胰凝乳蛋白酶抑制剂变体及其用途的制作方法
序列表的引用本申请包含计算机可读形式的序列表。该计算机可读形式通过引用并入本文。本发明涉及大麦胰凝乳蛋白酶抑制剂2a(ci-2a)的变体及其在蛋白酶发酵生产中的用途。
背景技术:
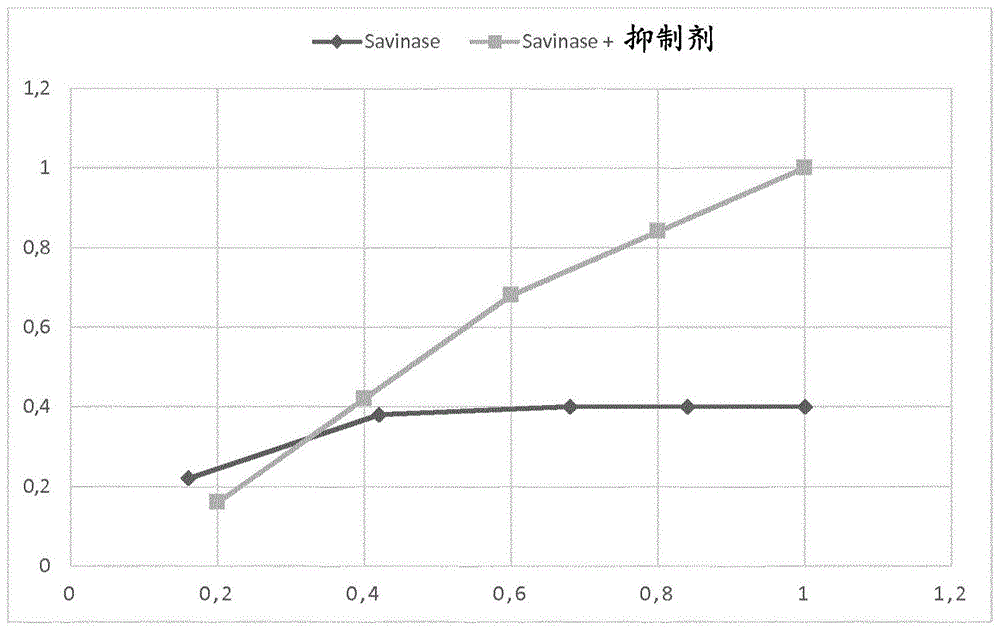
:蛋白酶成为酶工业的重要产品已经超过30年,并且蛋白酶已在许多不同的商业用途中使用,提及几种,例如用于洗涤剂,用于食品制造以及作为动物饲料组合物中的组分。蛋白酶具有降解蛋白质的能力,并且由于蛋白酶本身是蛋白质,这固有地意味着蛋白酶是自毁性的,因为一个活性蛋白酶分子将降解存在于同一组合物中的其他蛋白酶分子,从而降低该组合物的活性蛋白酶含量。这在本领域中称为自体蛋白水解(autoproteolysis),并且已采取各种措施来控制自体蛋白水解以改善生产期间的稳定性,并且改善中间和最终蛋白酶产物的保存期限。控制自体蛋白水解的一种措施是添加蛋白酶抑制剂。用添加的蛋白酶抑制剂来处理蛋白酶已描述于wo93/20175;wo93/13125;wo92/05239;wo93/17086(诺和诺德公司(novonordisk))中,或将蛋白酶与链霉菌ssi蛋白酶抑制剂共价融合描述于wo00/01831(宝洁公司(procter&gamble))和wo98/13483(宝洁公司)中。大麦的ci-2a胰凝乳蛋白酶抑制剂编码基因和携带由α前导序列翻译的基因的质粒描述于美国专利号5,674,833(1997)中。ci-2a(m59p)胰凝乳蛋白酶抑制剂描述于wo92/05239(诺和诺德公司)中。根据merops分类https://www.ebi.ac.uk/merops/cgi-bin/famsum?family=i13,ci-2a胰凝乳蛋白酶抑制剂属于i13家族抑制剂。已知该家族的几个成员,大多数成员衍生自植物,并且已经为一些成员阐明了结构。它抑制丝氨酸肽酶,这些肽酶主要属于丝氨酸蛋白酶的s1和s8家族。wo2002/016619披露了生产枯草杆菌蛋白酶的方法,其中将枯草杆菌蛋白酶(savinase)与ci-2a抑制剂共表达,导致产生savinase-ci-2a复合物。该申请还披露了savinase与ci-2a变体(m59p)的共表达,这导致产生savinase-ci-2a(m59p)复合物,该复合物与具有野生型ci-2a的、在洗涤剂组合物中解离从而释放savinase的复合物相反。使用抑制剂来控制蛋白酶的自体蛋白水解不可避免地导致在将蛋白酶用于其预期目的之前将抑制剂从蛋白酶解离的问题。因此,需要提供具有抑制蛋白酶活性并且防止自体蛋白水解的能力的新抑制剂,这些抑制剂可以以简单并且可控的方式从蛋白酶解离。技术实现要素:本发明提供了如下变体,这些变体具有蛋白酶抑制剂活性,与seqidno:1的成熟多肽具有至少60%序列同一性,并且在与seqidno:1的位置21、25、26、30、33、43、44和52对应的一个或多个位置处包含取代。本发明进一步提供编码此类变体和质粒的多核苷酸,包含此类多核苷酸的表达载体和宿主细胞,以及使用此类宿主细胞产生根据本发明的蛋白酶抑制剂的方法。本发明进一步涉及产生由s1或s8蛋白酶和根据本发明的蛋白酶抑制剂变体组成的复合物的方法,该方法包括以下步骤:a.提供表达s1或s8蛋白酶的微生物和表达蛋白酶抑制剂变体的微生物;b.在诱导该s1或s8蛋白酶和该蛋白酶抑制剂变体表达的条件下,培养表达该s1或s8蛋白酶的微生物和表达该蛋白酶抑制剂变体的微生物,从而形成由该s1或s8蛋白酶和该蛋白酶抑制剂变体组成的复合物;以及,任选地c.从发酵液中回收由该s1或s8蛋白酶和该蛋白酶抑制剂变体组成的复合物。最后,本发明涉及产生s1或s8蛋白酶的方法,该方法包括以下步骤:a.提供表达s1或s8蛋白酶的微生物和表达根据本发明的蛋白酶抑制剂变体的微生物;b.在诱导该s1或s8蛋白酶和该蛋白酶抑制剂变体表达的条件下,培养表达该s1或s8蛋白酶的微生物和表达该蛋白酶抑制剂变体的微生物,从而形成由该s1或s8蛋白酶和该蛋白酶抑制剂变体组成的复合物;c.调节ph至该s1或s8蛋白酶和该蛋白酶抑制剂变体的复合物解离并释放蛋白酶活性的低ph值,以及,任选地d.回收该s1或s8蛋白酶。附图说明图1显示savinase的发酵产量,将其与表达savinase的枯草芽孢杆菌(bacillussubtilis)宿主细胞以及表达savinase和ci-2a抑制剂的相同枯草芽孢杆菌宿主细胞的产量进行比较。更多细节,参见实例2。图2显示与ci-2a野生型相比,savinase从具有本发明变体的复合物的ph依赖性释放。更多细节,参见实例4。图3显示与ci-2a野生型相比,savinase变体从具有本发明变体的复合物的ph依赖性释放。更多细节,参见实例5。图4显示ty-145变体从具有本发明变体的复合物的ph依赖性释放。更多细节,参见实例6。序列简要说明seqidno:1是大麦胰凝乳蛋白酶抑制剂的氨基酸序列。seqidno:2是来自迟缓芽孢杆菌(bacilluslentus)的s8蛋白酶(也称为savinase)的氨基酸序列。seqidno:3:是来自解淀粉芽孢杆菌(bacillusamyloliquefaciens)的s8蛋白酶的氨基酸序列。seqidno:4是来自芽孢杆菌属物种(bacillussp.)的s8蛋白酶(也称为嘉士伯枯草杆菌蛋白酶(subtilisincarlsberg))的氨基酸序列。seqidno:5:是来自首次描述于wo92/17577中的芽孢杆菌属物种(bacillussp.)ty-145(ncimb40339)的s8蛋白酶的氨基酸序列。seqidno:6:是来自披露于wo01/58276中的拟诺卡氏菌属物种(nocardiopsissp.)nrrl18262的s1蛋白酶的氨基酸序列。定义丝氨酸蛋白酶:丝氨酸蛋白酶是催化肽键水解并且在活性位点处存在一个必需丝氨酸残基的酶(white,handler和smith,1973“principlesofbiochemistry[生物化学原理],”第五版,mcgraw-hillbookcompany[麦格劳-希尔图书公司],纽约州,第271-272页)。细菌丝氨酸蛋白酶的分子量范围为20,000至45,000道尔顿。它们可被二异丙基氟磷酸抑制。它们水解单末端酯并且在活性上类似于同样为丝氨酸蛋白酶的真核生物的胰凝乳蛋白酶。更狭义的术语,碱性蛋白酶(包括一个亚组)反映了一些丝氨酸蛋白酶从ph9.0至11.0的高最适ph(综述参见priest(1977)bacteriologicalrev.[细菌学综述]41711-753)。肽酶家族s1和s8的丝氨酸蛋白酶描述于biochem.j.[生物化学杂志]290:205-218(1993)和merops蛋白酶数据库发表版9.4(2011年1月31日)(www.merops.ac.uk)。该数据库描述于rawlings,n.d.,barrett,a.j.&bateman,a.(2010)‘merops:thepeptidasedatabase[merops:肽酶数据库]’,nucleicacidsres[核酸研究]38,d227-d233。暂时命名为枯草杆菌酶的丝氨酸蛋白酶的亚组已经由siezen等人,proteinengng.[蛋白质工程]4(1991)719-737以及siezen等人,proteinscience[蛋白质科学]6(1997)501-523提出。它们是通过对先前称为枯草杆菌蛋白酶样蛋白酶的丝氨酸蛋白酶的超过170种氨基酸序列进行同源性分析而定义的。枯草杆菌蛋白酶先前通常被定义为由革兰氏阳性细菌或真菌产生的丝氨酸蛋白酶,而现在根据siezen等人则为枯草杆菌酶的一个亚组。已鉴定了多种多样的枯草杆菌酶,并且已确定许多枯草杆菌酶的氨基酸序列。对于此类枯草杆菌酶及其氨基酸序列的更详细描述,参见siezen等人(1997)。枯草杆菌酶的一个亚组,i-s1或“真”枯草杆菌蛋白酶,包括“标准的”枯草杆菌蛋白酶,如枯草杆菌蛋白酶168(bss168)、枯草杆菌蛋白酶bpn’、嘉士伯枯草杆菌蛋白酶(subtilisincarlsberg)(诺和诺德公司)、以及枯草杆菌蛋白酶dy(bssdy)。siezen等人(见上文)识别了枯草杆菌酶的另一亚组,i-s2或高碱性枯草杆菌蛋白酶。亚组i-s2蛋白酶被描述为高碱性枯草杆菌蛋白酶,并且包括多种酶,如枯草杆菌蛋白酶pb92(baalkp)(吉斯特-布劳卡德斯公司(gist-brocadesnv))、枯草杆菌蛋白酶309(诺和诺德公司)、枯草杆菌蛋白酶147(bls147)(诺和诺德公司)、以及碱性弹性蛋白酶yab(bseyab)。编码序列:术语“编码序列”意指多核苷酸,该多核苷酸直接规定了变体的氨基酸序列。编码序列的边界通常由可读框确定,该可读框以起始密码子(例如atg、gtg或ttg)开始并且以终止密码子(例如taa、tag或tga)结束。编码序列可为基因组dna、cdna、合成dna或其组合。控制序列:术语“控制序列”意指对于表达编码本发明的变体的多核苷酸所必需的核酸序列。每个控制序列对于编码该变体的多核苷酸来说可以是天然的(即,来自相同基因)或外源的(即,来自不同基因),或相对于彼此是天然的或外源的。此类控制序列包括但不限于前导序列、多腺苷酸化序列、前肽序列、启动子、信号肽序列、以及转录终止子。最少,控制序列包括启动子、以及转录和翻译终止信号。出于引入促进控制序列与编码变体的多核苷酸的编码区域连接的特异性限制位点的目的,控制序列可以提供有接头。表达:术语“表达”包括涉及变体产生的任何步骤,包括但不限于转录、转录后修饰、翻译、翻译后修饰、以及分泌。表达载体:术语“表达载体”意指直链或环状dna分子,该分子包含编码变体的多核苷酸并且可操作地连接至提供用于其表达的控制序列。宿主细胞:术语“宿主细胞”意指易于用包含本发明的多核苷酸的核酸构建体或表达载体进行转化、转染、转导等的任何细胞类型。术语“宿主细胞”涵盖由于复制期间出现的突变而与亲本细胞不完全相同的任何亲本细胞子代。成熟多肽:术语“成熟多肽”意指在翻译和任何翻译后修饰如n-末端加工、c-末端截短、糖基化作用、磷酸化作用等之后处于其最终形式的多肽。在一方面,基于成熟多肽的n-末端测序,该成熟多肽是seqidno:1的氨基酸18或19至83。本领域描述了氨基酸n18和l19均被发现为n-末端氨基酸,因此据信宿主细胞可以产生具有seqidno:1的氨基酸序列的多肽的成熟多肽,其中seqidno:1具有n18和l19之一作为n-末端氨基酸,或宿主细胞可以根据生长条件和具体使用的表达构建体产生这两者的混合物。核酸构建体:术语“核酸构建体”意指单链或双链的核酸分子,该核酸分子是从天然存在的基因中分离的,或以原本不存在于自然界中的方式被修饰成包含核酸的区段,或者是合成的,该核酸分子包含一个或多个控制序列。可操作地连接:术语“可操作地连接”意指如下构型,在该构型中,控制序列被放置在相对于多核苷酸的编码序列适当的位置处,使得该控制序列指导该编码序列的表达。序列同一性:两个氨基酸序列之间或两个核苷酸序列之间的关联度通过参数“序列同一性”来描述。出于本发明的目的,使用如在emboss软件包(emboss:欧洲分子生物学开放软件包(theeuropeanmolecularbiologyopensoftwaresuite),rice等人,2000,trendsgenet.[遗传学趋势]16:276-277)(优选5.0.0版本或更新版本)的尼德尔程序中所实施的尼德曼-翁施算法(needleman-wunschalgorithm)(needleman和wunsch,1970,j.mol.biol.[分子生物学杂志]48:443-453)来确定两个氨基酸序列之间的序列同一性。所使用的参数是空位开放罚分10、空位延伸罚分0.5和eblosum62(emboss的blosum62版)取代矩阵。将needle标记的“最长同一性”的输出(使用非简化选项(nobriefoption)获得)用作同一性百分比并且计算如下:(相同的残基x100)/(比对长度-比对中的空位总数)变体:术语“变体”意指具有蛋白酶抑制剂活性的、在一个或多个(例如,若干个)位置处包含改变(即,取代、插入和/或缺失)的多肽。取代意指用不同的氨基酸替代占据某一位置的氨基酸;缺失意指去除占据某一位置的氨基酸;而插入意指在邻接并且紧随占据某一位置的氨基酸之后添加氨基酸。本发明的这些变体具有seqidno:1的成熟多肽的至少20%,例如至少40%、至少50%、至少60%、至少70%、至少80%、至少90%、至少95%、或至少100%的蛋白酶抑制剂活性。变体命名惯例出于本发明的目的,在seqidno:1中披露的成熟多肽用于确定另一种蛋白酶抑制剂中相应的氨基酸残基。