-100%,45-100%,50-100%,55-100%,或者约60-100%。在本文中,可以将如上所描述的保持gdh活力称为具有(包括)“储存稳定性”。然而,在本文中术语“储存”仅出于方便使用,并且不应以任何限制方式来看待或理解该术语。也就是说,短语“储存稳定性”仅意欲解释gdh的特性,即gdh在特定温度,在特定时间段内保持活力,并且不表示gdh应物理储存或放置。具有如上所描述的储存稳定性的gdh可以在电极、葡萄糖传感器或者cgm装置中使用或者在包括连续葡萄糖监测的方法中使用。在一个实施方式中,所述gdh可以是fad依赖性gdh。在一个实施方式中,所述gdh可以是可溶型gdh或者来源于可溶型gdh。在本文中,短语“可溶型gdh”是指在溶液中可溶并且不是膜结合蛋白的gdh(如wo2002/036779、us2004/0023330中所述的洋葱伯克霍尔德菌(b.cepacia)gdh)。然而,应指出在本文中,短语“可溶型gdh”仅描述gdh的“类型”(或者gdh的来源)并且不表示gdh必须物理地处于溶液中。所述可溶型gdh可以以干燥状态储存或者固定至电极上。在本文中,仍可以将这种固定化gdh类型称为可溶型。在另一个实施方式中,所述gdh不是膜结合蛋白并且不包含细胞色素域。在另一个实施方式中,所述gdh不是由能够形成非共价复合物的三个亚基组成的fad-gdh。来自洋葱伯克霍尔德菌(b.cepacia)的fad-gdh(wo2002/036779,us2004/0023330)由能够形成非共价复合物的三个亚基组成,并且这种洋葱伯克霍尔德菌(b.cepacia)gdh不包括在本发明方法与装置中所使用的gdh内。在另一个实施方式中,本发明所述的gdh由一个亚基组成,尽管这不排除共价融合至一个或多个细胞色素域。在另一个实施方式中,所述gdh不是pqq依赖性gdh。在另一个实施方式中,所述gdh不是nad依赖性gdh。在一个实施方式中,所述gdh是单体。在一个实施方式中,所述gdh不是二聚体。在一个实施方式中,可以在ph约7.0至7.5,例如,约ph7.4评价gdh的储存稳定性。所述gdh可以如以上(a)至(c)中所述,在约37-40℃的温度下,例如,在约37℃,约38℃,约39℃或者约40℃保持活力。优选地,可以使用在严苛的温度条件下相对稳定的gdh。在一个实施方式中,可以使用在57℃,ph7.0热处理15分钟之后保持其初始gdh活力的约20%或以上,例如,20-100%的gdh。在一个实施方式中,所述gdh可以来自属于毛霉属(mucor)、犁头霉属(absidia)、放射毛霉属(actinotnucor)和卷霉属(circinella)的微生物。在另一个实施方式中,所述gdh可以是来自普雷恩毛霉(mucorprainii)、卷枝毛霉(mucorcircinelloides)、卷枝毛霉f.cirinelloide变型(mucorcircinelioidesf.cirinelloides)、不明毛霉(mucorambiguus)、冻土毛霉(mucorhiemalis)、冻土毛霉林生变型(mucorhiemalisf.silvaticus)、细孢毛霉(mucorsubtilissimus)、季也蒙毛霉(mucorguilliermondii)、爪哇毛霉(mucorjavanicus)、两型孢毛霉(mucordimorphosporus)、毛霉属(mucor)rd056860、细孢毛霉(mucorsubtilissimus)、柱孢犁头霉(absidiacylindrospora)、透孢犁头霉(absidiahyalospora)、雅致放射毛霉(actinomucorelegans)、小卷霉(circinellaminor)、毛霉状卷霉(circinellamucoroides)、蝇卷霉(circinellamuscae)、硬卷霉(circinellarigida)、单卷霉(circinellasimplex)、伞形卷霉(circinellaumbellata)、parasitellaparasitica或者小孢根霉(rhizopusmicrosporus)的gdh。在另一个实施方式中,所述gdh可以包含改变其特性的突变。在一个实施方式中,与当前在常规cgm系统中使用的已知的god相比,在本发明的cgm方法中使用的gdh可以在较长的一段时间,如1至3周内保持较高的酶活力。在本文中,gdh或god酶的残余活力比的定义如下所示:残余活力比=(在约37-40℃保持3周后的残余活力a)/(初始活力b)×100应在相同的测定条件下测量相同酶的残余活力和初始活力。在一个实施方式中,可以在适合于cgm的ph,如ph6.9至7.4,例如,ph7.0下测量残余活力和初始活力。cgm方法的ph可以调节至约6.9至7.4,例如,约7.0。这是因为血液的ph为约7.0。可以使用磷酸盐缓冲液,如pbs或者其他常规缓冲液来调节ph。在一个实施方式中,在本发明中使用的gdh的残余活力比优选地高于在cgm系统中使用的已知的god酶。这种已知的god酶可以是来自属于曲霉属(aspergillussp.)的微生物,如黑曲霉(aspergillusniger)或曲霉属(aspergillussp.)的god。这些已知的god酶可以选自toyobogod(产品目录no.glo-201,来自曲霉属(aspergillussp.))、wakopurechemicalsgod(产品目录no.074-02401,来自黑曲霉(aspergillusniger))、sigma-aldrichgodvii型(来自黑曲霉(aspergillusniger)的god)和sigma-aldrichgodx-s型(来自黑曲霉(aspergillusniger)的god)。在一个实施方式中,可以在使用或不使用再校准的情况下进行连续葡萄糖监测。在一个实施方式中,可以在不经常地,例如,每1、2、3、4、5、6、7、8、9、10、11、12、13或14天再校准一次的情况下进行连续葡萄糖监测。在另一个实施方式中,本发明提供了筛选能够在连续葡萄糖监测中使用的gdh的方法。该方法包括以下步骤:(i)制备候选gdh,(ii)将所述候选gdh在约30-40℃,例如,约37-40℃保持预定时间段,(iii)在步骤(ii)之后确定所述gdh的残余活力,和(iv)将步骤(iii)中所确定的残余活力与所述候选gdh的初始活力相比较。当所述残余gdh活力(a)当在约30-40℃,例如,约37-40℃保持1周时,与初始gdh活力相比,为约60%或以上,例如,约65%或以上,约70%或以上,约75%或以上,约80%或以上,例如,约60-100%,约60-99%,约70-100%,75-100%,或者80-100%,(b)当在约30-40℃,例如,约37-40℃保持2周时,与初始gdh活力相比,为约40%或以上,例如,约45%或以上,约50%或以上,约55%或以上,约60%或以上,约65%或以上,例如,约40-100%,约40-99%,约45-100%,约50-100%,约55-100%,约60-100%,或者约65-100%,或者(c)当在约30-40℃,例如,约37-40℃保持3周时,与初始gdh活力相比,为约30%或以上,例如,约35%或以上,约40%或以上,约45%或以上,约50%或以上,约55%或以上,约60%或以上,例如,约30-100%,约30-99%,约40-100%,45-100%,50-100%,55-100%,或者约60-100%,则所述候选gdh可以选作适合于连续葡萄糖监测的gdh或者选作具有在cgm中使用,例如,在cgm中使用1至3周的潜力的gdh。在以上步骤(ii)中,当进行筛选时,所述候选gdh可以在约30-40℃,例如,约37-40℃保持一段时间,例如,0.5、1、2、3、4、5、6或12小时、1、2、3、4、5、6或7天,或者1、2或3周。通常在约30-40℃的温度下进行cgm,这部分是因为平均体温为约37℃。因此,gdh的筛选可以在约30-40℃,例如,约37-40℃,约30℃,约31℃,约32℃,约33℃,约34℃,约35℃,约36℃,约37℃,约38℃,约39℃或者约40℃进行。所述候选gdh可以是来自属于毛霉属(mucor)的微生物的gdh,或其衍生物、变体或突变体。所述候选gdh可以是对麦芽糖、d-半乳糖和/或d-木糖具有低特异性的gdh。所述候选gdh可以是热稳定的gdh(热稳定gdh),尽管在加速试验条件下,它不需要比在cgm装置中使用的god的热稳定性更好。在一个实施方式中,在57℃,ph7.0热处理15分钟之后,步骤(i)所述的候选gdh优选地保持了其初始gdh活力的约20%或以上,例如,约20-100%。为了筛选具有在cgm中使用的潜力的gdh,根据常规知识,对候选gdh酶实施加速热处理步骤来替代实施对于cgm装置的整个寿命(如1至3周)的完整稳定性测试可以是可行的。然而,本发明人意外地发现在相似条件下,与可商购的god相比,多种gdh在环境温度下,对于较长的一段时间保持了更高的稳定性。鉴于以下事实:在使用57℃下的热处理的加速试验中,已知的god保持了高于gdh的活力,这是特别意外的。因此,在本发明的一个实施方式中,可以优选地不用包括热处理步骤,如在约50-60℃的温度下热处理约5-30分钟的加速试验替代以上步骤(ii)。在一个实施方式中,本发明提供了连续葡萄糖监测装置,其包括具有以下能力的gdh(a)当保持在约37-40℃时,在1周内保持初始活力的60%或以上,例如,约65%或以上,约70%或以上,约75%或以上,约80%或以上,例如,约60-100%,约60-99%,约70-100%,75-100%,或者80-100%,(b)当保持在约37-40℃时,在2周内保持初始活力的40%或以上,例如,约45%或以上,约50%或以上,约55%或以上,约60%或以上,约65%或以上,例如,约40-100%,约40-99%,约45-100%,约50-100%,约55-100%,约60-100%,或者约65-100%,或者(c)当保持在约37-40℃时,在3周内保持初始活力的30%或以上,例如,约35%或以上,约40%或以上,约45%或以上,约50%或以上,约55%或以上,约60%或以上,例如,约30-100%,约30-99%,约40-100%,45-100%,50-100%,55-100%,或者约60-100%,其中所述gdh是fad依赖性gdh并且其中所述gdh不是膜结合蛋白。在一个实施方式中,所述gdh不是pqq依赖性gdh。就对于特定时间段的葡萄糖监测来说,连续葡萄糖监测(cgm)还可以称为快速葡萄糖监测(fgm)。照此,除非明确另外表示,否则对于本发明的实施方式,如本文所使用的短语“连续葡萄糖监测”涵盖了快速葡萄糖监测的实施方式。在一个实施方式中,装置包括在57℃,ph7.0热处理15分钟之后,优选地保持了其初始gdh活力的约20%或以上,例如,约20-100%的gdh。在一个实施方式中,cgm装置可以包括放置在皮肤下的葡萄糖传感器。传感器可以佩戴特定时间段,如1天至约3周。传感器部分可以是一次性的。传感器可以包含传感器膜层以防止组织与酶的直接接触。传感器可以设计以通过使用施用器插入腹部。在一个实施方式中,本发明提供了cgm装置,其还包括发射器以及从所述传感器至发射器的连接。发射器不必需植入。优选地,发射器能够与无线电接收器通讯。cgm装置还可以包含可以连续显示葡萄糖水平的电子接收器。cgm装置可以可选地包含用于校准的手指针刺(手指点刺)。cgm装置可以可选地包含使用手册。在一个实施方式中,cgm装置可以是自供电的(spgs)并且可以省去参比电极。fad-gdh催化其中在存在电子受体的情况下使葡萄糖的羟基氧化以形成葡萄糖酸-δ-内酯的反应。该反应可以用于测量fad-gdh的活力,例如,通过使用以吩嗪硫酸甲酯(pms)和2,6-二氯靛酚(dcip)作为电子受体的以下系统。d-葡萄糖+pms(氧化形式)→d-葡萄糖酸-δ-内酯+pms(还原形式)(通过fad-gdh催化)(反应1)pms(还原形式)+dcip(氧化形式)→pms(氧化形式)+dcip(还原形式)(反应2)在反应1中,葡萄糖氧化并且产生了还原形式的pms。在后续反应2中,还原形式的pms氧化,并且dcip还原。通过600nm波长下吸光值的变化,检测氧化形式的dcip的减少程度,可以确定gdh酶的活力。可以如下所示测量fad-gdh的活力。将2.05ml100mm的磷酸盐缓冲液(ph7.0)、0.6ml或1md-葡萄糖溶液和0.15ml2mmdcip溶液混合,然后在37℃培育5分钟。然后,加入0.1ml15mmpms溶液和0.1ml的酶样品溶液以起始反应。在初始条件并随时间测量吸光值,确定每分钟,600nm吸光值的减少(δa600),并因此可以计算gdh的活力。1u的gdh活力可以定义为在37℃,在存在浓度为200mm的d-葡萄糖的情况下,在1分钟内减少1μmoldcip的酶量。方程中数值3.0表示反应试剂和酶试剂的量(ml),16.3代表毫摩尔分子消光系数(cm2/μmol),0.1代表酶溶液的量(ml),1.0代表比色皿长度(cm),δ600空白代表600nm下每分钟吸光度减少的量,其中仅加入缓冲液来起始反应,而不是添加酶溶液,并且df代表稀释因子。