一种重组木聚糖酶及其应用
本发明属于基因工程领域,具体涉及一种重组木聚糖酶及其应用。
背景技术:
农业废弃物含有丰富的纤维素和半纤维素,木聚糖是植物半纤维素的主要组成部分,其含量仅次于纤维素,为第二大可再生资源,但未得到有效的开发及利用,造成我国农业发展面临着农业废弃物排放量大、环境污染严重等突出问题。充分利用农业废弃物中木聚糖有助于缓解环境、生态和能源问题,并增加农业废物的附加值。
木聚糖酶(endo-1,4-β-d-木聚糖酶,ec3.2.1.8)是一类水解酶,可水解木聚糖的β-1,4-连接的糖苷键以产生高附加值的产品,例如木寡糖(xos)和木糖(x1)。工业上利用木聚糖酶生产高附加值低聚木糖,要求酶具有耐热性、耐酸碱、高酶活、低聚木糖(以木二糖和木三糖为主)产率高等特性。目前对gh11木聚糖酶耐热性和耐酸碱性研究较多,但酶催化底物生成特异性水解产物-低聚木糖(主要是木二糖和木三糖)相关研究较少,且水解特异性机制不清楚,而gh11木聚糖酶利用低附加值的农业废弃物生产高附加值的低聚木糖,关键在于其水解特异性机制的研究。类似于n端改造可以引起g11木聚糖酶水解产物改变,c端对g11木聚糖酶水解特异性也存在一定的影响。li等报道了来自嗜热裸节菌f1208木聚糖酶(t-xyn)用序列fptgnttelekrqttp(1-16)替换aftvgngq并结合位于c末端193位丝氨酸替换苯丙氨酸的策略,产生杂合酶t-xynfm,其具有特殊水解特性:以木三糖(x3)、木四糖(x4)和木五糖(x5)为底物时,木糖均未被检出。t-xynfm水解木聚糖(桦木、榉木和燕麦木聚糖)时,产物中木糖生成量比t-xyn低,最大减少量约为90.22%(燕麦木聚糖)。t-xynfm水解木三糖的产物中出现木四糖,其生成量多于60%,表明t-xynfm具有较强的转糖苷酶活性,该活性对生产低聚木糖具有较强的吸引力。li等从酸性木聚糖酶pjxyn引入二硫键构建突变体pjxyns(27)s(186),以桦木、榉木和燕麦木聚糖为底物,与野生型pjxyn比,水解产物含有更多木糖和木二糖,分别为87.62%(榉木木聚糖)和69.91%(燕麦木聚糖)。由此可知,通过对木聚糖酶进行点突变可能会改变酶水解特性。
技术实现要素:
针对木聚糖酶在工业应用中存在的问题,本发明提供一种高效水解木聚糖的工程菌,诱导表达的木聚糖酶水解效率高,ph稳定性好,在酸性条件下ph耐受性增强。
为实现上述目的,本发明采用如下技术方案。
一种木聚糖酶,其氨基酸序列为seqidno:1-3所示;其中seqidno:2为seqidno:1所示序列121位精氨酸突变为丙氨酸,seqidno:3为seqidno:1所示序列179位的酪氨酸突变为丙氨酸。
优选的,编码上述木聚糖酶氨基酸序列的核苷酸序列如seqidno:4-6所示。
一种包含编码上述氨基酸的核苷酸序列的载体或工程菌。
一种上述工程菌在制备木聚糖酶中的应用。
一种上述木聚糖酶在制备低聚木糖中的应用。
本发明具有以下优点:
本发明提供了一种基因改造的木聚糖酶及生产该酶的工程菌。通过对特殊位点的改造,提高了木聚糖酶水解各种木聚糖的效率,且ph稳定性好,耐酸性增强,在工业上具有广泛的应用前景。
附图说明
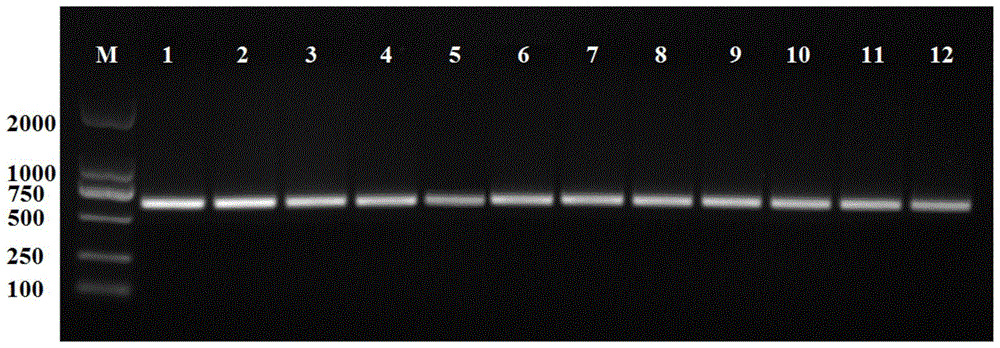
图1为xyna、xyna179和xyna121基因pcr产物1%琼脂糖电泳图;
图2为xyna、xyna179和xyna121粗酶液和纯化酶液的sds-page电泳图;
图3为xyna、xyna179和xyna121酶的最适ph和ph稳定性;
图4为xyna对不同木聚糖的水解特性;
图5为xyna121对不同木聚糖的水解特性;
图6为xyna179对不同木聚糖的水解特性。
具体实施方式
下面结合实施例和附图对本发明做进一步说明,但本发明不受下述实施例的限制。
实施例1重组酶xyna功能域突变体的构建
1.木聚糖酶xyna基因的获得
将源于streptomycerocheil2001g11木聚糖酶xyna基因切除信号肽序列,获得如seqidno:1所示的目的基因,将其连接入载体pmd18-t,转染e.colidh-5α,获得的阳性菌进行-80℃保存。
提取e.colidh-5α中含有xyna目的片段的质粒,将其作为pcr反应中的模板,以加酶切位点的引物1s11f2/1s11r1(见表1)进行pcr扩增,获得含酶切位点目的基因片段xyna。
