雄性不育基因ZmMYB84及其在创制玉米雄性不育系中的应用
本发明属于植物生物技术育种领域,具体涉及雄性不育基因zmmyb84及其在创制玉米雄性不育系中的应用。
背景技术:
玉米是我国第一大粮食作物,常年播种面积在5.5亿亩以上,玉米种业健康发展对保障国家粮食安全有着重大的战略意义。同时,玉米种业也是全球市值最大、商业化程度最高、科技含量最高的种业领域,是全球种业竞争的战略要地。我国玉米种业与国际领先水平相比,在科技创新和产业模式等方面,仍然存在巨大差距。一是受限于玉米雄性不育基础研究尚未取得根本性突破等因素,导致玉米自交系知识产权保护困难,造成近年来我国玉米种业长期存在跟随性、模仿性育种现象,重大新品种选育效率缓慢。二是玉米制种产业仍然处于依靠人工去雄为主的劳动密集型阶段,成本高,资源消耗巨大,种子质量难以保障。
玉米是杂种优势利用最成功的作物之一,雄性不育系是作物杂种优势利用和杂交制种的重要材料,主要包括细胞质雄性不育(cms)和细胞核雄性不育(gms)。cms是由线粒体基因和核基因共同控制的,虽然已应用于玉米育种和杂交种生产中,但存在资源利用率低、不育系胞质单一、易感病等问题。gms由核基因单独控制,可以克服cms缺陷,但很难通过常规育种方法大量繁殖纯合不育系。近年来,随着生物技术的进步,通过基因工程和分子设计育种相结合创制的玉米多控不育技术以及植物通用型显性不育技术,可以有效地解决玉米隐性核雄性不育系的保持和繁殖问题。实现上述技术应用的重要前提是获得大量功能明确的控制玉米雄性发育的gms基因和对应的雄性不育材料。
与模式植物拟南芥和模式作物水稻相比,玉米中已克隆和鉴定的gms基因和创制的雄性不育材料均相对较少。crispr/cas9(clustered,regularlyinterspaced,shortpalindromicrepeats-associatedendonuclease9)基因编辑技术由于具有成本低廉、操作简易和突变诱导率高等特点,被越来越广泛地应用于植物基因功能研究,作物遗传改良和育种等多个方面,应用前景十分广阔。利用crispr/cas9技术挖掘鉴定玉米雄性不育候选基因和创制雄性不育材料,可以快速丰富玉米gms基因和不育材料资源,从而促进玉米不育化育种和制种的推广和应用,最终可以有效解决我国玉米种业行业长期面临缺乏稳定的玉米不育系和突破性大品种的瓶颈问题。
技术实现要素:
针对现有技术的不足,本发明的目的是提供雄性不育基因zmmyb84及其在创制玉米雄性不育系中的应用,可以用来创制玉米雄性不育系,从而应用于玉米杂交育种和制种。
为实现上述目的,本发明提供了zmmyb84基因在控制玉米雄性生殖发育中的应用,其特征在于,所述基因的氨基酸序列如seqidno.2所示。一般可以预见这些来自不同植物或不同玉米材料中的同源基因具有相同或者相似的功能,因此同样可以利用这些基因改良植物的农艺性状。进一步,即使不能预见这些基因的功能,本领域的一般技术人员可以根据本发明提供的方法和现有技术测定它们是否具有控制植物雄性育性的功能。
在另一方面,本发明还提供了根据权利要求1所述的应用,其特征在于,所述基因的核苷酸序列为seqidno.1所示。
在另一方面,本发明还提供了一种创制玉米雄性不育系的方法,其特征在于,抑制玉米中权利要求1或2所述基因的表达和/或活性,选择玉米雄性不育的植株。
在一些实施方案中,上述抑制蛋白表达和/或活性的方法包括基因编辑、rna干扰、t-dna插入中任一种。
在一些实施方案中,上述基因编辑采用crispr/cas9方法。
在一些实施方案中,所述crispr/cas9方法包括:在所述基因第一外显子处设计crispr/cas9载体靶点,所述靶点的dna序列如seqidno.3和seqidno.4所示。
在另一方面,本发明还提供一种获得myb84雄性不育系的方法,其特征在于,通过权利要求3-6之任一所述方法获得的myb84雄性不育系与目标材料进行杂交和回交,从而使目标材料获得myb84雄性不育的性状和基因突变。
本发明还包括通过上述任一方法获得的myb84雄性不育系在杂交育种和制种中的应用。所述在杂交育种和制种中的应用是指将myb84雄性不育系作为母本与其他父本进行杂交,或是将获得的myb84雄性不育系与其他目标材料进行杂交和回交,从而使目标材料获得myb84雄性不育的性状和基因突变。
