水稻基因ORYsa;SQD1的基因工程应用的制作方法
本发明涉及植物基因工程技术领域,涉及水稻基因orysa;sqd1的基因工程应用。
背景技术:
水稻在全世界都有广泛种植,是亚洲最重要的粮食作物,更是我国主要的粮食作物之一,我国水稻种植面积占全球谷类作物种植面积的1/3,水稻产量占全国粮食总产的50%,是保证我国粮食安全的最重要作物(胡培松等,2002)。
磷是植物生长所需的大量营养元素之一,也是许多土壤中十分容易缺乏的营养元素之一。尽管土壤中的总磷含量往往很高,但磷素利用率才是限制植物生长与生产力的关键因素。这种矛盾的出现是因为土壤溶液中的可溶性磷浓度平均值约为1μm,很少超过10μm(bieleski,1973),远远满足不了植物的生长需要,使其成为植物生长的一大限制性因子。因此植物能否高效利用土壤中少量的可溶性磷对植物生长有着至关重要的影响。
长期以来,人们对磷这一不可再生资源的不断开采和利用,使得磷的可利用性将在2030年到达最大峰值。这将造成部分发展中国家在农业方面更大的经济投入。另一方面,在缓解农作物缺磷的同时,磷肥的不科学使用导致土壤质量的下降,虽然与过去相比,农业土壤中磷的绝对量已不再缺乏,但可利用效率很低。高效利用磷的作物新品种创制可以减少磷肥施用,提高磷素利用率,对农业生产与生态环境都具有具有十分重大的意义(wuetal.,2013;lopez-arredondoetal.,2014)。
在之前的研究中,拟南芥中的硫代异鼠李糖和硫代异鼠李糖甘油二酯合酶基因sqd1为受缺磷诱导的基因,参与到缺磷条件下磷脂与硫脂间动态平衡的调控过程,但并未公开该基因具有促进拟南芥生长的功能。
技术实现要素:
本发明的目的在于提供水稻缺磷响应基因ossqd1的基因工程应用。
本发明的目的可通过以下技术方案实现:
本发明提供了水稻缺磷响应基因ossqd1的基因工程应用,其基因序列登录号为os05g0387200。为研究ossqd1的功能,需要确定并克隆获得ossqd1准确的序列。通过根据数据库提供序列进行rt-pcr,克隆得到了ossqd1完整的编码区序列,编码区核苷酸序列如seqidno.1所示,编码的氨基酸序列如seqidno.2所示。ossqd1基因共含有2个外显子,1个内含子,编码区全长1440bp,编码479个氨基酸。根据克隆到的序列,我们构建了ossqd1的超表达转载体,并转化水稻,使其表达量上调。
将获得的纯合体株系进行水培实验,测定苗期不同部位总磷浓度,可提取磷浓度等生理指标。
将获得的纯合体株系进行盆栽实验,测定于成熟期分部位采样,测定各部位磷浓度差异。采样部位包括:倒一叶、倒二叶、叶鞘、茎、穗柄和种子。
上述基因orysa;sqd1的基因工程应用,是旨在提高水稻磷素吸收与利用,以及提高水稻对环境中磷素胁迫的耐受能力。
有益效果
水稻作为单子叶模式植物与双子叶模式植物拟南芥磷的响应机制都有许多不同。本发明基因orysa;sqd1的功能为单子叶植物中的首次报道。我们通过在多个基因组数据库中的检索比对,确定了水稻中的sqd1同源目标基因,并通过pcr的手段克隆获得了水稻中ossqd1基因的编码区完整序列。我们通过分子生物学方法,鉴定了ossqd1基因的表达模式和亚细胞定位特征。构建了sqd1-ox的载体,过量sqd1的表达,通过一系列生理实验的鉴定发现:ossqd1过量表达增加了种子的宽度的同时,提高了水稻产量;与此同时,在提高土壤中磷素吸收利用效率方面起着重要作用。还提高了水稻营养生长期缺磷条件下可溶性磷含量以及生殖生长期茎、穗以及种子的全磷含量,以及显著提高了水稻外部感受缺磷信号的敏感度,提高水稻中磷素再分配的效率,为提高植物耐低磷能力、养分利用效率,为培育适用于磷贫瘠土壤的水稻新品种提供了保障。
附图说明
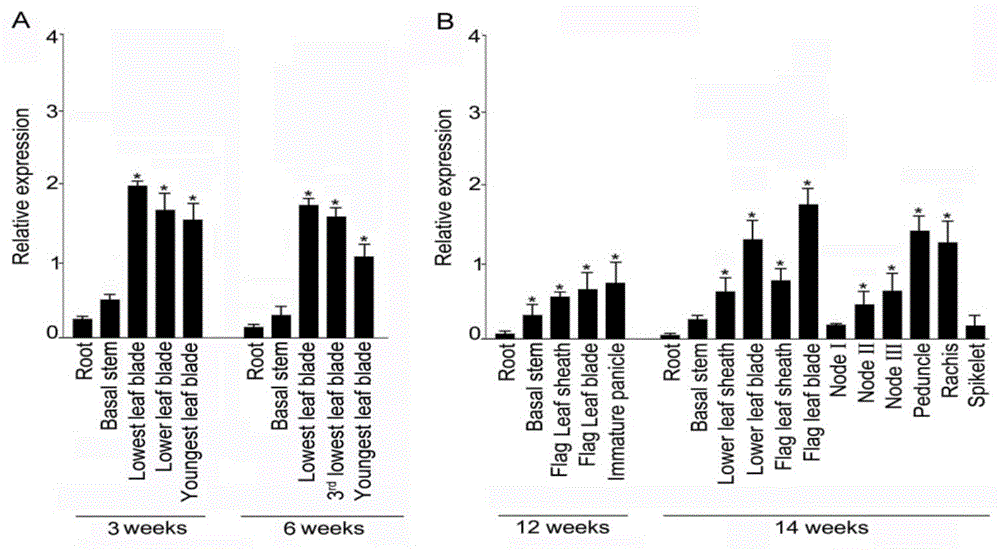
图1:水稻基因orysa;ossqd1在不同组织部位的表达模式。
图2:水稻基因orysa;ossqd1在不同缺磷时间点的表达模式。
图3:野生型和ossqd1超表达材料完熟期的表型。