将另一种蛋白酶抑制剂的氨基酸序列与seqidno:1中披露的成熟多肽进行比对,并且基于该比对,使用如在emboss软件包(emboss:欧洲分子生物学开放软件套件(theeuropeanmolecularbiologyopensoftwaresuite),rice等人,2000,trendsgenet.[遗传学趋势]16:276-277)(优选5.0.0版或更新版本)的尼德尔程序中所实施的尼德尔曼-翁施算法(needleman和wunsch,1970,j.mol.biol.[分子生物学杂志]48:443-453)来确定与seqidno:1中所披露的成熟多肽中的任何氨基酸残基对应的氨基酸位置编号。所使用的参数是空位开放罚分10、空位延伸罚分0.5和eblosum62(emboss的blosum62版)取代矩阵。可以通过使用若干计算机程序,使用其相应默认参数比对多个多肽序列来确定在另一种蛋白酶抑制剂中的对应氨基酸残基的鉴定,这些计算机程序包括但不限于muscle(通过对数预期的多序列比较;版本3.5或更新版本;edgar,2004,nucleicacidsresearch[核酸研究]32:1792-1797),mafft(版本6.857或更新版本;katoh和kuma,2002,nucleicacidsresearch[核酸研究]30:3059-3066;katoh等人,2005,nucleicacidsresearch[核酸研究]33:511-518;katoh和toh,2007,bioinformatics[生物信息学]23:372-374;katoh等人,2009,methodsinmolecularbiology[分子生物学方法]537:39-64;katoh和toh,2010,bioinformatics[生物信息学]26:1899-1900),以及采用clustalw的embossemma(1.83或更新版本;thompson等人,1994,nucleicacidsresearch[核酸研究]22:4673-4680)。当其他蛋白酶抑制剂与seqidno:1的成熟多肽相背离使得传统的基于序列的比较不能检测其相互关系时(lindahl和elofsson,2000,j.mol.biol.[分子生物学杂志]295:613-615),可使用其他成对序列比较算法。在基于序列的搜索中较高的敏感度可使用搜索程序来获得,这些搜索程序利用多肽家族的概率表现(谱)来搜索数据库。例如,psi-blast程序通过迭代数据库搜索过程来产生多个谱,并且能够检测远距离同源物(atschul等人,1997,nucleicacidsres.[核酸研究]25:3389-3402)。如果多肽的家族或超家族具有在蛋白结构数据库中的一个或多个代表,可以实现甚至更高的敏感度。程序例如genthreader(jones,1999,j.mol.biol.[分子生物学杂志]287:797-815;mcguffin和jones,2003,bioinformatics[生物信息学]19:874-881)利用来自多种来源(psi-blast、二级结构预测、结构比对谱、以及溶剂化势)的信息作为预测查询序列的结构折叠的神经网络的输入。类似地,gough等人,2000,j.mol.biol.[分子生物学杂志]313:903-919的方法可用于将未知结构的序列与存在于scop数据库中的超家族模型进行比对。这些比对进而可以用于产生多肽的同源性模型,并且使用出于该目的而开发的多种工具可以评估此类模型的准确度。对于已知结构的蛋白质,若干工具和资源可用于检索并产生结构比对。例如,蛋白质的scop超家族已经在结构上进行比对,并且那些比对是可访问且可下载的。可以使用多种算法如距离比对矩阵(holm和sander,1998,proteins[蛋白质]33:88-96)或组合延伸(shindyalov和bourne,1998,proteinengineering[蛋白质工程]11:739-747)比对两种或更多种蛋白质结构,并且这些算法的实施可以另外用于查询具有感兴趣结构的结构数据库,以便发现可能的结构同源物(例如,holm和park,2000,bioinformatics[生物信息学]16:566-567)。在描述本发明的变体中,为了便于参考,对以下所述的命名法进行了改编。采用了已接受的iupac单字母或三字母的氨基酸缩写。取代。对于氨基酸取代,使用以下命名法:原始氨基酸、位置、取代的氨基酸。相应地,将在位置226处的苏氨酸被丙氨酸取代表示为“thr226ala”或“t226a”。多个突变通过加号(“+”)分开,例如“gly205arg+ser411phe”或“g205r+s411f”代表在位置205和位置411处的甘氨酸(g)和丝氨酸(s)分别被精氨酸(r)和苯丙氨酸(f)取代。缺失。对于氨基酸缺失,使用以下命名法:原始氨基酸、位置、*。相应地,将在位置195处的甘氨酸的缺失表示为“gly195*”或“g195*”。多个缺失通过加号(“+”)分开,例如“gly195*+ser411*”或“g195*+s411*”。插入。于氨基酸插入,使用以下命名法:原始氨基酸、位置、原始氨基酸、插入的氨基酸。相应地,将在位置195处的甘氨酸之后插入赖氨酸表示为“gly195glylys”或“g195gk”。多个氨基酸的插入被表示为[原始氨基酸、位置、原始氨基酸、插入的氨基酸#1、插入的氨基酸#2;等]。例如,将在位置195处的甘氨酸之后插入赖氨酸和丙氨酸表示为“gly195glylysala”或“g195gka”。在此类情况下,通过将小写字母添加至在所插入的一个或多个氨基酸残基之前的氨基酸残基的位置编号而对所插入的一个或多个氨基酸残基进行编号。在以上实例中,该序列因此会是:亲本:变体:195195195a195bgg-k-a多种改变。包含多种改变的变体由加号(“+”)分开,例如,“arg170tyr+gly195glu”或“r170y+g195e”代表在位置170和位置195处的精氨酸和甘氨酸分别被酪氨酸和谷氨酸取代。不同改变。在可于一个位置处引入不同改变的情况下,不同的改变由逗号分开,例如,“arg170tyr,glu”代表用酪氨酸或谷氨酸取代在位置170处的精氨酸。因此,“tyr167gly,ala+arg170gly,ala”表示以下变体:“tyr167gly+arg170gly”、“tyr167gly+arg170ala”、“tyr167ala+arg170gly”、和“tyr167ala+arg170ala”。具体实施方式本发明涉及如下蛋白酶抑制剂,该蛋白酶抑制剂能够在约6.0至约9.0范围内的ph下抑制s1或s8蛋白酶的蛋白酶活性,但是其中该抑制剂还具有与s1或s8抑制剂的ph依赖性结合,这意味着当ph降低至低于6.0的ph,例如4.0至6.0范围内的ph值、例如4.5至6.0范围内的ph值、例如4.5至5.5范围内的ph值时,s1和s8蛋白酶和该抑制剂的复合物解离,从而释放蛋白酶活性。优选地,这些抑制剂是多肽。变体本发明还涉及蛋白酶抑制剂变体,这些蛋白酶抑制剂变体在与seqidno:1的成熟多肽的位置21、25、26、30、33、43、44和52对应的一个或多个(例如若干个)位置处包含改变取代,其中该变体具有蛋白酶抑制活性。本发明还提供了蛋白酶抑制剂变体,这些蛋白酶抑制剂变体在与21、25、26、30、33、43、44和52对应的一个或多个(例如,若干个)位置处包含改变取代,其中该变体具有蛋白酶抑制剂活性。在一个实施例中,该变体与亲本蛋白酶抑制剂的氨基酸序列具有至少60%,例如至少65%、至少70%、至少75%、至少80%、至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、或至少99%、但小于100%序列同一性。在另一个实施例中,该变体与seqidno:1的成熟多肽具有至少60%,例如至少65%、至少70%、至少75%、至少80%、至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%,如至少96%、至少97%、至少98%、或至少99%但小于100%序列同一性。在一方面,在本发明的变体中的改变数目是1-20个,例如1-10个和1-5个,如1、2、3、4、5、6、7、8、9或10个改变。在另一方面,变体在与位置21、25、26、30、33、43、44和52对应的一个或多个(例如若干个)位置处包含改变取代。在另一方面,变体在与位置21、25、26、30、33、43、44和52的任何一个对应的两个位置处包含改变。在另一方面,变体在与位置21、25、26、30、33、43、44和52的任何一个对应的三个位置处包含改变。在另一方面,变体在与位置21、25、26、30、33、43、44和52的任何一个对应的四个位置处包含改变。在另一方面,变体在与位置21、25、26、30、33、43、44和52的任何一个对应的五个位置处包含改变。在另一方面,变体在与位置21、25、26、30、33、43、44和52的任何一个对应的六个位置处包含改变。在另一方面,变体在与位置21、25、26、30、33、43、44和52的任何一个对应的七个位置处包含改变。在另一方面,变体在与位置21、25、26、30、33、43、44和52的任何一个对应的每个位置处包含改变。在另一方面,该变体在与位置21对应的位置处包含取代,或由其组成。在另一方面,在与位置21对应的位置处的氨基酸被ala、arg、asn、asp、cys、gln、glu、gly、his、ile、leu、met、phe、pro、ser、thr、trp、tyr或val,优选被his取代。在另一方面,该变体包含seqidno:1的成熟多肽的取代k21h,或由其组成。在另一方面,该变体在与位置25对应的位置处包含取代,或由其组成。在另一方面,在与位置25对应的位置处的氨基酸被ala、arg、asn、asp、cys、gln、glu、gly、his、ile、leu、lys、met、phe、ser、thr、trp、tyr、或val,优选被ser取代。在另一方面,该变体包含seqidno:1的成熟多肽的取代p25s,或由其组成。在另一方面,该变体在与位置26对应的位置处包含取代,或由其组成。在另一方面,在与位置26对应的位置处的氨基酸被ala、arg、asn、asp、cys、gln、gly、his、ile、leu、lys、met、phe、pro、ser、thr、trp、tyr或val,优选被his取代。在另一方面,该变体包含seqidno:1的成熟多肽的取代e26h,或由其组成。在另一方面,该变体在与位置30对应的位置处包含取代,或由其组成。在另一方面,在与位置30对应的位置处的氨基酸被ala、arg、asn、asp、cys、gln、glu、gly、his、ile、leu、met、phe、pro、ser、thr、trp、tyr或val,优选被his取代。在另一方面,该变体包含seqidno:1的成熟多肽的取代k30h,或由其组成。在另一方面,该变体在与位置33对应的位置处包含取代,或由其组成。在另一方面,在与位置33对应的位置处的氨基酸被ala、arg、asn、asp、cys、gln、gly、his、ile、leu、lys、met、phe、pro、ser、thr、trp、tyr或val,优选被his取代。在另一方面,该变体包含seqidno:1的成熟多肽的取代e33h,或由其组成。在另一方面,该变体在与位置43对应的位置处包含取代,或由其组成。在另一方面,在与位置43对应的位置处的氨基酸被ala、arg、asn、asp、cys、gln、glu、gly、his、ile、leu、met、phe、pro、ser、thr、trp、tyr或val,优选被his取代。在另一方面,该变体包含seqidno:1的成熟多肽的取代k43h,或由其组成。在另一方面,该变体在与位置44对应的位置处包含取代,或由其组成。在另一方面,在与位置44对应的位置处的氨基酸被ala、arg、asn、asp、cys、gln、glu、gly、his、ile、leu、lys、met、phe、ser、thr、trp、tyr、或val,优选被ala取代。在另一方面,该变体包含seqidno:1的成熟多肽的取代p44a,或由其组成。在另一方面,该变体在与位置52对应的位置处包含取代,或由其组成。在另一方面,在与位置52对应的位置处的氨基酸被ala、arg、asn、asp、cys、gln、glu、gly、his、ile、leu、lys、met、phe、ser、thr、trp、tyr、或val,优选被ala取代。在另一方面,该变体包含seqidno:1的成熟多肽的取代p52a,或由其组成。