CAS9-核酸复合物及其相关用途
本申请是申请日为2014年1月15日,申请号为“201480008880.5”,发明名称为“cas9-核酸复合物及其相关用途”的申请的分案申请。
相关申请的交叉引用
本申请要求2013年1月16日提交的美国临时申请号61/753,046和2013年11月18日提交的美国临时申请号61/905,368的优先权,两者都通过引用以其全文特此结合。
领域
本披露涉及cas9-核酸复合物及其相关用途。在某些实施例中,本披露设想了被遗传工程化以表达在此披露的cas9-核酸复合物的转基因植物和动物。在某些实施例中,本披露涉及使用被配置成表达在此披露的cas9-核酸复合物的载体治疗或预防疾病、病症、癌症、病毒感染、或其他病原性感染的方法。
背景
crispr(成簇的规律间隔的短回文重复序列)-cas(crispr-相关)基因提供针对外源核酸的防御。这些系统利用由重复序列侧翼间隔区组成的小crisprrna(crrna)的阵列来识别它们的靶标并且利用某些cas蛋白来介导靶向降解。参见黑尔(hale)等人,细胞(cell),2009,139,945-956;加西乌纳斯(gasiunas)等人,美国国家科学院院刊(procnatlacadsciusa),2012,109,e2579-2586;季聂克(jinek)等人,科学(science),2012,337,816-821;以及迪杜森科(datsenko)等人,自然通讯(natcommun),2012,3,945。加尔诺(garneau)等人,自然(nature),2010,468,67-71报道了crispr/cas细菌免疫系统裂解噬菌体和质粒dna。巴朗格(barrangou)等人,科学(science),2007,315,1709-1712报道了crispr在原核生物中提供针对病毒的获得性抗性。marraffmi(马拉弗米)和宗特海默尔(sontheimer),科学(science),2008,322,1843-1845报道了crispr干扰限制葡萄球菌中通过靶向dna进行的水平基因转移。
霍瓦特(horvath)等人,wo2007025097,报道了一种或多种cas基因或蛋白质用于调节细胞对靶核酸或其转录产物的抗性的用途。
黑尔等人报道了与casramp模块复合物一起作用来裂解rna的crisprrna的基本特征和合理设计。分子细胞(molecularcell)201245,292-302。
丘(cho)等人报道了使用cas9rna指导的内切核酸酶进行人细胞中靶向基因组工程化。自然生物技术(naturebiotechnology),2013,31,230-232。
马里(mali)等人报道了通过cas9进行的rna指导的人基因组工程化。科学(science),2013,339:823-26。还参见季聂克等人,elife,2013,2:e00471。
涅克拉索夫(nekrasov)等人报道了使用cas9rna指导的内切核酸酶进行模式植物本氏烟草的定向诱变。自然生物技术(natbiotechnol),2013,31(8):691-3。
在此引用的参考文献并非对现有技术的承认。
概述
本披露涉及cas9-核酸复合物及其相关用途。在某些实施例中,本披露设想了被遗传工程化以表达在此披露的cas9-核酸复合物的转基因植物和动物。在某些实施例中,本披露涉及使用被配置成表达在此披露的cas9-核酸复合物的载体治疗或预防疾病、病症、癌症、病毒感染、或其他病原性感染的方法。
在某些实施例中,本披露涉及使用被配置成表达靶向病毒或病原性核酸或与致癌基因相关联的rna的cas9-核酸复合物的载体治疗或预防癌症或病毒感染或其他病原性感染或其他遗传性疾病的方法。在某些实施例中,本披露设想了被遗传工程化以表达在此披露的cas9-核酸复合物,以用于癌症、遗传性疾病、预防或治疗病毒或其他病原性感染目的的转基因植物和动物。
在某些实施例中,本披露涉及分离或重组核酸、克隆载体、以及含有它们的重组细胞。在某些实施例中,本披露涉及治疗或预防病毒感染或癌症或其他遗传性疾病的方法,这些方法包括向对其有需要的受试者给予有效量的被配置成表达靶向病毒核酸或与致癌基因相关联的rna的cas9-核酸复合物的载体。
在某些实施例中,本披露设想了敲除内源性细菌或其他基因或防止在原核、真核、哺乳动物、人、昆虫或植物细胞中产生靶蛋白的组合物和方法。在某些实施例中,本披露涉及免疫刺激组合物以及如在此所述的用途。
在某些实施例中,本披露涉及包含以下的重组核酸:一个包含cas9或细菌cas9基因的序列;一个编码rna的序列,其中该rna包含被配置成在转录之后与该cas9结合的一个第一区段和被配置成结合靶核酸的一个第二区段。在某些实施例中,细菌cas9mrna翻译具有seqidno:1的cas9或其保守变体。在某些实施例中,cas9具有一个富含精氨酸的基序、一个ruvc-iii基序、以及一个ruvc-iv基序。在某些实施例中,cas9mrna翻译与seqidno:1具有大于约5%的一致性的cas9、与seqidno:6具有10%一致性的区段、与seqidno:7具有10%一致性的区段、以及与seqidno:8具有10%一致性的区段。在某些实施例中,该第一区段包含seqidno:5或seqidno:11或与其具有60%或更大的一致性。
在某些实施例中,第一区段包含与被配置成结合细菌cas9的tracrrna或scarna相关联的细菌衍生的序列。在某些实施例中,该第一区段形成一个发夹结构。在某些实施例中,该靶序列为病毒基因组或病毒rna,或与致癌基因相关联的mrna或微rna。在某些实施例中,rna的该第二区段是单链的。在某些实施例中,第二区段包含多于10、15、20、25、30、50、或100个被配置成与靶序列杂交的连续核苷酸。在某些实施例中,cas9基因为人、动物或植物编码优化序列。在某些实施例中,cas9基因包含(seqidno:9)或与其具有60%或更大的一致性。
在某些实施例中,本披露设想了包含以下的重组核酸:一个包含cas9或细菌cas9基因的序列;一个编码seqidno:5或seqidno:11或与其具有10%、30%、60%、70%、80%、90%、95%或更大一致性的缀合编码第三rna的序列的序列,其中该第三rna包含8个被配置成与靶序列杂交的连续核苷酸。
在某些实施例中,本披露设想了包含以下的重组核酸:一个编码单个嵌合rna5'-[x]n
在某些实施例中,靶序列为病毒基因组或rna,或与致癌基因相关联的mrna或微rna。在某些实施例中,第三rna包含多于10、15、20、25、30、50、或100个被配置成与靶序列杂交的连续核苷酸。在某些实施例中,cas9或细菌cas9基因为人密码子优化序列。在某些实施例中,cas9基因包含(seqidno:9)或与其具有10%、30%、50%、60%、70%、80%、90%、95%、98%或更大的一致性。
在某些实施例中,本披露涉及包含在此披露的核酸的重组载体。重组载体可选自遗传工程化的质粒、噬菌体、细菌人工染色体、酵母人工染色体、或遗传工程化的病毒。
在某些实施例中,本披露涉及用在此披露的重组载体转化的细菌、原核、真核、昆虫、哺乳动物、或植物细胞。
在某些实施例中,本披露涉及包含以下的分离或重组核酸:一个编码细菌或任何cas9mrna的序列、一个编码细菌scarna的序列、以及一个编码与启动子序列可操作结合的第三rna的序列,其中编码该第三rna的该序列的一部分与该scarna杂交并且其中编码该第三rna的该序列的第二部分与靶序列杂交。
在某些实施例中,本披露涉及包含以下的分离或重组核酸:一个编码cas9或细菌cas9mrna的序列和一个编码连接至编码第三rna的序列的细菌scarna的一部分的序列,该第三rna与靶序列杂交以提供rna嵌合体,其中该rna嵌合体提供scarna和靶向rna的功能。
在某些实施例中,分离核酸是cdna。
在某些实施例中,cas9mrna翻译具有seqidno:1的cas9或其变体。
在某些实施例中,cas9具有一个富含精氨酸的基序、一个ruvc-iii基序、以及一个ruvc-iv基序。
在某些实施例中,cas9mrna翻译与seqidno:1具有大于约5%、10%、20%、30%、40%、50%、60%、70%、80%、90%、或95%一致性的cas9。
在某些实施例中,cas9具有一个富含精氨酸的基序,该基序与mnnrtarrhqrrgidrkqlvk(seqidno:6)具有大于约10%、20%、30%、40%、50%、60%、80%、90%、或95%的一致性。
在某些实施例中,cas9具有一个ruvc-iii基序,该基序与knivddnwqnikqvlsakhqlhipiitesnafefe(seqidno:7)具有大于约10%、20%、30%、40%、50%、60%、80%、90%、或95%的一致性。
在某些实施例中,cas9具有一个ruvc-iv基序,该基序与akgdkpqasyshlidamlafciaadehrndg(seqidno:8)具有大于约10%、20%、30%、40%、50%、60%、80%、90%、或95%的一致性。
在某些实施例中,scarna包含guuguxuagauuauuugguauguacuuguguuaguuuaaaguagxxcuagaaaauucacuuuuagaccuacuuauuuu(seqidno:3),其中x在每次出现时独立地为任何核苷酸。
在某些实施例中,scarna与seqidno:3具有大于约50%、60%、70%、80%、90%、或95%的一致性。
在某些实施例中,rna的与scarna杂交的部分包含guaccaaauaauu(seqidno:5)。
在某些实施例中,该rna包含guaccaaauaauu[x]n(seqidno:14),其中x在每次出现时独立地为任何核苷酸,并且n为10、20、50、100、200、或更多个核苷酸,通常为少于100、200、或500个核苷酸。
在某些实施例中,本披露设想了包含在此披露的任何核酸序列的重组载体。
在某些实施例中,rna的与靶序列杂交的第二部分,例如[x]n大于约10、20、50、100、200、400、或800个核苷酸。
在某些实施例中,本披露涉及在此披露的分离核酸,其进一步编码标志物多肽,如抗体表位、配体、聚组氨酸、赋予抗生素抗性的蛋白质、分解抗生素的酶如β-内酰胺酶,或荧光蛋白如绿色荧光蛋白。
在某些实施例中,本披露涉及包含在此披露的核酸的克隆载体。在某些实施例中,该克隆载体选自遗传工程化的质粒、噬菌体、细菌人工染色体、酵母人工染色体、或病毒。
在某些实施例中,本披露涉及用在此披露的克隆载体转化的重组细菌细胞。
在某些实施例中,本披露设想了制备重组细菌细胞的方法,这些方法包括将在此披露的克隆载体与细菌细胞混合,其条件使得该克隆载体的核酸包含整合到该细菌细胞的基因组中的编码序列。
在某些实施例中,本披露涉及减少靶多肽的翻译的方法,这些方法包括将细菌、原核、真核、植物、昆虫、或哺乳动物细胞与在此披露的克隆载体混合,其条件使得发生编码序列的转录、发生cas9的翻译并且形成核酸复合物,其中细菌、原核、真核、植物、昆虫、或哺乳动物细胞翻译靶多肽,其中第三rna的与靶rna(例如rrna、非编码rna或编码靶多肽且翻译靶蛋白的mrna)杂交的第二部分被减少或靶向rna被降解。
在某些实施例中,靶多肽具有未知的功能。在某些实施例中,本披露设想可形成靶向rna和/或细菌的文库和阵列,以确定未知rna转录物的功能。第三rna的第二部分可被工程化,以与具有未知功能的靶rna序列(例如mrna、rrna或非编码rna)杂交。
在某些实施例中,本披露涉及编码包含以下的蛋白-核酸复合物的载体:cas9多肽、形成双链发夹且包含一部分单链rna的scarna;一部分包含该单链rna部分的互补序列的rna,并且该rna的第二部分与靶序列例如rna杂交。在某些实施例中,可将载体转移到细菌或原核或真核细胞中,其条件使得形成复合物。靶向序列的杂交防止具有未知功能的rna转录物例如mrna执行其预期功能并且对细菌的表型进行分析以确定敲除的功效。在某些实施例中,第三rna以及scarna和cas9复合物的靶向导致靶向rna的降解或杂交阻止翻译。独立地随机筛选大量的具有未知功能的rna转录物可用于鉴定生长、复制、或其他性状所必需的rna转录物。
在某些实施例中,本披露涉及包含以下的分离蛋白-核酸复合物:cas9或细菌cas9多肽、形成双链发夹且包含一部分单链rna的scarna;一部分包含该单链rna部分的互补序列的rna,并且该rna的第二部分与靶序列杂交,其中该单链rna部分与互补序列杂交以形成rna复合物;并且其中cas9或细菌cas9与该rna复合物结合,以形成蛋白-核酸复合物。
在某些实施例中,本披露涉及包含细菌菌株的免疫刺激组合物,该细菌菌株具有突变的cas9、scarna、或tracrrna基因、或其组合。在某些实施例中,突变处于cas9或细菌cas9富含精氨酸的基序、ruvc-iii基序以及ruvc-iv基序中。在某些实施例中,突变为氨基酸、多肽或区段的改变或缺失。在某些实施例中,突变是scarna或区段的缺失、tracrrna或区段的缺失、cas9或区段的缺失、或在scarna中产生反向互补序列或在tracrrna中产生反向互补序列突变。
在某些实施例中,本披露涉及免疫受试者以抗细菌菌株的方法,这些方法包括以有效量向受试者给予在此披露的免疫刺激组合物。
在某些实施例中,本披露设想了在此披露的cas9系统在任何原核、真核、人、哺乳动物、或植物细胞中的使用。
具体地,本申请提供了以下内容:
1.一种重组核酸,该核酸包含:
一个包含cas9基因的序列,
一个编码rna的序列,其中该rna包含被配置成在转录之后与该cas9结合的一个第一区段和被配置成结合靶核酸的一个第二区段。
2.如项目1所述的重组核酸,其中该cas9是一种细菌cas9。
3.如项目2所述的重组核酸,其中该细菌cas9mrna翻译具有seqidno:1的细菌cas9或其保守变体。
4.如项目3所述的重组核酸,其中该细菌cas9具有一个富含精氨酸的基序、一个ruvc-iii基序、以及一个ruvc-iv基序。
5.如项目1-4所述的重组核酸,其中该细菌cas9mrna翻译与seqidno:1具有大于约5%的一致性的一个细菌cas9、与seqidno:6具有10%一致性的一个区段、与seqidno:7具有10%一致性的一个区段、以及与seqidno:8具有10%一致性的一个区段。