图4:盆栽实验中,野生型与ossqd1超表达材料的种子形态与千粒重。
图5:正常供磷和缺磷条件下野生型与超表达材料的表型和生物量。
图6:不同磷处理条件下野生型和ossqd1-ox材料于营养生长阶段不同部位的全磷和可溶性磷含量。
图7:成熟期正常供磷和缺磷条件下野生型和ossqd1-ox株系不同部位中的全磷含量。
图8:正常供磷和缺磷条件下野生型和ossqd1-ox材料地上部和根部pht1家族成员的表达。
图9:ptck303质粒图谱。
具体实施方式
实施例1、基因序列的获得
atsqd1是一个定位在第五条染色体上,编码合成硫代异鼠李糖甘油二酯合酶sqd1的基因。sqd1酶本质为短链脱氢酶超家族(short-chaindehydrogenasesuperfamily;sdr)中的尿苷二磷酸硫代异鼠李糖合酶家族(udp-sulfoquinovosesynthasefamily;sdr52e)中的一员,具有nad+结合位点,可以与铁氧还原蛋白谷氨酰胺合酶结合,并接受从外界传递而来的亚硫酸根(so32-)。根据拟南芥sqd1基因序列在ncbi进行同源搜索,在ncbi数据库中获得水稻中同源基因ossqd1,基因号为(同rab_db)os05g0387200。通过blast和genomescan分析表明:ossqd1位于水稻第五条染色体上,具体位置为chr5:18738602..18741791。水稻sqd1基因含有2个外显子和1个内含子,基因的蛋白编码区为1440bp,编码479个氨基酸。
实施例2、orysa;sqd1基因的表达模式鉴定
2.1、总rna的提取和转录合成cdna第一链
选用水稻品种“日本晴”,待水稻幼苗长至10天后,完全营养液培养1周后采样(倒一叶,倒二叶,倒四叶,倒二叶叶鞘,倒四叶叶鞘,根茎结合部,根系根尖,根系伸长区和成熟区)。水稻苗龄至30天移栽至盆砵中进行盆栽处理,苗龄60天时进行采样(旗叶,倒三叶,旗叶叶鞘,倒三叶叶鞘,杆,茎结和穗柄)。分别采用trizol试剂抽提总rna,用琼脂糖凝胶电泳鉴定总rna质量,然后在分光光度计上测定rna含量。以获得的总rna为模版,经过逆转录获得水稻cdna第一链,供后续的实验使用。cdna第一链的合成步骤:用depc水处理过的pcr管(300μl),加入总rna5μg,oligodt1μl(25ng/μl),dntp(10mmol/l)2μl,65℃水浴5min,迅速置于冰上冷却,加5×反应缓冲液4μl,m-mlv逆转录酶(200u/μl)1μl,rnase抑制剂0.5μl,depc水至总体积为20μl(以上均在冰上操作)。稍离心后置于42℃水浴1h,70℃水浴10min,然后置于冰上迅速冷却。所得产物即为cdnas,置于-20℃保存。
2.2orysa;sqd1基因的组织特异性表达模式鉴定
以步骤2.1获得的“日本晴”cdna为模板,根据水稻ossqd1基因的3’utr部分的序列,设计以下ossqd1基因特异引物p1、p2扩增长度为117bp片段长度鉴定orysa;ossqd1基因的表达模式。
p1ctactacaacgccaagca(seqidno.3)
p2accagcctcctcttagac(seqidno.4)
pcr具体步骤为:以步骤1获得的cdna为模板,使用abisteponeplus(购自abi生物公司)和sybrpremixextaqkit(购自takara公司)进行基因定量pcr分析。根据takara试剂说明使用20μl的反应体系:sybrpremixextaq10μl,上下游引物各0.4μl,cdna模板2μl,roxreferencedye(50x)0.4μl,ddh2o6.8μl。反应条件为:40个循环,95℃1min,95℃5s,60℃30s,95℃15s,60℃1min,95℃5s。定量结果根据2-δct方法计算分析鉴定ossqd1基因的时空表达模式,结果见图1。委托南京金斯瑞生物技术公司测序确定序列为orysa;ossqd1片段。
