玉米ZmAKINβγ1基因在培育耐铅胁迫的玉米中的应用的制作方法
本发明涉及玉米zmakinβγ1基因在培育耐铅胁迫的玉米中的应用,属于分子生物学领域。
背景技术:
玉米是世界范围内重要的粮食作物之一。然而随着生态环境的不断恶化,土壤中重金属的含量日益增加,其生长发育和产量品质受到包括重金属在内的非生物胁迫的严重影响,这严重制约了作物产量。挖掘抗逆相关基因,研究其逆境应答机制,培育抗逆性较强的玉米新品种是应对逆境胁迫,提高玉米产量的最有效途径。目前,我国玉米抗逆种质资源十分匮乏,常规育种得到抗逆种质资源不仅使一个非常艰难的过程,而且育种周期也比较长,很难培育出理想的抗性品种。随着现代分子生物学的飞速发展,为解决这个问题提供了一个很好的契机。近些年的研究表明,蛋白激酶在植物逆境基因表达调控中具有重要作用,其家族成员广泛地参与植物对环境胁迫的应答反应、植物的生长发育、代谢调控等一些列生理生化活动,尤其是zmakinβγ1在玉米抗病中受到强烈的关注。因此,zmakinβγ1基因及其与之互作的基因都是潜在的重要抗逆基因。
植物受到非生物胁迫,蛋白质的可逆磷酸化是细胞内信号转导的主要机制之一,由此维持植物正常的新陈代谢、信号转导及生长发育。蛋白激酶(proteinkinases,pk)主要参与蛋白磷酸化,它将腺苷三磷酸(atp)上的磷酸基团转移到没有活性的蛋白质分子的氨基酸残基(一般是丝氨酸残基、苏氨酸残基及酪氨酸残基)上,使其具有生物学功能。可逆磷酸化是在生物新陈代谢中必不可少的过程,包括钙依赖型蛋白激酶(cdpk)、受体蛋白激酶(rpk)和转录调控蛋白激酶等,这些都是在植物受到逆境胁迫后起信号传递作用。其中cdpk类蛋白激酶的研究最为深入,其主要是在细胞膜上感受外界刺激并通过膜识别、接收以及转化将环境信号变为细胞内信号。一般而言,在植物中,蛋白激酶受到非生物胁迫的诱导表达。例如,在拟南芥中的两种钙依赖型蛋白激酶catcdpk1和catcdpk2在拟南芥受到盐胁迫的时候诱导表达。在玉米中,钙依赖型蛋白激酶cdpk1基因在玉米植株受到盐胁迫的时候诱导表达。snrk在植物抵抗逆境环境以及生长发育中都表现出调节作用,而且在黑麦中snf1的同系物能使酵母snf1rkin1的突变体恢复表型,而akin蛋白激酶一是snrk亚家族中的一个成员。目前,发现玉米zmakinβγ1基因与玉米病害有关,但无与耐铅性的相关报道。
技术实现要素:
本发明克服了上述现有技术的不足,提供玉米zmakinβγ1基因在培育耐铅胁迫的玉米中的应用,本发明确定了玉米zmakinβγ1基因与玉米耐铅胁迫的关系,验证了该基因在培育耐铅胁迫的植物中的重要性。
玉米zmakinβγ1基因在培育耐铅胁迫的玉米中的应用。
进一步的,上述应用中所述玉米zmakinβγ1基因的序列如seqidno.1所示,其氨基酸序列如seqidno.2所示。
一种培育耐铅胁迫的玉米的方法,所述方法包括:上调玉米zmakinβγ1基因的表达。
进一步的,上述方法中所述上调玉米zmakinβγ1基因的表达为:构建zmakinβγ1在玉米中的过表达载体,通过农杆菌介导的遗传转化法对玉米幼胚进行侵染,将玉米zmakinβγ1基因转化至玉米幼胚中,自交三代,获得纯合阳性转化植株,即为耐铅玉米植株。
进一步的,上述方法中所述的zmakinβγ1在玉米中的过表达载体是利用cub载体骨架,以ubi作启动子,nos做终止子,bar作为选择标记基因,选择bamhⅰ位点,采用同源重组法构建而成
有益效果:
首次明确了玉米zmakinβγ1基因与玉米耐铅胁迫的关系,验证了该基因在玉米耐铅应用的重要性,为利用该基因在玉米及其他植物上的应用从而提高抗逆性提供了理论依据和利用价值。玉米zmakinβγ1基因在植物抗重金属污染领域,特别是玉米抗土壤铅毒领域具有广阔的应用前景,其经济效益潜力巨大。
附图说明
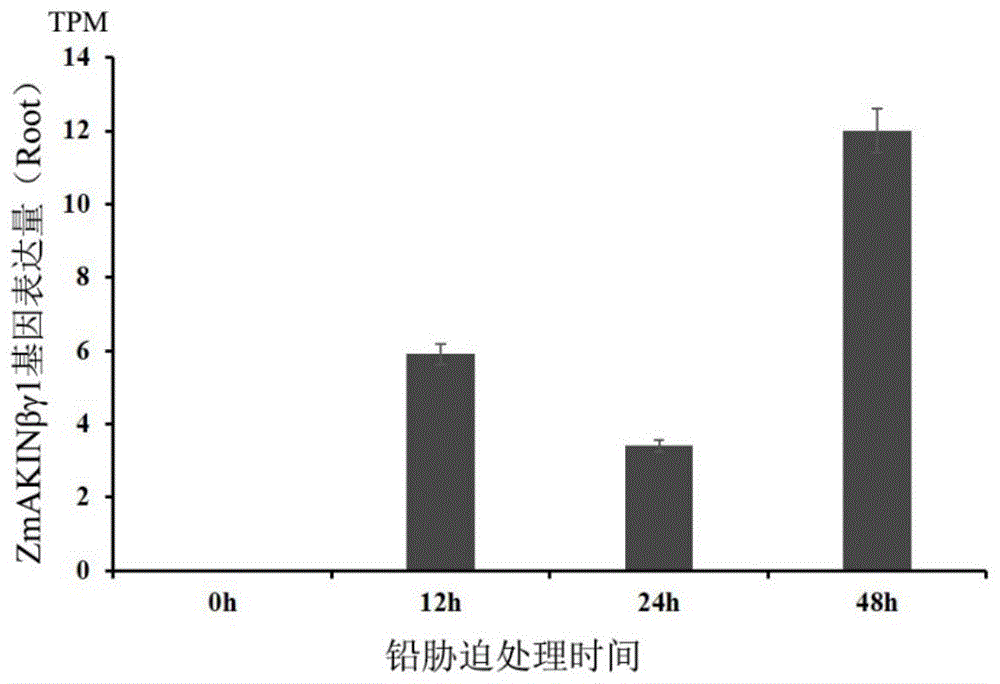
图1zmakinβγ1基因在玉米受到铅胁迫后的表达趋势。
图2全基因组关联分析显著位点。
图3拟南芥突变体与哥伦比亚野生型在铅胁迫下的长势情况。
图4哥伦比亚野生型以及突变体分别在正常条件(对照)以及0.15g/lpb(no3)2胁迫下根系长度对比。
图5zmakinβγ1的干涉载体图谱。
图6zmakinβγ1基因表达情况检测图。
图7转zmakinβγ1基因的过表达植株以及zmakinβγ1基因rna干扰植株中zmakinβγ1基因的表达水平。
图8转基因玉米受到铅胁迫后的表型。