表1带酶切位点羧肽酶基因克隆引物序列(下划线部分为酶切位点)
2.xyna突变体目的基因的获得
对木聚糖酶xyna121位点和179位点分别进行突变,将121位、179位的碳长链精氨酸、较大侧链的酪氨酸均突变为丙氨酸,获得突变酶xyna121和xyna179。采用重叠延伸pcr技术进行目的基因扩增,带有酶切位点的引物见表2,下划线部分为突变位点。
表2重叠延伸pcr引物序列
获得的pcr产物进行1%琼脂糖电泳,如图1所示,1-4泳道为xyna179,5-8泳道为xyna121,9-12泳道为xyna,将测序正确的pcr产物保存,备用。
3.重组质粒的构建
将目的基因xyna、xyna121、xyna179及表达载体pet-28a分别以ncoi和xhoi进行双酶切,之后用t4dna连接酶分别进行连接,获得带有目的片段的重组质粒pet-28a-xyna、pet-28a-xyna121、pet-28a-xyna179。
4.重组工程菌的获得
将构建好的重组质粒pet-28a-xyna、pet-28a-xyna121、pet-28a-xyna179分别转染感受态细胞e.colide3,随机挑取lb抗性筛选平板中的克隆子进行菌落pcr验证,产物测序验证,将验证正确的pet-28a-xyna/e.colide3及突变体重组菌甘油管保藏。
实施例2xyna、xyna121、xyna179的诱导表达、纯化
1.xyna、突变酶xyna121、xyna179粗酶液的获得
将实施例1中获得的重组菌液分别接种于含有40μg/mlkan+的5mllb液体培养基,37℃200r/min培养12h。然后以1%的接种量将活化后菌液转接至含40μg/mlkan+的100mllb液体培养基中,37℃2000r/min培养至od600值达0.6-0.8,再加入iptg至终浓度为0.5mmol/l,20℃200r/min振荡培养14-16h进行诱导表达。将发酵液于10000rpm、4℃离心5min,收集菌体。采用10ml0.05mol/lph6.0的柠檬酸钠缓冲液重悬菌体,超声破壁15min后10000r/min、4℃离心5min,所得上清液即为粗酶液。
2.xyna、xyna121、xyna179的纯化
重组木聚糖酶带有组氨酸标签能与金属离子ni2+特异性结合,咪唑也对ni2+具有结合作用,二者形成竞争性关系。因此,采用ni柱(nisepharosehpcolumn,1×10cm)及äktafplc蛋白纯化系统对重组木聚糖酶进行纯化。
(1)柱平衡:选用平衡缓冲液(bindingbuffer)冲柱,直至基线平稳;
(2)上样:将已过0.22μm水系膜样品先注入收集管中暂时储存样品,待柱平衡自动上样;
(3)除杂与洗脱:用平衡缓冲液冲洗层析柱,去除非特异性结合的蛋白,采用含不同咪唑浓度的洗脱缓冲液对重组蛋白进行梯度洗脱,同时检测a280值,收集洗脱峰处的液体,即纯化酶液,置于4℃保藏。
取10μg粗酶液和纯化的酶液进行sds-page聚丙烯酰胺凝胶电泳验证,分离胶浓度为12.5%(w/v),浓缩胶浓度为4.5%(w/v)。
经过sds-page电泳检测,如图2所示,其中,1、2泳道为xyna粗酶液和纯化酶液,3、4泳道为xyna179粗酶液和纯化酶液,5、6泳道为xyna121粗酶液和纯化酶液。由图可知,粗酶液在20kda左右有较粗的蛋白条带,与预测蛋白量20.8kda接近,表明xyna和xyna179、xyna121成功表达;且纯化的酶液蛋白目的条带单一,成功获得了电泳纯的酶液,适于用于酶学性质的研究。
实施例3xyna、xyna121和xyna179的最适ph和ph稳定性
测定ph对酶活力的影响,选择ph3.0、3.5、4、4.5、5、5.5、6、6.5、7、7.5、8、8.5、9.0的(50mm)缓冲液:柠檬酸盐缓冲液ph3.0-6.5、巴比妥钠缓冲液ph7.0-9.0,在37℃条件下测定酶活。最高的木聚糖酶活性定义为100%,计算在不同ph下的相对酶活力,绘制相对酶活力随ph值的变化曲线,如图3a(xyna121)和图3b(xyna179)所示。
测定酶的ph稳定性,将酶置于上述测定的不同ph缓冲液中保温,即37℃保温30min,冰水浴30min后,于37℃、最适ph条件下测定残余酶活。以未经保温处理的酶测定的酶活定义为100%,计算残余酶活力,绘制残余酶活力随ph值的变化曲线,如图3c(xyna121)和图3d(xyna179)所示。