更进一步地,本发明还提供了一种玉米雄性不育系myb84的分子标记引物,引物zmmyb84-f和zmmyb84-r序列分别如seqidno.5和seqidno.6所示。
本发明的优点及有益效果如下:zmmyb84(zm00001d025664)基因及编码的蛋白调控玉米雄性生殖发育是之前没有报道过的。本发明通过利用crispr/cas9方法突变玉米基因zmmyb84(zm00001d025664),发现了zmmyb84(zm00001d025664)基因对玉米雄穗发育的调控功能。利用crispr/cas9基因编辑的方法和编辑后获得的雄性不育突变体,可以创制玉米雄性不育系,从而可以应用于玉米杂交育种和制种。针对三种myb84的雄性不育系开发共分离分子标记,可用于植株的育性等位基因鉴定、分子标记辅助育种中目标单株的筛选和种子纯度鉴定等。
附图说明
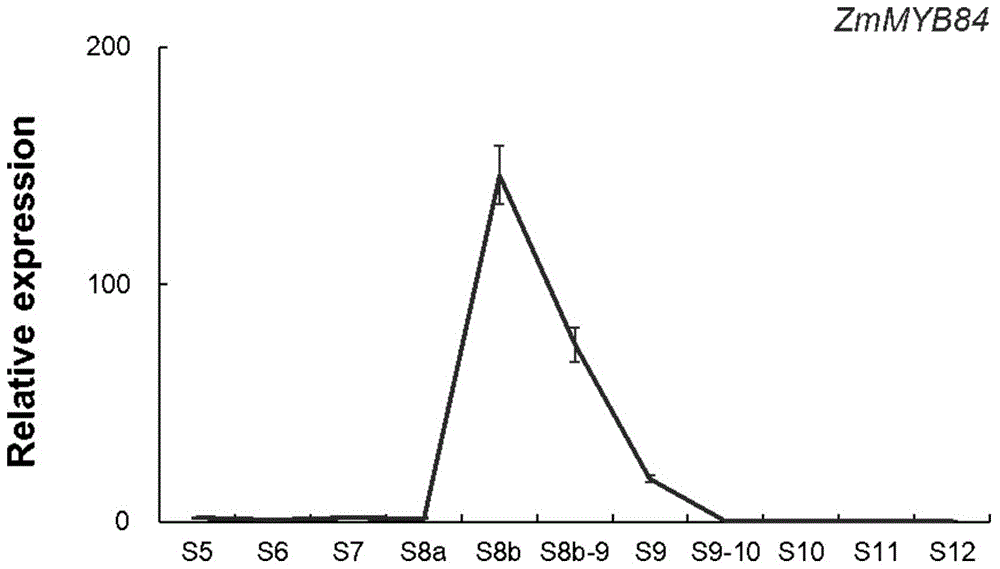
图1为zmmyb84基因在玉米不同发育时期花药中的表达模式分析
s5,造孢细胞时期;s6,小孢子母细胞时期;s7,减数分裂开始时期;s8a,减数分裂ⅰ,二分体时期;s8b,减数分裂ⅱ,四分体时期;s8b-9,四分体-单核小孢子时期;s9,单核小孢子时期;s9-10,单核小孢子-小孢子空泡化时期;s10,小孢子空泡化时期;s11,小孢子第一次不均等有丝分裂,二核小孢子时期;s12,小孢子第二次有丝分裂,三核小孢子时期。
图2为pcas9-zmmyb84定点突变表达载体的物理图谱
pcas9-zmmyb84:从t-dna的左边界到右边界分别是除草剂抗性基因bar的表达盒;核酸酶编码基因cas9的表达盒;zmmyb84基因靶标2(mt2)的表达盒;靶标1(mt1)的表达盒。
图3为野生型zmmyb84与其不育突变体的基因结构和dna序列分析
野生型zmmyb84(wt-zmmyb84):基因全长1241bp,包括3个外显子和2个内含子;myb84突变体zmmyb84-cas9-1:在第1个外显子40bp-92bp之间缺失53bp;突变体zmmyb84-cas9-2:在第1个外显子40bp-91bp之间缺失52bp;突变体zmmyb84-cas9-3:在第1个外显子40bp-93bp之间缺失54bp。
图4为野生型与myb84纯合突变体的雄穗、花药和花粉粒表型分析
上排为玉米野生型(wt)与zmmyb84-cas9-1、zmmyb84-cas9-2、zmmyb84-cas9-3突变体雄穗的表型比较;第二排为wt及与zmmyb84-cas9-1、zmmyb84-cas9-2、zmmyb84-cas9-3突变体花药的表型比较;下排为wt与zmmyb84-cas9-1、zmmyb84-cas9-2、zmmyb84-cas9-3突变体花粉粒的i2-ki染色比较。
图5为野生型与myb84纯合突变体的花药扫描电镜(sem)分析
从左至右依次为:野生型(wt)花药整体;myb84花药整体;剥开后的wt(上)和myb84(下)花药;wt的成熟花粉粒(上)和myb84无法扫描到花粉粒(下);wt(上)和myb84(下)花药的外表皮角质层;wt(上)和myb84(下)花药的内表皮乌氏体。
图6为利用共分离标记对zmmyb84-cas9-1不育系的f2代植株进行基因分型
共分离标记zmmyb84-f/r对7株zmmyb84-cas9-1不育系f2代植株的pcr和琼脂糖凝胶电泳鉴定结果:在纯合野生型(aa)植株中扩增出224bp条带;在myb84/myb84杂合型(aa)植株中扩增出224bp和171bp两条带;在myb84/myb84纯合突变型(aa)植株中扩增出171bp条带。
图7为利用共分离标记对zmmyb84-cas9-2不育系的f2代植株进行基因分型
共分离标记zmmyb84-f/r对6株zmmyb84-cas9-2不育系f2代植株的pcr和琼脂糖凝胶电泳鉴定结果:在纯合野生型(aa)植株中扩增出224bp条带;myb84/myb84杂合型(aa)植株中扩增出224bp和170bp两条带;在myb84/myb84纯合突变型(aa)植株中扩增出170bp条带。
图8为利用共分离标记对zmmyb84-cas9-3不育系的f2代植株进行基因分型