6.如项目1所述的重组核酸,其中该第一区段包含seqidno:5或seqidno:11或与其具有60%或更大的一致性。
7.如项目1所述的重组核酸,其中该第一区段形成一个发夹结构。
8.如项目1所述的重组核酸,其中该靶序列为一个病毒基因组或病毒rna,或与癌基因相关联的mrna或微rna。
9.如项目1所述的重组核酸,其中rna的该第二区段是单链的。
10.如项目1所述的重组核酸,其中该第二区段包含多于10、15、20、25、30、50、或100个被配置成与靶序列杂交的连续核苷酸。
11.如项目1所述的重组核酸,其中该cas9基因是人、动物、或植物编码优化序列。
12.如项目11所述的重组核酸,其中该cas9基因包含(seqidno:9)或与其具有60%或更大的一致性。
13.一种包含如项目1-12所述的核酸的重组载体。
14.如项目6所述的重组载体,选自遗传工程化的质粒、噬菌体、细菌人工染色体、酵母人工染色体、或病毒。
15.一种用如项目14所述的重组载体转化的细菌、原核、真核、昆虫、哺乳动物、或植物细胞。
附图简述
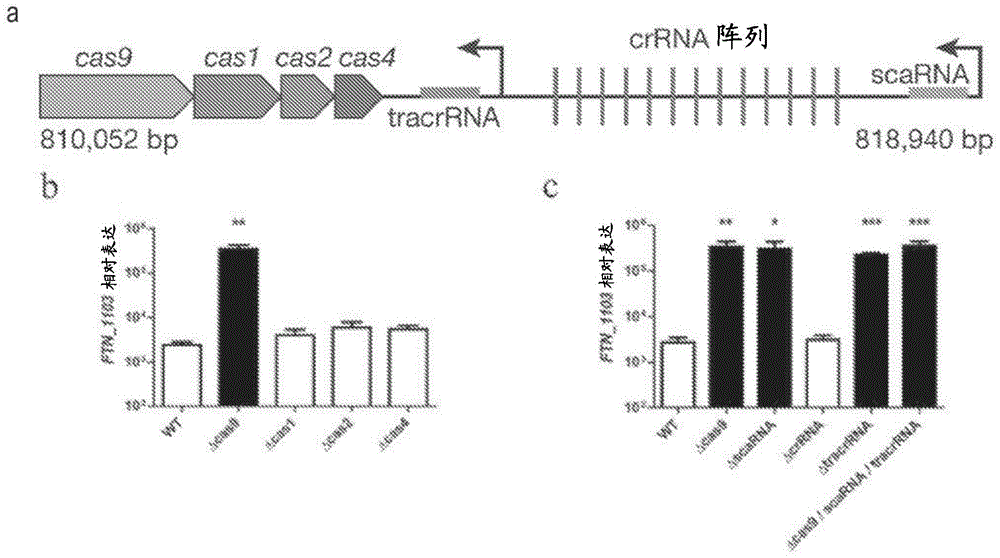
图1示出表明cas9、tracrrna、以及scarna对于ftn_1103阻遏非常重要的数据。(a)新杀手弗朗西斯菌(f.novicida)ii型crispr-cas基因座的示意图,其包含cas9、casl、cas2和cas4,以及crrna阵列(由竖直红线指示的重复序列)、tracrrna(蓝色)、scarna(灰色)、以及预测启动子(黑色箭头)。(b)野生型(wt)、δcas9、δcas1、δcas2、以及δcas4菌株中ftn_1103的相对表达,以及(c)wt、δcas9、δscarna、δcrrna、以及δtracrrna菌株中ftn_1103的相对表达(n=4,条线表示标准偏差)。
图2-1和2-2示出表明cas9、tracrrna、以及scarna与ftn_1103降解相关并介导ftn_1103降解的数据。(a)cas9结构域架构的示意图,指示五个内切核酸酶结构域(ruvc-i-ruvc-iv,hnh)和arm(富含精氨酸的基序)。(b)354野生型(wt)、δcas9、cas9:d11a(ruvc-i)、cas9:r59a(arm)、cas9:e86a(ruvc-ii)、cas9:r102a(ruvc-ii)、cas9:d876a(ruvc-iii)、cas9:h969a(hnh区域)、cas9:h1162a(ruvc-iv)、以及cas9:d1165a(ruvc-iv)菌株中ftn_1103的相对表达(n=4,条线表示标准偏差)。(c)在利福平处理后,在wt(黑色圆形)、δcas9(蓝色正方形)、δscarna(黄色三角形)、以及δtracrrna(绿色菱形)菌株中ftn_1103降解的时间过程(n=3,点表示平均值并且条线表示标准偏差,对于所有突变体,与野生型相比,在30分钟时间点处,p≤0.05)。(d)表示在tracrrna(米黄色)与scarna(绿色)、以及在scarna与ftn_1103(紫色)之间的预测杂交的示意图。与相邻的黑色条线区分开的绿色碱基表示在特定tracrrna和scarna突变体中改变的碱基对,并且红色碱基指示ftn_1103的起始密码子和rbs。(e、f)在来自wt、编码cas9-flag、或cas9:r59a-flag的菌株的裂解物上执行抗flag免疫沉淀,并且在来自(e)scarna和(f)tracrrna的沉淀的rna上执行qrt-pcr。(g)wt、δscarna、scarna:rc4-8(表达碱基4-8的反向互补序列)、scarna:rc48-54(表达碱基48-54的反向互补序列)、δtracrrna、以及tracrrna:rcl3-17(表达碱基13-17的反向互补序列)菌株中ftn_1103的相对表达(n=4,条线表示标准偏差)。
图3示出表明cas9、tracrrna、以及scarna通过ftn_1103的时间性阻遏而有利于避开tlr2信号传导的数据。(a)来自未刺激(un)的、或用来自野生型(wt)、δcas9、δscarna、以及δtracrrna菌株的膜蛋白制剂、或用还缺乏ftn_1103的双缺失菌株(δcas9/1103、δscarna/1103、以及δtracrrna/1103)在20:1的相对moi下刺激5小时的野生型(wt)和tlr2-/-骨髓衍生巨噬细胞(bmdm)的il-6分泌(n=3)。(b)来自未感染的、或用野生型(wt)、δcas9、δscarna、以及δtracrrna菌株、或用双缺失菌株δcas9/1103、δscarna/1103、以及δtracrrna/1103在20:1的moi下感染5小时的wt或tlr2-/-bmdm的il-6分泌(n=6)。在通过wt(黑色圆形)、δcas9(蓝色正方形)、δscarna(黄色三角形)、以及δtracrrna(绿色菱形)菌株感染bmdm的过程中(c)ftn_1103、(d)cas9、(e)scarna、以及(f)tracrrna的相对表达水平(n=3,点表示平均值并且条线表示标准偏差,对于所有突变体,与野生型相比,p≤0.05)。
图4示出表明cas9、tracrrna、以及scarna对于毒力非常重要的数据。(a)在感染后48小时,来自鼠脾脏的野生型和指示突变体或双突变株的竞争指数。条线表示几何平均数。(b)用107cfu的野生型、δcas9、δscarna、或δtracrrna菌株感染小鼠,并且随时间推移监测存活率。(c)用104cfu的δcas9、δscarna、或δtracrrna菌株、或pbs对小鼠进行预防接种。28天之后,用107cfu野生型激发小鼠。
图5示出某些细菌cas9、tracrrna、以及scarna的实施例。
图6示出细菌cas9富含精氨酸的基序、ruvc-iii基序、以及ruvc-iv基序的实施例。
图7示意性地示出fncas9与rna靶标的相互作用。a:fncas9与由两个小rna、tracrrna以及scarna形成的dsrna复合物缔合。共同地,这允许tracrrna靶向mrna转录物。随后,mrna靶标的稳定性降低并且转录物损失。这通过当前未鉴定的fncas9活性或通过内源性rna酶的作用而发生。b:已被重新编程以靶向新mrna的假设tracrrna:scarna杂交体的示意图。
图8示出表明在人细胞中表达和产生新杀手弗朗西斯菌cas9的数据。用含有ha表位标记的新杀手弗朗西斯菌cas9(fncas9)的开放阅读框、由cmv启动子驱动的pcdna3.3真核表达载体转染人肝细胞癌细胞(huh7.5细胞)。a)提取总rna,并且针对fncas9转录物执行qrt-pcr且标准化为gapdh。b)提取总蛋白,通过sds-page分离,并且使用抗ha通过蛋白质印记进行分析,以检测fncas9和作为上样对照的抗gapdh。
图9示出表明fncas9可涉及以序列特异性方式限制病毒感染的数据。a、b)靶向rgrna与hcv基因组的指示部分5'utr(a)或3'utr(b)的部分相互作用的示意图。灰色突出显示区是决定靶向特异性的可变区。双链区确定fncas9相互作用。c)用含有cas9、hcv5'和3'靶向rgrna、非特异性对照靶向rgrna、或两者的组合的指示质粒构建体转染huh7.5细胞。在转染之后,用hcv(菌株cp7)感染细胞并且在感染后48小时,用抗e2抗体将细胞染色,以测量病毒蛋白。d)e2染色的定量,报道为与非转染细胞相比的抑制百分比。在感染后48小时,将感染细胞裂解并且测量荧光素酶活性。报道了与非转染细胞相比荧光素酶活性的相对抑制。
图10示出fncas9靶向hcv病毒rna的数据。仅用ha表位标记的fncas9,或与hcv5'utr靶向rgrna或非特异性对照rna结合转染huh7.5细胞。然后用如上hcv感染经转染的细胞。在感染后48小时,将细胞裂解并且针对ha,使裂解物经受免疫沉淀(ip)。在ip之后,从沉淀中提取rna,通过taqmanqrt-pcr针对(a)总hcv基因组进行分析并且通过gapdh水平归一化。当通过hcv特异性rgrna,而不是用非特异性对照指导fncas9时,在沉淀中观察到hcv基因组的显著富集。(b、c)通过sybergreenqrt-pcr,针对靶向rgrna的存在对沉淀rna进行分析,归一化为gapdh。
图11示出表明靶向fncas9可拯救hcv病毒感染的数据。a)实验概要的示意图。首先用海肾荧光素酶转染huh7.5细胞,从而产生hcv(cp7:rluc)rna并且允许病毒感染进行72小时。然后用所指示的fncas9和如上rgrna质粒构建体转染经感染的细胞。(b)在感染后48小时,将感染细胞裂解并且测量荧光素酶活性。报道了与非转染细胞相比荧光素酶活性的相对抑制。
详细说明
在更详细地描述本披露之前,应理解的是本披露不限于所描述的具体实施例,因此这些当然可以改变。还应当理解,在此使用的术语仅是为了描述特定实施例的目的,而并不意图是限制性的,因为本披露的范围仅由所附权利要求限定。
除非另外定义,在此所用的全部技术术语和科学术语具有与本披露所属领域的普通技术人员通常所理解的相同意义。虽然与在此所述的那些方法和材料相似或等同的任意方法和材料也可以用于实施或测试本披露中,然而现在描述优选的方法和材料。
在本说明书中引用的所有公开物和专利通过引用结合于此,就好像每个单独的公开物或专利被确切地并单独地指示为通过引用结合,并且通过引用结合于此从而结合引用的公开物披露和描述这些方法和/或材料。任何公开物的引用内容是针对在提交日之前的披露,并且不能理解为承认因为先前披露而本披露不能获得比这些公开物更早的申请日。此外,所提供的公开日期可能与实际的公开日期不同,实际的公开日期可能需要单独地确认。
如将对于本领域技术人员清楚的是,在阅读本披露时,在此描述和展示的单独实施例的每一个具有离散的组成部分和特征,这些组成部分和特征可以在不偏离本披露的范围或精神的情况下易于与任何其他一些实施例的特征分离或组合。可以按照所叙述的事件的顺序或按照逻辑上可行的任何其他顺序来进行任何叙述的方法。
除非另外说明,本披露的实施例将采用医学、有机化学、生物化学、分子生物学、药理学等的技术,这些技术是在本领域的技术之内。此类技术在文献中得到充分解释。
必须指出,如在说明书和所附权利要求书中所使用,单数形式“一个/一种(a/an)”和“该(the)”包括复数指示物,除非上下文另外清楚地规定。因此,例如,提及“一种支撑件”包括一种或多种支撑件。在本说明书和以下权利要求书中,将参考应定义为具有下列含义的大量术语,除非明显是相反的意图。
在描述各种实施例之前,提供以下定义并且应使用这些定义,除非另外指明。
如在此所用的术语“受试者”是指任何动物、优选病人、牲畜或家养宠物。
如在此所用的术语“核酸”是指从5'至3'末端阅读的脱氧核糖核苷酸或核糖核苷酸碱基的单链或双链聚合物。“核酸”还可任选地含有允许通过聚合酶正确读通且不减少由该核酸编码的多肽的表达的非天然存在或经改变的核苷酸碱基。术语“核苷酸序列”或“核酸序列”是指呈单独的单链或双链体形式的核酸的有义链和反义链。术语“核糖核酸”(rna)包括rnai(抑制性rna)、dsrna(双链rna)、sirna(小干扰rna)、mrna(信使rna)、mirna(微rna)、trna(转运rna,无论是装载有还是卸载了相应的酰基化氨基酸)、以及crna(互补rna),并且术语“脱氧核糖核酸”(dna)包括cdna和基因组dna以及dna-rna杂交体。词语“核酸区段”、“核苷酸序列区段”或更一般地讲“区段”将由本领域技术人员理解为功能性术语,包括基因组序列、核糖体rna序列、转运rna序列、信使rna序列、小调控rna、操纵子序列以及表达或可被适配成表达蛋白质、多肽或肽的较小工程化的核苷酸序列。
本披露的核酸还可通过本领域已知的方法完整或部分地合成,尤其是在希望提供植物优选序列的情况下。因此,可使用选定宿主优选的密码子合成具有所呈现的密码子的核酸的全部或部分。物种优选的密码子可例如由在具体宿主物种中表达的蛋白质中最频繁使用的密码子确定。核苷酸序列的其他修饰可产生具有略微改变的活性的突变体。
术语指定的多肽“编码核酸序列”是指包括基因的编码区的核酸序列或换言之编码基因产物的核酸序列。编码区可以按cdna、基因组dna或rna形式存在。当以dna形式存在时,寡核苷酸、多核苷酸或核酸可以是单链的(即正义链)或双链的。如果需要允许适当地启动原始rna转录物的转录和/或正确的加工,则可以紧邻基因的编码区放置适合的控制元件,如增强子/启动子、剪接点、多腺苷酸化信号等。可替代地,在本披露的表达载体中使用的编码区可以包含内源性增强子/启动子、剪接点、间插序列、多腺苷酸化信号等或内源性和外源性的两种控制元件的组合。
术语“cdna”是指互补dna(cdna),即通常通过酶(逆转录酶和dna聚合酶)催化的由rna(例如mrna)模板合成的dna。
术语“基因”是指包含了产生rna、或多肽或其前体(例如,胰岛素原)所必需的编码序列的核酸(例如,dna或rna)序列。功能性多肽可以由一个全长编码序列或由该编码序列的任何部分来编码,只要保持了该多肽的所希望的活性或功能特性(例如,酶促活性、配体结合、信号转导等)即可。当关于基因使用时,术语“部分”是指该基因的片段。片段的大小可在几个核苷酸至整个基因序列减去一个核苷酸的范围内。因此,“包含一个基因的至少一部分的核苷酸”可包含该基因的片段或该整个基因。术语“基因”还涵盖一个结构基因的编码区并且包括邻近于该编码区在5'和3'末端上距任一端约1kb距离定位以使得该基因对应于全长mrna的长度的序列。定位在编码区5'并且存在于mrna上的序列称为5'非翻译序列。定位在编码区3'或下游并且存在于mrna上的序列称为3'非翻译序列。术语“基因”包括基因的cdna和基因组形式两者。一种基因的基因组形式或克隆包含了间杂有称为“内含子”或“插入区”或“插入序列”的非编码序列的编码区。内含子为转录成核rna(mrna)的基因的区段;内含子可以含有调控元件,如增强子。内含子被从核或初级转录物中移除或“剪切掉”;因此信使rna(mrna)转录物中不存在内含子。mrna在翻译期间起到指明新生多肽中氨基酸的序列或次序的作用。