由图3可知,突变酶xyna121与xyna最适反应ph相同为6.0,而突变酶xyna179的最适ph值相比原酶降低,更偏酸性,为4.5。突变酶xyna121和xyna179在ph3.0-5.0条件下,残余酶活均达到80%以上,是原酶的4-6倍,较原酶有大幅提高;在ph5.0-9.0的条件下,突变酶xyna121和xyna179与原酶残余酶活力变化趋势基本一致,均能保持80%以上。较原酶,xyna121的ph稳定性范围变广,而xyna121和xyna179在酸性条件下ph耐受性均增强。
实施例4xyna、xyna121、xyna179水解特性
1.xyna、xyna121、xyna179各酶的比活力
将采用bca蛋白浓度试剂盒,测定纯化前后未突变及突变的重组木聚糖酶的蛋白浓度。重组木聚糖酶的酶活力测定参照dns法。木聚糖酶酶活测定反应体系包括0.9ml相应缓冲液配制的1%(w/v)榉木木聚糖底物保温5min,加入0.1ml适当稀释的酶液,55℃反应10min。加入1mldns溶液终止反应,沸水浴15min,冷却并离心,加入1ml40%(w/v)四水合酒石酸钾钠溶液,540nm条件下测定吸光度值,采用木糖绘制标准曲线计算还原糖(以木糖计)的量。1个木聚糖酶活力单位(u)定义为:在上述实验条件下,每分钟水解木聚糖生成1μmol木糖所需要的酶量。
表3xyna、xyna121、xyna179纯酶液的比酶活
如表3所示,较原酶,突变酶xyna179、xyna121的比酶活降低。
2.xyna、xyna121、xyna179各酶的水解特性
分别以榉木木聚糖、桦木木聚糖、燕麦木聚糖为底物进行水解反应,由于各酶在30℃下残余酶活可达到90%以上,故选择30℃作为水解温度。
为了表征木聚糖酶的水解,将含有500μl反应混合物,包括50mmph6.0的柠檬酸缓冲液配置的10mg/ml木聚糖底物和5u/ml木聚糖酶,30℃下150rpm孵育0、15min、30min、1h、2h、4h、8h,然后煮沸10min并在冰水浴中冷却并离心。离心液测定不同低聚木糖和木糖的含量:将离心液用0.22μm膜滤器过滤,采用watershplc对样品进行分析,色谱柱为ks-802,配有waters2695示差检测器(ri)。色谱条件为:柱温65℃,流动相为高纯水,流速为0.6ml/min,进样20min,进样量10μl。木糖(x1)、木二糖(x2)、木三糖(x3)、木四糖(x4)和木五糖(x5)溶液作为标准溶液。所有实验一式三份进行。以水解时间为横坐标,以水解产物浓度为纵坐标做图4(xyna)、图5(xyna121)和图6(xyna179),其中a为不同酶水解桦木木聚糖,b为不同酶水解榉木木聚糖,c为不同酶水解燕麦木聚糖。
通过比较图5和图4,木聚糖酶xyna121水解桦木和榉木木聚糖,主要产生了x2、x3、x4,水解8h时,其占比分别为38.1%、51.4%、10.5%和40.2%、52.2%、7.6%,较原酶其产物组成比例发生变化。以桦木木聚糖为底物,x2比例减少,x3和x4比例增加。水解8h,低聚木糖(x2+x3+x4+x5)总量达到2.20mg/ml,xyna121水解30min时,低聚木糖(x2+x3+x4+x5)产量达到2.07mg/ml,相当于原酶水解8h(2.10mg/ml),因此,xyna121水解效率提高。以榉木木聚糖为底物,x2和x4占比降低,x3占比升高。水解8h,低聚木糖(x2+x3+x4+x5)总量达到2.81mg/ml,xyna121水解30min,其产量(2.36mg/ml)相当于原酶水解1h(2.38mg/ml)。由此可知,xyna121水解效率提高。以燕麦木聚糖为底物,主要产生了x2、x3、x4、x5。水解8h时,低聚木糖(x2+x3+x4+x5)总量(4.36mg/ml)较原酶(3.81mg/ml)增加了14%,xyna121水解2h,其产量(3.38mg/ml)相当于原酶水解4h(3.21mg/ml),由此可知,其水解效率较原酶明显提高,不仅体现在时间缩短,还有产量提高。