在本发明方法中使用的gdh可以具有如上所描述的葡糖脱氢酶活力和储存稳定性。还可以使用其等价形式。1-2.底物特异性在本发明方法中使用的gdh可以具有高底物特异性。在一个实施方式中,与对葡萄糖(d-葡萄糖)的反应性相比,所述gdh可以至少对麦芽糖具有低反应性。在另一个实施方式中,与对葡萄糖(d-葡萄糖)的反应性相比,所述gdh可以至少对麦芽糖和木糖(d-木糖)具有低反应性。在一个实施方式中,相对于对相同浓度的葡萄糖的反应性(100%),在本发明方法中使用的gdh对麦芽糖的反应性为(例如)2%或以下,优选地1%或以下,更优选地0.7%或以下,更优选地0.4%或以下。在一个实施方式中,相对于对相同浓度的葡萄糖的反应性(100%),在本发明方法中使用的gdh对木糖的反应性可以为(例如)3%或以下,优选地2%或以下,更优选地1%或以下,更优选地0.6%或以下。在另一个实施方式中,与对葡萄糖(d-葡萄糖)的反应性相比,所述gdh可以至少对半乳糖具有低反应性。在一个实施方式中,相对于对相同浓度的葡萄糖的反应性(100%),在本发明方法中使用的gdh对半乳糖的反应性可以为(例如)5%或以下,4%或以下,3%或以下,2%或以下或者1%或以下。可以通过在本文所述的测量葡糖脱氢酶活力的方法中用另一种糖如木糖或麦芽糖替代葡萄糖,并将所测量的活力与通过葡萄糖所获得的活力相比较来测量在本发明方法中使用的gdh对每种糖的反应性。例如,可以将每种糖的浓度设置为200mm。此外,当测量对葡萄糖的反应性时,反应液体的最终酶浓度可以设置为1.4μg/ml,并且当测量对木糖或麦芽糖的反应性时,反应液体的最终酶浓度可以设置为28μg/ml。在本发明方法中使用的具有这种高底物特异性的gdh可以用作准确测量样品葡萄糖浓度的酶,例如,即使当所述样品含有杂质,如麦芽糖或木糖时。1-3.最适ph在本发明方法中使用的gdh可以具有宽泛的最适ph活力范围。由于cgm是对血液和间质液实施的,因此优选地,在所述方法中使用的gdh酶具有中性ph(ph7.0)周围的最适ph。在一个实施方式中,在本发明方法中使用的gdh可以(例如)在ph5.0至7.5,优选地5.5至7.5,更优选地6.0至7.5,更优选地6.5至7.5具有最适ph活力。在另一个实施方式中,在本发明方法中使用的gdh可以在ph5.0至8,优选地5.5至7.5,更优选地6.0至7.5,更优选地6.5至7.5具有最适ph活力。在实施例中显示了这些gdh。在一个实施方式中,最适ph可以认为是相对于最高活力(100%),70%或以上的相对活力。在另一个实施方式中,最适ph可以认为是相对于最高活力(100%),75%或以上,例如,80%或以上的相对活力。在一个实施方式中,在本发明方法中使用的gdh具有最适ph6.0至7.5,其中最适活力是相对于最高活力(100%)的70%或以上。在初始条件并随时间测量吸光值,确定每分钟,520nm吸光值的减少(δa520),并因此可以计算gdh的活力。6.8代表毫摩尔分子消光系数(cm2/μmol)。已知基于ph,在600nm下,dcip的摩尔吸光系数在很大程度上是不同的。已知在520nm下,dcip的摩尔吸光系数相当恒定,与ph无关。通过对此进行考虑,可以在520nm进行测量。1-4.最适活力温度在本发明方法中使用的gdh可以在宽泛的范围,例如,30至50℃,30至55℃,35至50℃,35至55℃,35至60℃或者30至60℃具有最适活力温度。在本文中,30至60℃的最适活力温度通常是指所述温度大体上处于约30至60℃的范围内,并且所述范围还包括在某种程度上可接受的留量(allowance),同时所述活力是最适的。在本说明书中,可以通过在反应液体的最终酶浓度(1.4μg/ml)下,测量磷酸钾缓冲溶液(ph7.0)中的酶活力来计算最适活力温度。1-5.ph稳定性在本发明方法中使用的gdh可以在宽泛的ph范围内具有高ph稳定性。在一个实施方式中,在本发明方法中使用的gdh至少在5.0至7.5的整个ph范围内是稳定的。在本说明书中,当与相同的酶在处理前的初始酶活力(100%)相比,已在特定ph条件下,在25℃处理16小时的100u/ml的酶具有75%或以上的残余酶活力,则酶被认为在该ph条件下是稳定的。1-6.温度稳定性在本发明方法中使用的gdh可以具有温度稳定性。在一个实施方式中,在本发明方法中使用的gdh(例如)至少在50℃或以下(即在0至50℃的温度范围内),51℃或以下,52℃或以下,53℃或以下,54℃或以下,55℃或以下,56℃或以下或者57℃或以下稳定15分钟。在本说明书中,当与相同的酶在热处理前的初始酶活力(100%)相比,已在特定温度条件下,在适当缓冲溶液(例如,乙酸钾缓冲液(ph5.0))中处理15分钟的10u/ml的酶具有20%或以上的残余酶活力,则酶被认为在该温度条件下是稳定的。在本发明方法中使用的gdh可以具有至少1种或以上,2种或以上,3种或以上,4种或以上,5种或以上或者全部6种如上所描述的酶特性。在本发明方法中使用的gdh可以具有如上所描述的特性的任意组合。在一个实施方式中,在本发明方法中使用的gdh可以具有部分1-1中所述的特性,并且还具有选自以上部分1-2、1-3、1-4、1-5和1-6中所述的特性的至少一种特性。1-7.分子量在一个实施方式中,如通过sds-page所测量的,构成在本发明方法中使用的gdh的多肽部分可以具有约65至81kda,约65至80kda,约65至75kda,约65至70kda或者约70kda的分子量。短语“如通过sds-page所测量的约70kda”包括其中技术人员通常将确定当通过sds-page测量所述多肽的分子量时,条带出现在70kda的位置的范围。技术人员将理解根据条件不同,在sds-page上出现的条带可以比预期值高或低5kd。“多肽部分”是指gdh的多肽部分并且不包括所附接的糖链。在一个实施方式中,如根据氨基酸序列所推断的,构成在本发明方法中使用的gdh的多肽部分可以具有约65至80kda,约65至75kda,约65至70kda,约69至70kda或者约70kda的理论分子量。短语“如根据氨基酸序列所推断的约70kda的理论分子量”是指根据氨基酸序列计算的gdh的分子量。这不包括所附接的糖链。当使用某些微生物产生在本发明方法中使用的gdh时,所述gdh可能以糖基化形式表达。在一个实施方式中,对于gdh的糖基化形式,可以实施热处理或去糖基化处理以除去糖链以提供多肽部分。可以使用糖水解酶,如endoh内切糖苷酶进行去糖基化。当使用某些其他微生物产生在本发明方法中使用的gdh时,gdh可以以非糖基化形式表达。对于这种形式,所表达的gdh是多肽部分。已知用于从gdh的糖基化形式除去糖链的多种方法。一个实例可以是通过在100℃热处理10分钟使糖基化的gdh变性,然后用内切糖苷酶endoh,在37℃处理14小时。一个实例可以是通过用内切糖苷酶endoh在37℃处理24小时使糖基化的gdh变性。当在本发明方法中使用的gdh具有所附接的糖链时,对其分子量没有限制,只要对葡糖脱氢酶活力、底物特异性、比活等无副作用。例如,当在本发明方法中使用的gdh具有所附接的糖链时,如通过sds-page所测量的,其分子量可以是80至120kda。在本发明方法中使用的gdh可以或可以不是糖基化形式的。可以通过使用可商购的分子量标志物,使用常规方法和装置来实施通过sds-page的分子量测量。在一个实施方式中,在本发明方法中使用的gdh可以具有部分1-1中所描述的特性,并且还具有选自以上部分1-2至1-7中所描述的特性中的至少一种特性。1-8.来源未具体限制在本发明方法中使用的gdh的来源,只要gdh具有如上所描述的特性。在本发明方法中使用的gdh可以来源于(例如)属于毛霉亚门(mueoromyeotina)的成员、毛霉纲(mucoromycetes)的成员、毛霉目(mucorales)的成员、毛霉科(mucoraceae)的成员的微生物。具体实例包括毛霉属(mucor)种、犁头霉属(absidia)种、放射毛霉属(actinotnucor)种和卷霉属(circinella)种。毛霉属(mucor)种的实例包括普雷恩毛霉(mucorprainii)、爪哇毛霉(mucorjavanicus)、卷枝毛霉f.cirinelloide变型(mucorcircinelioidesf.cirinelloides)、不明毛霉(mucorambiguus)、季也蒙毛霉(mucorguilliermondii)、冻土毛霉林生变型(mucorhiemalisf.silvaticus)、细孢毛霉(mucorsubtilissimus)和两型孢毛霉(mucordimorphosporous)。犁头霉属(absidia)种的实例包括柱孢犁头霉(absidiacylindrospora)和透孢犁头霉(absidiahyalospora)。放射毛霉(actinomucor)种的实例包括雅致放射毛霉(actinomucorelegans)。卷霉属(circinella)种的实例包括小卷霉(circinellaminor)、毛霉状卷霉(circinellamucoroides)、蝇卷霉(circinellamuscae)、硬卷霉(circinellarigida)、单卷霉(circinellasimplex)和伞形卷霉(circinellaumbellata)。其他实例包括wo2010/140431、us2011/0318810中所描述的那些。其他实例包括parasitellaparasitica和小孢根霉(rhizopusmicrosporus)。其他来源可以是节菱孢属(arthrinium)和梨孢假壳属(apiospora)。节菱孢属(arthrinium)的实例包括但不限于:日本节菱孢(arthriniumjaponicum)、暗孢节菱孢(arthriniumphaeospermum)、arthriniumterminalis、蔗生节菱孢(arthriniumsaccharicola)、甘蔗节菱孢(arthriniumsacchari)、arthriniumserenense、芦竹节菱孢(arthriniumarundinis)、arthriniumeuphaubie。梨孢假壳属(apiospora)的实例包括但不限于:蒙塔涅梨孢假壳菌(apiosporamontagnei)、apiosporasetosa和apiosporatintinnabula以及甘蔗节菱孢(arthriniumsacchari)。对于节菱孢属(arthrinium)和梨孢假壳属(apiospora)的微生物,参见美国专利no.9,796,963,其以其全部内容并入本文。其他来源包括但不限于绿僵菌属(metarhizium)的成员,例如,绿僵菌属(metarhizium)种,例如,绿僵菌(metarhizium)种f2114(hyphagenesisinc.)(参见jp专利申请公开no.2016-116488,其以其全部内容并入本文);曲霉属(aspergillus)的成员,例如,曲霉(aspergillus)种rd009469(参见wo2015/060150,其以其全部内容并入本文)、aspergillusiizukae,例如,aspergillusiizukaeno.5453株(参见wo2017/077924,其以其全部内容并入本文)、杂色曲霉(aspergillusversicolor),例如,杂色曲霉(aspergillusversicolor)no.52439株(参见jp专利申请公开no.2017-221147,其以其全部内容并入本文);白曲霉(aspergilluskawachii)、泡盛曲霉(aspergillusawamori),例如,泡盛曲霉(aspergillusawamori)no.1731株(参见jp专利申请公开no.2017-112860,其以其全部内容并入本文);臭曲霉(aspergillusfoetidus),例如,臭曲霉(aspergillusfoetidus)nbrc4312株(nbrc)、黑曲霉(aspergillusniger)cbs513.88株(centraalbureauvoorschimmelcultures:cbs)、泡盛曲霉(aspergillusawamori)no.1751株、金黄曲霉(aspergillusaureus),例如,金黄曲霉(aspergillusaureus)no.2062株(参见jp专利申请公开no.2017-112858,其以其全部内容并入本文)、双孢曲霉(aspergillusbisporus),例如,双孢曲霉(aspergillusbisporus)nbrc32017株(nbrc);金黄色嗜热子囊菌(thermoascusaurantiacus),例如,金黄色嗜热子囊菌(thermoascusaurantiacus)6766株、金黄色嗜热子囊菌(thermoascusaurantiacus)9748株(参见jp专利申请公开no.2015-167506,其以其全部内容并入本文);和埃默森篮状菌(talaromycesemersonii),例如,埃默森篮状菌(talaromycesemersonii)31232株和9734株、thermoascuscrustaceus,例如,thermoascuscrustaceus9129株和9816株以及wo2014/045912的表1中所公开的其他微生物(参见wo2014/045912,其以其全部内容并入本文)以及botryothiniafuckeliana(参见wo2012/001976,其以其全部内容并入本文)。