除了含有内含子之外,基因的基因组形式还可以包括rna转录物上存在的定位在该序列的5'或3'末端的序列。这些序列称为“侧翼”序列或区域(这些侧翼序列定位在mrna转录物上存在的非翻译序列的5'或3')。5'侧翼区可以含有调控序列,如控制或影响该基因的转录的启动子和增强子。3'侧翼区可含有指导转录的终止、转录后裂解和聚腺苷酸化的序列。
术语“异源基因”是指编码不处于其天然环境(即,通过人手改变过的)的因子的基因。例如,异源基因包括从一个物种引入到另一个物种中的基因。异源基因还包括以一些方式改变(例如,突变、加入多个拷贝、连接至非天然启动子或增强子序列等)的生物体天然具有的基因。异源基因可包括包含细菌基因的cdna形式的细菌基因序列;cdna序列可以有义(以产生mrna)或反义取向(以产生与mrna转录物互补的反义rna转录物)表达。
术语“互补”和“互补性”是指通过碱基配对规则相关联的多核苷酸(即,一个核苷酸序列)。例如,对于序列“a-g-t”,与序列“t-c-a”互补。互补性可以是“部分的”,其中仅一些核酸的碱基根据碱基配对规则是匹配的。或者,在多个核酸之间可以存在“完全”或“全部”的互补性。核酸链之间的互补性程度对于核酸链之间杂交的效率和强度具有显著影响。这在扩增反应以及取决于核酸之间的结合的检测方法中特别重要。
在此披露的核酸分子或指导或靶向rna能够与靶核酸在某些情况下特异性杂交。如在此所使用,如果两个核酸分子能够形成氢键合核酸结构,那么这两个分子被称为能够与彼此特异性杂交。核酸分子可以表现出完全互补性。如果两个分子能够以足够的稳定性彼此杂交,以允许它们在至少常规的“低严格”条件下保持相互退火,那么称这两个分子是“最低度互补的”。类似地,如果两个分子能够以足够的稳定性彼此杂交,以允许它们在常规的“高严格”条件下保持相互退火,那么称这两个分子是互补的。常规严格条件由萨姆布鲁克(sambrook)等人(1989)和海姆斯(haymes)等人(1985)描述。
不是完全互补性因此是可能的,只要这没有完全排除rna分子与靶标形成氢键合结构的能力即可。因此,为了使rna充当靶标的指导序列,rna仅需要在能够在表达该rna的细胞的生理条件下形成稳定的氢键合结构的序列中具有足够的互补性即可。
当就核酸分子而言时,术语“重组”是指包含通过分子生物学技术而连接在一起的核酸区段的核酸分子。当就蛋白质或多肽而言时,术语“重组”是指使用重组核酸分子表达的蛋白质分子。
“克隆载体”或“载体”是指用作媒介物以将外源遗传物质运载到它可在其中复制和/或表达的另一个细胞中的核酸分子。含有外源核酸的克隆载体称为重组载体。载体的例子是质粒、病毒载体、粘粒、以及人工染色体。重组载体通常含有复制起点、多克隆位点、以及选择性标志物。核酸序列通常由插入序列(重组核酸或转基因)和充当载体“骨架”的较大序列组成。将遗传学信息转移至另一个细胞的载体的目的通常是分离、倍增、或表达靶细胞中的插入序列。表达载体(表达构建体)是用于表达靶细胞中的转基因,并且通常具有驱动转基因的表达的启动子序列。将载体插入靶细胞中是指细菌和真核细胞的转化或转染,但是病毒载体的插入通常称为转导。
术语“可操作组合”、“以可操作顺序”和“可操作地连接”是指核酸序列的键,其方式使得产生能够指导给定基因的转录和/或所期望蛋白分子的合成的核酸分子。该术语还指氨基酸序列的键,其方式使得产生功能蛋白。
术语“调控元件”是指控制核酸序列的表达的一些方面的遗传元件。例如,启动子是有利于可操作地连接的编码区的转录的起始的调控元件。其他调控元件是剪接信号、聚腺苷酸化信号、终止信号等。
真核生物中的转录控制信号包括“启动子”和“增强子”元件。启动子和增强子由与涉及转录的细胞蛋白特异性地相互作用的dna序列的短阵列组成(马尼亚蒂斯(maniatis)等人,科学236:1237,1987)。已从多种真核来源包括酵母、昆虫、哺乳动物或植物细胞中的基因中分离了启动子和增强子元件。已从病毒中分离了启动子和增强子,并且类似控制元件,如启动子,也存在于原核生物中。具体启动子和增强子的选择取决于表达目标蛋白所用的细胞类型。一些真核启动子和增强子具有广泛的宿主范围,而其他真核启动子和增强子在有限的细胞类型子集中是有功能的(综述参见沃斯(voss)等人,生化科技的趋势(trendsbiochem.sci.),11:287,1986;以及马尼亚蒂斯等人,同上,1987)。
如在此所使用的术语“启动子元件”、“启动子”、或“启动子序列”是指定位在dna聚合物的蛋白质编码区的5'末端(之前)的dna序列。自然界中已知的大多数启动子的位置在转录区之前。启动子用作开关,激活基因的表达。如果基因被激活,那么称为它被转录、或参与转录。转录涉及来自该基因的mrna的合成。因此,启动子充当转录调控元件并且还提供用于起始基因至mrna的转录的位点。如用于启动子的术语“细胞类型特异性”是指在相同组织中的不同细胞类型中相对不存在相同的目标核苷酸序列的表达的情况下,能够指导目标核苷酸序列在特定细胞类型中的选择性表达的启动子。启动子可为组成型或可调控的。当就启动子而言时,术语“组成型”是指启动子能够在不存在刺激(例如,热休克、化学品、光照等)的情况下指导可操作地连接的核酸序列的转录。典型地,组成型启动子能够在基本上任何细胞和任何组织中指导转基因的表达。相比之下,“可调节的”或“诱导型”启动子是能够在刺激(例如,热休克、化学品、光照等)的存在下指导可操作地连接的核酸序列的一定水平的转录的启动子,该水平不同于在不存在刺激的情况下该可操作地连接的核酸序列的转录的水平。
增强子和/或启动子可为“内源性的”或“外源性的”或“异源的”。“内源性”增强子或启动子是在基因组中与给定基因天然连接的增强子或启动子。“外源性”或“异源”增强子或启动子是通过遗传操纵的方法(即,分子生物学技术)与一个基因毗邻放置,使得该基因的转录由所连接的增强子或启动子指导的增强子或启动子。例如,与第一基因可操作组合的内源性启动子可被分离、移除、并且被放置与一个第二基因可操作组合,从而将其制备成与该第二基因可操作组合的“异源启动子”。
重组dna序列在真核细胞中的有效表达被认为包括指导所得转录物的有效终止和聚腺苷酸化的信号的表达。转录终止信号通常存在于聚腺苷酸化信号下游并且长度为几百个核苷酸。如在此所使用的术语“poly(a)位点”或“poly(a)序列”表示指导新生rna转录物的终止和聚腺苷酸化的dna序列。重组转录物的有效聚腺苷酸化是希望的,因为缺乏poly(a)尾巴的转录物是不稳定的并且被快速降解。表达载体中利用的poly(a)信号可以是“异源的”或“内源性的”。内源性poly(a)信号是在基因组中天然存在于给定基因的编码区的3'末端的poly(a)信号。异源poly(a)信号是从一个基因中分离出来且放置到另一个基因3'的poly(a)信号。
术语“标志物”是指编码具有活性的酶的基因,该活性给在其中表达选择性标志物的细胞赋予抗生素或药物抗性,或赋予可被检测的性状(例如,发光或荧光)的表达。选择性标志物可以是“阳性的”或“阴性的”。阳性选择性标志物的例子包括赋予对g418和卡那霉素的抗性的新霉素磷酸转移酶(nptii)基因,以及赋予对抗生素潮霉素的抗性的细菌潮霉素磷酸转移酶基因(hyg)。阴性选择性标志物编码酶活性,当在适当的选择性培养基中生长时,该酶活性的表达对细胞是细胞毒性的。例如,hsv-tk基因通常用作阴性选择性标志物。hsv-tk基因在于更昔洛韦或阿昔洛韦存在下生长的细胞中的表达是细胞毒性的;因此,细胞在含有更昔洛韦或阿昔洛韦的选择性培养基中的生长选择出能够表达功能性hsvtk酶的细胞。
术语“报道基因”是指编码可被测定的蛋白质的基因。报道基因的例子包括但不限于荧光素酶(参见,例如,德韦(dewet)等人,分子细胞生物学(mol.cell.biol.)7:725(1987)和美国专利号6,074,859;5,976,796;5,674,713;以及5,618,682;所有专利以引用方式结合在此)、绿色荧光蛋白(例如,genbank登录号u43284;多个gfp变体可从加利福尼亚州帕罗奥图(paloalto,calif)的clontechlaboratories公司商购获得)、氯霉素乙酰转移酶、β-半乳糖苷酶、碱性磷酸酶、以及辣根过氧化物酶。
“序列一致性”是指两个或更多个核酸或蛋白质之间相关性的量度,并且通常给出为相对于总比较长度的百分比。一致性计算考量一致的或在它们各自的较大序列中处于相同的相对位置的那些核苷酸或氨基酸残基。一致性的计算可通过计算机程序内包含的算法、使用默认参数来进行,这些程序如“gap”(遗传学计算机组(geneticscomputergroup),威斯康星州麦迪逊(madison,wis.))和“align”(dnastar,威斯康星州麦迪逊)。在某些实施例中,序列“一致性”是指在序列比对中在比对的两个序列之间精确匹配残基的数目(表示为百分比)。在某些实施例中,比对的一致性百分比可以使用一致位置的数目除以最短序列中的较大序列或排除突出端的等效位置的数目来计算,其中内部缺口计为等效位置。例如,多肽gggggg和ggggt具有五分之四或80%的序列一致性。例如,多肽gggppp和gggappp具有七分之六或85%的序列一致性。
在某些实施例中,对于任何设想的序列一致性百分比,还设想的是,序列可以具有相同的序列相似性百分比。“相似性”百分比用于定量相似性的程度,例如,比对的两个序列之间的氨基酸的疏水性、氢键合可能性、静电荷。该方法类似于测定一致性,不同的是,某些氨基酸不必须是一致的,以具有匹配。在某些实施例中,序列相似性可使用默认参数利用众所周知的计算机程序进行计算。典型地,如果氨基酸在具有相似特性的组之中,那么可根据匹配对它们分类,例如根据以下氨基酸组:芳族-fyw;疏水性-avil;带正电荷的:rkh;带负电荷的-de;极性的-stnq。
部分互补序列是至少部分抑制(或与其竞争)完全互补序列与靶核酸杂交的序列-也称为“基本上同源的”。完全互补序列与靶序列杂交的抑制可在具有低严格性的条件下使用杂交测定(southern或northern印迹、溶液杂交等)来检验。基本上同源的序列或探针在具有低严格性的条件下将竞争且抑制与靶标完全同源的序列的结合(即,杂交)。这并不是说具有低严格性的条件使得非特异性结合被允许;低严格条件要求两个序列彼此的结合是一种特异性(即,选择性)相互作用。可通过使用第二靶标测试非特异性结合的不存在,该第二靶标甚至缺乏部分程度的互补性(例如,小于约30%一致性);在不存在非特异性结合的情况下,探针将不与该第二非互补靶标杂交。
下列术语用来描述两种或更多种多核苷酸之间的序列关系:“参考序列”、“序列一致性”、“序列一致性百分比”、以及“基本一致性”。“参考序列(referencesequence)”是用作序列比较基础的限定序列;参考序列可以是较大序列的子集,例如,作为序列表中给定的全长cdna序列的区段或可包含完整的基因序列。一般地,参考序列的长度为至少20个核苷酸,常为至少25个核苷酸长,且通常为至少50个核苷酸长。因为两条多核苷酸各自可以(1)包含在两个多核苷酸之间类似的序列(即,完整的多核苷酸序列的一部分),以及(2)可进一步包含与两条多核苷酸相去甚远的序列,典型地通过在“比较窗”上比较两个多核苷酸的序列进行两个(或更多个)多核苷酸间的序列比较以鉴定和比较具有序列相似性的局部区域。如在此所使用,“比较窗”是指概念上至少20个连续核苷酸位置的区段,其中多核苷酸序列可以与至少20个连续核苷酸的参考序列进行比较,并且其中在该比较窗中的该多核苷酸序列部分与参考序列(其不包含添加或缺失)相比可以包含20%或更少的添加或缺失(即,缺口),从而对这两条序列进行最佳比对。用于比对比较窗的序列的最佳比对可通过史密斯(smith)和沃特曼(waterman)的局部同源性算法(史密斯和沃特曼,应用数学进展(adv.appl.math.)2:482(1981))、通过尼德曼(needleman)和翁施(wunsch)的同源性比对算法(尼德曼和翁施,分子生物学杂志(j.mol.biol.)48:443(1970))、通过皮尔森(pearson)和利普曼(lipman)的相似性搜索方法(皮尔森和利普曼,美国国家科学院院刊(proc.natl.acad.sci.(u.s.))85:2444(1988))、通过这些算法的计算机实现方式(威斯康星遗传学软件包发行版7.0(wisconsingeneticssoftwarepackagerelease7.0)中的gap、bestfit、fasta、以及tfasta,遗传学计算机组,575sciencedr.,威斯康星州麦迪逊)、或通过检查来进行,并且选择通过各种方法产生的最佳比对(即,在比较窗上得到最高同源性百分比)。在某些实施例中,术语“序列一致性”是指在比较窗上两个多核苷酸序列是一致的(即,在核苷酸紧接核苷酸的基础上)。在一些实施例中,术语在比较窗上的“序列一致性百分比”是通过比较在该比较窗上的两条最佳比对序列,确定在两条序列上出现一致的核酸碱基(例如,a、t/u、c、g、或i)位置的数目,从而获得匹配位置的数目,用匹配位置的数目除以在该比较窗(即,窗口大小)中的总位置数目,并将该结果乘以100从而得到序列一致性百分比来计算的。
当参考多肽使用时,术语“变体”是指在一个或多个氨基酸上彼此不同的氨基酸序列,通常为相关多肽。该变体可以具有“保守”变化,其中取代的氨基酸具有相似的结构或化学特性。一种类型的保守氨基酸取代是指具有相似侧链的残基的可互换性。例如,具有脂族侧链的一组氨基酸是甘氨酸、丙氨酸、缬氨酸、亮氨酸、以及异亮氨酸;具有脂族-羟基侧链的一组氨基酸是丝氨酸和苏氨酸;具有含酰胺侧链的一组氨基酸是天冬酰胺和谷氨酸;具有芳族侧链的一组氨基酸是苯丙氨酸、酪氨酸、以及色氨酸;具有碱性侧链的一组氨基酸是赖氨酸、精氨酸、以及组氨酸;并且具有含硫侧链的一组氨基酸是半胱氨酸和甲硫氨酸。优选的保守氨基酸取代组是:缬氨酸-亮氨酸-异亮氨酸、苯丙氨酸-酪氨酸、赖氨酸-精氨酸、丙氨酸-缬氨酸、以及天冬酰胺-谷氨酸。更为罕见的是,变体可具有“非保守”变化(例如,用色氨酸替代甘氨酸)。类似微小变异还可以包括氨基酸缺失或插入(换言之,添加)或两者。可以使用本领域中熟知的计算机程序(例如,dnastar软件)发现确定哪些或多少氨基酸残基可以被取代、插入或缺失而不消除生物活性的指导。可在功能测定中测试变体。某些变体具有小于10%、优选地小于5%、仍然更优选地小于2%的变化(取代、缺失等)。
crispr-cas系统介导细菌先天性免疫逃避和毒力