通过比较图6和图4,木聚糖酶xyna179水解桦木和榉木木聚糖,主要产生了x2、x3、x4。水解8h时,其占比为20.6%、61.1%、18.3%和19.7%、60.1%、20.2%,产生少量x1和x5,与原酶相比,水解产物的比例不同,x2比例明显下降,x3和x4比例明显上升,xyna179水解两种木聚糖水解产物及规律相似。以桦木木聚糖为底物,水解8h,低聚木糖(x2+x3+x4+x5)总量为1.33mg/ml,比原酶减少了37%,水解效率降低。以榉木木聚糖为底物,水解8h,x3达到1.04mg/ml,x4(0.35mg/ml)较原酶(0.23mg/ml)增加了50%,低聚木糖(x2+x3+x4+x5)总量(1.76mg/ml)比原酶(2.72mg/ml)减少了35%,水解效率降低。以燕麦木聚糖为底物,主要产生了x2、x3、x4、x5,产生少量x1。低聚木糖(x2+x3+x4+x5)总量(1.67mg/ml)比原酶(3.81mg/ml)减少了56%,水解更缓慢,水解效率显著降低。
这说明,通过对121位点和179位点的突变能够改变木聚糖降解产物的比例,相比于原酶,能够提高低聚木糖的比例,降低木糖比例,表现为突变酶的比酶活下降。对木聚糖酶xyna虽然都是进行了侧链较长或较大氨基酸替换成较小氨基酸的突变,但是突变位点对木聚糖酶的水解效率的影响却不同,相比于原酶,xyna179的水解效率降低,而xyna121的水解效率提高。
序列表
<110>北京工商大学
<120>一种重组木聚糖酶及其应用
<130>20201228
<160>12
<170>patentinversion3.5
<210>1
<211>191
<212>prt
<213>streptomycesrameus
<400>1
alathrvalvalthrthrasnglnthrglythraspasnglyphetyr
151015
tyrserphetrpthraspalaglnglythrvalsermetthrleugly
202530
serglyglyasntyrserthrsertrpargasnthrglyasnpheval
354045
alaglylysglytrpserthrglyalaargargasnvalthrtyrser
505560
glyserpheasnproserglyasnglytyrleuserleutyrglytrp
65707580
thrserasnproleuvalglutyrtyrilevalaspasntrpglythr
859095
tyrargprothrglythrtyrlysglyservalthrseraspglygly
100105110
thrtyraspiletyrglnthrthrargtyrasnalaproservalglu
115120125
glythrargthrpheasnglntyrtrpservalargglnserlysarg
130135140
thrglyglythrilethrthrglyasnhispheaspalatrpalaarg
145150155160
alaglymetproleuglyserphealatyrtyrmetileleualathr
165170175
gluglytyrglnserserglyasnserasnilethrvalserser
180185190
<210>2
<211>191
<212>prt
<213>streptomycesrameus
<400>2
alathrvalvalthrthrasnglnthrglythraspasnglyphetyr
151015
tyrserphetrpthraspalaglnglythrvalsermetthrleugly
202530
serglyglyasntyrserthrsertrpargasnthrglyasnpheval
354045
alaglylysglytrpserthrglyalaargargasnvalthrtyrser
505560
glyserpheasnproserglyasnglytyrleuserleutyrglytrp
65707580
thrserasnproleuvalglutyrtyrilevalaspasntrpglythr
859095
tyrargprothrglythrtyrlysglyservalthrseraspglygly
100105110