图1可以看出,在水稻营养生长阶段,生育期第3周和第6周ossqd1的相对表达量均是地上部高于根部,其中第3周不同叶片(第一、二和最新展开叶)的相对表达量没有明显差异,根茎结合处和根部表达量明显低于叶片(图1a);第6周中ossqd1同样在叶片中的表达量最高,其中老叶(第一叶和第三叶)表达量明显高于最新叶(图1a)。在生殖生长阶段,生育期第12周ossqd1主要是在根茎结合处、叶鞘、叶片以及未成熟的花穗中表达,而在根中的表达量比较低(图1b);生育期第14周,ossqd1在叶片、花梗和花轴中的表达量最高,除此之外,叶鞘、茎节次之,根系和根茎结合处的表达量最低(图1b)。
2.3orysa;sqd1基因的响应缺磷的表达模式鉴定
为了了解ossqd1表达水平对缺磷响应的敏感程度,设置不同的缺磷时间点(1天、3天、5天、7天和7天后回补磷1天),检测其相对表达量。发现ossqd1无论是在根部还是地上部,其表达量在缺磷处理1天时变化并不明显,从第三天开始ossqd1的相对表达量出现了显著的上调,且上调幅度随着处理时间的推移明显增加,处理7天时地上部ossqd1表达量上调了约5.5倍,根部约上调6倍(图2)。缺磷7天后回补磷素1天后,ossqd1表达量在地上部和根部的表达量又下降至正常供磷时的水平(图2)。这一结果说明ossqd1对磷素的供应水平较为敏感,并响应短期缺磷信号。
实施例3、orysa;sqd1超表达材料的获得与功能鉴定
3.1ossqd1超表达载体的构建
为了构建ossqd1超表达材料,我们选择本实验室常用载体ptck303作为表达载体(图9)。根据ncbi网站公布的ossqd1的cdna序列,用dnaman对其进行限制性内切酶分析,并选择cdna中没有且载体多克隆酶切位点具备的kpni和spei,可将目的片段直接与ptck303载体进行酶切和酶连操作。引物分别如表(表1)所示。以日本晴叶片cdna为模板,用高保真pcrmix进行pcr扩增得到ossqd1的全长cdna序列片段。纯化并回收pcr产物,连入到克隆载体peasy-blunt。经测序确定序列与日本晴ossqd1序列无差异后,以kpni和spei内切酶酶切重组的克隆载体,获得的酶切片段经纯化和回收后,连接至表达载体,并转化入大肠杆菌dh5α感受态细胞。提取阳性单克隆质粒,经酶切验证及测序确保正确后,将重组质粒经电转化导入农杆菌eha105中。
表1ossqd1过表达载体构建所用引物
3.2超表达转基因植株的获得和功能鉴定
将步骤1获得的转有表达载体的农杆菌,进一步转化至水稻(采用根癌农杆菌介导方法将构建的表达载体转入水稻日本晴品种)。诱导水稻成熟胚愈伤,将长到一定大小的水稻愈伤组织挑出,放入农杆菌悬浮液侵染5min(愈伤量没过50ml离心管锥形部位即可,不停的摇动);将愈伤组织取出,置于无菌的滤纸上沥干30-40分钟;愈伤组织置于共培养基上,28℃暗培养2.5天。然后愈伤转入含250mg/l羧苄青霉素(car)和50mg/l潮霉素的选择培养基上进行筛选。挑取颜色鲜黄的抗性愈伤移入装有分化培养基的培养皿或分化罐中,放入恒温培养室分化成苗。再放入生根培养基中壮苗一到两周,即获得转基因植株。
3.3ossqd1超表达影响了水稻产量
通过对超表达株系(ox1、ox2和ox3)生育期表型鉴定及穗形态的观察(图3),发现在水稻完熟期,超表达与野生型的表型存在差异(图3a)。ox1、ox2和ox3的株高均明显低于野生型(图3b),分蘖数则是明显比野生型的多(图3c)。对穗形态的观察发现,ossqd1超表达材料与野生型穗子间的表型无明显差异(图3d),且两者间的结实率也无明显差异(图3e)。
3.4ossqd1超表达影响了水稻种子发育
我们鉴定了野生型与超表达材料的种子表型(图4a),超表达ox1、ox2和ox3的种子宽度明显比野生型的宽(图4b),且千粒重也明显高于野生型(图4c)。这一结果说明,ossqd1的超表达可以增加种子宽度,进而影响种子重量,可知ossqd1在影响种子发育上为正调控。
3.5ossqd1对水稻不同磷素处理下生长发育的影响