已发现,新杀手弗朗西斯菌的cas蛋白cas9利用独特的小crispr-cas-相关rna(scarna)来介导编码细菌脂蛋白(blp)的内源性转录物的阻遏。由于blp触发意在对抗病原体的促炎性先天性免疫应答,crispr-cas介导的blp的阻遏对于新杀手弗朗西斯菌是关键的,以减弱宿主炎症性应答和提高毒力。cas9蛋白高度富含病原性和共生性细菌。在此披露的研究表明crispr-cas介导的基因调控可以广泛地促进这种细菌与真核宿主的相互作用。
新杀手弗朗西斯菌是在它通过真核细胞的吞噬体时避开宿主防御以在胞液内复制至高的数目的细胞内病原体。已开发出防止通过多种模式识别受体(prr)识别的机制,这些模式识别受体检测细菌并且定位至宿主吞噬细胞的表面和吞噬体。一种prr,即toll样受体2(tlr2)识别blp并且对于防御新杀手弗朗西斯菌是重要的。通过减弱tlr2活化,新杀手弗朗西斯菌在胞液中到达其复制场所(replicativeniche),而无需诱导显著的炎症性信号传导,从而促进其发病。
新杀手弗朗西斯菌基因ftn_0757参与由基因ftn_1103编码的blp的阻遏,但是其作用机制尚不明确。参见琼斯(jones)等人,标题为“通过新杀手弗朗西斯菌阻遏细菌脂蛋白的产生有利于避开先天性免疫识别(repressionofbacteriallipoproteinproductionbyfrancisellanovicidafacilitatesevasionofinnateimmunerecognition)”,细胞微生物学(cellmicrobiol),2012。出人意料地,生物信息学分析揭示,ftn_0757与crispr-cas系统蛋白cas9具有显著的序列相似性,(参见图5)通常已知为介导外源dna的降解并且在内源性基因调控中起到的作用当前尚未知。
此外,ftn_0757存在于完整的ii型crispr-cas系统基因座中。ii型crispr-cas系统存在于病原体和共生生物如链球菌属物种、李斯特氏菌属物种、奈瑟氏菌属物种、以及乳杆菌属物种的基因组中。参见图6。该基因座含有cas1、cas2、以及cas4,均预测为参与通过获得新靶向crrna以及预测的反式激活rna(tracrrna)(crrna活性所需的一种附属小rna)来实现的适应性抗性。它还包含先前在crispr基因座中未描述的、不同于crrna和tracrrna的独特的小rna,在此称为小的crispr-cas相关rna(scarna)(图1的a图)。细菌cas9同系物包括在(切里尼斯基(chylinski),2013,rna生物学(rnabiol))中提及的那些和以下列出的那些:
脆弱拟杆菌(bacteroidesfragilis)脓肿分枝杆菌(mycobacteriumabscessus)
齿双歧杆菌(bifidobacteriumdentium)鸡败血支原体(mycoplasmagallisepticum)
空肠弯曲杆菌(campylobacterjejuni)运动支原体(mycoplasmamobile)
红嘴鸥弯曲杆菌(campylobacterlari)渗透支原体(mycoplasmapenetrans)
黄褐二氧化碳嗜纤维菌(capnocytophagaochracea)关节液支原体(mycoplasmasynoviae)
肉毒梭状芽孢杆菌(clostridiumbotulinum)气味类香菌(myroidesodoratus)
白喉棒状杆菌(corynebacteriumdiphtheria)灰色奈瑟氏菌(neisseriacinerea)
谷氨酸棒状杆菌(corynebacteriumkroppenstedtii)浅黄色奈瑟氏菌(neisseriaflavescens)
粪肠球菌(enterococcusfaecalis)乳糖奈瑟氏菌(neisserialactamica)
人费克兰姆菌(facklamiahominis)脑膜炎奈瑟氏菌(neisseriameningitidis)
大芬戈尔德菌(finegoldiamagna)鼻疽诺卡氏菌(nocardiafarcinica)
嗜冷黄杆菌(flavobacteriumpsychrophilum)齿龈欧氏菌(olsenellauli)
b型土拉弗朗西斯菌(francisellaholarctica)出血败血性巴士杆菌(pasteurellamultocida)
新杀手弗朗西斯菌(francisellanovicida)大西洋假交替单胞菌(pseudoalteromonasatlantica)
土拉弗朗西斯菌(francisellatularensis)红平红球菌(rhodococcuserythropolis)
溶血孪生球菌(gemellahaemolysans)韦格斯卡多维亚菌(scardoviawiggsiae)
副流感嗜血杆菌(haemophilusparainfluenzae)食醇鞘氨醇杆菌(sphingobacteriumspiritivorum)
pittmaniae嗜血杆菌(haemophiluspittmaniae)金黄色葡萄球菌(staphylococcusaureus)
肝螺杆菌(helicobacterhepaticus)念珠状链杆菌(streptobacillusmoniliformis)
干酪乳杆菌(lactobacilluscasei)无乳链球菌(streptococcusagalactiae)
发酵乳杆菌(lactobacillusfermentum)停乳链球菌类马亚种(streptococcusdysgalactiaeequisimilis)
鼠李糖乳杆菌(lactobacillusrhamnosus)马链球菌兽疫亚种(streptococcusequizooepidemicus)
嗜肺军团菌(legionellapneumophila)gallolyticus链球菌(streptococcusgallolyticus)
莱姆血清型稻田钩端螺旋体(leptospirainadaiserovarlyme)戈登氏链球菌(streptococcusgordonii)
无害李斯特氏菌(listeriainnocua)马其顿链球菌(streptococcusmacedonicus)
单核细胞增生利斯特菌(listeriamonocytogenes)缓症链球菌(streptococcusmitis)
变形链球菌(streptococcusmutans)氯酚节杆菌(arthrobacterchlorophenolicus)
口腔链球菌(streptococcusoralis)纤维素降解梭菌(clostridiumcellulolyticum)
副血链球菌(streptococcusparasanguinis)efficiens棒状杆菌(corynebacteriumefficiens)
巴氏链球菌(streptococcuspasteurianus)谷氨酸棒状杆菌(corynebacteriumglutamicum)
假豕链球菌(streptococcuspseudoporcinus)需盐脱硫弧菌(desulfovibriosalexigens)
化脓性链球菌(streptococcuspyogenes)絮凝剂产生菌属tpsy(diaphorobactertpsy)
牙垢密螺旋体(treponemadenticola)elusimicrobiumminutum
小韦荣球菌(veillonellaparvula)黄色韩国生工菌(kribbellaflavida)
有毒威克斯菌(weeksellavirosa)汉氏硝化细菌(nitrobacterhamburgensis)
琥珀酸放线菌(actinobacillussuccinogenes)食清洁剂细小棒菌(parvibaculumlavamentivorans)
减肥菌(akkermansiamuciniphila)persephonellamarina
固氮螺旋菌属b510(azospirillumb510)推定的γ-变形菌纲(putativegammaproteobacterium)
barnesiellaintestinihominishtcc5015
长双歧杆菌(bifidobacteriumlongum)jostii红球菌(rhodococcusjostii)
慢生根瘤菌(bradyrhizobium)混浊红球菌(rhodococcusopacus)
伯克氏菌(burkholderialesbacterium)深红红螺菌(rhodospirillumrubrum)
溶纤维丁酸弧菌(butyrivibriofibrisolvens)绿色玫瑰弯菌(roseiflexuscastenholzii)
恒雄芝氏沟鞭藻玫瑰杆菌(dinoroseobactershibae)玫瑰弯菌属rs-1(roseiflexusrs-1)
直肠真杆菌(eubacteriumrectale)蓝细菌pcc6803(synechocystispcc6803)
尤里真杆菌珍珠亚种(eubacteriumyuriisubsp.margaretiae)弯曲热单孢菌(thermomonosporacurvata)
产琥珀酸拟杆菌(fibrobactersuccinogenes)auensis甲苯单胞菌(tolumonasauensis)
重氮营养葡糖杆菌(gluconacetobacterdiazotrophicus)产琥珀酸沃廉菌(wolinellasuccinogenes)。
唾液乳杆菌(lactobacillussalivarius)
parasutterellaexcrementihominis
肠道罗斯氏菌(roseburiaintestinalis)
inulinivorans罗斯氏菌(roseburiainulinivorans)
slackiaheliotrinireducens
嗜热链球菌(streptococcusthermophilus)
parvirubra萨特菌(sutterellaparvirubra)
wadsworthensis萨特菌(sutterellawadsworthensis)
未经培养的白蚁菌群1细菌
verminephrobactereiseniae
解纤维热酸菌(acidothermuscellulolyticus)
脂环酸芽孢杆菌(alicyclobacillushesperidum)
在此的研究表明,cas9系统具有超过外源dna防御的现有形态的非规范功能,以与非crrna、tracrrna以及scarna一起作用,以通过内源性mrna的靶向调控基因表达,从而引起先天性免疫逃避和毒力。参见图7。这种令人惊讶的观察显示,crispr-cas组分已共同操作,以执行不同于抵御外源核酸的功能。cas组分在通过细胞内病原体感染宿主细胞的过程中被有效地诱导。当宿主吞噬体中存在新杀手弗朗西斯菌时,通过激活cas9、tracrrna、以及scarna来阻遏ftn_1103,该病原体暂时性阻遏其blp表达并从而避开tlr2信号传导。已知编码cas9的一组109种细菌中的85种为病原体或共生生物,表明crispr-cas组分介导的调控机制可在与真核细胞相互作用的其他生物体中起作用。
用于改变多种生物学系统中的基因表达的cas9和靶向核酸复合物
在某些实施例中,本披露涉及使用在此披露的cas9系统,例如,cas9、tracrrna、以及scarna在各种生物学系统的背景中靶向目标rna的组合物和方法。这允许cas9系统用作rna干扰形式。cas9能够在真核胞液中起作用。通过使用较长的靶向rna,可以提高特异性。在某些实施例中,本披露设想了大于10、20、30、40、50、60、70、80、90、或100个核苷酸的靶向rna的区段。在此披露的cas9系统由所靶向的rna产生较低水平的蛋白质。参照受权利要求书保护的实施例,通过任何具体机制降低蛋白质结果是不预期的。认为在一些情况下,rna可能被降解,但是还可能的是,cas9只是置于靶rna上,以便阻断核糖体的接近,从而阻断翻译或通过一些其他未认识到的机制实现。
认为cas9不依赖于任何规范的rnai宿主因子,如dicer或risc复合物的组分,从而允许在可具有rnai的内在抑制的系统中使用。因此,cas9作为可编程的rna指导的rna靶向系统的工程化在多种生物学系统中是有益的。虽然在一些情况下,通过与靶序列杂交的“指导rna”(grna或靶向rna,或rna靶向的指导rna或rgrna)将cas9引导至其结合位点,但是设想指导序列可以含有一定数量的错配或二级结构。在某些实施例中,rgrna是tracrrna和scarna或其变体序列的融合。为了防止非靶标相互作用,可使用某些策略,例如,形成抑制非靶标相互作用的rgrna二级结构或改变rgrna的长度。
哺乳动物中的cas9被靶向以识别病毒rna阻止了生产性病毒复制。可通过改变rna靶向指导rna的序列使cas9靶向任何rna,作为能够抵抗任何病毒的抗病毒策略。对于治疗或预防病毒感染,cas9系统优于常规rnai。病毒可突变,以避开常规rnai系统。宿主rnai机器依赖于19-21bp的序列与有待被降解的靶标相互作用。在一些情况下,靶标中的即使单个碱基对突变也可完全消除通过宿主rnai机器进行的降解。具有cas9rna靶向指导rna的rnai不依赖于宿主的内源性rna沉默机器,即,自给自足。
病毒可直接遏制rnai机器,但是不认为遏制cas9活性,因为cas9来源于细菌,即,由于病毒病原体未进化成具有cas9,所以病毒可能不可离开这种系统。在某些实施例中,设想可利用同时(多重)靶向病毒rna的不同区域例如hcvrna的多个rgrna,从而限制病毒突变将有利于离开该靶向系统的机会。
用于与本披露一起使用的用于转化宿主细胞的适合的方法被认为实际上包括任何方法,通过这些方法,可将核酸(例如dna)引入细胞中,如通过原生质体的转化(美国专利号5,508,184)、通过干燥/抑制介导的dna摄取、通过电穿孔、通过用碳化硅纤维搅拌(美国专利号5,302,523;以及5,464,765);通过农杆菌介导的转化(美国专利号5,563,055;5,591,616;5,693,512;5,824,877;5,981,840;6,384,301)以及通过dna包被颗粒的加速(美国专利号5,015,580;5,550,318;5,538,880;6,160,208;6,399,861;6,403,865)等。尽管应用了如那些的技术,但是实际上可稳定地转化任何种属的细胞。在多细胞物种的情况下,可将转基因细胞再生到转基因植物和生物体中。