共分离标记zmmyb84-f/r对6株zmmyb84-cas9-3不育系f2代植株的pcr和琼脂糖凝胶电泳鉴定结果:在纯合野生型(aa)植株中扩增出224bp条带;在myb84/myb84杂合型(aa)植株中扩增出224bp和172bp两条带;在myb84/myb84纯合突变型(aa)植株中扩增出172bp条带。
具体实施方式
下述实施例用来说明本发明,但不限制本发明的范围。在不背离本发明精神和实质的情况下,对本发明方法、步骤或条件所作的修改或替换,均属于本发明的范围。如无特殊说明,实施例中所用引物及基因的合成和测序均由生工生物工程(上海)股份有限公司完成。其他生化试剂非特别注明外均为常规市售试剂,实施例中所用的技术手段为本领域技术人员所熟知的常规手段。
实施例一玉米zmmyb84(zm00001d025664)基因序列和表达模式分析
在maizegdb库(https://www.maizegdb.org/)中,查询到玉米zmmyb84(zm00001d025664,grmzm2g173633)基因,在b73中基因的核酸序列见seqidno.1所示,基因功能注释为myb84转录因子(myb-transcriptionfactor84-myb84),其编码蛋白包含356个氨基酸,序列如seqidno.2所示。
myb类转录因子参与植物中众多生理过程的调节,由于zmmyb84基因在玉米中的实际功能还没有公开的研究资料,为了研究该基因与玉米雄性生殖发育的关系,本发明首先利用qrt-pcr分析了该基因在玉米花药发育不同阶段的表达模式。具体步骤如下:
1、玉米花药的取样与发育时期鉴定
从玉米自交系b73处于不同发育阶段的雄穗中,按照花药的长度,收集不同长度的花药样品;每份样品采集20个长度相近的新鲜花药,其中3个固定在faa溶液中(coolaber,中国)通过树脂半薄切片实验确定具体的发育阶段,其余17个花药立即冷冻在液氮中,用于rna的提取。
利用梯度乙醇(50%、70%、90%、100%)对用于树脂切片的固定花药进行脱水,每步15-30分钟。脱水过程中花药可置于70%乙醇中长期保存;为便于后期包埋,可在90%乙醇中加入0.1%的伊红对材料进行染色;为保证脱水彻底,材料须在无水乙醇中脱水2-3次。随后进行树脂替换,将花药依次置于乙醇与spurr树脂体积比为3:1、1:1、1:3液体中2-4小时,最后置于纯树脂中过夜。树脂置换完成后,将花药置于模具中,加入200µlspurr树脂,置于烘箱中,70℃聚合过夜。随后进行修块,然后可利用德国莱卡切片机进行切片,切片厚度为2µm;用镊子夹取切好的片子,置于载玻片中央的无菌水中,42℃条件下展片过夜。将固定有样品的载玻片浸入0.1%的甲苯胺蓝染液中,染色1分钟,然后用去离子水冲洗,再置于展片台上,烘干后即可用于显微观察;也可以封片后长期保存。分析树脂切片的结果,根据玉米14个不同发育时期(stage1-stage14:s1-s14)的细胞学特点,确定每份样品的具体发育时期。
、qrt-pcr分析
用trizol试剂(invitrogen,美国)提取上述鉴定的处于不同发育时期(s5-s12)的玉米花药总rna;然后使用5xall-in-onertmastermix(abm,加拿大)合成cdna;采用tbgreen™premixextaq™(takara,日本)在quantstudio5quantstudio5real-timepcrsystem(abi,美国)上进行定量逆转录聚合酶链反应检测,扩增引物为:qmyb84-f(5’-cggagaaaggcaacgacagc-3’)和qmyb84-r(5’-cgacttcgcacgaacggtatt-3’);zmactin1为参照基因,其扩增引物为:actin1-f(5’-aaatgacgcagattatgtttga-3’)和actin1-r(5’-gctcgtagtgagggagtacc-3’);每个发育时期包括三个生物学重复,每个样品有三个技术重复;数据用2-δδct方法进行分析,并以均值±标准差(means±sd)的形式给出定量结果。
zmmyb84基因呈现花药发育时期特异表达的模式:在玉米花药发育的早期和晚期几乎没有表达,而在玉米花药发育的中后期(s8b)有一个非常高的表达峰值,随后表达迅速减弱(图1)。
实施例二玉米zmmyb84(zm00001d025664)基因的功能以及利用crispr/cas9方法创制玉米雄性不育系