在另一方面,该变体在与位置21和25对应的位置处包含改变,或由其组成,如以上所述的那些。在另一方面,该变体在与位置21和26对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置21和30对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置21和33对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置21和43对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置21和44对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置21和52对应的位置处包含改变,或由其组成,如以上所述的那些。在另一方面,该变体在与位置25和26对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置25和30对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置25和33对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置25和43对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置25和44对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置25和52对应的位置处包含改变,或由其组成,如以上所述的那些。在另一方面,该变体在与位置26和30对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置26和33对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置26和43对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置26和44对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置26和52对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置30和33对应的位置处包含改变或由其组成,如以上所述的那些。在另一方面,该变体在与位置30和43对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置30和44对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置30和52对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置33和43对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置33和44对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置33和52对应的位置处包含改变或由其组成,如以上所述的那些。在另一方面,该变体在与位置43和44对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置43和52对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置44和52对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置21、25和26对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置21、25和30对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置21、25和43对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置21、25和44对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置21、25和52对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置21、26和30对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置21、26和33对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置21、26和43对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置21、26和44对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置21、26和52对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置21、30和33对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置21、30和43对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置21、30和44对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置21、30和52对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置21、33和43对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置21、33和44对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置21、33和52对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置21、43和44对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置21、43和52对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置21、44和52对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置25、26和30对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置25、26和33对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置25、26和43对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置25、26和44对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置25、26和52对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置25、30和33对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置25、30和43对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置25、30和44对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置25、30和52对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置25、33和43对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置25、33和44对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置25、33和52对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置25、43和44对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置25、43和52对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置25、44和52对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置26、30和33对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置26、30和43对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置26、30和44对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置26、30和52对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置26、33和43对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置26、33和44对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置26、33和52对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置26、43和44对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置26、43和52对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置26、44和52对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置30、33和43对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置30、33和44对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置30、33和52对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置30、43和44对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置30、43和52对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置30、44和52对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置33、43和44对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置33、43和52对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置33、44和52对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体在与位置43、44和52对应的位置处包含改变,或由其组成,如上文描述的那些。在另一方面,该变体包含选自由以下组成的组的一个或多个(例如若干个)取代或由其组成:k21h、p25s、e26h、k30h、e33h、k43h、p44a和p52a。在另一方面,该变体包含seqidno:2的成熟多肽的取代k21h+e26h或由其组成,或者包含与seqidno:1的成熟多肽具有至少65%、至少70%、至少75%、至少80%、至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%同一性的具有蛋白酶抑制剂活性的多肽或由其组成,并且另外与seqidno:1的成熟抑制剂相比,该变体在较高ph下与蛋白酶解离。