具体实施方式
为了使本技术领域人员更好地理解本申请中的技术方案,下面结合实施例对本发明作进一步说明,所描述的实施例仅是本申请一部分实施例,而不是全部,本发明不受下述实施例的限制。
实施例1.玉米zmakinβγ1基因在铅胁迫下的表达模式分析
1-1、玉米自交系材料的铅胁迫处理
选择饱满的玉米骨干自交系178的种子用75%乙醇消毒1min,10℅的h2o2溶液浸泡15min并且消毒期间不断摇晃,然后用去离子水漂洗3~5次直到将残留的h2o2漂洗干净,再用去离子水浸泡4个小时,最后在28℃黑暗条件下用滤纸发芽。大约2-3天后,将发芽的玉米转移到漂浮板上,在28℃光照(16h)/黑暗(8h)条件下用hoagland营养液培养进行水培,一周换两次培养液,待幼苗长至三叶期进行胁迫处理。
1-2、铅胁迫处理:
选取30株长势一致的幼苗,去除胚乳后移入装有营养液的塑料容器中。本实验设置2个处理:正常水分供给组(即全营养液,ck)和铅胁迫处理组(即全营养液+3mmol/lpb(no3)2,t),每组12株幼苗。营养液要现用现配,并且使用前将营养液ph调至6.0左右。两组分别在0h,处理12h,24h及48h取玉米根系组织并提取rna,每次取样取3株,检测rna浓度及质量。用于基因数字表达谱分析。如图1所示,0h代表没有受铅胁迫两叶一心的玉米幼苗,12h,24h及48h分别代表处理时间。图中的玉米幼苗在0h的时候zmakinβγ1基因表达基本为0,在受到铅胁迫后诱导表达,并且在胁迫48h的时候表达量达到最高。表明zmakinβγ1基因受到重金属铅的诱导表达,并且在48h的时候表达量最高。
1-3、玉米总rna的提取
参照trizol试剂盒(invitrogen公司)操作手册对2-3中取的样品进行总rna的提取。具体步骤如下:(1)用液氮冷却研磨器:将已称量好的材料迅速放入研钵中并快速研磨,直至材料研磨成极细粉末;(2)按照每0.1克材料1ml的用量,将trizol加入研钵中;(3)20分钟,拆开研钵继续研磨,直至研钵内trizol呈透明状,其分装到2ml离心管中;(4)在上述离心管中再加入氯仿300μl,颠倒1分钟充分混匀,并静置5分钟后离心15min(4℃,12000rpm),小心吸取上清液至另一2ml离心管中;(5)重复第4步加氯仿及之后步骤,再次吸取上清液转至另一1.5ml离心管中;(6)再次吸取上清后,使用北京天恩泽基因科技有限公司的trizol伴侣提取总rna。(7)待rna沉淀晾干后,加入适量depc处理水溶解。
实施例2.全基因组关联分析鉴定到候选基因。
利用312份玉米自交系,处理方法以及表型鉴定同1-2,再结合56110个snp基因型数据,进行全基因组关联分析结果如图2所示,其中图标识的地方为zmakinβγ1基因所在的位置。
在第1号染色体上检测到4个snp位点(put-163a-60399874-3004,syn7988,syn7984,pze-101256211,分别位于1号染色体298967209bp,298967713bp,298969383bp,299548320bp,maizeb73refgen_v3),与根表面积,根干重,次生根长,地上干重,生物量,总根长等显著关联。其中,关联分析的结果如表1所示。zmakinβγ1(299461477-299469668bp)基因落在这四个snp的ld(500kb)区段里。进一步验证了该基因在玉米铅胁迫过程中的重要性。
表1全基因组关联分析与zmakinβγ1相关联的snp位点及关联的性状
实施例3.拟南芥同源基因突变体植株耐铅性分析
拟南芥同源基因salk_074210在tair拟南芥突变体库中订购,其中在图3和图4中标注为mutant。该突变体是t-dna插入突变体,在该突变体中zmakinβγ1在拟南芥中的同源基因功能丧失。首先利用75%无水乙醇对突变体和哥伦比亚野生型种子消毒1min,然后用1%naclo消毒15min,最后用无菌水清洗3~5遍后播种在1/2ms培养基上培养7天,然后将野生型植株以及突变体植株分别移植在0g/l以及0.15g/lpb(no3)2的1/2ms培养基上继续培养,10天后观察植株生长状况,结果如图3和图4所示,图3中的右侧图为哥伦比亚野生型和拟南芥突变体分别在正常条件下的长势(右侧图的左边的拟南芥为突变体,右边的拟南芥为野生型),图3中的左侧图为哥伦比亚野生型和拟南芥突变体分别在0.15g/lpb(no3)2胁迫下的长势(左侧图的左边的拟南芥为突变体,右边的拟南芥为野生型)。在正常条件下突变体植株的生长与野生型相差不大,在铅胁迫条件下,铅胁迫明显抑制了植物根系的生长,但是突变体的生长受到了更明显的抑制。野生型植株被铅胁迫后根系平均长度减少32.5%,而突变体植株的根系平均长度减少60%。