为了明确玉米zmmyb84(zm00001d025664)在玉米中的功能,本发明采用crispr/cas9基因编辑方法突变zm00001d025664基因序列,敲除该基因在玉米中的功能。本发明选取玉米杂交种hiii作为基因编辑的受体材料。本发明分别选取基因保守区的seqidno.3和seqidno.4所示序列作为crispr/cas9基因编辑的靶标区域。
、zmmyb84的crispr/cas9基因编辑载体的构建
本发明的基因编辑载体为pbue411-mt1t2-cas9,该载体的基础载体为pbue411-cas9,中间载体为pcbcmt1t2,提供grna。本发明通过在引物上设计靶点然后通过pcr获得mt-sgrna进而通过酶切连接到基础载体中,具体的构建流程如下:
(1)靶标grna的设计。将zmmyb84(zm00001d025664)的基因序列输入http://crispr.hzau.edu.cn/cgi-bin/crispr2/crispr进行靶标设计。本发明选择的两个靶标区域的dna序列如seqidno.3和seqidno.4所示。本发明的sgrna骨架序列从中间载体pcbcmt1t2直接扩增获得。
(2)通过在引物上设计靶点然后pcr扩增获得mt-sgrna。引物zmmyb84-mt1-f和引物zmmyb84-mt2-r扩增中间载体pcbcmt1t2,用于获得包含第一个和第二个靶标的sgrna的片段,产物长度为891bp。pcr体系和条件如下:模板dna(中间载体pcbcmt1t2≥30ng/μl)1.2μl;primerf/r:各1.2μl;灭菌ddh2o:11.4μl;2xmclab酶(产品编号:i5hm-200):15μl。pcr的温度程序如下:①98℃2分钟;②98℃10秒;③58℃30秒;④72℃30秒;⑤从②-④循环34次;⑥72℃5分钟;⑦25℃10分钟。最后回收pcr产物。载体构建所需的引物序列如下:
zmmyb84-mt1-f:5’-atatatggtctctggcgagaaggacagcgtcac
aagcgcgttttagagctagaaatagcaa-3’
zmmyb84-mt2-r:5’-attattggtctctaaactactgggtgatgtagg
agatgcttcttggtgccgc-3’
(3)通过酶切连接构建到骨架载体。将pbue411-cas9载体和回收的带有靶标的sgrna片段用bsai消化,同时加入t4连接酶将载体和sgrna片段连接。15μl的酶切连接体系如下,sgrna片段:2μl,pbue411-cas9载体(≥60ng/μl):2μl,10xnebbuffer:1.5μl,bsai内切酶(产品编号:#r3733s):1μl,t4连接酶(产品编号:#m0202m):1μl,灭菌ddh2o:6μl。
图2所示为目的基因zmmyb84(zm00001d025664)的双靶标(对应第一个靶标和第二个靶标),标记基因cas9和bar与骨架载体pbue411-cas9构建的表达载体pcas9-zmmyb84。
、农杆菌介导的玉米遗传转化
将上述构建的pcas9-zmmyb84通过热激法转入农杆菌eha105中,pcr进行鉴定;然后将含有敲除载体的农杆菌加甘油混合,于-80℃保存菌液。以新鲜剥离的1.5mm左右的玉米杂交种hiii的幼胚作为受体材料,将剥离的玉米胚放入含有1.8ml悬浮液的2ml塑料离心管中,放置时间不超过1小时,每个离心管中大约放入100个幼胚;吸去悬浮液,并用新的悬浮液将幼胚清洗2遍,管底保留少量可没过幼胚的悬浮液,然后43℃热激2分钟,之后再冰浴1分钟,用移液枪吸尽管底残留的洗液,并加入1.0ml的农杆菌侵染液,轻摇30秒,然后黑暗静置8分钟。接下来将离心管中的幼胚和侵染液倒入共培养基上,晃匀后用移液枪吸出多余的侵染液,所有幼胚的盾片朝上,于23℃黑暗共培养3天。共培养结束后,用无菌的镊子将幼胚转移至恢复培养基,于28℃培养7-14天,中间过程需留意及时去掉幼胚上长出的幼芽。恢复培养结束后将幼胚放至含1.5mg/lbialaphos筛选培养基上筛选培养3轮,每轮筛选2周,然后转到2mg/lbialaphos筛选培养基上筛选培养2轮,每轮筛选2周。将抗性愈伤组织转至扩繁培养基,28℃,暗培养2周。随后将扩繁好的抗性愈伤组织转移至诱导培养基,于28℃黑暗环境下培养2周。然后转到分化培养基中,25℃,5000lx,光照培养2周。培养结束后将分化出的苗簇分离出单个幼苗放置于生根培养基中,25℃,5000lx,光照培养直到生根;将小苗转入小营养钵中生长,生长成活后移栽于温室中,3-4个月后收获后代种子。