为了探究ossqd1营养生长阶段在正常供磷(+p)和缺磷(-p)条件下的功能,我们对野生型和超表达株系长至10天的幼苗分别进行+p(200μm)和-p(10μm)水培处理2周。结果显示,超表达株系的根系与野生型相比变得更加发达(图5a和b)。超表达的株高与野生型差异并不明显(图5c),根长有明显差异,其中+p条件下超表达上升约13.8%,-p条件下超表达约上升17.5%(图5d)。超表达生物量与野生型相比,在地上部和根部均有明显上升(图5e和f)。
3.6ossqd1超表达提高了水稻缺磷条件下可溶性磷含量
为了验证ossqd1对水稻营养生长阶段体内磷含量的影响,超表达材料中的磷含量在不同处理下也与野生型略有差异(图6)。在+p条件下,超表达中的可溶性磷和全磷含量都没有变化,但在-p条件下,与野生型相比,超表达中的可溶性磷含量明显上升,而全磷含量并无明显变化(图6)。这一结果说明-p条件下ossqd1的超表达可能促进了体内有机磷转变为无机磷的过程,才导致在体内总磷没有变化的前提下,可溶性磷出现明显上升。综合以上结果,推测ossqd1主要是在-p条件下对水稻地上部和根部磷浓度产生影响,并可能促进体内有机磷向无机磷的转变。
3.7ossqd1影响水稻成熟期磷素再分配
通过对+p和-p条件下处于成熟期的野生型和ossqd1超表达材料(ox1、ox2和ox3)新叶(第六叶)、老叶(第二叶)、茎、旗叶叶鞘、穗以及种子中磷含量的测定,发现+p处理下的新叶,老叶和旗叶叶鞘中磷浓度均没有差异,而超表达材料茎中的磷浓度均高于野生型(图7);-p条件下超表达材料老叶中磷浓度下降,新叶和旗叶叶鞘中的磷浓度没有变化,而茎秆、穗以及种子中的磷浓度明显高于野生型(图7),这些结果说明ossqd1超表达促进繁育器官磷素积累,且-p条件下抑制老叶中磷素积累。
3.8ossqd1过量表达提高了水稻对环境缺磷胁迫的响应
为了探究ossqd1的过量表达对水稻磷酸盐转运蛋白pht1家族成员的影响,通过荧光定量pcr的手段,我们检测了pht1家族成员在野生型和ossqd1-ox材料(ossqd1-ox1、ox2和ox3)在正常供磷(+p)和缺磷(-p)地上部以及根部的相对表达量(图8)。结果发现,+p处理的地上部中pt基因的相对表达量与野生型相比无明显差异,超表达材料(ossqd1-ox1、ox2和ox3)-p处理地上部的ospt1和ospt4的相对表达量与野生型相比明显上调(图8);与野生型相比,+p处理的超表达材料根部ospt1和ospt2的相对表达量明显上升,-p处理下的超表达根部ospt2、4和6均出现了不同程度的上调(图8)。由此可见ossqd1的过表达对pht1家族基因的表达整体呈现促进的趋势,且促进pht1家族基因表达的作用在-p条件下更加显著。这一结果也部分解释了为何-p条件下的超表达中磷含量上升。
综上所述,本发明人提供的orysa;ossqd1的工程应用为水稻中首次报道。orysa;ossqd1的超表达基因导入植物,提高水稻产量的同时增加了植物磷素利用效率和吸收速率,为培育高产、高磷素吸收及其体内磷素高效再分配的水稻新品种提供了保障。为了简单起见,本发明中orysa;ossqd1有时标注为ossqd1。
序列表
<110>南京农业大学
<120>水稻基因orysa;sqd1的基因工程应用
<160>6
<170>siposequencelisting1.0
<210>1
<211>1440
<212>dna
<213>水稻(oryzasativa)
<400>1
atgaaaatggcacatatggtaactaactgtagtttcagcccttcgcctgcagttaagacg60
tactcgaaatctcctggttattgctgcaatgtgacccaattccagagctcaaaatgttcc120
aatctagtgctcaaatcatgtactgcgacaagaccgaacaggccatttgtcgctcgtgct180
agcgctgctgtacaaggacagacacaaacaccccttactggaagtcagcaagcatctggg240