在另一方面,该变体包含seqidno:2的成熟多肽的取代k21h+k30h或由其组成,或者包含与seqidno:1的成熟多肽具有至少65%、至少70%、至少75%、至少80%、至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%同一性的具有蛋白酶抑制剂活性的多肽或由其组成,并且另外与seqidno:1的成熟抑制剂相比,该变体在较高ph下与蛋白酶解离。在另一方面,该变体包含seqidno:1的成熟多肽的取代k21h+e33h或由其组成,或者包含与seqidno:1的成熟多肽具有至少65%、至少70%、至少75%、至少80%、至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%同一性的具有蛋白酶抑制剂活性的多肽或由其组成,并且另外与seqidno:1的成熟抑制剂相比,该变体在较高ph下与蛋白酶解离。在另一方面,该变体包含seqidno:1的成熟多肽的取代k21h+k43h或由其组成,或者包含与seqidno:1的成熟多肽具有至少65%、至少70%、至少75%、至少80%、至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%同一性的具有蛋白酶抑制剂活性的多肽或由其组成,并且另外与seqidno:1的成熟抑制剂相比,该变体在较高ph下与蛋白酶解离。在另一方面,该变体包含seqidno:1的成熟多肽的取代e26h+k30h或由其组成,或者包含与seqidno:1的成熟多肽具有至少65%、至少70%、至少75%、至少80%、至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%同一性的具有蛋白酶抑制剂活性的多肽或由其组成,并且另外与seqidno:1的成熟抑制剂相比,该变体在较高ph下与蛋白酶解离。在另一方面,该变体包含seqidno:1的成熟多肽的取代e26h+e33h或由其组成,或者包含与seqidno:1的成熟多肽具有至少65%、至少70%、至少75%、至少80%、至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%同一性的具有蛋白酶抑制剂活性的多肽或由其组成,并且另外与seqidno:1的成熟抑制剂相比,该变体在较高ph下与蛋白酶解离。在另一方面,该变体包含seqidno:1的成熟多肽的取代e26h+k43h或由其组成,或者包含与seqidno:1的成熟多肽具有至少65%、至少70%、至少75%、至少80%、至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%同一性的具有蛋白酶抑制剂活性的多肽或由其组成,并且另外与seqidno:1的成熟抑制剂相比,该变体在较高ph下与蛋白酶解离。在另一方面,该变体包含seqidno:1的成熟多肽的取代k30h+e33h或由其组成,或者包含与seqidno:1的成熟多肽具有至少65%、至少70%、至少75%、至少80%、至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%同一性的具有蛋白酶抑制剂活性的多肽或由其组成,并且另外与seqidno:1的成熟抑制剂相比,该变体在较高ph下与蛋白酶解离。在另一方面,该变体包含seqidno:1的成熟多肽的取代k30h+k43h或由其组成,或者包含与seqidno:1的成熟多肽具有至少65%、至少70%、至少75%、至少80%、至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%同一性的具有蛋白酶抑制剂活性的多肽或由其组成,并且另外与seqidno:1的成熟抑制剂相比,该变体在较高ph下与蛋白酶解离。在另一方面,该变体包含seqidno:1的成熟多肽的取代e33h+k43h或由其组成,或者包含与seqidno:1的成熟多肽具有至少65%、至少70%、至少75%、至少80%、至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%同一性的具有蛋白酶抑制剂活性的多肽或由其组成,并且另外与seqidno:1的成熟抑制剂相比,该变体在较高ph下与蛋白酶解离。在另一方面,该变体包含seqidno:1的成熟多肽的取代k21h+e26h+k30h或由其组成,或者包含与seqidno:1的成熟多肽具有至少65%、至少70%、至少75%、至少80%、至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%同一性的具有蛋白酶抑制剂活性的多肽或由其组成,并且另外与seqidno:1的成熟抑制剂相比,该变体在较高ph下与蛋白酶解离。在另一方面,该变体包含seqidno:1的成熟多肽的取代k21h+e26h+e33h或由其组成,或者包含与seqidno:1的成熟多肽具有至少65%、至少70%、至少75%、至少80%、至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%同一性的具有蛋白酶抑制剂活性的多肽或由其组成,并且另外与seqidno:1的成熟抑制剂相比,该变体在较高ph下与蛋白酶解离。在另一方面,该变体包含seqidno:1的成熟多肽的取代k21h+e26h+k43h或由其组成,或者包含与seqidno:1的成熟多肽具有至少65%、至少70%、至少75%、至少80%、至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%同一性的具有蛋白酶抑制剂活性的多肽或由其组成,并且另外与seqidno:1的成熟抑制剂相比,该变体在较高ph下与蛋白酶解离。在另一方面,该变体包含seqidno:1的成熟多肽的取代k21h+k30h+e33h或由其组成,或者包含与seqidno:1的成熟多肽具有至少65%、至少70%、至少75%、至少80%、至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%同一性的具有蛋白酶抑制剂活性的多肽或由其组成,并且另外与seqidno:1的成熟抑制剂相比,该变体在较高ph下与蛋白酶解离。在另一方面,该变体包含seqidno:1的成熟多肽的取代k21h+k30h+k43h或由其组成,或者包含与seqidno:1的成熟多肽具有至少65%、至少70%、至少75%、至少80%、至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%同一性的具有蛋白酶抑制剂活性的多肽或由其组成,并且另外与seqidno:1的成熟抑制剂相比,该变体在较高ph下与蛋白酶解离。在另一方面,该变体包含seqidno:1的成熟多肽的取代k21h+e33h+k43h或由其组成,或者包含与seqidno:1的成熟多肽具有至少65%、至少70%、至少75%、至少80%、至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%同一性的具有蛋白酶抑制剂活性的多肽或由其组成,并且另外与seqidno:1的成熟抑制剂相比,该变体在较高ph下与蛋白酶解离。在另一方面,该变体包含seqidno:1的成熟多肽的取代e26h+k30h+e33h或由其组成,或者包含与seqidno:1的成熟多肽具有至少65%、至少70%、至少75%、至少80%、至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%同一性的具有蛋白酶抑制剂活性的多肽或由其组成,并且另外与seqidno:1的成熟抑制剂相比,该变体在较高ph下与蛋白酶解离。在另一方面,该变体包含seqidno:1的成熟多肽的取代e26h+k30h+k43h或由其组成,或者包含与seqidno:1的成熟多肽具有至少65%、至少70%、至少75%、至少80%、至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%同一性的具有蛋白酶抑制剂活性的多肽或由其组成,并且另外与seqidno:1的成熟抑制剂相比,该变体在较高ph下与蛋白酶解离。在另一方面,该变体包含seqidno:1的成熟多肽的取代e26h+e33h+k43h或由其组成,或者包含与seqidno:1的成熟多肽具有至少65%、至少70%、至少75%、至少80%、至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%同一性的具有蛋白酶抑制剂活性的多肽或由其组成,并且另外与seqidno:1的成熟抑制剂相比,该变体在较高ph下与蛋白酶解离。在另一方面,该变体包含seqidno:1的成熟多肽的取代k30h+e33h+k43h或由其组成,或者包含与seqidno:1的成熟多肽具有至少65%、至少70%、至少75%、至少80%、至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%同一性的具有蛋白酶抑制剂活性的多肽或由其组成,并且另外与seqidno:1的成熟抑制剂相比,该变体在较高ph下与蛋白酶解离。在另一方面,该变体包含seqidno:1的成熟多肽的取代k21h+e26h+k30h+e33h或由其组成,或者包含与seqidno:1的成熟多肽具有至少65%、至少70%、至少75%、至少80%、至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%同一性的具有蛋白酶抑制剂活性的多肽或由其组成,并且另外与seqidno:1的成熟抑制剂相比,该变体在较高ph下与蛋白酶解离。在另一方面,该变体包含seqidno:1的成熟多肽的取代k21h+e26h+k30h+k43h或由其组成,或者包含与seqidno:1的成熟多肽具有至少65%、至少70%、至少75%、至少80%、至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%同一性的具有蛋白酶抑制剂活性的多肽或由其组成,并且另外与seqidno:1的成熟抑制剂相比,该变体在较高ph下与蛋白酶解离。在另一方面,该变体包含seqidno:1的成熟多肽的取代k21h+e26h+e33h+k43h或由其组成,或者包含与seqidno:1的成熟多肽具有至少65%、至少70%、至少75%、至少80%、至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%同一性的具有蛋白酶抑制剂活性的多肽或由其组成,并且另外与seqidno:1的成熟抑制剂相比,该变体在较高ph下与蛋白酶解离。在另一方面,该变体包含seqidno:1的成熟多肽的取代k21h+k30h+e33h+k43h或由其组成,或者包含与seqidno:1的成熟多肽具有至少65%、至少70%、至少75%、至少80%、至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%同一性的具有蛋白酶抑制剂活性的多肽或由其组成,并且另外与seqidno:1的成熟抑制剂相比,该变体在较高ph下与蛋白酶解离。在另一方面,该变体包含seqidno:1的成熟多肽的取代e26h+k30h+e33h+k43h或由其组成,或者包含与seqidno:1的成熟多肽具有至少65%、至少70%、至少75%、至少80%、至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%同一性的具有蛋白酶抑制剂活性的多肽或由其组成,并且另外与seqidno:1的成熟抑制剂相比,该变体在较高ph下与蛋白酶解离。在另一方面,该变体包含seqidno:1的成熟多肽的取代k21h+e26h+k30h+h33h+k43h,或由其组成。变体在一个或多个(例如,若干个)其他位置上可进一步包含一个或多个另外的改变。氨基酸改变可以具有微小性质,即,不会显著地影响蛋白质的折叠和/或活性的保守氨基酸取代或插入;典型地为1-10个氨基酸的小缺失;小的氨基末端或羧基末端延伸,如氨基末端的甲硫氨酸残基;多达20-25个残基的小接头肽;或小的延伸,其通过改变净电荷或另一功能(例如聚组氨酸段、抗原表位或结合结构域)来促进纯化。保守取代的实例是在下组之内:碱性氨基酸(精氨酸、赖氨酸及组氨酸)、酸性氨基酸(谷氨酸和天冬氨酸)、极性氨基酸(谷氨酰胺和天冬酰胺)、疏水性氨基酸(亮氨酸、异亮氨酸及缬氨酸)、芳香族氨基酸(苯丙氨酸、色氨酸及酪氨酸)及小氨基酸(甘氨酸、丙氨酸、丝氨酸、苏氨酸及甲硫氨酸)。通常不会改变比活性的氨基酸取代是本领域已知的并且例如由h.neurath和r.l.hill,1979,于theproteins[蛋白质],academicpress[学术出版社],纽约中描述。