实验表明,哥伦比亚野生型在铅胁迫下的根系长度明显高于突变植株的根系长度,地上部分的长势也明显好于突变植株地上部分的长势。证实zmakinβγ1基因与植物耐铅胁迫密切相关。
实施例4.过表达及rnai玉米植株耐铅分析。
4-1、基因扩增
利用pcr对zmakinβγ1基因进行扩增,zmakinβγ1基因的核苷酸序列如seqidno.1所示,其编码蛋白氨基酸序列如seqidno.2所示。
其中反应体系如表2所示:
表2pcr扩增体系
反应程序为:95℃30s;98℃10s,tm(引物最适退火温度)30s,68℃1min10s,40cycles;68℃10min,4℃保存。所用基因pcr引物序列如下表所示:
表3pcr引物
4-2、转化玉米rnai载体及过表达载体构建及农杆菌介导的遗传转化
利用cub载体骨架,以ubi作启动子,nos做终止子,bar作为选择标记基因,选择bamhⅰ位点,采用同源重组法构建zmakinβγ1在玉米中的过表达载体。同时利用cub载体骨架,利用bamhⅰ和smalⅰ双酶切构建zmakinβγ1的干涉载体,载体图谱如图5所示。其中,图5中的targetgene就是目标基因以及rnai片段所在位置。rnai序列片段为:ggatccgcgacctaatgcatcacttagttcaagagactaagtgatgcattaggtcgctttttcccggg(seqidno.5)。将构建好的过表达和干涉载体分别转化eha105农杆菌感受态细胞。利用农杆菌介导的遗传转化法对玉米c01幼胚进行侵染,并培养得到t0代转zmakinβγ1玉米。自交三代,获得纯合阳性转化植株。其中阳性检测图片如图6所示。其中图6中的m代表2000bp的marker,“+”和“-”分别代表阳性对照和阴性对照,图6左图代表过表达检测条带,右图代表rnai检测条带。
其中阳性检测反应体系如下:
表4pcr扩增体系
反应程序为:95℃30s;95℃15s,tm(引物最适退火温度)30s,60℃40s,38cycles;72℃10min,4℃保存。所用基因pcr引物序列如下表所示:
表5pcr引物
4-3、耐铅性分析
利用已获得的过表达载体及rnai玉米植株(处理方法以及表型鉴定同4-2),用于做定量的取样时间为胁迫第三天的没有铅处理的玉米幼苗,表型鉴定时间为铅处理第七天。zmakinβγ1基因表达量结果如图7所示,在正常条件下,rnai玉米植株无论是叶片还是根系,zmakinβγ1基因的表达量均显著低于野生型,而过表达植株无论是叶片还是根系,zmakinβγ1基因的表达量均显著高于野生型。表型鉴定结果如图8所示,图8中左一到左三分别为对照条件下未转化的玉米幼苗,过表达zmakinβγ1基因的玉米幼苗以及zmakinβγ1基因rna干扰玉米幼苗的生长状况;右三到右一分别为铅胁迫条件下未转化的玉米幼苗,过表达zmakinβγ1基因的玉米幼苗以及zmakinβγ1基因rna干扰玉米幼苗的生长状况。过表达植株在受到铅胁迫后,叶片发育并没有停滞,而野生型叶片发育受到铅胁迫过后生长明显受到抑制,并且叶片更加萎蔫叶片变黄。干涉zmakinβγ1基因后,根系生长以及叶片发育在受到铅胁迫后受到比野生型更明显的抑制。证实zmakinβγ1基因与玉米耐铅胁迫密切相关。
虽然,上文已经用一般性说明、具体实施方式及试验,对本发明作了详尽的描述,但在本发明基础上,可以对之做出一些修改或改进,这对本领域技术人员而言是显而易见的。因此,在不偏离本发明精神的基础上所做的这些修改或改进,均属于本发明要求保护的范围。
sequencelisting
<110>四川农业大学
<120>玉米zmakinβγ1基因在培育耐铅胁迫的玉米中的应用
<130>2020
<160>9
<170>patentinversion3.3
<210>1
<211>1494
<212>dna
<213>序列
<400>1
atgttctcgcacggcgctgattccgcccacgacgccgggaccgttggcgtcagcagcggc60
gtcggggccaccgtccccgcccgcttcgtctggccctacggtgggaagagggtcttcgtc120
tcaggctccttcaccaggtggtcggaacatttgccgatgtctcctgtcgaaggctgcccc180