cactcatcctctaaacccaaaaaggtcatggttatcggcggagatggctactgcggctgg300
gcaaccgcacttcatctctccaataaaggatacgaggttgccattgttgacaatcttgtg360
cggcgcctttttgatcaccaacttggcctggattccctcacacccatagcctccatccag420
aatcgcatccgtcggtggaaggctctaaccggaaagacgattcagctctatgttggtgat480
atatgtgactttgatttcctttcagaagccttcaagtctttcgagccggattctgctgtc540
cactttggtgagcagagatcagcgccatattctatgattgatcgttcccgtgcggtattc600
actcagcataacaatgttatcggaactcttaatgtattatttgccattaaggagttcagt660
gaagaatgccatctggtcaaactaggaaccatgggtgagtatggaactccaaatattgac720
attgaagaagggttcatcactattacccacaatggaagaactgataccttgccttaccca780
aagcaagcgagctccttctaccatctaagcaaagtgcatgactcgcacaatatagcattt840
acatgcaaggcttggggtataagggccacggatcttaaccaaggtgttgtatatggagtg900
agaacagatgaaactgcaatgcatgaagaactatccaacaggtttgattatgatggcgtc960
tttgggacagcactgaataggttctgtgttcaggctgctgtaggtcatccacttacagtt1020
tatggaaaaggtggtcagacccgtggatatctggacatcagggatacagtgcaatgcgta1080
gagctcgcaatcgccaacccagccaaaccaggtgagttcagggtcttcaaccagttcacc1140
gagcagttctcggtcaacgagctggcaaagctggttacggccgccggtgcaaagcttggg1200
ctggaggtgcagaccaagtcggtgcccaacccgcgggtggaagcggaggaacactactac1260
aacgccaagcacaccaagctcatggagctcggcctggagccccacctgctgtcggactcg1320
ctcctcgactcgctgctcaacttcgccgtccagtacaaggacagggtcgacacggcgcag1380
atcatgcccagcgtgtcgtggaagaagatgggggcgaagccgaagacggtgtccgtctaa1440
<210>2
<211>479
<212>prt
<213>水稻(oryzasativa)
<400>2
metlysmetalahismetvalthrasncysserpheserproserpro
151015
alavallysthrtyrserlysserproglytyrcyscysasnvalthr
202530
glnpheglnserserlyscysserasnleuvalleulyssercysthr
354045
alathrargproasnargprophevalalaargalaseralaalaval
505560
glnglyglnthrglnthrproleuthrglyserglnglnalasergly
65707580
hisserserserlysprolyslysvalmetvalileglyglyaspgly
859095
tyrcysglytrpalathralaleuhisleuserasnlysglytyrglu
100105110