常见取代为ala/ser、val/ile、asp/glu、thr/ser、ala/gly、ala/thr、ser/asn、ala/val、ser/gly、tyr/phe、ala/pro、lys/arg、asp/asn、leu/ile、leu/val、ala/glu和asp/gly。可替代地,这些氨基酸改变具有这样一种性质:改变多肽的物理化学特性。例如,氨基酸改变可以提高多肽的热稳定性、改变底物特异性、改变最适ph,等等。可以根据本领域中已知的程序,如定点诱变或丙氨酸扫描诱变(cunningham和wells,1989,science[科学]244:1081-1085)来鉴定多肽中的必需氨基酸。在后一项技术中,在该分子中的每个残基处引入单个丙氨酸突变,并且对所得突变体分子的蛋白酶抑制剂活性进行测试以鉴定对于该分子的活性至关重要的氨基酸残基。还参见,hilton等人,1996,j.biol.chem.[生物化学杂志]271:4699-4708。酶或其他生物学相互作用的活性部位还可通过对结构的物理分析来确定,如由下述技术确定:核磁共振、晶体学(crystallography)、电子衍射、或光亲和标记,连同对推定的接触位点(contactsite)氨基酸进行突变。参见例如,devos等人,1992,science[科学]255:306-312;smith等人,1992,j.mol.biol.[分子生物学杂志]224:899-904;wlodaver等人,1992,febslett.[欧洲生化学会联合会快报]309:59-64。还可以从与相关多肽的比对来推断必需氨基酸的身份。这些变体可以由60至90个氨基酸,例如70至85和75至83个氨基酸组成。根据本发明的变体的优选实例包括如下变体,这些变体包含使用seqidno:1进行编号的以下取代:k43h+p44a+p52a;k21h+p25s+k30h;k21h+p25s+e26h+k30h+e33h;以及k30h+e33h+k43h+p44a+p52a;k21h+p25s+e26h+k30h+e33h+d42n+e45g;k21h+p25s+e26h+k30h+e33h+e45g;k21h+p25s+e26h+k30h+e33h+q47h;k21h+p25s+e26h+k30h+e33h+d42n+q47h;以及k21h+p25s+e26h+k30h+e33h+e45g+q47h;并且与seqidno:1具有至少60%序列同一性。其他实例包括如下变体,这些变体包含使用seqidno:1进行编号的以下取代:p25s+p44a+p52a+p80v;p25s+p44a+p52a+p80i;k21h+e26h+d41n;e33h;k43h+e45g,k43h+e45a;q47h;i49h;以及l73h。根据本发明的特别优选的变体包括如下变体,这些变体具有带有以下取代的seqidno:1的序列:k43h+p44a+p52a;k21h+p25s+k30h;k21h+p25s+e26h+k30h+e33h;以及k30h+e33h+k43h+p44a+p52a;k21h+p25s+e26h+k30h+e33h+d42n+e45g;k21h+p25s+e26h+k30h+e33h+e45g;k21h+p25s+e26h+k30h+e33h+q47h;k21h+p25s+e26h+k30h+e33h+d42n+q47h;以及k21h+p25s+e26h+k30h+e33h+e45g+q47h。本发明的变体具有当ph降低至低于6的ph值(例如,至4.5的ph)时,从由该变体和s8或s1蛋白酶(例如,具有seqidno:2的迟缓芽孢杆菌蛋白酶(也称为savinase))组成的复合物解离的能力。技术人员将理解,由s1或s8蛋白酶和本发明的变体组成的复合物解离时的确切ph值取决于s1或s8蛋白酶和本发明的特定变体两者的特定序列,但是对于由给定的s1或s8蛋白酶和本发明的变体组成的复合物,该复合物解离时的ph值比由相同的s1或s8蛋白酶和野生型ci-2a抑制剂组成的复合物解离时的ph值高至少0.4个ph单位,优选至少0.5个ph单位,优选至少0.6个ph单位,优选至少0.7个ph值单位,优选至少0.8个ph单位,优选至少0.9个ph单位,或优选至少1.0个ph单位。这可以通过以下过程方便地进行分析:a)通过混合等摩尔量的蛋白酶和蛋白酶抑制剂制备s8或s1蛋白酶和本发明的蛋白酶抑制剂变体的复合物;b)将a)中制备的复合物的等分试样在6至3.5范围内的各种ph值,例如ph6、5.5、5.0、4.5、4.0和3.5的缓冲溶液中在25°-50°范围内的温度下孵育;c)孵育60分钟后,使用标准蛋白酶测定法分析每个样品中的蛋白酶活性;以及d)使用野生型ci-2a作为参考重复步骤a)至c)。用于确定变体从蛋白酶解离的能力的优选测定法是以下过程:a)通过混合等摩尔量的savinase和蛋白酶抑制剂制备本发明给定的蛋白酶抑制剂变体和savinase的复合物;b)将a)中制备的复合物的等分试样在6至3.5范围内的各种ph值,例如ph6、5.5、5.0、4.5、4.0和3.5的缓冲溶液中在25°-50°范围内的温度下孵育;c)孵育60分钟后,使用标准蛋白酶测定法分析每个样品中的蛋白酶活性;以及d)使用野生型ci-2a作为参考重复步骤a)至c)。实例3中举例说明了该测定法。对于本发明的变体,可以看出,蛋白酶活性在比野生型ci-2a高至少0.5个ph单位处达到峰值,优选在4.0至4.5范围内的ph处达到峰值。当ph降低时,野生型ci-2a也可以被释放,但这需要在由于低ph而释放抑制剂或使其变性之前将ph降低至低得多的ph值,例如低于4.0或甚至低于3.5。因此,这些变体具有在中性至碱性区域的ph值(通常5.5至9.0范围内的ph值)下抑制s1和s8蛋白酶的能力,并且进一步地,这些变体可以通过将ph降低至5.5至4.0范围内的ph值容易地与s1和s8蛋白酶解离,从而释放s1和s8蛋白酶。这样做的好处是,可以在中性至碱性ph值下将本发明的变体添加到包含一种或多种s1和/或s8的组合物中,从而抑制蛋白酶的活性,从而保护组合物中的蛋白酶免受自体蛋白水解,并且保护组合物中的其他蛋白质免受蛋白酶降解,并且进一步地当需要时可以通过降低ph来释放ph活性。野生型ci-2a抑制剂还可以抑制s1和s8蛋白酶并保护其免受自体蛋白水解和蛋白酶降解,但是野生型ci-2sa不能像本发明的变体一样容易地从s1和s8蛋白酶中除去。野生型ci-2a要求ph比变体所需的ph单位降低至少0.5个ph单位。当ph降低与从s1和/或s8蛋白酶释放野生型ci-2a抑制剂所需的量同样多时,所存在的s1和/或s8蛋白酶的很大一部分将由于低ph条件而变性,但是使用本发明的变体,不需要将ph降低与使复合物和野生型ci-2a解离所需的如此低的ph值,因此使用本发明的变体将显著降低蛋白酶活性的损失。因此,本发明的变体可用于保护s1和/或s8蛋白酶免于自体蛋白水解,并保护包含一种或多种s1和/或s8蛋白酶和一种或多种其他蛋白酶的组合物中的蛋白质在中性和碱性区域ph下免于蛋白水解,在该ph下蛋白酶抑制剂复合物能够容易解离,并在需要时通过降低ph来释放蛋白酶活性。本发明的变体的用途本发明还涉及用于生产、回收和纯化s1和/或s8蛋白酶和/或本发明的抑制剂,优选本发明的变体和s1和/或s8蛋白酶的复合物的方法。在一个优选的实施例中,本发明涉及生产和回收s1或s8蛋白酶或由s1或s8蛋白酶和本发明的变体组成的复合物。在该实施例中,通过发酵过程制备由s1或s8蛋白酶和本发明的变体组成的复合物,其中产生s1或s8蛋白酶和本发明的变体并将其分泌到发酵液中。发酵可以是其中宿主细胞表达s1或s8蛋白酶和本发明变体两者的发酵,或者它可以是其中表达s1或s8蛋白酶的微生物与表达本发明变体的微生物共培养的发酵过程。优选地,s1或s8蛋白酶和本发明的变体以大约等摩尔量,例如以1:3至3:1的范围内的摩尔比、例如以1:2至2:1的范围内的摩尔比、例如以1:1.5至1.5:1的范围内的摩尔比、例如以1:1.3至1.3:1的范围内的摩尔比、例如以1:1.2至1.2:1的范围内的摩尔比、例如以1:1.1至1.1:1的范围内的摩尔比、或者以大约1:1的摩尔比表达。发酵应在s1或s8蛋白酶和本发明的变体形成复合物的ph值下进行,通常在高于5.5的ph值下,例如在ph5.5至9的范围内,例如在6.0至9.0的范围内。当发酵完成时,通过本领域已知的一系列分离步骤,例如过滤、离心、沉淀、微滤、浓缩等,回收由s1或s8蛋白酶和本发明的变体组成的复合物。在一些实施例中,由s1或s8蛋白酶和本发明的变体组成的复合物在发酵过程之后以沉淀形式存在于发酵液中。在此类实施例中,用于回收在发酵期间沉淀的酶的技术,例如披露于wo2008/110498、pct/ep2018/058387(10月初公开的14385)和wo2003/050274中的技术可以用于回收过程。如果需要在没有本发明的变体的情况下回收游离的s1或s8蛋白酶,则在回收过程中应包括低ph步骤,以使s1或s8蛋白酶和本发明的变体的复合物解离。用于解离复合物的低ph步骤可以在发酵后立即进行,或者可以在从发酵液中回收了复合物的回收过程之后进进行,或者可以在回收过程中的任何阶段进行。在同一发酵罐中发酵一种或多种s1和/或s8蛋白酶和本发明的一种或多种变体的好处是,可以完全或部分保护s1和/或s8蛋白酶免受蛋白水解,这通常意味着s1和/或s8蛋白酶的产量增加。回收后,可以如本领域已知的使用s1和/或s8蛋白酶。s1和/或s8蛋白酶从已测序的蛋白质数量和独特的肽酶活性数量来看,s1家族是所有肽酶家族中最大的家族。s1家族的肽酶包含在pa/s亚族的所有成员中发现的催化三联体his、asp和ser。已宣称丝氨酸被苏氨酸取代的人类睾丸特异性蛋白质tsp50的活性,尽管丝氨酸对于来自许多其他脊椎动物的该蛋白质来说是保守的。胰凝乳蛋白酶是众所周知的s1蛋白酶的一个实例。肽酶家族s8包含丝氨酸内肽酶枯草杆菌蛋白酶及其同源物。s8家族的成员具有催化三联体,其顺序为asp、his和ser,这与s1家族的顺序不同。在s8a亚家族中,活性位点残基经常发生在基序asp-thr/ser-gly、his-gly-thr-his和gly-thr-ser-met-ala-xaa-pro中。在s8b亚家族中,催化残基经常发生在基序asp-asp-gly、his-gly-thr-arg和gly-thr-ser-ala/val-ala/ser-pro中。在确定枯草杆菌蛋白酶的序列和结构之前,人们认为所有丝氨酸型肽酶都与胰凝乳蛋白酶同源。枯草杆菌蛋白酶显然非常不同,其与胰凝乳蛋白酶无关。就序列数目和特征化肽酶两者而言,s8家族也称为枯草杆菌酶家族,是丝氨酸肽酶的第二大家族。该家族分为两个亚家族,其中枯草杆菌蛋白酶是s8a亚家族的模范实例,而kexin是s8b亚家族的模范实例。根据本发明的s1蛋白酶的实例包括10r蛋白酶,以及与10r具有至少60%序列同一性,例如与seqidno:6具有至少70%序列同一性、例如至少80%序列同一性、例如至少90%序列同一性、例如至少95%序列同一性、例如至少96%序列同一性、例如至少97%序列同一性、例如至少98%序列同一性、例如至少99%序列同一性的蛋白酶。根据本发明的s8蛋白酶的实例包括枯草杆菌酶、s8a亚家族,例如衍生自解淀粉芽孢杆菌并且具有seqidno:3的序列的蛋白酶,衍生自迟缓芽孢杆菌并且具有seqidno:2的序列的蛋白酶;称为嘉士伯枯草杆菌蛋白酶并且具有seqidno:4的序列的芽孢杆菌属物种蛋白酶,以及衍生自芽孢杆菌属物种ty145并且具有seqidno:5的序列的蛋白酶,以及与seqidno:2、3、4或5之一具有至少60%序列同一性,例如至少70%序列同一性、例如至少80%序列同一性、例如至少90%序列同一性、例如至少95%序列同一性、例如至少96%序列同一性、例如至少97%序列同一性、例如至少98%序列同一性、例如至少99%序列同一性的枯草杆菌酶。核酸序列本发明还涉及分离的核酸序列,其编码本发明的蛋白酶抑制剂变体。用来分离或克隆编码多肽的核酸序列的技术在本领域中是已知的,并且包括从基因组dna分离、从cdna制备或其组合。可以例如通过使用熟知的聚合酶链式反应(pcr)或表达文库的抗体筛选来检测具有共有结构特征的克隆dna片段,实现从这样的基因组dna克隆本发明的核酸序列。参见例如,innis等人,1990,pcr:aguidetomethodsandapplication[pcr:方法和应用指南],academicpress[学术出版社],纽约。还可以使用其他核酸扩增程序,如连接酶链式反应(lcr)、连接激活转录(lat)和基于核苷酸序列的扩增(nasba)。