、t0代植株crispr/cas9突变结果检测
为确定t0代植株crispr/cas9突变结果,采取如下步骤进行:
本发明首先采用ctab法提取玉米叶片dna,具体方法如下:剪取2厘米左右长度的幼苗叶片,放入装有钢珠的2ml离心管;将装有叶片的离心管放入液氮浸没5分钟,然后利用研磨仪打碎叶片样品;向离心管中加入700μlctab提取缓冲液(其中含有1%的β-巯基乙醇),并用力震荡混匀,65℃恒温水浴中预热20-30min,(期间取出颠倒1-2次,并注意实验样品编号的对应);离心管冷却至室温后加入700μl氯仿:异戊醇(24:1)萃取液,用力摇晃30s之后在室温条件下静置片刻;在4℃条件下,以12000rpm速度离心5min,取离心完成后的500μl上清液于新的1.5ml离心管中;将等体积的异丙醇加入到盛有上清液的离心管中轻轻震荡混匀,室温条件下静置10min左右;随后将盛有样品的离心管放入4℃离心机中,以12000rpm速度离心10min之后,轻轻吸取上清液,弃上清,保留沉淀;加入800μl75%乙醇,洗涤沉淀两次,以10000rpm速度离心5min,弃掉上清;将样品放置在室温下自然干燥2-4小时,得到dna沉淀,加入适量无菌水溶解,轻微震荡,充分溶解dna。dna样品存于-20℃保存。用nanodrop检测dna浓度,稀释至10ng/l用作pcr模板。
然后根据zmmyb84(zm00001d025664)基因序列设计pcr引物。
检测靶标:mt1和mt2;产物大小:224bp;引物序列如下:
zmmyb84-t-f:5’-taccactccgagacagcag-3’;
zmmyb84-t-r:5’-gaggaactacacgacacgc-3’。
提取基因组dna,按照以下pcr参数扩增:
反应体系:15μlmix常规pcr体系,0.5μlforwardprimer,0.5μlreverseprimer,1μldna,5.5μl灭菌ddh2o,7.5μl2xtaqmix(产品编号:10103es)。
反应程序:常规pcr:58℃退火、延伸30s、32轮循环。
接着将pcr产物回收并连接t载体测序,通过测序多个t0代独立阳性转化事件靶标区域的dna序列,明确靶标区域是否发生基因编辑,最终发现3个t0转化事件靶标区域序列发生了变化,且均为纯合突变,编辑前后的序列如图3所示,对应3个myb84纯合突变体:zmmyb84-cas9-1、zmmyb84-cas9-2、zmmyb84-cas9-3。与野生型序列比对显示zmmyb84-cas9-1、zmmyb84-cas9-2和zmmyb84-cas9-3在靶标1和2处都发生了缺失突变。
对3个myb84纯合突变体中氨基酸序列进行比对分析发现,与未编辑的wt相比,突变后的品系zmmyb84-cas9-1和zmmyb84-cas9-2,其编码的的核苷酸在靶标1或2处的缺失使其氨基酸发生了移码突变,且随后的氨基酸发生提前终止。另外zmmyb84-cas9-3品系中编码的核苷酸在靶标1和2处的缺失使得发生了18个氨基酸的丢失。因此这些转化体中zm00001d025664蛋白的功能均表现缺失。
、f1代植株的基因分型
由于在温室生长的玉米t0代植株,经常雌穗和雄穗发育不协调,同时当编辑的基因与雄性发育相关时也会影响育性,因此为了繁殖t0代植株,并使获得的基因编辑类型遗传下去,本发明使用玉米自交系郑58的野生型花粉为上述获得的zmmyb84-cas9-1、zmmyb84-cas9-2、zmmyb84-cas9-3的t0代植株授粉,进而获得f1代种子,生长的植株为f1代植株。
f1代植株包括2种分离类型,一种为cas9-阳性植株(转基因植株),另一种为cas9-阴性植株(非转基因植株),为了避免sgrna和cas9对杂交授粉引入的郑58野生型等位基因进行持续编辑,从而造成突变类型的复杂性,我们需要通过基因分型从f1代植株中挑选出不含有cas9基因,但含有t0代突变类型的植株,这类植株自交后可以得到非转基因的f2代。f1代植株的基因分型步骤如下:
按照上述的ctab法提取叶片dna后,首先利用cas9基因的特异引物cas9-f(5’-cccggacaatagcgatgt-3’)和cas9-r(5’-gagtgggccgacgtagta-3’)进行pcr扩增。pcr反应体系同上;反应程序:常规pcr:58℃退火、延伸1分钟、32轮循环。pcr产物进行琼脂糖凝胶电泳后,根据结果区分出cas9-阳性植株和cas9-阴性植株。
进一步针对cas9-阴性植株,采用上述的检测mt1和mt2靶标的引物zmmyb84-t-f和zmmyb84-t-r进行pcr扩增;pcr产物纯化后,连接t载体,进行测序;根据测序结果分析确定t0代突变类型的遗传情况。
实施例三myb84不育系的表型分析
上述实施例二鉴定的不含有cas9基因的f1代植株自交后获得f2代种子,三种myb84突变类型(zmmyb84-cas9-1、zmmyb84-cas9-2和zmmyb84-cas9-3)各取1个自交单穗进行穗行播种,在成熟期进行表型的调查。三种f2株系中,可育株与不育株的比例,均符合3:1分离,进一步表明myb84不育系的不育性状由单个隐性基因控制,然后针对f2代获得的稳定非转基因myb84不育系与野生型进行详细的表型比较。
、雄穗、花药和花粉活力的观察
在营养生长和雌穗的发育方面,myb84不育系(zmmyb84-cas9-1、zmmyb84-cas9-2和zmmyb84-cas9-3)的植株与野生型相比基本无差异;在雄穗发育方面,野生型能够正常抽雄,花药能够正常开裂、散粉,自交后可正常结实,而myb84不育系虽能正常抽雄,但是不能正常开花,花药颖壳不开裂,花药明显较小,并且发白干瘪不外露(图4);进一步对野生型和突变体的花粉进行i2-ki染色,发现野生型花粉发育正常,花粉粒染色后呈黑色,但突变体无花粉粒形成,并且突变体的花药在长度和体积上也小于野生型(图4)。这表明zmmyb84(zm00001d025664)基因控制玉米的雄性发育,通过基因编辑方法创制的myb84不育系为无花粉型不育系,呈现完全败育的特点。
、花药的扫描电镜(sem)观察
为了深入解析myb84的细胞学特征,对野生型和突变体花药内外壁进行扫描电镜(sem)分析。剥取处于成熟期(s13)的野生型和突变体花药,然后立即固定在faa(coolaber,中国)溶液中,固定液的体积不少于所取研究材料体积的20倍;对于突变体花药,可用解剖针在花药壁穿孔以提高固定液的渗透效果,或者反复抽真空至花药沉入固定液底部;室温固定2小时后,将材料置于4℃保存,或依次置于50%、60%、70%、80%、90%、100%的乙醇中进行脱水,每个梯度保持15分钟;材料可置于70%的乙醇中过夜或保存。脱水后的样品进行二氧化碳临界点干燥,然后镀金即可进行观察。发现myb84突变体的花药外表皮的角质层结构相比于野生型的外表皮更加致密;但是myb84突变体花药的内表皮表现的十分光滑,没有致密的颗粒状的乌氏小体形成(图5)。花药角质层是覆盖在花药表面的细胞外脂质层,保护花药免受外部非生物胁迫、内部组织失水和病原体的侵袭,位于花药内壁的乌氏体,被认为是孢粉素前体从绒毡层细胞到小孢子的转运载体。上述结果表明zmmyb84(zm00001d025664)基因突变后,可以阻断绒毡层内孢子花粉素前体物质的合成,并且该基因也可能参与调节花药角质层的形成。
实施例四myb84不育系鉴定的共分离分子标记开发和应用
1、共分离分子标记的开发
在本发明中,针对获得的三种myb84不育系的突变位点,利用primer5.0软件进行引物设计,开发出一对共分离分子标记:zmmyb84-f/r,结合pcr与琼脂糖凝胶电泳检测的方法,根据获得的条带及大小即可分离出突变体的基因型。
共分离分子标记zmmyb84-f/r包括第一引物zmmyb84-f和第二引物zmmyb84-r;该标记能够特异性检测玉米zmmyb84-cas9-1、zmmyb84-cas9-2和zmmyb84-cas9-3突变体及由它们转育的玉米不育材料中的突变基因myb84,并能同时区分野生型myb84基因和突变型myb84基因;针对突变基因myb84分别扩增出171bp、170bp和172bp的条带,而对野生型myb84基因扩增出224bp的条带。引物序列如下:
zmmyb84-f:5’-taccactccgagacagcag-3’
zmmyb84-r:5’-gaggaactacacgacacgc-3’