可使用被遗传工程化以表达具有对于不同的病毒或有害生物有特异性的rna靶向(rgrna)或多个rna靶向rna的cas9的植物和动物,以产生抗有害生物的子代。在某些实施例中,本披露涉及产生耐受病毒感染的转基因昆虫载体。
在某些实施例中,本披露设想cas9和grna在真核细胞中的表达用于靶向病毒,例如,c型肝炎(hcv)rna并且防止病毒复制。进行cas9对真核细胞胞液的靶向,以便靶向hcvrna(hcv为rna病毒,并且不具有dna期)。哺乳动物细胞中的cas9工程化研究通常包括至蛋白质的nls(核定位信号)并且使其靶向核,以便靶向dna。在某些实施例中,本披露的重组产生的cas9不含有nls序列。cas9在真核细胞的胞液中具有活性。真核细胞胞液中的cas9可用于靶向rna或可用于防止其翻译成蛋白质。通过改变“指导”rna的序列,cas9核酸复合物可被配置成靶向任何rna。
cas9系统对mrna的靶向可使用也可忍受不完全杂交(错配、成环等)的具有互补性的更大区域(在50bp的范围内)。这可用于产生“可调”系统,在该系统中,可控制敲除多少给定rna。在某些实施例中,本披露设想了在25至50个核苷酸、或25至100或更多个核苷酸、或35至65个核苷酸或更多个核苷酸、或40至60个核苷酸或更多个核苷酸范围内的单链靶向核酸。
在某些实施例中,本披露设想了同时靶向多个基因或靶rna,例如,同时靶向多个宿主基因、同时靶向多个病毒基因、或同时靶向病毒基因和宿主基因。在某些实施例中,本披露设想了cas9系统可用于靶向宿主rna。在某些实施例中,靶向病毒rna和宿主rna编码因子的组合促进病毒感染。
在某些实施例中,本披露设想了可以使免疫应答倾斜(例如,向th1、th2或th17表型)。可用以rgrna诱导th2应答的病原体处理感染,这将使该应答倾斜回到th1并且导致病原体的清除。
表达cas9和靶向核酸复合物的转基因植物
在某些实施例中,本披露设想了被遗传工程化以表达在此披露的cas9-核酸复合物,例如以用于预防病毒或其他有害生物感染目的的植物。在某些实施例中,本披露涉及通过用被配置成表达在此披露的cas9-核酸复合物的异源核酸转化宿主植物细胞来对植物进行基因修饰,以赋予有害生物抗性。
在某些实施例中,本披露提供重组核酸构建体,以用于实现具体宿主靶标(例如植物和植物细胞)的稳定转化。经转化的宿主靶标可由重组核酸构建体表达有效水平的在此披露的cas9系统。根据本披露提供的是表达某些cas9或细菌cas9核苷酸序列的核酸和结合缀合至一个核酸序列的cas9的rna,该核酸序列与植物或植物有害生物或其组合中的靶向基因的rna分子杂交。
在某些实施例中,本披露提供能够在细胞中表达为rna以抑制植物、植物有害生物或其组合的细胞或组织中的靶基因表达的核酸序列。这些序列包括编码一个或多个不同的核苷酸序列的核酸分子,其中不同核苷酸序列中的每一个靶向植物有害生物rna分子。这些序列可通过间隔区序列相连。编码cas9且靶向rna的核酸分子可以可操作地放置,受在宿主的细胞或组织中起作用的启动子序列的控制。
在某些实施例中,靶向序列处于有害生物的基因组中或处于有害生物的基因组中的一个基因的rna中。在某些实施例中,靶向序列经过选择,基本上参与有害生物的生长和发育,例如,在有害生物的存活力、生长、发育、感染性中起重要作用的蛋白质的mrna。这些mrna靶标可以是管家基因、转录因子等中的一个。
在某些实施例中,本披露提供用于在cas9表达和靶向rna以及植物有害生物摄取时在植物的细胞中表达,实现细胞或组织中靶标的遏制的核酸序列。在植物中表达基因遏制分子的方法是已知的(例如,wo06073727a2;美国公开2006/0200878a1),并且可用于表达在此披露的核苷酸序列。
可将核酸序列克隆在两个组织特异性启动子之间,如两个根特异性启动子,它们在转基因植物细胞中是可操作的并且在其中表达以在转基因植物细胞中产生mrna。根特异性启动子的例子是本领域已知的(例如,线虫诱导的rb7启动子;美国专利号5,459,252)。
在不同的植物物种中起作用的启动子也是本领域中众所周知的。可用于在植物中表达多肽的启动子包括为诱导型的、病毒的、合成的、或组成型的那些,和/或为时序调控的、空间调控的、以及时空调控的启动子。优选的启动子包括增强的camv35s启动子和fmv35s启动子。表现出根特异性的camv35s启动子的片段也可以是优选的。出于本披露的目的,可为优选的是在植物的根组织内实现这些基因的最高水平的表达。已鉴定了多个根特异性启动子并且是本领域中已知的(例如,美国专利号5,110,732;5,837,848;5,459,252)。
本披露的重组载体或克隆载体还可包括可筛选标志物。可筛选标志物可用于监测表达。示例性可筛选标志物包括β-葡糖苷酸酶或编码各种显色底物已知的酶的uida基因(gus);r-基因座基因,其编码调控植物组织中花青苷色素(红色)的产生的产物;β-内酰胺酶基因,编码各种显色底物(例如,padac、显色先锋霉素)已知的酶的基因;荧光素酶基因;xyle基因,其编码可转化显色儿茶酚的儿茶酚加双氧酶;α-淀粉酶基因;酪氨酸酶基因,其编码能够将酪氨酸氧化成dopa和多巴醌(其进而凝结为黑色素)的酶;以及α-半乳糖苷酶,其催化显色α-半乳糖底物。
优选的植物克隆或转化载体包括来源于根癌农杆菌的ti质粒的那些(例如,美国专利号4,536,475、4,693,977、4,886,937、5,501,967以及ep0122791)。发根农杆菌质粒(或“ri”)也是可用的并且是本领域已知的。使用农杆菌转化法形成的转基因植物通常含有插入到一个染色体中的单个简单的重组dna序列并且称为转基因事件。这种转基因植物可称为对于所插入的外源性序列是杂合的。相对于转基因纯合的转基因植物可通过以下方式获得:使含有单个外源性基因序列的独立的分离转基因植物与自身(例如f0植物)进行有性杂交(自交),以产生f1种子。四分之一的所产生的f1种子相对于转基因将为纯合的。使f1种子发芽得到植物,可针对杂合性对这些植物进行测试,通常使用snp测定或允许在杂合体与纯合体之间做出区分的热扩增测定(即,接合性测定)。使一个杂合性植株与自身或另一个杂合性植株杂交通常仅得到杂合性子代。
一般来讲,可为优选的是,在植物基因组中的非特异性位置处引入功能性重组dna。在特殊情况下,可为有用的是,通过位点特异性整合插入重组核酸构建体。存在已知在植物中起作用的若干位点特异性重组系统,包括如美国专利号4,959,317中所披露的cre-lox和如美国专利号5,527,695中所披露的flp-frt。
在某些实施例中,能够表达在此披露的cas9系统的种子也具有转基因事件,这提供除草剂耐受性。除草剂耐受性基因的一个有益的例子提供对草甘膦,即n-(膦酰基甲基)甘氨酸(包括这种除草剂的异丙胺盐形式)的抗性。
除了用重组dna构建体直接转化植物之外,可以通过将具有重组dna构建体的第一植株与缺乏该构建体的第二植株杂交来制成转基因植株。例如,可将用于基因遏制的重组dna引入第一株系中,该第一株系可适于转化,以产生转基因植株,该转基因植株可与第二株系杂交,以使用于基因遏制的重组dna渐渗进入第二株系中。
在某些实施例中,本披露可用于任何植物的转化,包括但不限于:玉米(zeamays)、卡罗拉油菜(canola)(欧洲油菜、芜菁亚种(brassicarapassp.))、苜蓿(紫苜蓿(medicagosativa))、水稻(oryzasativa)、黑麦(secalecereale)、高粱(两色蜀黍(sorghumbicolor)、sorghumvulgare)、向日葵(helianthusannuus)、小麦(triticumaestivum)、大豆(glycinemax)、烟草(nicotianatabacum)、马铃薯(阳芋(solanumtuberosum))、花生(arachishypogaea)、棉花(陆地棉(gossypiumhirsutum))、甘薯(ipomoeabatatus)、木薯(manihotesculenta)、咖啡(咖啡属亚种)、椰子(cocosnucifera)、菠萝(ananascomosus)、柑桔树(柑橘属物种)、可可(theobromacacao)、茶(茶树(camelliasinensis))、香蕉(芭蕉属物种)、鳄梨(perseaamericana)、无花果(ficuscasica)、番石榴(psidiumguajava)、芒果(mangiferaindica)、橄榄(油橄榄(oleaeuropaea))、番木瓜(caricapapaya)、腰果树(anacardiumoccidental)、澳洲坚果(macadamiaintegrifolia)、扁桃(巴旦杏(prunusamygdalus))、甜菜(betavulgaris)、燕麦、大麦、蔬菜类、观赏物、以及松柏类植物。
在某些实施例中,设想了作物植物(例如,谷类以及干豆、玉米、小麦、马铃薯、树薯(tapioca)、水稻、高粱、粟、木薯、大麦、豌豆、以及其他根、块茎、或种子作物)。用于本披露的重要的种子作物是油籽油菜、甜菜、玉米、向日葵、大豆、以及高粱。在某些实施例中,设想了园艺植物,包括莴苣、菊苣、以及蔬菜芸苔属(包括卷心菜、西建兰以及花椰菜),以及香石竹、天竺葵、喇叭花、以及秋海棠。本披露可以应用于烟草、瓜类蔬菜、胡萝卜、草莓、向日葵、番茄、胡椒、菊、杨木、桉树、以及松木。在某些实施例中,设想了如谷物种子的植物,如玉米、小麦、大麦、水稻、高粱、裸麦。在某些实施例中,设想了如油籽植物的植物。油籽植物包括卡罗拉油菜、棉花、大豆、红花、向日葵、芸苔、玉米、苜蓿、棕榈、椰子等。在某些实施例中,设想了如豆科植物的植物。豆科植物包括菜豆和豌豆。豆类包括瓜尔豆、槐豆、胡芦巴、大豆、四季豆、豇豆、绿豆、利马豆、蚕豆、滨豆、鹰嘴豆、等等。
在某些实施例中,植物是单子叶植物和/或双子叶植物。有用的单子叶植物的非限制性例子是水稻、玉米、小麦、棕榈树、草坪草、大麦、以及燕麦。有用的双子叶植物的非限制性例子是大豆、棉花、苜蓿、卡罗拉油菜、亚麻、番茄、甜菜、向日葵、马铃薯、烟草、玉米、小麦、水稻、莴苣、芹菜、黄瓜、胡萝卜、花椰菜、葡萄、以及草坪草。在某些实施例中,设想了如有花植物、树、草、阴生植物、以及开花的和不开花的观赏植物的植物。
可用于本披露的植物有害生物(即,可变得为非致病的或减小病原性)包括真菌、线虫动物、细菌、以及寄生植物,如独脚金(striga)、菟丝子以及槲寄生。通过本披露有用地治疗的植物有害生物包括霜霉菌(downymildews)。
技术人员可易于鉴定有待靶向的有害生物基因。这种基因可以是在这种有害生物对宿主植物的有害作用中起着直接或间接作用的任何有害生物基因。仅通过举例的方式,这种基因可以是在有害生物生长、发育、复制和繁殖、以及侵入或者侵染中起作用的基因。
在某些实施例中,该有害生物是一种植物病毒。这种植物病毒的例子是大豆花叶病毒、豆荚色斑病毒、烟草环斑病毒、大麦黄矮病毒、小麦主轴条斑病毒、土传花叶病毒、玉米中的小麦条斑病毒、玉米矮缩花叶病毒、玉米褪绿矮缩病毒、黄瓜花叶病毒、烟草花叶病毒、苜蓿花叶病毒、马铃薯x病毒、马铃薯y病毒、马铃薯卷叶病毒以及番茄金色花叶病毒。在这些之中,针对玉米矮缩花叶病毒、大麦黄矮病毒、小麦条斑花叶病毒、土传花叶病毒、马铃薯卷叶病毒以及黄瓜花叶病毒的保护是尤其重要的。
在某些实施例中,有害生物是草莓灰霉病菌(botrytiscinerea),一种具有异常广泛的宿主范围的死体营养型病原性真菌。栽培番茄(主要为lycopersiconesculentum)也易受灰霉菌(botrytis)的感染,并且真菌一般影响番茄植株的茎、叶以及果实。
表达cas9和靶向核酸复合物的转基因动物
除了转基因植物之外,本披露的某些实施例设想了表达在此披露的cas9系统以防止病原性感染(例如病毒)的转基因动物。所设想的转基因动物的非限制性例子包括鱼、牲畜和宠物。在某些实施例中,本披露设想了用编码或表达在此披露的cas9系统的期望核酸转化在组织培养基中生长的胚胎干细胞(es细胞)。在某些实施例中,本披露设想了将在此披露的克隆载体注入到人或非人动物的分离的胚胎干细胞中。
可通过将胚胎干细胞与编码在此披露的cas9系统的载体在es细胞将核酸掺入到该es细胞的基因组中的条件下混合来在培养基中转化es细胞。可通过将转化细胞注入到生物催化剂的内细胞团(icm)中,接着例如通过使雌性与输精管切除的雄性交配制备假孕动物来分离和选择成功转化的细胞。交配的刺激引发进行子宫接纳通常所需的激素变化。可替代地,可利用激素的直接给药。将胚胎植入子宫中为发育具有表达在此披露的cas9系统的核酸的转基因动物提供条件。
作为产生转基因动物的替代方法,可通过将克隆载体注入到精子原核中来转化受精卵。在融合之后,合子将分裂形成两个胚胎细胞。可以如上所述将胚胎植入假孕代母中。
在某些实施例中,本披露设想了包含表达在此披露的cas9系统与另一种蛋白质的组合(例如,生长激素)的核酸的转基因动物。在此披露的克隆载体可被配置成替代靶基因。在某些实施例中,本披露涉及包含表达在此披露的cas9系统的核酸和在它们的奶中表达重组蛋白的核酸的转基因绵羊或山羊。
在某些实施例中,本披露设想了包含表达在此披露的cas9系统的核酸和在它们的蛋(例如,蛋白)中表达重组蛋白的核酸的转基因鸡。
基因疗法
在某些实施例中,本披露涉及治疗或预防疾病、病症、或感染的方法,这些方法包括向对其有需要的受试者给予有效量的重组载体,该重组载体编码在此披露的cas9和核酸复合物。
在某些实施例中,本披露涉及治疗或预防病毒感染或其他病原性感染的方法,这些方法包括给予有效量的被配置成表达靶向病毒或病原性核酸的cas9-核酸复合物的载体。
在某些实施例中,本披露设想了与其他治疗剂、抗病原剂、抗病毒剂、抗细菌剂或疫苗结合给药。在某些实施例中,抗病毒剂选自阿巴卡韦、阿昔洛韦、阿昔洛韦、阿德福韦、金刚胺、安普那韦、安普利近、阿比朵尔、阿扎那韦、立普妥(atripla)、波普瑞韦、西多福韦、双汰芝、complera、达芦那韦、地拉韦啶、去羟肌苷、二十二醇、度鲁特韦、依度尿苷(edoxudine)、依非韦伦、恩曲他滨、恩夫韦地、恩替卡韦、泛昔洛韦、福米韦生、福沙那韦、膦甲酸钠、膦乙酸钠、更昔洛韦、伊巴他滨、伊姆诺韦(imunovir)、碘苷、咪喹莫特、茚地那韦、肌苷、iii型干扰素、ii型干扰素、i型干扰素、拉米夫定、洛匹那韦、洛韦胺、马拉韦罗、吗啉胍、美替沙腙、奈非那韦(nelfmavir)、奈韦拉平、奈沙韦(nexavir)、奥司他韦、聚乙二醇干扰素α-2a、喷昔洛韦、帕拉米韦、普利康那利(pleconaril)、鬼臼毒素、雷特格韦、利巴韦林、金刚乙胺、利托那韦、嘧啶、沙奎那韦、司他夫定、stribild、替诺福韦、替诺福韦地索普、替诺福韦艾拉酚胺富马酸酯(taf)、替拉那韦、三氟尿苷、三协唯(trizivir)、曲金刚胺、特鲁瓦达(truvada)、伐昔洛韦、缬更昔洛韦、韦克利韦洛克(vicriviroc)、阿糖腺苷、韦拉米啶(viramidine)、扎西他滨、扎那米韦、或齐多夫定、以及其组合。