在一个实施方式中,可以(例如)通过初始获得已知的gdh基因,如专利文献1(wo2015/099112、us20160319246)、专利文献8(wo2012/169512、us20140287445)或者专利文献9(wo2015/129475、ep3112461)中所描述的gdh基因来获得fad-gdh。所获得的基因可以或可以不是突变的。突变可以是任何突变,如这些文档中所描述的那些或相应突变。在一个实施方式中,可以通过向其中引入突变n66y/n68g/c88a/q233r/t387c/e554d/l557v/s559k来修饰来自普雷恩毛霉(mucorprainii)的gdh(mpgdh,seqidno:1)。还可以应用其他突变,如文献中所描述的突变。在本文中,可以用缩写“axxxb”表示氨基酸替代(其中a是替代之前的氨基酸,xxx是要替代的具体氨基酸序列内的位置,并且b是替代后的氨基酸)。例如,对于seqidno:1,a175c表示其中seqidno:1的位置175处的丙氨酸替代为半胱氨酸的替代。在另一个实施方式中,可以在cgm中使用表示为gld3(funakoshi)的gdh或其等价形式。在另一个实施方式中,可以在cgm中使用来源于冻土毛霉(mucorhiemalis)gdh的gdh。在另一个实施方式中,可以在cgm中使用表示为mpgdh-m2的gdh(seqidno:11)。引入靶标氨基酸替代的方法的实例包括其中随机引入突变的方法和其中在指定位置引入定点突变的方法。随机突变法的实例包括易错pcr法(techniques,1,11-15(1989))和使用xl1-red感受态细胞的方法,其中在质粒复制期间经常发生错误并且其对细胞增殖期间的修饰发生敏感(stratagenecorp.)。定点突变法的实例包括包含选择可以赋予靶标作用的氨基酸和使用常规试剂盒,如可商购的快速变换定点突变试剂盒(stratagenecorp.)引入定点突变的方法。为了获得fad-gdh基因,可以使用常规方法,如currentprotocolsinmolecularbiology,wileyinterscience(1989)中所描述的那些,从已知的微生物细胞或者具有产生fad-gdh的能力的多种其他细胞提取染色体dna或mrna。此外,可以通过使用mrna作为模板合成cdna。然后,可以使用如此所获得的染色体dna或cdna制备染色体dna或cdna文库。随后,可以扩增含有编码具有高底物特异性的fad-gdh的靶标基因片段的dna,可以连接所述dna片段,并且可以获得含有整个fad-gdh基因的dna。可以通过基于已知fad-gdh的氨基酸序列信息合成适合的探针dna,使用探针dna从染色体dna或cdna文库选择具有高底物特异性的fad-gdh基因,或者通过基于氨基酸序列制备适合的引物dna,使用聚合酶链反应(pcr)法,如5'race或者3'race并且连接dna片段来进行dna片段的连接。编码fad-gdh的基因可以用作起始材料并且可以通过选择表达优选的fad-gdh的克隆来进行突变体筛选。可以通过将fad-gdh基因或者包含所述基因的细胞暴露于诱变剂或者紫外辐射,或者通过实施引起突变的基因工程方法来产生突变体。诱变剂的实例包括n-甲基-n'-硝基-n-亚硝基胍、羟胺、亚硝酸、亚硫酸、肼、甲酸和5-溴尿嘧啶等。就通过紫外线辐照的情况来说,可以使用如chemistrytoday,24-30,jun.1989中所描述的常规方法进行辐照。定点突变的实例包括kunkel法(proc.natl.acad.sci.u.s.a.,82,488(1985);methodsenzymol.,154,367(1987))、eckstein法(nucleicacidsres.,13,8749(1985);nucleicacidsres.,13,8765(1985);nucleicacidsres.,14,9679(1986))和kramer法(nucleicacidsres.,12,9441(1984);methodsenzymol.,154,350(1987);gene,37,73(1985))。可商购的试剂盒(如transformer突变试剂盒(clontechlaboratories,inc.)、exoiii/绿豆缺失试剂盒(stratagenecorp.)或者快速变换定点突变试剂盒(stratagenecorp.))可以用于转化dna序列。此外,可以通过使用dna合成法获得fad-gdh基因。基因可以经设计以包含天然存在的gdh的基因序列或者可以具有突变。可以使用常规测序技术对gdh基因的dna序列测序,并且可以使用可商购系统,如多毛细管dna分析系统ceq2000(beckmancoulterinc.)。可以通过获得天然存在的fad-gdh或者通过修饰已知的fad-gdh获得fad-gdh。包含fad-gdh的微生物的实例包括毛霉亚门(mueoromyeotina)的成员、毛霉纲(mucoromycetes)的成员、毛霉目(mucorales)的成员、毛霉科(mucoraceae)的成员。具体实例包括毛霉属(mucor)种、犁头霉属(absidia)种、放射毛霉属(actinotnucor)种和卷霉属(circinella)种。毛霉属(mucor)种的实例包括普雷恩毛霉(mucorprainii)、爪哇毛霉(mucorjavanicus)、卷枝毛霉f.cirinelloide变型(mucorcircinelioidesf.cirinelloides)、不明毛霉(mucorambiguus)、季也蒙毛霉(mucorguilliermondii)、冻土毛霉林生变型(mucorhiemalisf.silvaticus)、细孢毛霉(mucorsubtilissimus)和两型孢毛霉(mucordimorphosporous)。犁头霉属(absidia)种的实例包括柱孢犁头霉(absidiacylindrospora)和透孢犁头霉(absidiahyalospora)。放射毛霉(actinomucor)种的实例包括雅致放射毛霉(actinomucorelegans)。卷霉属(circinella)种的实例包括小卷霉(circinellaminor)、毛霉状卷霉(circinellamucoroides)、蝇卷霉(circinellamuscae)、硬卷霉(circinellarigida)、单卷霉(circinellasimplex)和伞形卷霉(circinellaumbellata)。其他实例包括wo2010/140431、us2011/0318810中所描述的那些。其他实例包括parasitellaparasitica和小孢根霉(rhizopusmicrosporus)。其他实例包括如上所描述的那些。可以将fad-gdh基因引入载体,如噬菌体、粘粒或质粒,并且使用常规方法,这些可以用于转化原核细胞或真核细胞。原核宿主细胞的实例包括属于埃希氏菌属(escherichia)的微生物,如大肠杆菌(escherichiacoli)k-12株、大肠杆菌bl21(de3)、大肠杆菌dh5α、大肠杆菌w3110或者大肠杆菌c600(takarabioinc.)。可以使用cacl2法或者电穿孔法转化或转导细胞。可以使用可商购的感受态细胞(如ecos感受态大肠杆菌bl21(de3),nippongeneco.,ltd.)。真核宿主细胞的实例包括酵母,如接合酵母属(zygosaccharomyces)、酵母菌属(saccharomyces)、毕赤氏酵母属(pichia)和假丝酵母属(candida)的那些。标志物基因可以用于鉴别转化的细胞。标志物基因的实例包括补充宿主营养需求的基因,如ura3或trp1。此外,所插入的基因可以包含启动子或其他控制序列(如增强子序列、终止子序列或多腺苷酸化序列)。启动子的实例包括gal1启动子和adh1启动子。可以使用常规方法,如(无限制地)醋酸锂法(methodsmol.cell.biol.,5,255-269(1995))或电穿孔法(j.microbiol.methods,55,481-484(2003))转化酵母。作为另外一种选择,可以使用如原生质球状体法或玻璃珠法的方法。真核宿主细胞的其他实例包括霉菌,如曲霉属(aspergillus)种和木霉属(tricoderma)种的那些。所插入的基因可以包含启动子,如tef1启动子,及其他控制序列(如分泌信号序列、增强子序列、终止子序列或多腺苷酸化序列)。此外,所插入的基因还可以包含标志物基因,如niad或pyrg以选择转化的细胞。此外,所插入的基因可以包含用于插入到染色体中的同源重组域。在原生质体形成后,可以使用已知的方法,如unkles等人(mol.gen.genet.,218,99-104(1989))所述的方法,使用聚乙二醇和氯化钙来转化霉菌。可以通过培养能够产生fad-gdh的宿主细胞并且从所述培养物中收集fad-gdh来产生fad-gdh。培养宿主细胞的培养基的实例包括包含一种或多种类型的无机盐,如氯化钠、磷酸二氢钾、磷酸氢二钾、硫酸镁、氯化镁、氯化铁、硫化铁或硫酸锰和一种或多种类型的氮源,如酵母提取物、胰蛋白胨、蛋白胨、牛肉膏、玉米浆或者大豆或麦麸提取物的培养基。必要时,所述培养基还可以包含碳水化合物源或维生素。培养基的初始ph可以调节至ph6至9,尽管不限于该范围。可以通过在(例如)10-42℃,如25℃或37℃的培养温度,通风搅拌深层培养、振荡培养或稳态培养1小时至4天或以上来进行培养。在培养之后,可以从培养物中收集fad-gdh。这可以使用常规方法进行。在一个实例中,可以使用超声破碎使微生物细胞破碎或者将其研磨或压碎,可以使用裂解酶,如溶菌酶或丝状真菌破壁酶(yatalase)提取酶,或者可以通过搅拌细胞或者将细胞暴露于甲苯来使细胞裂解从而使酶从所述细胞中释放。然后,可以通过过滤所述溶液来获得粗fad-gdh,可以通过离心除去固体部分,必要时可以通过使用盐酸链霉素、硫酸鱼精蛋白或硫酸锰除去核酸,硫酸铵、乙醇或丙酮可以用于分级分离,并且可以收集沉淀物。可以使用传统方法纯化粗fad-gdh酶。可以使用以下方法:使用凝胶,如sephadex(注册商标)、ultrogel(注册商标)或者biogel(registeredtrademark)的凝胶过滤法,使用离子交换树脂的吸附洗脱法、使用聚丙烯酰胺凝胶的电泳法,使用羟基磷灰石的吸附洗脱法,沉淀法,如蔗糖密度梯度离心、亲和色谱法或者使用分子筛膜或中空纤维膜的分级分离法或它们的组合进行纯化。可以使用比色葡萄糖测定试剂盒进行葡萄糖浓度测量。可以使用含有fad-gdh、电子受体和反应加速剂的组合物。选自n-(2-乙酰胺基)亚氨基二乙酸(ada)、双(2-羟乙基)亚胺基三(羟甲基)甲烷(bis-tris)、碳酸钠和咪唑的试剂可以保留在葡萄糖测定试剂盒的反应层中。可选地,可以添加ph缓冲液或显色试剂。然后,添加含有葡萄糖的样品,并使反应发生一定时间量。在反应期间,监测吸光度。吸光度对应于由于还原而褪色的电子受体或者由于从电子受体接受电子而通过聚合反应所形成的色素的吸收波长,优选地最大吸收波长。可以参考通过使用具有已知标准浓度的葡萄糖溶液所产生的标准曲线计算样品中的葡萄糖浓度。在变化率法中,可以基于每单位时间吸光度的变化率计算浓度。在终点法中,可以基于样品中所有葡萄糖已消耗(氧化)的点计算浓度。在使用介体和显色试剂的一个实例中,可以通过添加作为电子受体的2,6-二氯靛酚(dcip),然后监测600nm吸光度的减少来定量葡萄糖。此外,可以通过添加吩嗪硫酸甲酯(pms)形式的电子受体和硝基四唑蓝(ntb)形式的显色试剂,然后通过测量570nm的吸光度确定二甲瓒的量来计算葡萄糖浓度。当然可以使用其他电子受体和显色试剂。包含fad-gdh的葡萄糖传感器在一个实施方式中,提供了包含fad-gdh的葡萄糖传感器。在一个实施方式中,葡萄糖传感器包含含有fad-gdh的工作电极、参比电极和反电极。电极,如碳电极、钯电极、金电极或铂电极可以用作工作电极,并且fad-gdh可以固定化在工作电极上。固定化的fad-gdh的类型可以是可溶型fad-gdh。作为另外一种选择或者另外地,可以将电子介体固定化在工作电极上。反电极可以是常规电极,如铂电极或pt/c电极。参比电极可以是常规电极,如ag/agcl电极。固定化方法的实例包括使用交联剂的方法、包括包埋在聚合物基质中的方法、包括用透析膜覆盖的方法和使用光致可交联聚合物、导电聚合物和氧化还原聚合物的方法。作为另外一种选择,可以将fad-gdh在聚合物中与电子介体,如二茂铁或其衍生物一起固定化,通过吸附至电极固定化,或者它们的组合。在典型的方法中,可以使用戊二醛将fad-gdh固定化在碳电极上,然后使用包含氨基的试剂封闭戊二醛。另一种方法可以使用聚乙二醇二缩水甘油醚(pegde)作为交联剂。