actgtatttcaggctatttgcagcctgtctccagggattcatgagtacaagttctatgtg240
gacggggagtggcgtcatgatgagcgccaacctactatatctggggagttcggcatagtt300
aacacactatacttgacaagggaatttaatcaaataaacgccttattaaatccaagcaca360
cctggaagcaggatgaacatggatgtggataatgaaaattttcaacatacggttacattg420
tcggatggtaccattccagaaggtactgtgagagtttcagaggctgcaatacaaatctca480
aggtgccgcgtttctgagtatctcaatttgcatacatgctatgatttactcccggattcg540
ggcaaggttattgctctggacattaatttacctgtgaagcaatcttttcatattctgcat600
gaacaggggattcctgtagctcctctctgggactcattcagaggtcaatttgtcggcctt660
ctgagcccattggatttcatacttatattgcgggagctagaaactcatggctcgaacttg720
acagaagatcagcttgaaacacacactatatctgcatggaaagaggctaagcggcaaact780
tgtggaagaaatgatggtcagtggcgagcacatcagcatctagtgcatgccaccccttat840
gagtccttgagggacattgcagtaaagcttttgcaaaatgacatttctacagtgccagtt900
atttattcatcatcatcagatggatcattccctcagttattgcaccttgcatccctttct960
ggaattttgaaatgtatttttaggtattttaaaaactcaactggtaatttgcctattctg1020
aaccaaccggtgtgctccattccgctgggttcctgggttccgaaaatcggtgatccaaac1080
agtcgtccattggctatgttgcgacctaatgcatcacttagctctgcccttaacatgttg1140
gttcaagctggagtgagctcaataccaattgtggatgaaaacgactccctgcttgacact1200
tactctagaagtgacatcacagccctagctaaagacaaggtctacacacatgttcgcctg1260
gatgagatgaccattcatcaggctttacagcttggacaagatgccaatacgccttttggt1320
ttttttaacggtcagagatgccagatgtgcctccggtctgatcctttgctgaaggtgatg1380
gagcgactggctaatccgggggtgcggcgggtgttcatagtagaagctgggagcaaacgt1440
gtggagggtattatatcactgagtgatattttcaagttcttgctgagcttgtga1494
<210>2
<211>497
<212>prt
<213>序列
<400>2
metpheserhisglyalaaspseralahisaspalaglythrvalgly
151015
valserserglyvalglyalathrvalproalaargphevaltrppro
202530
tyrglyglylysargvalphevalserglyserphethrargtrpser
354045
gluhisleuprometserprovalgluglycysprothrvalphegln
505560
alailecysserleuserproglyilehisglutyrlysphetyrval
65707580
aspglyglutrparghisaspgluargglnprothrileserglyglu
859095
pheglyilevalasnthrleutyrleuthrargglupheasnglnile
100105110
asnalaleuleuasnproserthrproglyserargmetasnmetasp