在某些实施例中,本披露设想了通过靶向rna和dna病毒,例如,靶向rna病毒的基因组和/或转录物或dna病毒的病毒转录物来治疗和/或预防病毒感染。在一些实施例中,该病毒为或受试者被诊断患有甲型流感病毒(包括亚型h1n1),乙型流感病毒,丙型流感病毒,轮状病毒a,轮状病毒b,轮状病毒c,轮状病毒d,轮状病毒e,sars冠状病毒,人类腺病毒类型(hadv-1至55),人乳头瘤病毒(hpv)类型16、18、31、33、35、39、45、51、52、56、58、59,细小病毒b19,触染性软疣病毒,jc病毒(jcv),bk病毒,默克尔(merkel)细胞多瘤病毒,甲型柯萨奇病毒,诺如病毒,风疹病毒,淋巴细胞性脉络丛脑膜炎病毒(lcmv),黄热病毒,麻疹病毒,腮腺炎病毒,呼吸道合胞病毒,牛瘟病毒,加利福尼亚脑炎病毒,汉坦病毒,狂犬病病毒,埃博拉病毒,马尔堡病毒,单纯性疱疹病毒-1(hsv-1),单纯性疱疹病毒-2(hsv-2),水痘带状疱疹病毒(vzv),eb病毒(epstein-barrvirus)(ebv),巨细胞病毒(cmv),疱疹淋巴细胞病毒,玫瑰疹病毒(roseolovirus),卡波氏肉瘤相关疱疹病毒,甲型肝炎病毒(hav),乙型肝炎病毒(hbv),丙型肝炎病毒(hcv),丁型肝炎病毒(hdv),戊型肝炎病毒(hev)、人类免疫缺陷病毒(hiv),i型人嗜t淋巴细胞病毒(htlv-1),弗罗德脾脏病灶形成病毒(sffv)或异嗜性mulv相关病毒(xmrv)。
在某些实施例中,本披露设想了靶向rna病毒的rna基因组中的多个位点,或dna病毒的rna转录物,以用于防止病毒产生抗性的目的。
在某些实施例中,本披露设想了可使用cas9和靶向不同病毒的grna的混合物作为一种“一次性(one-shot)”治疗。
在某些实施例中,本披露设想了使用在此披露的cas9系统来改善受试者处理并且对疫苗产生应答的能力,这通过将在此披露的克隆载体与疫苗结合给药来实现,其中cac9核酸复合物被配置成具有grna,以靶向il-10和/或其他抗炎性细胞因子的mrna表达、和/或靶向mrna表达pd-1/pd-l1。
在某些实施例中,本披露设想了使用cas9系统治疗癌症。例如,grna可被配置成靶向在癌细胞中过表达的mrna或微rna或控制癌基因的表达。一些癌症遏制rnai机器,但是在具有在此披露的cas9系统的情况下,将可能不能这样做。用在此披露的cas9系统靶向mrna通常导致基因产物的表达降低,而靶向微rna通常导致基因产物的表达增加。
在某些实施例中,本披露涉及治疗或预防癌症,包括给予表达在此披露的cas9和指导核酸复合物的载体,其中癌症选自脑癌、肺癌、子宫颈癌、卵巢癌、结肠癌、乳腺癌、胃癌、皮肤癌、卵巢癌、胰腺癌、前列腺癌、颈癌、以及肾癌。
在某些实施例中,本披露涉及治疗癌症的方法,包括给予有效量的在此披露的克隆载体,该克隆载体被配置成表达靶向与癌基因相关联的mrna或微rna的cas9和指导核酸复合物。在某些实施例中,靶mrna或微rna与以下相关联:k-ras、含有3个的杆状病毒iap重复序列、含有7个的杆状病毒iap重复序列、肿瘤蛋白p53、肿瘤蛋白p53调控的细胞凋亡诱导蛋白1、肿瘤蛋白p73、血管内皮生长因子a、v-akt鼠胸腺瘤病毒癌基因、磷酸酶和紧张素、b细胞cll/淋巴瘤2、信号转导和转录活化蛋白3、表皮生长因子受体、v-erb-b2禽成红细胞白血病病毒癌基因、肿瘤坏死因子、肿瘤坏死因子超家族成员14、b细胞1中的κ轻多肽基因增强子的核因子、连环蛋白(钙粘蛋白相关蛋白)β1、转化生长因子β1、细胞周期蛋白依赖性激酶抑制剂1a、半胱天冬酶3、半胱天冬酶8、半胱天冬酶9、端粒酶逆转录酶、低氧诱导因子1α亚基、atp结合盒亚家族b、细胞周期蛋白依赖性激酶抑制剂2a、v-myc禽成髓细胞瘤病毒癌基因、胰岛素样生长因子1、基质金属蛋白酶7、基质金属蛋白酶9、白细胞介素8、细胞周期蛋白b1、细胞周期蛋白d1、趋化因子(c-c基序)配体2、钙粘蛋白1、e-钙粘蛋白、促细胞分裂原活化蛋白激酶1、γ干扰素、肿瘤坏死因子(配体)超家族成员10、微管相关蛋白tau、细胞凋亡的x连锁抑制剂、fas细胞表面死亡受体、成视网膜细胞瘤1、bcl-2、bcl2样2、bcl2相关x蛋白、bcl2拮抗剂/杀手1、小窝蛋白1、小窝蛋白、雷帕霉素的机制靶标、v-kit哈迪-朱克曼(hardy-zuckerman)4猫科肉瘤病毒癌基因、促细胞分裂原活化的蛋白激酶14、腺瘤性结肠息肉蛋白、极光激酶b、细胞周期蛋白依赖性激酶1、细胞周期蛋白依赖性激酶4、细胞周期蛋白依赖性激酶抑制剂1b、血红素加氧酶(脱环)1、notch1、notch2、分泌型磷蛋白1、促细胞分裂原活化的蛋白激酶3、矮小相关转录因子1、叉头框o3、叉头框p3、jun原癌基因、多腺苷二磷酸核糖聚合酶1、哈维(harvey)大鼠肉瘤病毒癌基因、糖原合酶激酶3β、氧化氮合酶2、ras相关c3肉毒杆菌毒素底物1、e1a结合蛋白p300、fas配体、atp结合盒g2、creb结合蛋白、蛋白激酶cα、fms相关酪氨酸激酶3、成纤维细胞生长因子2、o-6-甲基鸟嘌呤-dna甲基转移酶、关卡激酶2、diabloiap-结合线粒体蛋白、帕金森蛋白2、polo样激酶1、转录因子7样2、e2f转录因子1、高迁移率族匣1、早幼粒细胞性白血病、bcl2样1、尿激酶纤溶酶原活化蛋白、肿瘤坏死因子受体超家族成员1a、增殖细胞核抗原、尿激酶受体纤溶酶原活化蛋白、apex核酸酶、凝集素半乳糖苷结合可溶物3、骨髓细胞白血病序列1、大麻素受体1、缝隙连接蛋白α1、由单克隆抗体ki-67识别的抗原、钙感应受体、血小板反应蛋白1、pou种类5同源异型框1、肝细胞核因子4α、转化生长因子β受体ii、血小板衍生的生长因子受体α多肽、runt相关转录因子2、血管内皮生长因子c、早期生长应答1、血管生成素2、bmi1多梳环指癌基因、帕金森蛋白7、v-myc禽成髓细胞瘤病毒癌基因成神经细胞瘤、v-akt鼠胸腺瘤病毒癌基因同系物2、h2a组蛋白家族成员x、结节性硬化2、输出蛋白1、肽酰脯氨酰顺/反异构酶nima-相互作用1、dickkopfwnt信号传导途径抑制剂1、beclin1、血小板衍生的生长因子β多肽、肌动蛋白结合蛋白、融合在肉瘤中的集落刺激因子2、ets变体6、gata结合蛋白1、ran成员ras致癌基因、kruppel样因子4、kruppel样因子5、淋巴样增强子-结合因子1、组蛋白脱乙酰酶6、驿蛋白1、叶酸盐水解酶1、rasp21蛋白活化蛋白1、富含丝氨酸/精氨酸的剪接因子1、磷脂酰肌醇蛋白聚糖3、细胞粘附分子1、无翼型mmtv整合位点家族成员1、血小板衍生的生长因子α多肽、连接桥粒斑珠蛋白、蛋白精氨酸甲基转移酶1、白细胞介素11、成视网膜细胞瘤样2、e2f转录因子3、肿瘤相关钙信号转导物2、xiap相关因子1、微管相关蛋白4、长寿蛋白6、韦母氏肿瘤1相关蛋白、或其组合。
在某些实施例中,本披露涉及治疗癌症的方法,这些方法包括向对其有需要的受试者给予有效量的在此披露的克隆载体,该克隆载体被配置成表达靶向与生长因子或促细胞分裂原(例如c-sis)相关的mrna或微rna的cas9和指导核酸复合物。在某些实施例中,该癌症选自或该受试者被诊断患有成胶质细胞瘤、纤维肉瘤、骨肉瘤、乳腺癌、或黑素瘤。
在某些实施例中,本披露涉及治疗癌症的方法,这些方法包括向对其有需要的受试者给予有效量的在此披露的克隆载体,该克隆载体被配置成表达靶向与受体酪氨酸激酶(例如,表皮生长因子受体(egfr)、血小板衍生的生长因子受体(pdgfr)、以及血管内皮生长因子受体(vegfr)、her2/neu)相关的mrna或微rna的cas9和指导核酸复合物。在某些实施例中,该癌症选自或该受试者被诊断患有乳腺癌、胃肠道基质肿瘤、非小细胞肺癌、或胰腺癌。
在某些实施例中,本披露涉及治疗癌症的方法,这些方法包括向对其有需要的受试者给予有效量的在此披露的克隆载体,该克隆载体被配置成表达靶向与胞质酪氨酸激酶(例如,酪氨酸激酶的src-家族、syk-zap-70家族、以及btk家族)相关的mrna或微rna的cas9和指导核酸复合物。在某些实施例中,该癌症选自或该受试者被诊断患有结肠直肠癌、乳腺癌、黑素瘤、卵巢癌、胃癌、头颈癌、胰腺癌、肺癌、脑癌、或血癌。
在某些实施例中,本披露涉及治疗癌症的方法,这些方法包括向对其有需要的受试者给予有效量的在此披露的克隆载体,该克隆载体被配置成表达靶向与胞质丝氨酸/苏氨酸激酶及其调控亚基(例如,raf激酶和细胞周期蛋白依赖性激酶)相关的mrna或微rna的cas9和指导核酸复合物。在某些实施例中,该癌症选自或该受试者被诊断患有恶性黑素瘤、乳头状甲状腺癌、结肠直肠癌、或卵巢癌。
在某些实施例中,本披露涉及治疗癌症的方法,这些方法包括向对其有需要的受试者给予有效量的在此披露的克隆载体,该克隆载体被配置成表达靶向与调控gtp酶(例如,ras蛋白)相关的mrna或微rna的cas9和指导核酸复合物。在某些实施例中,该癌症选自或该受试者被诊断患有胰腺和结肠的腺癌、甲状腺肿瘤、或骨髓性白血病
在某些实施例中,本披露涉及治疗癌症的方法,这些方法包括向对其有需要的受试者给予有效量的在此披露的克隆载体,该克隆载体被配置成表达靶向与转录因子(例如,myc)相关的mrna或微rna的cas9和指导核酸复合物。在某些实施例中,该癌症选自或该受试者被诊断患有恶性t细胞淋巴瘤和急性骨髓性白血病、乳腺癌、胰腺癌、成视网膜细胞瘤、以及小细胞肺癌
在某些实施例中,本披露设想了靶向癌症癌基因或癌细胞中希望敲除的任何基因中的多个位点,以用于防止癌细胞产生抗性的目的。
在某些实施例中,本披露涉及治疗癌症的方法,这些方法包括与化学疗法结合给予有效量的在此披露的克隆载体,该克隆载体被配置成表达cas9和指导核酸复合物。在某些实施例中,化学疗法包括施用在某些实施例中
本披露设想了使用在此披露的cas9系统来改善受试者处理并且对化学疗法产生应答的能力,这通过与化学疗法结合给予在此披露的克隆载体来实现,其中cac9核酸复合物被配置成具有grna,以靶向il-10和/或其他抗炎性细胞因子的mrna表达、和/或靶向mrna表达pd-1/pd-l1。
实例
细菌和巨噬细胞感染
使用引物通过等位基因替代构建新杀手弗朗西斯菌u112和突变株。突变株类似于野生型在液体培养基中生长。由野生型和tlr2-/-c57bl/6小鼠制备来源于鼠骨髓的巨噬细胞并培养。在20:1细菌/巨噬细胞的感染复数(moi)下用细菌感染巨噬细胞。通过elisa(bd生物科技公司(bdbiosciences))对培养基上清液中il-6的浓度进行定量。对于使用细菌膜蛋白级份的刺激,轻柔洗涤细胞并且在20:1的相对moi下添加含有膜蛋白级份的培养基并且对il-6进行定量。
细菌cas9(ftn_0757)处于crispr-cas基因座中
对ftn_0757是否需要规范的crispr-cas系统来阻遏ftn_1103(细菌脂蛋白,blp)的表达进行测试(图1的a图)。cas9而不是其他cas基因的缺失导致ftn_1103转录物的水平增加100倍(图1的b图)。由于cas9降解被crrna靶向的dna,对crrna阵列或tracrrna是否为ftn_1103的阻遏所需要进行测试。crrna阵列的缺失不改变ftn_1103转录水平(图1的c图);然而,tracrrna的缺失导致ftn_1103转录增加,类似于cas9突变体(图1的c图)。另外,scarna的缺失导致ftn_1103转录的增加,表明它对ftn_1103阻遏也是重要的。cas9、tracrrna、以及scarna突变体的互补使ftn_1103表达恢复到接近野生型水平,并且突变体中ftn_1103的转录水平与蛋白质产量的增加相关。此外,缺乏cas9、tracrrna、以及scarna的三突变体表达与单突变体类似水平的ftn_1103mrna,从而提供以下遗传学证据:这些组分可以在相同的调控途径中共同作用来阻遏ftn_1103的表达。
突变型cas9、tracrrna和scarna,以及cas9中的基序参与blpmrnaftn_1103的阻遏
cas9蛋白含有四个ruvc内切核酸酶结构域(ruvc-i至ruvc-iv),以及hnh内切核酸酶结构域(图6)。虽然已知ruvc-i和hnh是靶dna的降解所必需的,但是其他结构域的功能是未知的。为了确定这些结构域中的哪些是ftn_1103mrna的阻遏所必需的,在每个结构域中构建缺乏保守残基的点突变株(图2-1的a图)。令人惊讶的是,ruvc-i和hnh催化突变体保持阻遏ftn_1103的野生型能力,证明cas9介导的ftn_1103的阻遏不需要这些结构域并且将该过程与dna的靶向区分开来。同时ruvc-ii点突变体也具有野生型的ftn_1103转录水平(图2-1的b图)。此外,发现不同的rna酶蛋白在ftn_1103阻遏中没有作用,支持了cas9能够介导靶向mrna的降解的假设。因此,阻遏ftn_1103的能力需要两个cas9内切核酸酶结构域,它们不同于介导靶dna降解的那些。
由于被cas9靶向可导致dna的降解,所以接下来通过降解对cas9、tracrrna、以及scarna是否参与ftn_1103mrna的沉默进行分析。在用利福平处理以阻断转录且阻止mrna的产生之后,ftn_1103转录物在野生型细胞中被快速耗尽(图2-1的c图)。相比之下,ftn_1103转录物在缺乏cas9、tracrrna、或scarna的突变体中未被降解(图2-1的c图)。因此,这三种crispr-cas系统组分中的每一种通过促进ftn_1103mrna的降解来参与其阻遏。