未限制pegde的分子量,并且可以使用任何类型。用于产生葡萄糖传感器的方法可见于文献,如liu等人,anal.chem.2012,84,3403-3409和tsujimura等人,j.am.chem.soc.2014,136,14432-14437(以上两篇文献以其全部内容作为参考并入)。可以通过使用gdh代替god来修改所公开的方法。例如,氧化还原聚合物,如含有锇络合物的聚合物可以与另一种聚合物,如聚乙二醇以及gdh在适合的缓冲液,如hepes缓冲液中混合以制备葡萄糖传感试剂。锇络合物可以是与一种或几种有机分子,包括二吡啶分子,如2,2'-二吡啶、联咪唑分子,如2,2'-联咪唑、吡啶-咪唑化合物,如2-(2-吡啶基)咪唑或它们的组合络合的锇。形成络合物的有机分子可以可选地被烷基,如c1-c6烷基,如甲基或乙基取代。在一个实施方式中,锇络合物可以如下所示:(os有机分子络合物)-(聚合物)形成络合物的有机分子之一可以可选地通过接头附接至聚合物。因此,在另一个实施方式中,锇络合物可以如下所示:(os有机分子络合物)-(接头)-(聚合物)在文献中公开了示例性分子,如ohara等人,analchem.1993dec1;65(23):3512-7和antiochia等人,materialssciencesandapplicationsvol.4no.7a2(2013)(以上两篇文献以其全部内容作为参考并入)。含有锇络合物的聚合物的实例包括锇双(2,2'-二吡啶)氯化物络合的聚(1-乙烯基咪唑)、锇双(2,2'-二吡啶)氯化物络合的聚(4-乙烯基吡啶)、聚(1-乙烯基咪唑)n-[锇(4,4'-二甲基-2,2'-双吡啶基)2cl2]2+/+及其衍生物。溶液可以沉积在碳传感器上以形成用作包含氧化还原聚合物-丝网连接的gdh的工作电极的有效电极区。可以加入膜聚合物和交联剂溶液的混合物以在传感器上形成膜。膜可以包含聚合物,如聚(1-乙烯基咪唑)、聚(4-乙烯吡啶)、衍生物或它们的组合。上述含有锇络合物的聚合物可以与这些膜聚合物组合使用以形成水凝胶膜。在一个实施方式中,葡萄糖传感器可以包含印刷电极。在此情况下,电极可以在绝缘基底上形成。具体地,可以通过光刻法或印刷技术,如丝网印刷、凹板印刷或胶版印刷在基底上形成电极。构成绝缘基底的材料的实例包括硅、玻璃、陶瓷、聚氯乙烯、聚乙烯、聚丙烯和聚酯。可以使用对多种溶剂或化学品显示出高耐受性的材料。在一个实施方式中,fad-gdh电极或葡萄糖传感器可以包含离子型聚合物,例如,聚乙烯亚胺(pei)、聚丙烯酸(pa)、ε-聚赖氨酸、α-聚赖氨酸或聚苯乙烯。未限制这些聚合物的分子量,并且可以使用任何类型。可以如下所示进行葡萄糖测量。将缓冲液置于恒温池中并保持在特定温度。铁氰化钾或吩嗪硫酸甲酯可以用作介体。制备了包含固定化fad-gdh的工作电极、反电极(如铂电极)和参比电极(如ag/agcl电极)。将恒定电压施加于碳电极,并且在电流稳定之后,添加含有葡萄糖的样品,接着测量电流的升高。然后,可以对照使用具有已知标准浓度的葡萄糖浓度所制备的标准曲线计算样品中的葡萄糖浓度。在一个实例中,将1.5u的fad-gdh固定化在玻璃碳(gc)电极上并且测量相对于葡萄糖浓度的响应电流值。将1.8ml50mm磷酸钾缓冲液(ph6.0)和0.2ml1m铁(iii)氰化钾(铁氰化钾)水溶液加入至电解池。将gc电极连接至bas100b/w恒电位器(basco.,ltd.),并在37℃搅拌溶液,然后将+500mv电压施加于ag/agcl参比电极。在另一个实例中,施加于ag/agcl参比电极的电压可以选自(但不限于)-100至+550之间的电压,例如,+400mv。将最终浓度为(例如)1mm、2mm、3mm、4mm、5mm、10mm、20mm、30mm和40mm的葡萄糖溶液加入至这些系统,然后测量每次添加的稳态电流值。将这些电流值对已知的葡萄糖浓度(例如,1mm、2mm、3mm、4mm、5mm、10mm、20mm、30mm和40mm)作图以制备标准曲线。可以通过将测量值与标准曲线相比较来定量葡萄糖浓度在另一个实例中,可以使用本文的固定化fad-gdh和传统方法进行循环伏安法。可以通过从0至+600mv扫描来进行测量。然后,可以选择特定电压来测量样品的葡萄糖浓度。这可以通过制备如上所描述的标准曲线,然后测量具有未知葡萄糖浓度的响应电流来进行。在一个实施方式中,包含fad-gdh的葡萄糖传感器或包含fad-gdh的工作电极可以包含保护薄膜(保护膜)。可以使用与用于评价gdh活力的系统相同的测定系统,如其中存在过量介体以接受电子的测定系统来确定god的酶活力。作为另外一种选择,可以使用其中氧为受体的测定系统来确定god的酶活力。作为另外一种选择,可以按照生产商的说明书来确定god的酶活力。对于初始活力和对于残余活力,应使用相同系统测量酶活力以计算残余活力比。以下是测量god活力的示例性测定法。还可以使用其他方法。god测定原理:β-d-葡萄糖+o2+h2o→d-葡萄糖酸-δ-内酯+h2o2(通过god催化)2h2o2+4-aa+ehspt→醌亚胺染料+4h2o(通过pod催化)(其中4-aa是4-氨基安替比林的缩写,ehspt是n-乙基-n-(2-羟基-3-磺丙基)-m-甲苯胺的缩写)可以通过分光光度法测量醌亚胺的产生。单位的定义:god单位可以定义为在以下条件下,能够引起每分钟形成1微摩尔过氧化氢(1/2微摩尔醌亚胺染料)的酶活力。god测定法:试剂a-f如下所示a.mes-na缓冲液ph5.7:0.1m(将2.13g2-(n-吗啉代)乙烷磺酸(mw=213.25)溶解在约60mlh2o中,在25℃,用1nnaoh将ph调解至5.7,用h2o补充至100ml)b.葡萄糖溶液:15%(溶解1.5gβ-d-葡萄糖并用h2o补充至10ml)(应新鲜制备)c.4-aa溶液:0.5%(50mg4-氨基安替比林(mw=203.25)/10mlh2o)d.ehspt溶液:40mm(118mgn-乙基-n-(2-羟基-3-磺丙基)-m-甲苯胺(mw=295.3)/10mlh2o)e.过氧化物酶溶液:500u(以purpurogalin单位表示)/ml的h2o溶液f.酶稀释剂:10mmmes-na缓冲液,ph5.7,包含0.1%tritonx-100测定混合物中的浓度:mes缓冲液:79mmd-葡萄糖:131mm4-aa:0.2mmehspt:0.3mmpod:约4u/ml程序:步骤1.在棕色瓶中新鲜制备以下工作溶液并在冰上储存。30ml缓冲溶液(a)6ml底物溶液(b)0.3ml4-aa溶液(c)0.3mlehspt溶液(d)0.3mlpod溶液(e)步骤2.将3.0ml工作溶液吸取至比色皿(d=1.0cm)并在37℃平衡约5分钟。步骤3.添加0.1ml酶溶液*并通过倒转轻轻混合。(*在测定之前,立即将酶制剂在冰冷的酶稀释剂(f)中溶解并用相同缓冲液稀释至0.05-0.2u/ml。)步骤4.在37℃恒温的分光光度计中,对水记录2-3min,555nm下的光密度增加,并根据曲线初始直线部分计算每分钟的δod(δod测试)。另外,通过使用与测试相同的方法测量空白率(δod空白),但条件是添加酶稀释剂(f)来替代酶溶液。可以如下所示计算god活力:δod/min(δod测试-δod空白)×vt×df重量活力(u/mg)=(u/ml)×1/cvt:总体积(3.1ml)vs:样品体积(0.1ml)32.8:测定条件下醌亚胺染料的毫摩尔消光系数(cm2/毫摩尔)1/2:基于1摩尔h2o2产生半摩尔醌亚胺染料的事实的因子。1.0:光程长(cm)df:稀释因子c:溶解的酶浓度(cmg/ml)。以下实施例详细地描述了本发明的方面。然而,不应以任何方式将本发明的技术范围视为或限制于以下实施例。本领域技术人员可以采取本文中所公开的概念的可选的特性、改变和变化,并且将这些改变和变化考虑为在该技术的范围内。实施例1除非另外说明,否则根据以下测试方法进行gdh和god的热稳定性和储存稳定性评价。测试方法(1)引入来自属于毛霉属(mucor)的微生物的gdh基因和转化体中gdh活力的确认简要地,通过向其中引入突变n66y/n68g/c88a/q233r/t387c/e554d/l557v/s559k来修饰来自属于毛霉属(mucor)的微生物的gdh(mpgdh,seqidno:1)以获得编码修饰的gdh的基因(其在本文中可以称为mpgdh-m1,或者gdh-m1)。seqidno:3中显示了mpgdh-m1的氨基酸序列,而seqidno:4中显示了碱基序列。通过将mpgdh-m1基因引入到质粒puc19的多克隆位点获得了dna构建体。更具体地,将in-fusionhd克隆试剂盒(clontech)中的puc19线性化载体用作puc19载体。根据试剂盒说明书中的规程,通过使用所述in-fusionhd克隆试剂盒将mpgdh-m1基因连接至puc19的多克隆位点的in-fusion克隆位点以获得构建体质粒(puc19-mpgdh-m1)。将所获得的重组质粒puc19-mpgdh-m1用作模板,并且以下列条件实施使用seqidno:5至10所示的合成寡核苷酸和kod-plus-(toyobo)的pcr反应。加入5μl1×kod-plus-缓冲液、5μl包含2mm每种dntp的混合物溶液、2μl25mmmgso4溶液、50ng包含连接的模板mpgdh-m1基因的dna构建体、15pmol的以上每种合成寡核苷酸和1个单位kod-plus-,并用无菌水将总体积调节至50μl。将所制备的反应溶液应用于热循环仪(eppendorf)并在94℃培育2min,然后将循环“94℃,15秒”-“50℃,30秒”-“68℃,8分钟”重复30次。将反应溶液部分应用于1.0%的琼脂糖凝胶并进行电泳,并且确认具体地扩增了约8,000bp的dna片段。用限制性内切酶dpni(newenglandbiolabs)消化所得dna,并在切割剩余的模板dna之后,转化大肠杆菌(ecoli.)jm109株并涂布在lb-amp琼脂培养基上。将生长的菌落接种至2.5mllb-amp培养基(含有1%(w/v)细菌用胰蛋白胨、0.5%(w/v)蛋白胨、0.5%(w/v)nacl、50μg/ml氨苄西林)中并在37℃振荡培养20小时以获得培养物。通过7,000rpm离心5分钟获得细菌细胞以收集细胞。然后,使用qiagentip-100(qiagen)对细菌细胞进行重组质粒提取以提取和纯化重组质粒,并且获得了2.5μgdna。使用多毛细管dna分析系统appliedbiosystems3130xl基因分析仪(lifetechnologies)确定质粒中编码mpgdh-m1的dna碱基序列,并因此获得了编码突变体mpgdh-m1/a175c/n214c/g466d(seqidno:12)(在本文中也称为mpgdh-m2,或者gdh-m2)的dna构建体(seqidno:11)。在酱油曲霉(aspergillussojae)中表达该基因,并确认其gdh活力。更具体地,将来自属于毛霉属(mucor)的微生物的gdh基因用作起始材料并设计修饰的gdh基因以获得适合于在酱油曲霉(aspergillussojae)中重组表达的gdh。更具体地,基于seqidno:1的gdh基因序列,设计了具有适合于在宿主中表达的密码子频率的密码子优化的基因序列并合成了整个基因。seqidno:2中显示了该完全合成的dna序列。实施双-连接pcr(fungalgeneticsandbiology,2004,41卷,973-981页)来构建由5'臂区-pyrg基因(尿嘧啶营养缺陷型标志物)-tef1启动子基因-黄素结合gdh基因-3'臂区所组成的盒。如下所示,该盒将转化至来自酱油曲霉(aspergillussojae)nbrc4239株的pyrg缺陷型株(其中pyrg基因上游48个碱基对、896bp编码区和下游240个碱基对缺失的菌株)。将来自酱油曲霉(aspergillussojae)nbrc4239的pyrg-打断菌株的分生孢子接种至处于500ml锥形烧瓶中的100ml包含20mm尿嘧啶核苷的多聚蛋白胨糊精液体培养基中,并且在30℃振荡培养约20小时之后,回收细胞。从所回收的细胞制备原生质体。通过所获得的原生质体和20μg靶标基因-插入的dna构建体,使用原生质体peg法进行转化。随后,将细胞在30℃用含有0.5%(w/v)琼脂和1.2m山梨糖醇的czapek-dox基本培养基(difcoco.,ph6)一起培育5天或以上以获得具有形成菌落能力的转化的酱油曲霉(aspergillussojae)。在所得转化的酱油曲霉(aspergillussojae)中,引入作为能够补充尿嘧啶核苷营养缺陷型的基因的pyrg,并因此转化体可以在不含尿嘧啶核苷的培养基上生长,并且可以选作其中引入了所关心的基因的那些菌株。从所得菌株中,通过pcr确认选择所期望的转化体。使用通过mpgdh-m2基因转化的酱油曲霉(aspergillussojae)并产生gdh。将每个菌株的分生孢子接种至200ml锥形烧瓶中的40mldpy液体培养基(1%(w/v)多聚蛋白胨、2%(w/v)糊精、0.5%(w/v)酵母提取物、0.5%(w/v)kh2po4、0.05%(w/v)mgso4·7h2o;ph未调节)中并在30℃,以160rpm振荡培养4天。