115120125
valaspasngluasnpheglnhisthrvalthrleuseraspglythr
130135140
ileprogluglythrvalargvalserglualaalaileglnileser
145150155160
argcysargvalserglutyrleuasnleuhisthrcystyraspleu
165170175
leuproaspserglylysvalilealaleuaspileasnleuproval
180185190
lysglnserphehisileleuhisgluglnglyileprovalalapro
195200205
leutrpaspserpheargglyglnphevalglyleuleuserproleu
210215220
asppheileleuileleuarggluleugluthrhisglyserasnleu
225230235240
thrgluaspglnleugluthrhisthrileseralatrplysgluala
245250255
lysargglnthrcysglyargasnaspglyglntrpargalahisgln
260265270
hisleuvalhisalathrprotyrgluserleuargaspilealaval
275280285
lysleuleuglnasnaspileserthrvalprovaliletyrserser
290295300
serseraspglyserpheproglnleuleuhisleualaserleuser
305310315320
glyileleulyscysilepheargtyrphelysasnserthrglyasn
325330335
leuproileleuasnglnprovalcysserileproleuglysertrp
340345350
valprolysileglyaspproasnserargproleualametleuarg
355360365
proasnalaserleuserseralaleuasnmetleuvalglnalagly
370375380
valserserileproilevalaspgluasnaspserleuleuaspthr
385390395400
tyrserargseraspilethralaleualalysasplysvaltyrthr
405410415
hisvalargleuaspglumetthrilehisglnalaleuglnleugly
420425430
glnaspalaasnthrpropheglyphepheasnglyglnargcysgln
435440445
metcysleuargseraspproleuleulysvalmetgluargleuala
450455460
asnproglyvalargargvalpheilevalglualaglyserlysarg
465470475480
valgluglyileileserleuseraspilephelyspheleuleuser
485490495
leu
<210>3
<211>21
<212>dna
<213>人工序列
<400>3
atgttctcgcacggcgctgat21
<210>4
<211>19
<212>dna
<213>人工序列
<400>4
tcacaagctcagcaagaac19
<210>5
<211>68
<212>dna
<213>人工序列
<400>5
ggatccgcgacctaatgcatcacttagttcaagagactaagtgatgcattaggtcgcttt60
ttcccggg68
<210>6
<211>18
<212>dna
<213>人工序列
<400>6
gccctgccttcatacgct18
<210>7
<211>18
<212>dna
<213>人工序列
<400>7
gcaaatgttccgaccacc18
<210>8
<211>22
<212>dna
<213>人工序列
<400>8
ggctttacactttatgcttccg22
<210>9
<211>24
<212>dna
<213>人工序列
<400>9
gccttcatacgctatttatttgct24
- 还没有人留言评论。精彩留言会获得点赞!