分离的核酸序列可以例如通过基因工程中使用的标准克隆程序获得,以将核酸序列从其天然位置重新定位到将要复制的不同位点。克隆程序可以涉及切除和分离包括编码枯草杆菌酶的核酸序列的希望的核酸片段,将该片段插入载体分子,并将该重组载体掺入宿主细胞中,其中将会复制该核酸序列的多个拷贝或克隆。该核苷酸序列可以是基因组、cdna、rna、半合成、合成来源的、或其何组合。出于本发明的目的,如上所述确定两个核酸序列之间的同一性程度。修饰编码本发明的枯草杆菌酶的核酸序列对于合成与该枯草杆菌酶基本上相似的枯草杆菌酶可能是必需的。术语与枯草杆菌酶“基本上相似”是指枯草杆菌酶的非天然存在形式。这些枯草杆菌酶可以因某种工程化方式而与从其天然来源分离的枯草杆菌酶不同,例如在比活性、热稳定性、最适ph等方面不同的变体。对于核苷酸取代的一般描述,参见例如ford等人,1991,proteinexpressionandpurification[蛋白质表达与纯化]2:95-107。对于本领域技术人员显而易见的是,这些取代能够在对于分子功能重要的区域之外进行,并且仍然产生活性枯草杆菌酶。对于由本发明的分离的苷酸序列编码的多肽活性关键的并且因此优选不进行取代的氨基酸残基,可以根据本领域公知的方法,例如定点诱变或丙氨酸扫描诱变法(参见,例如,cunningham和wells,1989,science[科学]244:1081-1085)来鉴定。在后一技术中,将突变引入到分子中的每个荷正电的残基处,并且测试所得突变分子的蛋白水解活性,以鉴定对于所述分子的活性关键的氨基酸残基。底物-酶相互作用的位点也能够通过分析三维结构来确定,如通过核磁共振分析、晶体学或光亲和标记这样的技术所确定的(参见,例如,devos等人,1992,science[科学]255:306-312;smith等人,1992,journalofmolecularbiology[分子生物学杂志]224:899-904;wlodaver等人,1992,febsletters[欧洲生化学会联合会快报]309:59-64)。核酸构建体本发明还涉及核酸构建体,这些构建体包含与一个或多个能够指导多肽在合适的宿主细胞中表达的控制序列可操作地连接的本发明的核酸序列。可以用许多方式操作编码本发明的蛋白酶抑制剂复合物的分离的核酸序列,以提供枯草杆菌酶的表达。取决于表达载体,在将核酸序列插入载体之前对其进行操作可能是令人希望的或必需的。使用重组dna方法修饰核酸序列的技术是本领域熟知的。控制序列包括表达本发明的枯草杆菌酶所必需或有利的所有组分。每个控制序列对于编码枯草杆菌酶的核酸序列可以是天然的或外来的。此类控制序列包括但不限于前导序列、多腺苷酸化序列、前肽序列、启动子、信号肽序列、以及转录终止子。最少,控制序列包括启动子、以及转录和翻译终止信号。控制序列可以和用于引入特异性限制位点的接头一起提供,这些特异性限制位点促进控制序列与编码枯草杆菌酶的核酸序列编码区的连接。该控制序列可以是适当的启动子序列,即由宿主细胞识别以表达核酸序列的一种核酸序列。启动子序列包括介导枯草杆菌酶表达的转录控制序列。该启动子可以是在所选择的宿主细胞中显示出转录活性的任何核酸序列,包括突变的、截短的及杂合的启动子,并且可以是从编码与该宿主细胞同源或异源的细胞外或细胞内枯草杆菌酶的基因获得。用于指导本发明的核酸构建体转录,特别是在细菌宿主细胞中转录的合适启动子的实例是从以下获得的启动子:大肠杆菌lac操纵子、天蓝链霉菌(streptomycescoelicolor)琼脂糖酶基因(daga)、枯草芽孢杆菌果聚糖蔗糖酶基因(sacb)、地衣芽孢杆菌(bacilluslicheniformis)α-淀粉酶基因(amyl)、嗜热脂肪芽孢杆菌(bacillusstearothermophilus)产麦芽糖淀粉酶基因(amym)、解淀粉芽孢杆菌α-淀粉酶基因(amyq)、地衣芽孢杆菌青霉素酶基因(penp)、枯草芽孢杆菌(xyla和xylb基因和原核β-内酰胺酶基因(villa-kamaroff等人,1978,proceedingsofthenationalacademyofsciencesusa[美国国家科学院院刊]75:3727-3731),以及tac启动子(deboer等人,1983,proceedingsofthenationalacademyofsciencesusa[美国国家科学院院刊]80:21-25)。另外的启动子描述于scientificamerican[科学美国人],1980,242:74-94和sambrook等人,1989,同上,中的"usefulproteinsfromrecombinantbacteria[重组细菌中有用的蛋白质]"。调控序列也可以是合适的转录终止子序列,是由宿主细胞识别以终止转录的序列。该终止子序列被可操作地连接至编码该枯草杆菌酶的核酸序列的3’末端。在所选择的宿主细胞中发挥功能的任何终止子均可在本发明中使用。调控序列还可以是合适的前导序列,其为对于宿主细胞的翻译重要的mrna非翻译区。该前导子序列被可操作地连接至编码该多肽的核酸序列的5’末端。可以将在所选宿主细胞中有功能的任何前导序列用在本发明中。该控制序列也可以是聚腺苷酸化序列,即可操作地连接至该核酸序列的3’末端并且当转录时由宿主细胞识别成将多腺苷酸残基添加到所转录的mrna上的一个信号的一种序列。在所选择的宿主细胞中发挥功能的任何聚腺苷酸化序列均可在本发明中使用。该控制序列还可以是一种信号肽编码区,其编码连接至枯草杆菌酶的氨基末端并指导编码的枯草杆菌酶进入细胞分泌途径的氨基酸序列。核酸序列的编码序列的5’端可固有地含有在翻译阅读框中与编码分泌的枯草杆菌酶的编码区的区段天然地连接的一个信号肽编码区。可替代地,编码序列的5’端可以包含有对编码序列而言为外源的信号肽编码区。当编码序列天然地不包含信号肽编码区时,外源信号肽编码区可能是需要的。可替代地,外源信号肽编码区可简单地替换天然信号肽编码区以增强枯草杆菌酶的分泌。然而,在本发明中可以使用指导表达的枯草杆菌酶进入所选择的宿主细胞的分泌途径中的任何信号肽编码区。对于细菌宿主细胞有效的信号肽编码区是从如下酶的基因获得的信号肽编码区:芽孢杆菌属ncib11837产麦芽糖淀粉酶、嗜热脂肪芽孢杆菌(α-淀粉酶、地衣芽孢杆菌枯草杆菌蛋白酶、地衣芽孢杆菌β-内酰胺酶、嗜热脂肪芽孢杆菌中性蛋白酶(nprt、nprs、nprm)和枯草芽孢杆菌prsa。另外的信号肽由simonen和palva,1993,microbiologicalreviews[微生物评论]57:109-137描述。控制序列也可以是编码位于枯草杆菌酶氨基末端的氨基酸序列的前肽编码区。所得的多肽被称为前体酶(proenzyme)或多肽原(或在一些情况下被称为酶原(zymogen))。多肽原通常是无活性的并且可以通过催化切割或自身催化切割来自多肽原的前肽而转化为成熟的活性多肽。该前肽编码区可以获自以下各项的基因:枯草芽孢杆菌碱性蛋白酶(apre)、枯草芽孢杆菌中性蛋白酵(nprt)、酿酒酵母(saccharomycescerevisiae)α-因子、曼赫根毛霉(rhizomucormiehei)天冬氨酸蛋白酶、以及嗜热毁丝霉(myceliophthorathermophila)漆酶(wo95/33836)。当枯草杆菌酶的氨基末端同时存在信号肽和前肽区时,前肽区位于紧邻枯草杆菌酶的氨基末端并且信号肽区紧邻位于前导肽区的氨基末端。也可能令人希望的是添加调节序列,该调节序列允许相对于宿主细胞的生长而调节多肽的表达。调节系统的实例是引起基因表达响应化学或物理刺激物,包括调节化合物的存在而开启或关闭的那些系统。原核系统中的调节系统包括lac、tac和trp操纵子系统。在酵母中,可以使用adh2系统或gal1系统。表达载体包含编码本发明酶的dna构建体的重组表达载体可以是可方便地进行重组dna程序的任何载体,并且载体的选择通常取决于要引入的宿主细胞。因此,该载体可以是自主复制载体,即,以染色体外实体形式存在的载体,其复制与染色体复制无关,例如质粒。可替代地,该载体可以是这样的载体,当它引入宿主细胞中时部分或全部整合入宿主细胞基因组中并与其中已整合了它的一个或多个染色体一起复制。该载体优选是表达载体,其中编码本发明酶的dna序列可操作地连接至dna转录所需的其他区段。通常,该表达载体衍生自质粒或病毒dna,或可以包含这两者的元件。术语“可操作地链接”表示区段的排列方式使其可以按预期目的协同发挥发作,例如转录起始于启动子,并贯穿编码该酶的dna序列进行。启动子可以是在所选宿主细胞中显示转录活性的任何dna序列,并且可以衍生自编码与宿主细胞同源或异源的蛋白质的基因。用于细菌宿主细胞的合适启动子的实例包括嗜热脂肪芽孢杆菌产麦芽糖淀粉酶基因、地衣芽孢杆菌α-淀粉酶基因、解淀粉芽孢杆菌α-淀粉酶基因、枯草芽孢杆菌碱性蛋白酶基因或短小芽孢杆菌(bacilluspumilus)木糖苷酶基因的启动子,或噬菌体λpr或pl启动子或大肠杆菌lac、trp或tac启动子。如果需要,编码本发明酶的dna序列也可以可操作地连接至合适的终止子。本发明的重组载体可以进一步包含使载体能够在所讨论的宿主细胞中复制的dna序列。载体还可以包含选择标记,例如,其产物与宿主细胞中的缺陷互补的基因,或编码对例如抗生素(如卡那霉素、氯霉素、红霉素、四环素、大观霉素等)有抗性或对重金属或除草剂有抗性的基因。为了将本发明的酶引导至宿主细胞的分泌途径中,可以在重组载体中提供分泌信号序列(也称为前导序列,prepro序列或pre序列)。分泌信号序列在正确的阅读框中与编码酶的dna序列连接。分泌信号序列通常位于编码酶的dna序列的5'端。分泌信号序列可以是通常与酶相关的序列,也可以来自编码另一种分泌蛋白的基因。用于分别连接编码本发明酶的dna序列、启动子和任选地终止子和/或分泌信号序列、或通过合适的pcr扩增方案组装这些序列、并将其插入包含复制或整合所需信息的合适载体中的程序是本领域技术人员众所周知的(参见,例如,sambrook等人,同上)。宿主细胞编码引入宿主细胞中的本发明酶的dna序列可以与所讨论的宿主同源或异源。如果与宿主细胞同源,即由宿主细胞自然产生,则其通常将与其天然环境中的另一个启动子序列或,如果适用,另一个分泌信号序列和/或终止子序列可操作地连接。术语“同源”旨在包括编码原产自所讨论的宿主生物的酶的dna序列。术语“异源”旨在包括不由宿主细胞自然表达的dna序列。因此,dna序列可以来自另一种生物体,或者可以是合成序列。引入本发明的dna构建体或重组载体的宿主细胞可以是能够产生本发明酶的任何细胞,包括细菌、酵母、真菌和高等真核细胞。在培养时能够产生本发明酶的细菌宿主细胞的实例是革兰氏阳性菌,例如芽孢杆菌属的菌株,例如枯草芽孢杆菌、地衣芽孢杆菌、迟缓芽孢杆菌、短芽孢杆菌、嗜热脂肪芽孢杆菌、嗜碱芽孢杆菌(b.alkalophilus)、解淀粉芽孢杆菌、凝结芽孢杆菌(b.coagulans)、环状芽孢杆菌(b.circulans)、灿烂芽孢杆菌(b.lautus)、巨大芽孢杆菌(b.megatherium)或苏云金芽孢杆菌(b.thuringiensis)的菌株,或链霉菌属的菌株,例如变铅青链霉菌(s.lividans)或鼠灰链霉菌(s.murinus);或革兰氏阴性菌,例如大肠杆菌。细菌的转化可以通过原生质体转化、电穿孔、缀合或通过以本身已知的方式使用感受态细胞来实现(参见sambrook等人,同上)。当在诸如大肠杆菌的细菌中表达酶时,酶通常可以以不溶性颗粒(称为包涵体)的形式保留在细胞质中,或者可以通过细菌分泌序列导向至周质空间。在前一种情况下,将细胞裂解,将颗粒回收并变性,然后通过稀释变性剂使酶重新折叠。在后一种情况下,可通过破坏细胞,例如通过通过超声或渗透压休克,从周质空间中回收酶以释放周质空间的内容物并回收酶。当在革兰氏阳性菌,例如芽孢杆菌属或链霉菌属菌株中表达酶时,酶可保留在细胞质中,或可通过细菌分泌序列导向至细胞外培养基。在后一种情况下,可以如下所述从培养基中回收酶。产生蛋白酶和/或蛋白酶抑制剂复合物的方法本发明提供了产生根据本发明的分离的蛋白酶和/或变体抑制剂复合物的方法,其中在允许产生复合物的条件下,培养已被编码蛋白酶和/或变体抑制剂复合物的dna序列转化的合适的宿主细胞,并从培养物中回收所得的复合物或蛋白酶。当将包含编码蛋白质的dna序列的表达载体转化到异源宿主细胞中时,可以实现本发明蛋白质的异源重组产生。由此可以制备特征在于不含同源杂质的高度纯化的枯草杆菌酶组合物。在本文中,同源杂质是指源自同源细胞(最初从其获得本发明的蛋白质)的任何杂质(例如本发明的复合物或蛋白酶以外的其他多肽)。用于培养转化的宿主细胞的培养基可以是适合于生长所讨论的宿主细胞的任何常规培养基。