valalailevalaspasnleuvalargargleupheasphisglnleu
115120125
glyleuaspserleuthrproilealaserileglnasnargilearg
130135140
argtrplysalaleuthrglylysthrileglnleutyrvalglyasp
145150155160
ilecysasppheasppheleuserglualaphelysserpheglupro
165170175
aspseralavalhispheglygluglnargseralaprotyrsermet
180185190
ileaspargserargalavalphethrglnhisasnasnvalilegly
195200205
thrleuasnvalleuphealailelysgluphesergluglucyshis
210215220
leuvallysleuglythrmetglyglutyrglythrproasnileasp
225230235240
ileglugluglypheilethrilethrhisasnglyargthraspthr
245250255
leuprotyrprolysglnalaserserphetyrhisleuserlysval
260265270
hisaspserhisasnilealaphethrcyslysalatrpglyilearg
275280285
alathraspleuasnglnglyvalvaltyrglyvalargthraspglu
290295300
thralamethisglugluleuserasnargpheasptyraspglyval
305310315320
pheglythralaleuasnargphecysvalglnalaalavalglyhis
325330335
proleuthrvaltyrglylysglyglyglnthrargglytyrleuasp
340345350
ileargaspthrvalglncysvalgluleualailealaasnproala
355360365
lysproglyglupheargvalpheasnglnphethrgluglnpheser
370375380
valasngluleualalysleuvalthralaalaglyalalysleugly
385390395400
leugluvalglnthrlysservalproasnproargvalglualaglu
405410415
gluhistyrtyrasnalalyshisthrlysleumetgluleuglyleu
420425430
gluprohisleuleuseraspserleuleuaspserleuleuasnphe
435440445
alavalglntyrlysaspargvalaspthralaglnilemetproser
450455460
valsertrplyslysmetglyalalysprolysthrvalserval
465470475
<210>3
<211>18
<212>dna
<213>人工序列(artificialsequence)
<400>3
ctactacaacgccaagca18
<210>4
<211>18
<212>dna
<213>人工序列(artificialsequence)
<400>4
accagcctcctcttagac18
<210>5
<211>25
<212>dna
<213>人工序列(artificialsequence)
<400>5
tcggtaccatgaaaatggcacatat25
<210>6
<211>25
<212>dna
<213>人工序列(artificialsequence)
<400>6
tcactagttagacggacaccgtctt25
- 还没有人留言评论。精彩留言会获得点赞!