2、共分离分子标记的应用
为了验证上述标记的有效性,以实施例三中获得的f2株系为材料,进行myb84等位基因的检测。dna提取方法、pcr扩增体系和条件同实施例二,pcr产物进行琼脂糖凝胶电泳分离。
理论上,zmmyb84-f/r在myb84/myb84纯合野生型(aa)dna中能扩增224bp的条带,在myb84/myb84纯合突变型材料(aa)dna中分别扩增出171bp、170bp和172bp的条带,而在myb84/myb84杂合型(aa)材料中,则能分别同时扩增出对应的两条条带。针对三种突变类型的f2植株,zmmyb84-f/r分子标记的验证结果如图6、图7和图8所示,结果显示设计的功能性分子标记对f2植株的检测结果完全符合预期,在myb84/myb84纯合野生型(aa)、myb84/myb84杂合型(aa)和myb84/myb84纯合突变型材料(aa)中分别扩增出对应大小的条带,可以作为myb84,myb84等位基因检测的理想标记。
这些分子标记有助于在开花和授粉前确定突变基因型,以便在不同遗传背景下进行杂交和回交选育雄性不育系,具有重要的应用价值。
序列表
<110>北京科技大学
北京中智生物农业国际研究院
北京首佳利华科技有限公司
<120>雄性不育基因zmmyb84及其在创制玉米雄性不育系中的应用
<160>6
<170>siposequencelisting1.0
<210>1
<211>1241
<212>dna
<213>玉米(zeamays)
<400>1
atggggcggatcccgtgctgcgagaaggacagcgtcaagcgcgggcagtggacgcccgag60
gaggacaacaagctgctctcctacatcacccagtacggcacgcgcaactggcgcctcatc120
cccaagaatgccggtacgtcctagcgaccgtccaatgtccgcgtgtcgtgtagttcctct180
caagtgtgaccgagtccgcgtgatgggattgcaggactgcagcgatgcgggaagagctgc240
cggctccggtggaccaactacctgcgtcccgacctcaagcacggtgagttcacggacacc300
gaggagcagaccatcatcaagctgcactccgtcgttggcaacaggtggcccgcccgtcct360
cccctctcgtcgtgcctcttctgatgggtgcctgttctgactacaagcaatgtgcaacaa420
acgcgcgcgcgtgcaggtggtcggtgatcgcggcgcagctgccgggtcggacggacaacg480
acgtcaagaaccactggaacaccaagctgaagaagaagctgtccgggatgggcatcgacc540
ccatcacgcacaagtccttctcgcacctcatggccgagatcgccaccacgctggcgccgc600
cgcaggtggcccacctcgccgaggccgcgctggggtgcttcaaggacgagatgctccacc660
tcctcaccaagaagcgccccaccgacttcccgtcgcccgcggtgcccgacatgtcggcga720
tcgcgggcggctccggcgtcgcggcgccctgcggcttcccggcgccgccccagaccgacg780
acaccatcgagcgcatcaagctgggcctgtcccgcgccatcatgagcgagcccgccgcgc840
cccccggcaagcaggagcagccctgggcgccggccgacttgccggaggggctgccgggga900
tgtacgccacgtacaatcccgcctcgcacggacacgaagagttccgctacgacaacggga960
cagtgccggagtacgtcctcggcggcggcggcggcgcggaccagggcacgtcgatgtgga1020
gccaccagagcatgtacagcgggagttcggccacggaggccgcgcccaggccggcggagg1080
tgttgccggagaaaggcaacgacagcgtcgggagcagcggcggcggcgaggaggcggacg1140