表达的枯草杆菌酶复合物可以方便地分泌到培养基中,并且可以通过众所周知的方法从培养基中回收,包括通过离心或过滤从培养基中分离细胞,通过盐(例如硫酸铵)沉淀培养基的蛋白质在组分,然后进行色谱程序,例如离子交换色谱、亲和色谱等。优选的实施例本发明进一步由以下编号的实施例描述:实施例1.一种蛋白酶抑制剂,该蛋白酶抑制剂在6.0至9.0范围内的ph下具有抑制s1或s8蛋白酶的能力,其中当ph降低至低于6.0的ph时,s1或s8蛋白酶和该抑制剂的复合物解离。实施例2.如实施例1所述的蛋白酶抑制剂,其中该抑制剂是多肽。实施例3.如实施例1或2所述的蛋白酶抑制剂,其中当将ph调节至4.0至6.0范围内的ph值时,s1或s8蛋白酶和该抑制剂的复合物解离。实施例4.如实施例3所述的蛋白酶抑制剂,其中当将ph调节至4.5至6.0范围内的ph值时,s1或s8蛋白酶和该抑制剂的复合物解离。实施例5.如实施例4所述的蛋白酶抑制剂,其中当将ph调节至4.5至5.5范围内的ph值时,s1或s8蛋白酶和该抑制剂的复合物解离。实施例6.如实施例2所述的蛋白酶抑制剂,其中该蛋白酶抑制剂是如下变体,该变体具有蛋白酶抑制剂活性并且与seqidno:1的成熟多肽具有至少60%序列同一性,并且在与seqidno:1的位置21、25、26、30、33、43、44和52对应的一个或多个位置处包含取代。实施例7.如实施例6所述的变体,其中与包含具有seqidno:1的成熟多肽的序列的蛋白酶抑制剂和具有seqidno:2的序列的蛋白酶的复合物相比,包含该变体和具有seqidno:2的序列的蛋白酶的复合物在更高ph下解离。实施例8.如实施例7所述的变体,其中包含该变体和具有seqidno:2的序列的蛋白酶的复合物在解离时的ph值比包含具有seqidno:1的成熟多肽的序列的蛋白酶抑制剂和具有seqidno:2的序列的蛋白酶的复合物在解离时的ph值高至少0.4个ph单位,例如至少0.5个ph单位,例如至少0.6个ph单位,例如至少0.7个ph单位,例如至少0.8个ph单位,例如至少0.9个ph单位,例如至少1.0个ph单位。实施例9.如实施例6-8中任一项所述的变体,其中这些取代选自与seqidno:1中的以下取代相对应的取代:k21h、p25s、e26h、k30h、e33h、k43h、p44a和p52a。实施例10.如实施例6-9中任一项所述的变体,这些变体包含以下取代:k21h、p25s、e26h、k39h和e33h。实施例11.如实施例10所述的变体,这些变体进一步包含选自以下的一个或多个取代:d42n、e45g和q47h。实施例12.如实施例6至9中任一项所述的变体,这些变体包含以下取代:k43h+p44a+p52a;k21h+p25s+k30h;k21h+p25s+e26h+k30h+e33h;k30h+e33h+k43h+p44a+p52a;k21h+p25s+e26h+k30h+e33h+d42n+e45g;k21h+p25s+e26h+k30h+e33h+e45g;k21h+p25s+e26h+k30h+e33h+q47h;k21h+p25s+e26h+k30h+e33h+d42n+q47h;以及k21h+p25s+e26h+k30h+e33h+e45g+q47h。实施例13.如实施例12所述的变体,这些变体具有带有以下取代的seqidno:1的序列:k43h+p44a+p52a;k21h+p25s+k30h;k21h+p25s+e26h+k30h+e33h;k30h+e33h+k43h+p44a+p52a;k21h+p25s+e26h+k30h+e33h+d42n+e45g;k21h+p25s+e26h+k30h+e33h+e45g;k21h+p25s+e26h+k30h+e33h+q47h;k21h+p25s+e26h+k30h+e33h+d42n+q47h;或k21h+p25s+e26h+k30h+e33h+e45g+q47h。实施例14.一种多核苷酸,该多核苷酸编码如实施例1-13中任一项所述的蛋白酶抑制剂。实施例15.一种质粒、表达构建体或宿主细胞,该质粒、表达构建体或宿主细胞包含如实施例14所述的多核苷酸。实施例16.一种产生如实施例1-13中任一项所述的蛋白酶抑制剂的方法,该方法包括以下步骤:a.提供如实施例15所述的宿主细胞,b.在导致该蛋白酶变体表达的条件下培养该宿主细胞,产生发酵液;以及c.从该发酵液中回收该蛋白酶抑制剂变体。实施例17.一种产生由s1或s8蛋白酶和如实施例1-13所述的蛋白酶抑制剂组成的复合物的方法,该方法包括以下步骤:a.提供表达s1或s8蛋白酶的微生物和表达如实施例1-13所述的蛋白酶抑制剂的微生物;b.在诱导该s1或s8蛋白酶和该蛋白酶抑制剂变体表达的条件下,培养表达该s1或s8蛋白酶的微生物和表达该蛋白酶抑制剂变体的微生物,从而形成由该s1或s8蛋白酶和该蛋白酶抑制剂变体组成的复合物;以及c.从发酵液中回收由该s1或s8蛋白酶和该蛋白酶抑制剂变体组成的复合物。实施例18.如实施例17所述的方法,其中表达该s1或s8蛋白酶的微生物与表达该蛋白酶抑制剂的微生物相同。实施例19.一种产生s1或s8蛋白酶的方法,该方法包括以下步骤:a.提供表达s1或s8蛋白酶的微生物和表达如实施例1-13所述的蛋白酶抑制剂的微生物;b.在诱导s1或s8蛋白酶和蛋白酶抑制剂变体表达的条件下,培养表达该s1或s8蛋白酶的微生物和表达该蛋白酶抑制剂变体的微生物,从而形成由该s1或s8蛋白酶和该蛋白酶抑制剂变体组成的复合物;c.调节ph至该s1或s8蛋白酶和该蛋白酶抑制剂变体的复合物解离并释放蛋白酶活性的低ph值,以及d.回收该s1或s8蛋白酶。实施例20.如实施例19所述的方法,其中将步骤c中的ph调节至4.0至4.5范围内的ph值。实施例21.如实施例19或20所述的方法,其中表达该s1或s8蛋白酶的微生物与表达该蛋白酶抑制剂的微生物相同。实施例22.如实施例19至21中任一项所述的方法,其中步骤c在由该s1或s8蛋白酶和该蛋白酶抑制剂变体组成的复合物的回收过程之前、回收过程中或在回收之后进行。实施例23.如实施例17至22中任一项所述的方法,其中该s1或s8蛋白酶选自如下多肽,这些多肽具有蛋白酶活性并且与seqidno:2、seqidno:3、seqidno:4、seqidno:5或seqidno:6之一具有至少60%序列同一性,例如至少70%序列同一性;具有至少80%序列同一性,例如至少90%序列同一性;具有至少95%序列同一性,例如至少96%序列同一性;具有至少97%序列同一性,例如至少98%序列同一性。实例材料与方法缓冲液:通用缓冲液:100mm琥珀酸、100mmhepes、100mmchas、100mmcabs、1mmcacl2、150mmkcl、0.01%tritonx-100,ph2.0-6.0实例1ci-2a变体的制备使用基本上如wo2002/016619的实例1中所披露的技术,由包含编码具有seqidno:2的氨基酸序列的迟缓芽孢杆菌蛋白酶(savinase)的基因和编码变体的基因的枯草芽孢杆菌宿主细胞产生ci-2a变体。ci-2a变体的以下变体具有带有以下取代的seqidno:1的成熟多肽的序列:k43h+p44a+p52a;k21h+p25s+k30h;k21h+p25s+e26h+k30h+e33h;k30h+e33h+k43h+p44a+p52a;k21h+p25s+e26h+k30h+e33h+d42n+e45g;k21h+p25s+e26h+k30h+e33h+e45g;k21h+p25s+e26h+k30h+e33h+q47h;k21h+p25s+e26h+k30h+e33h+d42n+q47h;以及k21h+p25s+e26h+k30h+e33h+e45g+q47h。将表达savinase和变体的宿主菌株接种到蛋白酶敏感板上,其中这些菌株没有在菌落周围产生清除区,表明这些菌株没有分泌活性蛋白酶。产生savinase而不产生ci2a变体的对照菌株在菌落周围具有清除区。该实验表明,产生的变体是有活性的,并且能够抑制产生的savinase,从而防止形成清除区。实例2.savinase和ci-2a的共表达将枯草芽孢杆菌宿主菌株用3个拷贝的具有seqidno:2(savinase)的迟缓芽孢杆菌蛋白酶转化,并进一步用2个拷贝的、具有seqidno:1的成熟蛋白质的氨基酸序列的ci-2a野生型转化。在实验室规模的发酵罐中在37℃下分别发酵包含3个拷贝的迟缓芽孢杆菌蛋白酶的菌株、以及包含3个拷贝的迟缓芽孢杆菌蛋白酶和2个拷贝的ci-2a抑制剂的菌株,记录蛋白酶的产率并在图1中显示。结果表明,在表达迟缓芽孢杆菌蛋白酶但不表达ci-2a抑制剂的菌株的发酵液中的蛋白酶水平在发酵时间的一半之前达到了平台期,而表达迟缓芽孢杆菌蛋白酶和ci-2a抑制剂的菌株在整个发酵过程中继续生长,并达到不含该抑制剂的菌株所达到水平的超过2倍水平。实例3.savinase和ci-2a变体的共表达和蛋白酶活性的释放将枯草芽孢杆菌宿主菌株用3个拷贝的具有seqidno:2(savinase)的迟缓芽孢杆菌蛋白酶转化,并进一步用2个拷贝的、具有带有以下取代的seqidno:1的成熟蛋白质的氨基酸序列的ci-2a变体转化:(ci5.3)k21h+p25s+k30h;(ci08)k21h+p25s+e26h+k30h+e33h;或(ci10)k30h+e33h+k43h+p44a+p52a。基本上如实例2中所述制备这些菌株。在实验室规模的发酵罐中在37℃下分别发酵3个拷贝的迟缓芽孢杆菌蛋白酶和2个拷贝的ci-2a变体的菌株,评估蛋白酶的产率,发现其基本上与如图1所示的ci-2a野生型抑制剂的情况一样发育。实例4.从savinase和本发明变体的复合物释放蛋白酶活性使用以下程序测试实例3的发酵液在不同ph值下的蛋白酶活性的释放:将发酵液以14.000g离心3分钟,并回收上清液;将50μl上清液与200μl调节至ph3.0、3.5、4.0、4.5和5.0的通用缓冲液混合,并在30℃下孵育1小时;将每种孵育混合物在0.01%triton中稀释50倍,并使用suc-aapf-pna作为底物在ph9.0的通用缓冲液中测量蛋白酶活性。结果如图2所示。结果表明,对于ci-2a野生型以及每种变体,蛋白酶活性在ph5时以较低水平开始,并且蛋白酶水平随着ph降低而升高,因为蛋白酶从抑制剂释放。当ph值低于4-4.5时,蛋白酶活性可能再次下降,这是由于在如此低的ph值下蛋白酶的变性,并且当ph降至3.0时,所有蛋白酶活性都丧失。对于这些变体,可以看出,蛋白酶活性在比ci-2a野生型(在ph3.5附近达到峰值)更高的ph值处(在4和4.5之间)达到峰值,表明这些变体在比野生型ci-2a更高的ph下与蛋白酶解离。此外,可以看出,这些变体的蛋白酶活性明显高于野生型ci-2a,这可能是因为该蛋白酶在使野生型ci-2a抑制剂与该蛋白酶解离所必需的非常低的ph下不是非常稳定。实例5.savinase变体和ci-2a变体的共表达和蛋白酶活性的释放用3个拷贝的、具有带有取代s9en43rn76dv205iq206ly209ws259dn261wl262e(使用seqidno:3进行编号,披露于wo2016/087617中)的seqidno:2的氨基酸序列的迟缓芽孢杆菌蛋白酶变体转化枯草芽孢杆菌宿主菌株,并进一步用2个拷贝的、具有带有以下取代的seqidno:1的成熟蛋白的氨基酸序列的ci-2a变体转化:(ci08)k21h+p25s+e26h+k30h+e33h;或(ci10)k30h+e33h+k43h+p44a+p52a。基本上如实例2中所述制备这些菌株。基本上如实例4中所述,所不同的是使用2.0和6.0之间的ph值,测试发酵液在不同ph值下的蛋白酶活性的释放。结果如图3所示。结果表明,具有本发明变体的发酵液中的蛋白酶活性在比ci-2a野生型(在ph3.0附近达到峰值)更高的ph值处(在3.5和4.0之间)达到峰值,表明这些变体在比野生型ci-2a更高的ph下与该蛋白酶解离。在低于3的ph下,由于在此种苛刻条件下失活,蛋白酶活性迅速丧失。实例6.ty145蛋白酶和ci-2a变体的共表达和蛋白酶活性的释放用3个拷贝的、具有带有取代s27kn109ks111es171es173pg174ks175pf180yg182al184fq198en199kt297p(披露于wo2016/097350和wo2016/097354)的seqidno:5的氨基酸序列的蛋白酶变体转化枯草芽孢杆菌宿主菌株,并进一步用2个拷贝的、具有带有如下取代的seqidno:1的成熟蛋白的氨基酸序列的ci-2a变体转化:(ci08)k21h+p25s+e26h+k30h+e33h;或(ci10)k30h+e33h+k43h+p44a+p52a。基本上如实例2中所述制备这些菌株。