thrtyraspiletyrglnthrthralatyrasnalaproservalglu
115120125
glythrargthrpheasnglntyrtrpservalargglnserlysarg
130135140
thrglyglythrilethrthrglyasnhispheaspalatrpalaarg
145150155160
alaglymetproleuglyserphealatyrtyrmetileleualathr
165170175
gluglytyrglnserserglyasnserasnilethrvalserser
180185190
<210>3
<211>191
<212>prt
<213>streptomycesrameus
<400>3
alathrvalvalthrthrasnglnthrglythraspasnglyphetyr
151015
tyrserphetrpthraspalaglnglythrvalsermetthrleugly
202530
serglyglyasntyrserthrsertrpargasnthrglyasnpheval
354045
alaglylysglytrpserthrglyalaargargasnvalthrtyrser
505560
glyserpheasnproserglyasnglytyrleuserleutyrglytrp
65707580
thrserasnproleuvalglutyrtyrilevalaspasntrpglythr
859095
tyrargprothrglythrtyrlysglyservalthrseraspglygly
100105110
thrtyraspiletyrglnthrthrargtyrasnalaproservalglu
115120125
glythrargthrpheasnglntyrtrpservalargglnserlysarg
130135140
thrglyglythrilethrthrglyasnhispheaspalatrpalaarg
145150155160
alaglymetproleuglyserphealatyrtyrmetileleualathr
165170175
gluglyalaglnserserglyasnserasnilethrvalserser
180185190
<210>4
<211>576
<212>dna
<213>streptomycesrameus
<400>4
gccacggtcgtcaccacgaaccagaccggcaccgacaacggcttctactactcgttctgg60
accgacgcgcagggcacggtctcgatgaccctgggctccggtggcaactacagcaccagc120
tggcgcaacaccggcaacttcgtggccggcaagggctggagcaccggcgcccgcaggaac180
gtgacctactccggcagcttcaacccgtccggcaacggctacctgtcgctctacggctgg240
acgtcgaacccgctcgtggagtactacatcgtcgacaactggggcacctaccggcccacg300
gggacgtacaagggcagtgtcaccagtgacggcggaacgtacgacatctaccagacgacg360
cggtacaacgccccgtccgtcgagggcaccaggaccttcaaccagtactggagcgtgcgg420
cagtcgaagcgcaccggcggcaccatcaccaccggcaaccacttcgacgcctgggcccgc480
gccgggatgcccctcggcagcttcgcgtactacatgatcctcgcgaccgaggggtaccag540
agcagcggcaactccaatatcacggtgtcgtcgtga576
<210>5
<211>576
<212>dna
<213>streptomycesrameus
<400>5
gccacggtcgtcaccacgaaccagaccggcaccgacaacggcttctactactcgttctgg60
accgacgcgcagggcacggtctcgatgaccctgggctccggtggcaactacagcaccagc120