然后,在培养之后,从培养物中过滤出细胞,并通过amiconultra-15,30knmwl(millipore)将所得培养基的上清液部分浓缩至10ml。然后,将其上样至通过含有150mmnacl的20mm磷酸钾缓冲液(ph6.5)平衡的hiload26/60superdex200pg(gehealthcare)柱并用相同缓冲液洗脱。收集显示出gdh活力的部分以获得mpgdh-m2的纯化产物。(2)gdh和god的热稳定性评价来自黑曲霉(aspergillusniger)的god购自sigma-aldrich(x-s型,在本文中称为god-1)。来自曲霉属(aspergillus)菌株的另一种god购自toyobo(产品目录no.glo-201,在本文中称为god-2)。来自黑曲霉(aspergillusniger)的god购自wakopurechemicalindustries,ltd(产品目录no.074-02401,在本文中称为god-3)。来自黑曲霉(aspergillusniger)的另一种god购自sigma-aldrich(vii型,在本文中称为god-4)。表示为gld3的gdh购自funakoshi(发现该酶的氨基末端与来自冻土毛霉(mucorhiemalis)的gdh相同,并且该酶非常可能来自属于毛霉属(mucor)的微生物,冻土毛霉(mucorhiemalis))(在本文中gld3还可以称为gdh-gld3)。当使用gld3时,使用amiconultra0.5ml过滤器(merck,截留分子量30kda)进行超滤以除去低分子量成分,并且用pbs(ph7.4)替换溶液。如上所描述,制备mpgdh-m2。将酶稀释以重新获得约1u/ml的稀酶溶液(100mm磷酸钾缓冲液,ph7.0)。制备了分别为0.1ml的一对这种酶溶液,并且将一个保持在4℃,而将另一个在57℃,ph7热处理15分钟。在热处理之后,确定每种酶的活力,并且作为与保持在4℃的酶的酶活力(100%)相比,在57℃,ph7热处理15分钟之后的残余活力的百分比(%)计算“残余活力比(%)”。图1中显示了结果。与gdh相比,god显示出优良的热稳定性并且在严苛的温度条件下是稳定的。(3)gdh和god的储存稳定性评价用pbs溶液替换分别为1mg/ml的gdh和god溶液,并且在40℃储存持续的一段时间。在1天、4天、1周、2周和3周后,测量每种酶的gdh活力或god活力。通过将在40℃储存之前的活力设置为100%,将“剩余活力率(%)”计算为在40℃储存后的活力值。图2至图4中显示了结果。因此,在1周之后,god-3的残余活力比为22%,god-4为22%,god-2为30%,并且god-1为41%。也就是说,在40℃保持1周之后,所有god显示出显著降低的活力。此外,在2周之后,god-3的残余活力比为9%,god-4为8%,god-2为13%,并且god-1为30%,并且所有god再次显示出显著降低的活力。另一方面,在1周之后,mpgdh-m2的残余活力比为80%,并且gld3为88%。也就是说,gdh将它们的活力保持了持续的一段时间。在2周之后,mpgdh-m2的残余活力比为72%,gld3为83%,并且gdh再次将它们的活力保持了持续的一段时间。即使在3周后,mpgdh-m2保持了其初始活力的约73%,然而god-2在相同时间段内丧失了其大部分活力(4%)。参见图2。当比较在57℃、ph7处理15分钟后的残余活力时,每种god均具有比mpgdh-m2和gld3更高的残余活力和更高的稳定性。然而,意外地发现与gdh相比,在40℃储存试验中god的稳定性较低,并且发现尽管它们在严苛的温度条件下具有优良的热稳定性,但是god可能对于在cgm中的使用不是最优的。相反地,证明gdh适合于长期使用,如在cgm中的长期使用。由于gdh甚至在40℃储存试验中保持了相对高的活力,因此非常可能当在37℃储存时,gdh还将保持相对高的活力。实施例2gdh/god固定化电极的储存稳定性测试如下所示,产生了gdh/god固定化电极并对其测试了储存稳定性。产生电极的方法将3μl5%的聚乙烯亚胺(pei)(产品目录号no.164-17821,平均分子量10000,来自wako,阳离子聚合物)加入到基于碳的丝网印刷电极上,然后干燥。然后,加入0.5μl在98℃处理1小时的锇聚合物(产品目录号no.002096,bas),然后干燥。然后,加入3μl35mg/mlgdh/god,然后干燥。然后,加入3μl2%的聚乙二醇二缩水甘油醚(pegde,平均分子量500,sigma-aldrich生产)并在4℃储存过夜。*还使用1%聚丙烯酸(pa)(产品目录号no.416037-100ml,平均分子量15000,来自sigma-aldrich,阴离子聚合物)代替pei来产生电极。储存稳定性测试用超纯水清洗如上所产生的电极并在30℃干燥1小时。然后,每2小时,取出电极并进行电化学测量。在电化学测量之后,将电极再次以干燥状态储存在30℃的培育箱中。电化学测量将参比电极ag/agcl、反电极(铂)和作为工作电极的基于gdh或god的丝网印刷电极置于100mm磷酸钾缓冲液(ph7)中。然后,实施从0至+600mv扫描的循环伏安法测量。对于其中未添加葡萄糖的条件和其中添加了5mm葡萄糖的条件,比较+500mv的氧化电流值,并将差异作为响应电流值。在0、2和4hr进行测量,并且以0hr时的响应电流值作为100%,记录相对值。结果对于通过使用pei所产生的电极,在2小时之后,gdh-m2电极的响应电流为67.3%。通过比较,god-3丧失了较多的活力并且在2小时,god-3电极的剩余响应电流仅为30.8%。在存储4小时之后观察到了类似的结果,并且god-3的相对响应电流值低于gdh-m2。参见图5。对于通过使用pa所产生的电极,即使在0小时,使用god-3固定化电极也不可以观察到响应电流。通过比较,对于使用pa的gdh-m2固定化电极,可以观察到响应电流。在储存2小时之后,保持了68.3%的活力,并且在4小时之后,保持了47.1%的初始活力,其表明稳定性优于通过使用pei固定化的电极。参见图6。应注意在本实验中评价的电极没有保护膜并因此条件比实际cgm中的条件更严苛和严重。显示出比当前cgm装置中使用的具有葡萄糖氧化酶的传感器更高的储存稳定性的具有葡糖脱氢酶的传感器可以有利地用于长期测量。实施例3最适ph评价了葡糖脱氢酶gdh-m2和gld3的最适ph。如下所示进行活力测定。通过测量520nm下dcip的时间依赖性减小来进行2,6-二氯-靛酚(dcip)测定(300mmdcip/1mmpms/400mm葡萄糖)。对于ph5.5至6.0,使用柠檬酸-钠缓冲液,而对于ph6至8,使用磷酸-钾缓冲液。参见图7(gdh-m2)和图8(gld3)。通过比较,根据生产商的产品目录,god-3的最适ph报告为ph5.6。实施例4葡萄糖浓度依赖性如下所示,评价了具有固定化酶的电极的葡萄糖浓度依赖性。如以上实施例2中所示,使用pei、gdh-m2、os聚合物和pegde制备传感器。然后,将电极与ag/agcl参比电极和铂反电极(辅助电极)一起置于10ml100mm磷酸钾缓冲溶液(ph7)中以组成三电极系统。通过该三电极系统进行测量。在以350rpm搅拌的同时,加入预定浓度的葡萄糖。然后,进行从0至+600mv扫描的循环伏安法测量,并记录+400mv的氧化电流值。图9和10中显示了结果。据证实可以从1至30.1mm葡萄糖确认响应电流的葡萄糖浓度依赖性。实施例5分子量如下所示,通过sds-page评价酶的分子量。在37℃,endoh内切糖苷酶(newenglandbiolabs)处理24小时。然后,通过5-20%梯度凝胶进行sds-page。m2的分子量为约70kda,并且gld3的分子量为约65至80kda。参见图11。实施例6底物特异性如下所示,评价gdh-m2的底物特异性。更具体地,底物糖的浓度为200mm。将对葡萄糖的活力设置为100%,并且以百分比表示对其他底物的相对活力。表1gdh-m2gld3葡萄糖100%100%麦芽糖0.37%0.67%木糖0.55%1.60%*gld3的值为根据生产商所提供的产品目录的那些在本文中参考的每篇专利、专利申请、专利公开和文档以其全部内容作为参考并入。对以上专利、专利申请、专利公开和文档的引用或提及不是对任何上述内容是相关现有技术的承认,并且它也不构成有关这些专利公开或文档的内容或日期的任何承认。在本说明书中所描述的说明和定义优先于作为参考并入本文的文档中的那些。可以在不存在本文具体公开的任何元素的情况下适当地实践本文说明性地描述的技术。已使用的术语和表达用作说明术语而非限制术语。这些术语和表达的使用不排除所示或特征或其部分的任何等价形式,并且在所主张的技术范围内,多种修改是可能的。除非上下文明确表示描述了任一种元素或不止一种元素,否则术语“一个(a)”或“一种(an)”可以表示一个或多个它所修饰的元素(例如,“一个试剂”可以表示一个或多个试剂)。在本文中的每种情况下,术语“包括”、“基本由……组成”和“由……组成”中的任一个可以用其他两个术语替换。工业实用性在一些实施方式中,本发明的方法可以用于(例如)连续葡萄糖监测。序列的简要说明seqidno:1普雷恩毛霉(mucorprainii)wtgdhaaseqidno:2普雷恩毛霉(mucorprainii)wtgdhdnaseqidno:3mpgdh-m1aaseqidno:4mpgdh-m1dnaseqidno:5-10引物seqidno:11mpgdh-m2aaseqidno:12mpgdh-m2dnaseqidno:13mhgdhaaseqidno:14mhgdhdna序列表<110>龟甲万株式会社(kikkomancorporation)<120>使用fad依赖性葡糖脱氢酶的连续葡萄糖监测<130>ph-6896-pct<150>us62/479442<151>2017-03-31<160>14<170>patentinversion3.5<210>1<211>641<212>prt<213>普雷恩毛霉(mucorprainii)<400>1metlysilethralaalaileilethrvalalathralaphealaser151015phealaseralaglnglnaspthrasnserserserthraspthrtyr202530asptyrvalilevalglyglyglyvalalaglyleualaleualaser354045argilesergluasnlysaspvalthrvalalavalleuglusergly505560proasnalaasnaspargphevalvaltyralaproglymettyrgly65707580glnalavalglythraspleucysproleuileprothrthrprogln859095gluasnmetglyasnargserleuthrilealathrglyargleuleu100105110glyglyglyseralaileasnglyleuvaltrpthrargglyglyleu115120125lysasptyraspalatrpglugluleuglyasnproglytrpasngly130135140alaasnleuphelystyrphelyslysvalgluasnphethrpropro145150155160thrproalaglnileglutyrglyalathrtyrglnlysseralahis165170175glylyslysglyproileaspvalserphethrasntyrglupheser180185190glnseralasertrpasnalaserleugluthrleuaspphethrala195200205leuproaspileleuasnglythrleualaglytyrserthrthrpro210215220asnileleuaspprogluthrvalglnargvalaspsertyrthrgly225230235240tyrilealaprotyrthrserargasnasnleuasnvalleualaasn245250255histhrvalserargileglnphealaprolysasnglyserglupro260265270leulysalathrglyvalglutrptyrprothrglyasnlysasngln275280285lysglnileilelysalaargtyrgluvalileileserserglyala290295300ileglyserprolysleuleugluileserglyileglyasnlysasp305310315320ilevalseralaalaglyvalgluserleuileaspleuproglyval325330335glyserasnmetglnasphisvalhisalailethrvalserthrthr340345350asnilethrglytyrthrthrasnservalphevalasngluthrleu355360365alaglngluglnarggluglutyrglualaasnlysthrglyiletrp370375380alathrthrproasnasnleuglytyrprothrprogluglnleuphe385390395400asnglythrgluphevalserglylysgluphealaasplysilearg405410415asnserthraspglutrpalaasntyrtyralaserthrasnalaser420425430asnvalgluleuleulyslysglntyralailevalalaserargtyr435440445glugluasntyrleuserproilegluileasnphethrproglytyr450455460gluglyserglyasnvalaspleuglnasnasnlystyrglnthrval465470475480asnhisvalleuilealaproleuserargglytyrthrhisileasn485490495serseraspvalgluasphisservalileasnproglntyrtyrser500505510hisprometaspileaspvalhisilealaserthrlysleualaarg515520525gluileilethralaserproglyleuglyaspileasnserglyglu530535540ilegluproglymetasnilethrsergluaspaspleuargsertrp545550555560leuserasnasnvalargserasptrphisprovalglythrcysala565570575metleuprolysgluleuglyglyvalvalserproalaleumetval580585590tyrglythrserasnleuargvalvalaspalaserilemetproleu595600605gluvalserserhisleumetglnprothrtyrglyilealaglulys610615620alaalaaspileilelysasnphetyrlysthrglnhislysasngln625630635640asn<210>2<211>1926<212>dna<213>普雷恩毛霉(mucorprainii)<400>2atgaagatcacagctgccattatcactgttgccacagcatttgcttcttttgcttctgct60caacaagacacaaattcttcctcaactgatacttatgattatgttatcgttggcggcggt120gtagctggtttggctttggctagtcgtatctctgaaaacaaggatgtcactgttgctgtt180ctcgagtccggtcctaatgccaatgatagatttgttgtttatgctcctggcatgtatggc240caagctgttggcactgatctctgtcctctcattcctactactcctcaagaaaatatgggc300aacagaagtctcacaatcgctactggtagattgctcggtggtggcagtgctattaatggt360ctcgtttggacccgtggtggcttgaaggattacgatgcttgggaggagctcggtaaccct420ggatggaacggtgccaacttgttcaagtactttaagaaggtcgaaaacttcactcctcct480actcctgcccaaattgaatacggcgctacttatcagaaaagtgctcatggcaagaaggga540cctattgatgtctctttcacgaactacgagttctctcaatctgctagctggaacgcctca600ctcgaaacccttgatttcactgcacttcctgatatcttgaacggtactttggccggttac660tctaccactcccaacattttggaccctgagactgttcaacgtgttgattcctatactggt720tacattgctccttacactagccgtaacaacctcaatgttttggccaaccataccgtctcc780cgcattcaatttgctcccaagaatggtagcgaacctctcaaggctaccggtgttgaatgg840tatcccactggcaacaagaatcaaaagcaaattatcaaggcccgttatgaagttatcatc900tcatctggtgccattggtagtcctaagcttttggaaatctctggtatcggtaataaggat960atcgtctctgctgctggtgtcgagtccttgattgacttgcctggcgttggttccaacatg1020caagatcacgttcatgctatcactgtctctactaccaatattactggctatactaccaac1080agcgtctttgtcaatgaaacccttgcccaagaacaaagagaagaatatgaagccaacaag1140actggtatctgggctactactcccaacaacctcggttatcctacgcccgaacaactcttc1200aatggcaccgaattcgtttctggaaaggagtttgctgacaagattcgtaactctactgat1260gaatgggccaactattatgcttccaccaacgcctccaatgtcgagttattaaagaagcaa1320tatgctattgtcgcctctcgttacgaagagaactacttgtctcctattgaaatcaacttc1380actcctggttatgagggtagcggtaatgtcgatttgcaaaacaacaagtaccaaactgtc1440aaccatgtcttgattgctcctttaagtcgtggttatactcacattaactcttctgatgtg1500gaggatcattctgtcattaatccccaatactactctcatcctatggatattgatgtccat1560atcgcttccactaaacttgctcgcgaaatcatcactgcctctcccggtcttggtgacatt1620aacagtggcgaaatcgaacccggtatgaatattacttctgaagacgaccttagatcttgg1680ttgagtaataatgtccgttctgactggcatcctgttggtacttgtgctatgcttcccaag1740gaattaggtggtgttgtcagccccgctctcatggtttacggcacttccaacttgcgtgtt1800gttgatgcttcgattatgcccctcgaagtctcttctcatttgatgcaacccacctacggt1860attgctgagaaggctgctgacattattaagaatttctacaagactcaacacaagaaccaa1920aattag1926<210>3<211>641<212>prt<213>普雷恩毛霉(mucorprainii)<400>3metlysilethralaalaileilethrvalalathralaphealaser151015phealaseralaglnglnaspthrasnserserserthraspthrtyr202530asptyrvalilevalglyglyglyvalalaglyleualaleualaser354045argilesergluasnlysaspvalthrvalalavalleuglusergly505560protyralaglyaspargphevalvaltyralaproglymettyrgly65707580glnalavalglythraspleualaproleuileprothrthrprogln859095gluasnmetglyasnargserleuthrilealathrglyargleuleu100105110glyglyglyseralaileasnglyleuvaltrpthrargglyglyleu115120125lysasptyraspalatrpglugluleuglyasnproglytrpasngly130135140alaasnleuphelystyrphelyslysvalgluasnphethrpropro145150155160thrproalaglnileglutyrglyalathrtyrglnlysseralahis165170175glylyslysglyproileaspvalserphethrasntyrglupheser180185190glnseralasertrpasnalaserleugluthrleuaspphethrala195200205leuproaspileleuasnglythrleualaglytyrserthrthrpro210215220asnileleuaspprogluthrvalargargvalaspsertyrthrgly225230235240tyrilealaprotyrthrserargasnasnleuasnvalleualaasn245250255histhrvalserargileglnphealaprolysasnglyserglupro260265270leulysalathrglyvalglutrptyrprothrglyasnlysasngln275280285lysglnileilelysalaargtyrgluvalileileserserglyala290295300ileglyserprolysleuleugluileserglyileglyasnlysasp305310315320ilevalseralaalaglyvalgluserleuileaspleuproglyval325330335glyserasnmetglnasphisvalhisalailethrvalserthrthr340345350asnilethrglytyrthrthrasnservalphevalasngluthrleu355360365alaglngluglnarggluglutyrglualaasnlysthrglyiletrp370375380alathrcysproasnasnleuglytyrprothrprogluglnleuphe385390395400asnglythrgluphevalserglylysgluphealaasplysilearg405410415asnserthraspglutrpalaasntyrtyralaserthrasnalaser420425430asnvalgluleuleulyslysglntyralailevalalaserargtyr435440445glugluasntyrleuserproilegluileasnphethrproglytyr450455460gluglyserglyasnvalaspleuglnasnasnlystyrglnthrval465470475480asnhisvalleuilealaproleuserargglytyrthrhisileasn485490495serseraspvalgluasphisservalileasnproglntyrtyrser500505510hisprometaspileaspvalhisilealaserthrlysleualaarg515520525gluileilethralaserproglyleuglyaspileasnserglyglu530535540ilegluproglymetasnilethrseraspaspaspvalarglystrp545550555560leuserasnasnvalargserasptrphisprovalglythrcysala565570575metleuprolysgluleuglyglyvalvalserproalaleumetval580585590tyrglythrserasnleuargvalvalaspalaserilemetproleu595600605gluvalserserhisleumetglnprothrtyrglyilealaglulys610615620alaalaaspileilelysasnphetyrlysthrglnhislysasngln625630635640asn<210>4<211>1926<212>dna<213>普雷恩毛霉(mucorprainii)<400>4atgaagatcacagctgccattatcactgttgccacagcatttgcttcttttgcttctgct60caacaagacacaaattcttcctcaactgatacttatgattatgttatcgttggcggcggt120gtagctggtttggctttggctagtcgtatctctgaaaacaaggatgtcactgttgctgtt180ctcgagtccggtccttatgccggtgatagatttgttgtttatgctcctggcatgtatggc240caagctgttggcactgatctcgctcctctcattcctactactcctcaagaaaatatgggc300aacagaagtctcacaatcgctactggtagattgctcggtggtggcagtgctattaatggt360ctcgtttggacccgtggtggcttgaaggattacgatgcttgggaggagctcggtaaccct420ggatggaacggtgccaacttgttcaagtactttaagaaggtcgaaaacttcactcctcct480actcctgcccaaattgaatacggcgctacttatcagaaaagtgctcatggcaagaaggga540cctattgatgtctctttcacgaactacgagttctctcaatctgctagctggaacgcctca600ctcgaaacccttgatttcactgcacttcctgatatcttgaacggtactttggccggttac660tctaccactcccaacattttggaccctgagactgttcgacgtgttgattcctatactggt720tacattgctccttacactagccgtaacaacctcaatgttttggccaaccataccgtctcc780cgcattcaatttgctcccaagaatggtagcgaacctctcaaggctaccggtgttgaatgg840tatcccactggcaacaagaatcaaaagcaaattatcaaggcccgttatgaagttatcatc900tcatctggtgccattggtagtcctaagcttttggaaatctctggtatcggtaataaggat960atcgtctctgctgctggtgtcgagtccttgattgacttgcctggcgttggttccaacatg1020caagatcacgttcatgctatcactgtctctactaccaatattactggctatactaccaac1080agcgtctttgtcaatgaaacccttgcccaagaacaaagagaagaatatgaagccaacaag1140actggtatctgggctacttgtcccaacaacctcggttatcctacgcccgaacaactcttc1200aatggcaccgaattcgtttctggaaaggagtttgctgacaagattcgtaactctactgat1260gaatgggccaactattatgcttccaccaacgcctccaatgtcgagttattaaagaagcaa1320tatgctattgtcgcctctcgttacgaagagaactacttgtctcctattgaaatcaacttc1380actcctggttatgagggtagcggtaatgtcgatttgcaaaacaacaagtaccaaactgtc1440aaccatgtcttgattgctcctttaagtcgtggttatactcacattaactcttctgatgtg1500gaggatcattctgtcattaatccccaatactactctcatcctatggatattgatgtccat1560atcgcttccactaaacttgctcgcgaaatcatcactgcctctcccggtcttggtgacatt1620aacagtggcgaaatcgaacccggtatgaatattacttctgacgacgacgttagaaaatgg1680ttgagtaataatgtccgttctgactggcatcctgttggtacttgtgctatgcttcccaag1740gaattaggtggtgttgtcagccccgctctcatggtttacggcacttccaacttgcgtgtt1800gttgatgcttcgattatgcccctcgaagtctcttctcatttgatgcaacccacctacggt1860attgctgagaaggctgctgacattattaagaatttctacaagactcaacacaagaaccaa1920aattag1926<210>5<211>30<212>dna<213>人工<220><223>引物<400>5tatcagaaaagttgtcatggcaagaaggga30<210>6<211>30<212>dna<213>人工<220><223>引物<400>6agagacatcaataggtcccttcttgccatg30<210>7<211>30<212>dna<213>人工<220><223>引物<400>7cctgatatcttgtgcggtactttggccggt30<210>8<211>30<212>dna<213>人工<220><223>引物<400>8gggagtggtagagtaaccggccaaagtacc30<210>9<211>30<212>dna<213>人工<220><223>引物<400>9cctggttatgaggacagcggtaatgtcgat30<210>10<211>30<212>dna<213>人工<220><223>引物<400>10cttgttgttttgcaaatcgacattaccgct30<210>11<211>641<212>prt<213>普雷恩毛霉(mucorprainii)<400>11metlysilethralaalaileilethrvalalathralaphealaser151015phealaseralaglnglnaspthrasnserserserthraspthrtyr202530asptyrvalilevalglyglyglyvalalaglyleualaleualaser354045argilesergluasnlysaspvalthrvalalavalleuglusergly505560protyralaglyaspargphevalvaltyralaproglymettyrgly65707580glnalavalglythraspleualaproleuileprothrthrprogln859095gluasnmetglyasnargserleuthrilealathrglyargleuleu100105110glyglyglyseralaileasnglyleuvaltrpthrargglyglyleu115120125lysasptyraspalatrpglugluleuglyasnproglytrpasngly130135140alaasnleuphelystyrphelyslysvalgluasnphethrpropro145150155160thrproalaglnileglutyrglyalathrtyrglnlyssercyshis165170175glylyslysglyproileaspvalserphethrasntyrglupheser180185190glnseralasertrpasnalaserleugluthrleuaspphethrala195200205leuproaspileleucysglythrleualaglytyrserthrthrpro210215220asnileleuaspprogluthrvalargargvalaspsertyrthrgly225230235240tyrilealaprotyrthrserargasnasnleuasnvalleualaasn245250255histhrvalserargileglnphealaprolysasnglyserglupro260265270leulysalathrglyvalglutrptyrprothrglyasnlysasngln275280285lysglnileilelysalaargtyrgluvalileileserserglyala290295300ileglyserprolysleuleugluileserglyileglyasnlysasp305310315320ilevalseralaalaglyvalgluserleuileaspleuproglyval325330335glyserasnmetglnasphisvalhisalailethrvalserthrthr340345350asnilethrglytyrthrthrasnservalphevalasngluthrleu355360365alaglngluglnarggluglutyrglualaasnlysthrglyiletrp370375380alathrcysproasnasnleuglytyrprothrprogluglnleuphe385390395400asnglythrgluphevalserglylysgluphealaasplysilearg405410415asnserthraspglutrpalaasntyrtyralaserthrasnalaser420425430asnvalgluleuleulyslysglntyralailevalalaserargtyr435440445glugluasntyrleuserproilegluileasnphethrproglytyr450455460gluaspserglyasnvalaspleuglnasnasnlystyrglnthrval465470475480asnhisvalleuilealaproleuserargglytyrthrhisileasn485490495serseraspvalgluasphisservalileasnproglntyrtyrser500505510hisprometaspileaspvalhisilealaserthrlysleualaarg515520525gluileilethralaserproglyleuglyaspileasnserglyglu530535540ilegluproglymetasnilethrseraspaspaspvalarglystrp545550555560leuserasnasnvalargserasptrphisprovalglythrcysala565570575metleuprolysgluleuglyglyvalvalserproalaleumetval580585590tyrglythrserasnleuargvalvalaspalaserilemetproleu595600605gluvalserserhisleumetglnprothrtyrglyilealaglulys610615620alaalaaspileilelysasnphetyrlysthrglnhislysasngln625630635640asn<210>12<211>1926<212>dna<213>普雷恩毛霉(mucorprainii)<400>12atgaagatcacagctgccattatcactgttgccacagcatttgcttcttttgcttctgct60caacaagacacaaattcttcctcaactgatacttatgattatgttatcgttggcggcggt120gtagctggtttggctttggctagtcgtatctctgaaaacaaggatgtcactgttgctgtt180ctcgagtccggtccttatgccggtgatagatttgttgtttatgctcctggcatgtatggc240caagctgttggcactgatctcgctcctctcattcctactactcctcaagaaaatatgggc300aacagaagtctcacaatcgctactggtagattgctcggtggtggcagtgctattaatggt360ctcgtttggacccgtggtggcttgaaggattacgatgcttgggaggagctcggtaaccct420ggatggaacggtgccaacttgttcaagtactttaagaaggtcgaaaacttcactcctcct480actcctgcccaaattgaatacggcgctacttatcagaaaagttgtcatggcaagaaggga540cctattgatgtctctttcacgaactacgagttctctcaatctgctagctggaacgcctca600ctcgaaacccttgatttcactgcacttcctgatatcttgtgcggtactttggccggttac660tctaccactcccaacattttggaccctgagactgttcgacgtgttgattcctatactggt720tacattgctccttacactagccgtaacaacctcaatgttttggccaaccataccgtctcc780cgcattcaatttgctcccaagaatggtagcgaacctctcaaggctaccggtgttgaatgg840tatcccactggcaacaagaatcaaaagcaaattatcaaggcccgttatgaagttatcatc900tcatctggtgccattggtagtcctaagcttttggaaatctctggtatcggtaataaggat960atcgtctctgctgctggtgtcgagtccttgattgacttgcctggcgttggttccaacatg1020caagatcacgttcatgctatcactgtctctactaccaatattactggctatactaccaac1080agcgtctttgtcaatgaaacccttgcccaagaacaaagagaagaatatgaagccaacaag1140actggtatctgggctacttgtcccaacaacctcggttatcctacgcccgaacaactcttc1200aatggcaccgaattcgtttctggaaaggagtttgctgacaagattcgtaactctactgat1260gaatgggccaactattatgcttccaccaacgcctccaatgtcgagttattaaagaagcaa1320tatgctattgtcgcctctcgttacgaagagaactacttgtctcctattgaaatcaacttc1380actcctggttatgaggacagcggtaatgtcgatttgcaaaacaacaagtaccaaactgtc1440aaccatgtcttgattgctcctttaagtcgtggttatactcacattaactcttctgatgtg1500gaggatcattctgtcattaatccccaatactactctcatcctatggatattgatgtccat1560atcgcttccactaaacttgctcgcgaaatcatcactgcctctcccggtcttggtgacatt1620aacagtggcgaaatcgaacccggtatgaatattacttctgacgacgacgttagaaaatgg1680ttgagtaataatgtccgttctgactggcatcctgttggtacttgtgctatgcttcccaag1740gaattaggtggtgttgtcagccccgctctcatggtttacggcacttccaacttgcgtgtt1800gttgatgcttcgattatgcccctcgaagtctcttctcatttgatgcaacccacctacggt1860attgctgagaaggctgctgacattattaagaatttctacaagactcaacacaagaaccaa1920aattag1926<210>13<211>635<212>prt<213>冻土毛霉(mucorhiemalis)<400>13metlysileservalalailevalthrilealaalaalaphealaser151015phealaasnalaglnlysthralathrserasnthrtyrasptyrval202530ilevalglyglyglyvalglyglyleualaleualaserargleuser354045gluasplysservalthrvalalavalleuglualaglyproasnala505560aspgluglnphevalvaltyralaproglymettyrglyglnalaval65707580glythraspleucysproleuargprothrvalproglnglualamet859095asnasnargthrleuthrilealathrglylysleuleuglyglygly100105110seralaileasnglyleuvaltrpthrargglyalaleulysaspphe115120125aspalatrpglugluleuglyasnproglytrpasnglyargthrmet130135140phelystyrphelyslysvalgluargphehisproprothrlysala145150155160glnvalglntyrglyalathrtyrglnlysglyvalhisglylysasn165170175glyargileaspileserpheproglupheglnpheproglnserala180185190asntrpasnalaserleualathrleuaspphethrhisglnglnasp195200205leuleuasnglyserleuhisglytyrserthrthrproasnthrleu210215220aspprolysthralaargargvalaspsertyrthrglytyrileala225230235240prophevalserarglysasnleuphevalleualaasnhisthrval245250255serargileglnphelysprolysasnglythrgluleuleulysala260265270valglyvalglutrptyrthrthrglyaspasnserasnlysglnthr275280285ilelysalaargarggluvalilevalserserglyserileglyser290295300prolysleuleugluileserglyileglyasnlysaspilevalthr305310315320alaalaglyvalglnserleuileaspleuproglyvalglyserasn325330335metglnasphisvalhisalavalthrvalserthrthrasnilethr340345350glyphethrthraspservalpheglnasngluthrleualagluglu355360365glnargglnglntyrtyrasnasnlysthrglyiletrpthrthrthr370375380proasnasnleuglytyrproserproserglnleupheaspglythr385390395400serphegluserglyglnalaphealaasnargileargasnserthr405410415aspglntrpalaglutyrtyralaserthrasnalathrasnileglu420425430leuleulyslysglntyralailevalalaserargtyrglugluasn435440445tyrleuserproilegluileasnphethrproglytyrglyglythr450455460thraspvalaspleulysasnasnlystyrglnthrvalasnhisval465470475480leuilealaproleuserargglytyrthrhisileasnserserasn485490495ilegluaspprovalvalileasnproglntyrtyrthrhispromet500505510aspvalaspvalhisilealaserthrlysleualaargargileleu515520525glyalagluproglyleualaserileasnserglygluileglnpro530535540glyserasnilethrseraspgluaspvallysglntrpleualaasp545550555560asnvalargserasptrphisprovalglythrcysalametleupro565570575arggluleuglyglyvalvalaspproasnleuleuvaltyrglythr580585590alaasnleuargvalvalaspalaserilemetproleugluileser595600605serhisleumetglnprothrtyrglyvalalaglulysalaalaasp610615620ileilelysmetserarglysasnasnasnasn625630635<210>14<211>1908<212>dna<213>冻土毛霉(mucorhiemalis)<400>14atgaaaatctcagtagccattgtcactatcgccgccgccttcgcttccttcgccaacgcc60caaaagaccgcaaccagtaatacctatgattatgtaattgttggtggtggtgttggtggt120ttggcattagcctctagattgtcagaagataaatctgtcacagtagctgttttagaagca180ggtcctaatgccgacgaacaatttgttgtctatgccccaggcatgtacggtcaagctgtt240ggtaccgatttgtgtccattaagacctactgtcccacaagaagctatgaataacagaaca300ttgaccatagcaaccggtaaattgttaggtggtggttcagctatcaatggtttggtttgg360actagaggtgcattaaaggattttgacgcctgggaagaattaggtaatccaggttggaac420ggtagaactatgttcaagtacttcaaaaaggttgaaagattccatccacctacaaaggct480caagtccaatatggtgcaacctaccaaaaaggtgtacacggtaaaaatggtagaatcgat540atttcttttcctgaatttcaattcccacaatctgctaattggaacgcctcattggctacc600ttagatttcactcatcaacaagacttgttaaatggttccttgcacggttatagtactaca660cctaacacattagatccaaaaaccgccagaagagttgactcctatacaggttacattgct720cctttcgttagtagaaagaatttgttcgtcttagcaaaccatactgtatctagaatacaa780ttcaaaccaaagaatggtacagaattgttgaaggctgtcggtgtagaatggtacaccact840ggtgacaactcaaacaaacaaacaattaaggcaagaagagaagttatcgtctcttcaggt900tccattggtagtcctaaattgttggaaatatccggtatcggtaataaggatatcgtaact960gctgcaggtgttcaatctttgattgatttgccaggtgtaggttcaaacatgcaagaccat1020gttcacgctgtaactgtttccacaaccaacataacaggttttactacagatagtgttttc1080caaaacgaaacattggcagaagaacaaagacaacaatactacaacaacaaaaccggtatc1140tggaccactacaccaaataacttgggttatccatcccctagtcaattatttgatggtaca1200tctttcgaatcaggtcaagcatttgcaaacagaattagaaactctaccgaccaatgggct1260gaatattacgcatcaactaatgccacaaacatcgaattgttgaagaaacaatacgcaatc1320gttgcctccagatacgaagaaaactacttgagtcctatcgaaatcaacttcactccaggt1380tatggtggtaccactgatgttgatttgaaaaataacaagtaccaaactgttaatcatgtc1440ttgatcgctcctttatcaagaggttatacacacatcaattccagtaacatagaagatcct1500gtagttataaatccacaatactacacccatccaatggatgtcgacgtacacattgcttct1560actaaattggcaagaagaatattaggtgccgaacctggtttggcttccataaatagtggt1620gaaatccaaccaggttctaacattacatcagatgaagacgttaagcaatggttagcagat1680aatgttagatctgactggcatcctgtcggtacatgcgccatgttgccaagagaattaggt1740ggtgtcgtagatccaaatttgttggtttacggtactgcaaacttaagagttgtcgacgcc1800tctataatgcctttggaaatctcttcacatttgatgcaaccaacttacggtgttgctgaa1860aaagccgctgatattattaagatgtctagaaagaataacaataactaa1908当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1