acgtcaaggacggcgggaaaggcggctccgatatgtccggcctgtttggatccgactgcg1200
tactttgggacttgcccgacgagctgaccaatcacatggtg1241
<210>2
<211>356
<212>prt
<213>玉米(zeamays)
<400>2
metglyargileprocyscysglulysaspservallysargglygln
151015
trpthrproglugluaspasnlysleuleusertyrilethrglntyr
202530
glythrargasntrpargleuileprolysasnalaglyleuglnarg
354045
cysglylyssercysargleuargtrpthrasntyrleuargproasp
505560
leulyshisglygluphethraspthrglugluglnthrileilelys
65707580
leuhisservalvalglyasnargtrpservalilealaalaglnleu
859095
proglyargthraspasnaspvallysasnhistrpasnthrlysleu
100105110
lyslyslysleuserglymetglyileaspproilethrhislysser
115120125
pheserhisleumetalagluilealathrthrleualaproprogln
130135140
valalahisleualaglualaalaleuglycysphelysaspglumet
145150155160
leuhisleuleuthrlyslysargprothrasppheproserproala
165170175
valproaspmetseralailealaglyglyserglyvalalaalapro
180185190
cysglypheproalaproproglnthraspaspthrilegluargile
195200205
lysleuglyleuserargalailemetsergluproalaalapropro
210215220
glylysglngluglnprotrpalaproalaaspleuprogluglyleu
225230235240
proglymettyralathrtyrasnproalaserhisglyhisgluglu
245250255
pheargtyraspasnglythrvalproglutyrvalleuglyglygly
260265270
glyglyalaaspglnglythrsermettrpserhisglnsermettyr
275280285
serglyserseralathrglualaalaproargproalagluvalleu
290295300
proglulysglyasnaspservalglyserserglyglyglygluglu
305310315320
alaaspaspvallysaspglyglylysglyglyseraspmetsergly
325330335
leupheglyseraspcysvalleutrpaspleuproaspgluleuthr
340345350
asnhismetval
355
<210>3
<211>19
<212>dna
<213>人工序列(artificialsequence)
<400>3
gaaggacagcgtcaagcgc19
<210>4
<211>19
<212>dna
<213>人工序列(artificialsequence)
<400>4
tctcctacatcacccagta19
<210>5
<211>19
<212>dna
<213>人工序列(artificialsequence)
<400>5
taccactccgagacagcag19
<210>6
<211>19
<212>dna
<213>人工序列(artificialsequence)
<400>6
gaggaactacacgacacgc19
- 还没有人留言评论。精彩留言会获得点赞!