如实例4中所述,测试发酵液在3.0至6范围内的不同ph值下的蛋白酶活性的释放。结果如图4所示。结果表明,对于包含本发明变体的发酵液,蛋白酶活性可以在ph4.0-4.5下释放。在ph3.5和更低下,蛋白酶活性在此种苛刻条件下完全失活。序列表<110>诺维信公司(novozymesa/s)<120>胰凝乳蛋白酶抑制剂变体及其用途<130>14786-ep-epa<160>6<170>patentin版本3.5<210>1<211>83<212>prt<213>大麦(hordumvulgaris)<400>1serservalglulyslysprogluglyvalasnthrglyalaglyasp151015arghisasnleulysthrglutrpprogluleuvalglylysserval202530gluglualalyslysvalileleuglnasplysproglualaglnile354045ilevalleuprovalglythrilevalthrmetglutyrargileasp505560argvalargleuphevalasplysleuaspasnilealaglnvalpro65707580argvalgly<210>2<211>269<212>prt<213>迟缓芽孢杆菌<400>2alaglnservalprotrpglyileserargvalglnalaproalaala151015hisasnargglyleuthrglyserglyvallysvalalavalleuasp202530thrglyileserthrhisproaspleuasnileargglyglyalaser354045phevalproglygluproserthrglnaspglyasnglyhisglythr505560hisvalalaglythrilealaalaleuasnasnserileglyvalleu65707580glyvalalaproseralagluleutyralavallysvalleuglyala859095serglyserglyservalserserilealaglnglyleuglutrpala100105110glyasnasnglymethisvalalaasnleuserleuglyserproser115120125proseralathrleugluglnalavalasnseralathrserarggly130135140valleuvalvalalaalaserglyasnserglyalaglyserileser145150155160tyrproalaargtyralaasnalametalavalglyalathraspgln165170175asnasnasnargalaserpheserglntyrglyalaglyleuaspile180185190valalaproglyvalasnvalglnserthrtyrproglyserthrtyr195200205alaserleuasnglythrsermetalathrprohisvalalaglyala210215220alaalaleuvallysglnlysasnprosertrpserasnvalglnile225230235240argasnhisleulysasnthralathrserleuglyserthrasnleu245250255tyrglyserglyleuvalasnalaglualaalathrarg260265<210>3<211>275<212>prt<213>解淀粉芽孢杆菌<400>3alaglnservalprotyrglyvalserglnilelysalaproalaleu151015hisserglnglytyrthrglyserasnvallysvalalavalileasp202530serglyileaspserserhisproaspleulysvalalaglyglyala354045sermetvalprosergluthrasnpropheglnaspasnasnserhis505560glythrhisvalalaglythrvalalaalaleuasnasnserilegly65707580valleuglyvalalaproseralaserleutyralavallysvalleu859095glyalaaspglyserglyglntyrsertrpileileasnglyileglu100105110trpalailealaasnasnmetaspvalileasnmetserleuglygly115120125proserglyseralaalaleulysalaalavalasplysalavalala130135140serglyvalvalvalvalalaalaalaglyasngluglythrsergly145150155160serserserthrvalglytyrproglylystyrproservalileala165170175valglyalavalaspserserasnglnargalaserpheserserval180185190glyprogluleuaspvalmetalaproglyvalserileglnserthr195200205leuproglyasnlystyrglyalatyrasnglythrsermetalaser210215220prohisvalalaglyalaalaalaleuileleuserlyshisproasn225230235240trpthrasnthrglnvalargserserleugluasnthrthrthrlys245250255leuglyaspserphetyrtyrglylysglyleuileasnvalglnala260265270alaalagln275<210>4<211>274<212>prt<213>芽孢杆菌属物种<400>4alaglnthrvalprotyrglyileproleuilelysalaasplysval151015glnalaglnglyphelysglyalaasnvallysvalalavalleuasp202530thrglyileglnalaserhisproaspleuasnvalvalglyglyala354045serphevalalaglyglualatyrasnthraspglyasnglyhisgly505560thrhisvalalaglythrvalalaalaleuaspasnthrthrglyval65707580leuglyvalalaproservalserleutyralavallysvalleuasn859095serserglyserglysertyrserglyilevalserglyileglutrp100105110alathrthrasnglymetaspvalileasnmetserleuglyglyala115120125serglyserthralametlysglnalavalaspasnalatyralaarg130135140glyvalvalvalvalalaalaalaglyasnserglyserserglyasn145150155160thrasnthrileglytyrproalalystyraspservalilealaval165170175glyalavalaspserasnserasnargalaserpheserservalgly180185190alagluleugluvalmetalaproglyalaglyvaltyrserthrtyr195200205prothrasnthrtyralathrleuasnglythrsermetalaserpro210215220hisvalalaglyalaalaalaleuileleuserlyshisproasnleu225230235240seralaserglnvalargasnargleuserserthralathrtyrleu245250255glyserserphetyrtyrglylysglyleuileasnvalglualaala260265270alagln<210>5<211>311<212>prt<213>芽孢杆菌属物种<400>5alavalproserthrglnthrprotrpglyilelysseriletyrasn151015aspglnserilethrlysthrthrglyglyserglyilelysvalala202530valleuaspthrglyvaltyrthrserhisleuaspleualaglyser354045alagluglncyslysaspphethrglnserasnproleuvalaspgly505560sercysthraspargglnglyhisglythrhisvalalaglythrval65707580leualahisglyglyserasnglyglnglyvaltyrglyvalalapro859095glnalalysleutrpalatyrlysvalleuglyaspasnglysergly100105110tyrseraspaspilealaalaalailearghisvalalaaspgluala115120125serargthrglyserlysvalvalileasnmetserleuglyserser130135140alalysaspserleuilealaseralavalasptyralatyrglylys145150155160glyvalleuilevalalaalaalaglyasnserglyserglyserasn165170175thrileglypheproglyglyleuvalasnalavalalavalalaala180185190leugluasnvalglnglnasnglythrtyrargvalalaasppheser195200205serargglyasnproalathralaglyasptyrileileglngluarg210215220aspilegluvalseralaproglyalaservalgluserthrtrptyr225230235240thrglyglytyrasnthrileserglythrsermetalathrprohis245250255valalaglyleualaalalysiletrpseralaasnthrserleuser260265270hisserglnleuargthrgluleuglnasnargalalysvaltyrasp275280285ilelysglyglyileglyalaglythrglyaspasptyralasergly290295300pheglytyrproargvallys305310<210>6<211>188<212>prt<213>拟诺卡氏菌属物种<400>6alaaspileileglyglyleualatyrthrmetglyglyargcysser151015valglyphealaalathrasnalaalaglyglnproglyphevalthr202530alaglyhiscysglyargvalglythrglnvalthrileglyasngly354045argglyvalphegluglnservalpheproglyasnaspalaalaphe505560valargglythrserasnphethrleuthrasnleuvalserargtyr65707580asnthrglyglytyralathrvalalaglyhisasnglnalaproile859095glyserservalcysargserglyserthrthrglytrphiscysgly100105110thrileglnalaargglyglnservalsertyrprogluglythrval115120125thrasnmetthrargthrthrvalcysalagluproglyaspsergly130135140glysertyrileserglythrglnalaglnglyvalthrserglygly145150155160serglyasncysargthrglyglythrthrphetyrglngluvalthr165170175prometvalasnsertrpglyvalargleuargthr180185当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1