tggcgcaacaccggcaacttcgtggccggcaagggctggagcaccggcgcccgcaggaac180
gtgacctactccggcagcttcaacccgtccggcaacggctacctgtcgctctacggctgg240
acgtcgaacccgctcgtggagtactacatcgtcgacaactggggcacctaccggcccacg300
gggacgtacaagggcagtgtcaccagtgacggcggaacgtacgacatctaccagacgacg360
gcgtacaacgccccgtccgtcgagggcaccaggaccttcaaccagtactggagcgtgcgg420
cagtcgaagcgcaccggcggcaccatcaccaccggcaaccacttcgacgcctgggcccgc480
gccgggatgcccctcggcagcttcgcgtactacatgatcctcgcgaccgaggggtaccag540
agcagcggcaactccaatatcacggtgtcgtcgtga576
<210>6
<211>576
<212>dna
<213>streptomycesrameus
<400>6
gccacggtcgtcaccacgaaccagaccggcaccgacaacggcttctactactcgttctgg60
accgacgcgcagggcacggtctcgatgaccctgggctccggtggcaactacagcaccagc120
tggcgcaacaccggcaacttcgtggccggcaagggctggagcaccggcgcccgcaggaac180
gtgacctactccggcagcttcaacccgtccggcaacggctacctgtcgctctacggctgg240
acgtcgaacccgctcgtggagtactacatcgtcgacaactggggcacctaccggcccacg300
gggacgtacaagggcagtgtcaccagtgacggcggaacgtacgacatctaccagacgacg360
cggtacaacgccccgtccgtcgagggcaccaggaccttcaaccagtactggagcgtgcgg420
cagtcgaagcgcaccggcggcaccatcaccaccggcaaccacttcgacgcctgggcccgc480
gccgggatgcccctcggcagcttcgcgtactacatgatcctcgcgaccgagggggcccag540
agcagcggcaactccaatatcacggtgtcgtcgtga576
<210>7
<211>30
<212>dna
<213>artificialsequence
<220>
<223>1s11f2
<400>7
catgccatggccacggtcgtcaccacgaac30
<210>8
<211>34
<212>dna
<213>artificialsequence
<220>
<223>1s11r1
<400>8
ccgctcgagcgacgacaccgtgatattggagttg34
<210>9
<211>24
<212>dna
<213>artificialsequence
<220>
<223>121f
<400>9
atctaccagacgacggcgtacaac24
<210>10
<211>24
<212>dna
<213>artificialsequence
<220>
<223>121r
<400>10
gttgtacgccgtcgtctggtagat24
<210>11
<211>21
<212>dna
<213>artificialsequence
<220>
<223>s11tf1
<400>11
gcacggtctcgatggagctgg21
<210>12
<211>50
<212>dna
<213>artificialsequence
<220>
<223>179r
<400>12
tcacgacgacaccgtgatattggagttgccgctgctctgggccccctcgg50
- 还没有人留言评论。精彩留言会获得点赞!