cas9含有先前未表征的、保守的、富含精氨酸的基序(arm)(图6)。由于cas9和两种srna(tracrrna和scarna)参与ftn_1103mrna的阻遏和降解,所以该推定的rna结合区对于cas9的功能可能是重要的。实际上,arm中的点突变完全消除cas9阻遏ftn_1103表达的能力(图2-1的b图),暗示了该区域在cas9与rna相互作用中的能力。对tracrrna和scarna的序列进行分析,并且确定了tracrrna可与scarna中的简并重复区杂交(图2-2的d图),类似于在tracrrna与crrna的重复区之间的相互作用,这是靶向dna所必需的。分析还表明,tracrrna的不同区域可与ftn_1103转录物的涵盖起始密码子和核糖体结合位点(rbs)的区域杂交(图2-2的d图)。为了确定cas9和rna是否缔合在一起,我们将cas9从表达这种蛋白质的flag-标记型式的菌株中免疫沉淀。tracrrna、scarna、以及ftn_1103mrna显著富集,与cas9缔合(图2-2的e、f图)。然而,这种缔合在cas9arm突变体(r59a)中被消除,暗示cas9的这个rna结合结构域与这些rna相互作用。
为了进一步确定这些组分之间的预测的相互作用是否是复合物的形成所必需的,在被预测为与scarna相互作用的tracrrna区域(碱基13-17)以及被预测为与tracrrna(碱基4-8)或与ftn_1103mrna(碱基48-54)相互作用的scarna区域中产生反向互补突变。所有三种突变导致完全不能阻遏ftn_1103转录物(图2-2的g图)。此外,被指示为破坏在scarna与tracrrna之间的相互作用的突变显著减弱任一小rna与cas9缔合的能力。因此,cas9、tracrrna、以及scarna的序列特异性缔合对于靶向和阻遏转录是重要的。
cas9氨基酸序列
cas9核苷酸
scarna序列
tracrrna序列
crispr-cas组分参与tlr2的逃避
由于cas9、tracrrna、以及scarna调控blpftn_1103的表达,并且blp是宿主tlr2的配体,所以对这些crispr-cas组分是否参与tlr2的逃避进行研究。tracrrna和scarna突变体的膜蛋白级份刺激促炎性细胞因子il-6的tlr2依赖性分泌的增加,类似于来自cas9突变体的那些(图3的a图)。这种应答在缺乏ftn_1103的双突变体中得到拯救,表明ftn_1103在这些菌株中的过表达很大程度上决定了tlr2信号传导的增加(图3的a图)。与依赖于ftn_1103的野生型新杀手弗朗西斯菌相比,缺乏cas9、tracrrna、或scarna的突变体也在巨噬细胞感染期间引发增强的tlr2依赖性il-6分泌(图3的b图)。这与不改变tlr2信号传导的其他cas基因、crrnaarray的突变体、或仅缺乏ftn_1103的突变体是相反的。这些数据综合表明,crispr-cas组分介导的blp的遏制有利于tlr2的逃避。
当细菌处于吞噬体中时cas9、tracrrna、以及scarna表达的诱导
为了确定ftn_1103的阻遏是否是一个有效逃避过程,我们分析了胞内感染期间crispr-cas组分的时间性表达。我们发现,当细菌处于吞噬体中时,ftn_1103表达显著降低(图3的c图),与cas9、tracrrna、以及scarna的大致100倍诱导直接相关(图3的d至f图)。在不存在cas9、tracrrna、或scarna的情况下,ftn_1103的时间性表达被消除(图3的c图)。这些数据表明在胞内感染期间一起诱导了cas9、tracrrna、以及scarna,从而在细菌在吞噬体中接近tlr2时允许ftn_1103的暂时遏制,由此有利于避开这种先天性免疫途径。
具有突变型cas9的细菌作为疫苗
进行野生型新杀手弗朗西斯菌与cas9、tracrrna或scarna缺失突变体的比较性感染。用1×105cfu的在1:1比率下在无菌pbs中的新杀手弗朗西斯菌的野生型和指示突变株皮下感染雌性c57bl/6小鼠。在感染后48小时,收获脾脏并且对细菌计数。对于存活率实验,用1×105cfu皮下感染小鼠并且在它们看起来濒死时将它们处死。对于预防接种实验,用l×105cfu的无菌pbs中的新杀手弗朗西斯菌的指示突变株皮下感染小鼠,并且28天之后,用1×107cfu野生型新杀手弗朗西斯菌皮下激发。
与野生型相比,全部三种突变体均高度减毒(1,000至10,000倍)(图4的a图),表明全部三种组分对于新杀手弗朗西斯菌毒力都是重要的。通过从突变体中缺失ftn_1103,这种减毒得以显著拯救。值得注意的是,缺乏crrna阵列或其他cas基因的突变体不减毒,这与它们阻遏ftn_1103的能力相关。cas9、tracrrna、以及scarna突变体在单独接种时也高度减毒,因为它们即使在100×ld50剂量下也不能导致死亡,而用野生型感染的小鼠则快速死于疾病(图4的b图)。在这种初始感染中存活的小鼠可以被保护免于受到随后新杀手弗朗西斯菌的致死激发。虽然初试小鼠快速死于激发,但是用cas9、tracrrna或scarna突变体免疫的小鼠完全被保护(图4的c图)。这证明了缺乏这些crispr-cas组分的突变体可有效地预防接种小鼠。鉴于其他病原体的crispr系统也可以通过调控内源性mrna促进毒力,这些基因的突变体可代表多种毒性细菌中的有吸引力的疫苗菌株。
在人细胞中表达和产生新杀手弗朗西斯菌cas9
用含有ha表位标记的人密码子优化的新杀手弗朗西斯菌cas9(fncas9)的开放阅读框、由cmv启动子驱动的pcdna3.3真核表达载体转染人肝细胞癌细胞(huh7.5细胞)。a)提取总rna,并且针对fncas9转录物执行qrt-pcr且标准化为gapdh。b)提取总蛋白,通过sds-page分离,并且使用抗ha通过蛋白质印记进行分析,以检测fncas9和作为上样对照的抗gapdh。人密码子优化的新杀手弗朗西斯菌cas9(基因座ftn_0757)
fncas9可涉及以序列特异性方式限制病毒感染。
靶向rgrna与hcv基因组的指示部分5'utr(a)或3'utr(b)的部分的相互作用在图9中示出。靶向rgrna为5'-guaucaggcaguaccacaag
加下划线指示靶向区域(seqidno:10)(参见图9),它可修饰为19个碱基(或更多个),以与期望的rna靶标形成碱基配对。加双下划线(seqidno:11)指示新杀手弗朗西斯菌cas9结合区域。这形成双链结构(参见图9)
加单下划线区域是决定靶向特异性的可变区。双链区确定fncas9相互作用。用含有cas9、hcv5'和3'靶向rgrna、非特异性对照靶向rgrna、或两者的组合的指示质粒构建体转染huh7.5细胞。在转染之后,用hcv(菌株cp7)感染细胞并且在感染后48小时,用抗e2抗体将细胞染色,以测量病毒蛋白(图9的c图)。
用所指示的fncas9和如上rgrna质粒构建体转染huh7.5细胞。然后用海肾荧光素酶感染细胞,从而产生hcv(cp7:rluc)。在感染后48小时,将感染细胞裂解并且测量荧光素酶活性。
序列表
<110>埃默里大学(emoryuniversity)
<120>cas9-核酸复合物及其相关用途
<130>12224us
<150>61/753,046
<151>2013-01-16
<150>61/905,368
<151>2013-11-18
<150>pct/us2014/011716
<151>2014-01-15
<160>14
<170>patentin3.5版
<210>1
<211>1628
<212>prt
<213>人工序列
<220>
<223>合成构建体
<400>1
metasnphelysileleuproilealaileaspleuglyvallysasn
151015
thrglyvalpheseralaphetyrglnlysglythrserleugluarg
202530
leuaspasnlysasnglylysvaltyrgluleuserlysaspsertyr
354045
thrleuleumetasnasnargthralaargarghisglnargarggly
505560
ileasparglysglnleuvallysargleuphelysleuiletrpthr
65707580
gluglnleuasnleuglutrpasplysaspthrglnglnalaileser
859095
pheleupheasnargargglypheserpheilethraspglytyrser
100105110
proglutyrleuasnilevalprogluglnvallysalaileleumet
115120125
aspilepheaspasptyrasnglygluaspaspleuaspsertyrleu
130135140
lysleualathrgluglngluserlysilesergluiletyrasnlys
145150155160
leumetglnlysileleugluphelysleumetlysleucysthrasp
165170175
ilelysaspasplysvalserthrlysthrleulysgluilethrser
180185190
tyrgluphegluleuleualaasptyrleualaasntyrsergluser
195200205
leulysthrglnlysphesertyrthrasplysglnglyasnleulys
210215220
gluleusertyrtyrhishisasplystyrasnileglnglupheleu
225230235240
lysarghisalathrileasnaspargileleuaspthrleuleuthr
245250255
aspaspleuaspiletrpasnpheasnpheglulyspheasppheasp
260265270
lysasngluglulysleuglnasnglngluasplysasphisilegln
275280285
alahisleuhishisphevalphealavalasnlysilelysserglu
290295300
metalaserglyglyarghisargserglntyrpheglngluilethr
305310315320
asnvalleuaspgluasnasnhisglngluglytyrleulysasnphe
325330335
cysgluasnleuhisasnlyslystyrserasnleuservallysasn
340345350
leuvalasnleuileglyasnleuserasnleugluleulysproleu
355360365
arglystyrpheasnasplysilehisalalysalaasphistrpasp
370375380
gluglnlysphethrgluthrtyrcyshistrpileleuglyglutrp
385390395400
argvalglyvallysaspglnasplyslysaspglyalalystyrser
405410415
tyrlysaspleucysasngluleulysglnlysvalthrlysalagly
420425430
leuvalasppheleuleugluleuaspprocysargthrilepropro
435440445
tyrleuaspasnasnasnarglysproprolyscysglnserleuile
450455460
leuasnprolyspheleuaspasnglntyrproasntrpglnglntyr
465470475480
leuglngluleulyslysleuglnserileglnasntyrleuaspser
485490495
phegluthraspleulysvalleulysserserlysaspglnprotyr
500505510
phevalglutyrlysserserasnglnglnilealaserglyglnarg
515520525
asptyrlysaspleuaspalaargileleuglnpheilepheasparg
530535540
vallysalaseraspgluleuleuleuasngluiletyrpheglnala
545550555560
lyslysleulysglnlysalasersergluleuglulysleugluser
565570575
serlyslysleuaspgluvalilealaasnserglnleuserglnile
580585590
leulysserglnhisthrasnglyilephegluglnglythrpheleu
595600605
hisleuvalcyslystyrtyrlysglnargglnargalaargaspser
610615620
argleutyrilemetproglutyrargtyrasplyslysleuhislys
625630635640
tyrasnasnthrglyargpheaspaspaspasnglnleuleuthrtyr
645650655
cysasnhislysproargglnlysargtyrglnleuleuasnaspleu
660665670
alaglyvalleuglnvalserproasnpheleulysasplysilegly
675680685
seraspaspaspleupheileserlystrpleuvalgluhisilearg
690695700
glyphelyslysalacysgluaspserleulysileglnlysaspasn
705710715720
argglyleuleuasnhislysileasnilealaargasnthrlysgly
725730735
lyscysglulysgluilepheasnleuilecyslysilegluglyser
740745750
gluasplyslysglyasntyrlyshisglyleualatyrgluleugly
755760765
valleuleupheglygluproasnglualaserlysproglupheasp
770775780
arglysilelyslyspheasnseriletyrserphealaglnilegln
785790795800
glnilealaphealagluarglysglyasnalaasnthrcysalaval
805810815
cysseralaaspasnalahisargmetglnglnilelysilethrglu
820825830
provalgluaspasnlysasplysileileleuseralalysalagln
835840845
argleuproalaileprothrargilevalaspglyalavallyslys
850855860
metalathrileleualalysasnilevalaspaspasntrpglnasn
865870875880
ilelysglnvalleuseralalyshisglnleuhisileproileile
885890895
thrgluserasnalaphegluphegluproalaleualaaspvallys
900905910
glylysserleulysaspargarglyslysalaleugluargileser
915920925
progluasnilephelysasplysasnasnargilelysglupheala
930935940
lysglyileseralatyrserglyalaasnleuthraspglyaspphe
945950955960
aspglyalalysglugluleuasphisileileproargserhislys
965970975
lystyrglythrleuasnaspglualaasnleuilecysvalthrarg
980985990
glyaspasnlysasnlysglyasnargilephecysleuargaspleu
99510001005
alaaspasntyrlysleulysglnphegluthrthraspaspleu
101010151020
gluileglulyslysilealaaspthriletrpaspalaasnlys
102510301035
lysaspphelyspheglyasntyrargserpheileasnleuthr
104010451050
proglngluglnlysalaphearghisalaleupheleualaasp
105510601065
gluasnproilelysglnalavalileargalaileasnasnarg
107010751080
asnargthrphevalasnglythrglnargtyrphealagluval
108510901095
leualaasnasniletyrleuargalalyslysgluasnleuasn
110011051110
thrasplysileserpheasptyrpheglyileprothrilegly
111511201125
asnglyargglyilealagluileargglnleutyrglulysval
113011351140
aspseraspileglnalatyralalysglyasplysproglnala
114511501155
sertyrserhisleuileaspalametleualaphecysileala
116011651170
alaaspgluhisargasnaspglyserileglyleugluileasp
117511801185
lysasntyrserleutyrproleuasplysasnthrglygluval
119011951200
phethrlysaspilepheserglnilelysilethraspasnglu
120512101215
pheserasplyslysleuvalarglyslysalailegluglyphe
122012251230
asnthrhisargglnmetthrargaspglyiletyralagluasn
123512401245
tyrleuproileleuilehislysgluleuasngluvalarglys
125012551260
glytyrthrtrplysasnserglugluilelysilephelysgly
126512701275
lyslystyraspileglnglnleuasnasnleuvaltyrcysleu
128012851290
lysphevalasplysproileserileaspileglnileserthr
129513001305
leuglugluleuargasnileleuthrthrasnasnilealaala
131013151320
thralaglutyrtyrtyrileasnleulysthrglnlysleuhis
132513301335
glutyrtyrilegluasntyrasnthralaleuglytyrlyslys
134013451350
tyrserlysglumetglupheleuargserleualatyrargser
135513601365
gluargvallysilelysserileaspaspvallysglnvalleu
137013751380
asplysaspserasnpheileileglylysilethrleuprophe
138513901395
lyslysglutrpglnargleutyrargglutrpglnasnthrthr
140014051410
ilelysaspasptyrglupheleulysserphepheasnvallys
141514201425
serilethrlysleuhislyslysvalarglysasppheserleu
143014351440
proileserthrasngluglylyspheleuvallysarglysthr
144514501455
trpaspasnasnpheiletyrglnileleuasnaspseraspser
146014651470
argalaaspglythrlyspropheileproalapheaspileser
147514801485
lysasngluilevalglualaileileaspserphethrserlys
149014951500
asnilephetrpleuprolysasnilegluleuglnlysvalasp
150515101515
asnlysasnilephealaileaspthrserlystrpphegluval
152015251530
gluthrproseraspleuargaspileglyilealathrilegln
153515401545
tyrlysileaspasnasnserargprolysvalargvallysleu
155015551560
asptyrvalileaspaspaspserlysileasntyrphemetasn
156515701575
hisserleuleulysserargtyrproasplysvalleugluile
158015851590
leulysglnserthrileilegluphegluserserglypheasn
159516001605
lysthrilelysglumetleuglymetlysleualaglyiletyr
161016151620
asngluthrserasnasn
1625
<210>2
<211>4889
<212>dna
<213>人工序列
<220>
<223>合成构建体
<400>2
atgaatttcaaaatattgccaatagcaatagatttaggtgttaaaaatactggtgtcttt60
agcgcattttatcaaaaaggaacttctcttgagagattggataataaaaatggcaaagta120
tatgaactatcaaaagattcttatactttattgatgaataatagaacagcaagaagacat180
caaagaagagggatagatagaaagcagctagtcaaaaggctctttaagcttatttggaca240
gagcagctaaatttagagtgggataaagacactcaacaagcaattagctttttatttaat300
cgtagaggttttagttttattactgatggttattcgcctgaatatttaaatattgttcca360
gagcaagtaaaagcgatacttatggatatatttgatgattacaacggtgaagatgattta420
gacagttatttaaaattagctactgagcaagaaagcaaaatttctgaaatttataacaag480
ctaatgcaaaaaatattagagtttaaattaatgaaattatgtactgatattaaggatgat540
aaagtaagtactaaaacgcttaaagaaatcacaagctatgaatttgagttattagctgat600
tatttagcaaactatagcgagagtttaaaaacacaaaaatttagttatacagataaacaa660
ggtaatttaaaagagctaagctactatcatcatgataaatataatattcaagaatttcta720
aagcgacatgctactataaatgatcgaattttagatactcttttaactgatgatttagat780
atttggaattttaattttgagaaatttgattttgataagaatgaagaaaagcttcagaat840
caggaagataaagatcatatacaagcgcatttacatcattttgtttttgcagtaaataaa900
ataaaaagtgaaatggcaagtggtggtcgtcatcgtagccaatattttcaagagataaca960
aatgtgctagatgaaaataatcatcaagagggatatctcaagaatttctgtgaaaatttg1020
cataataaaaaatattcaaatttaagtgttaaaaatttagttaatctaattggtaaccta1080
agtaatttagagctaaaaccgctaagaaaatattttaatgacaaaattcacgcaaaagct1140
gatcattgggatgagcaaaagtttacagaaacttattgccactggatattaggagagtgg1200
cgagtaggtgtcaaagatcaagataagaaagatggcgctaaatatagttataaagatctg1260
tgtaatgaattaaaacaaaaagttactaaggctggtttggtagattttttattagagtta1320
gatccatgtagaactataccaccatatctggataacaataaccgtaaaccaccaaaatgt1380
caaagtttgattttaaatccgaagtttttagataatcaatatccaaactggcaacaatat1440
ttacaagaattaaagaaactacaaagtattcaaaattatttagacagttttgaaactgat1500
ttaaaagtcttaaagtcaagtaaagatcaaccatattttgttgaatacaagagttcaaat1560
cagcaaatagcaagtggtcaaagagattataaagatttagatgctcgaatattacagttt1620
atatttgatagggtaaaagctagtgatgagttgcttttgaatgagatttattttcaggct1680
aaaaaacttaaacaaaaagctagctctgagttagaaaaactcgagtcgagcaaaaagcta1740
gatgaagttatagcaaatagtcaactatcacagatactaaagtctcaacatacaaatggt1800
atttttgaacagggtacttttttgcatttggtttgtaaatattataaacaaagacaaaga1860
gcgagagactctaggctatatattatgcctgaatatcgttatgataaaaaactacataaa1920
tataacaatacaggcaggtttgatgatgataatcagctgctaacatattgtaatcataag1980
ccaagacaaaaaagataccaattgttaaatgatttagctggggtgttgcaggtatcacct2040
aattttttgaaagataaaattggttctgatgatgatctatttattagcaaatggttggta2100
gagcatattagaggatttaaaaaagcttgtgaagatagtttaaaaatacaaaaagacaat2160
agaggattattaaatcataaaataaatatagctaggaatacaaaaggcaaatgtgaaaaa2220
gaaatatttaatttaatatgtaaaatagaaggttcagaagataaaaaaggtaattacaag2280
catggtttagcttacgaattaggagtacttttatttggtgaacctaatgaagctagtaaa2340
cctgagttcgatagaaaaattaaaaaatttaactcaatatacagttttgcacagattcaa2400
caaattgcttttgcagagcgtaaaggcaatgctaacacttgtgcagtttgtagtgctgat2460
aatgctcatagaatgcaacaaattaagatcactgagcctgtagaggacaataaagataga2520
taatcttaagtgccaaagctcagagactaccagcgattccaactagaatagttgacggtg2580
cggttaagaaaatggcaactatattagctaaaaatatagttgatgataattggcagaata2640
tcaaacaagttttatcagcaaaacatcagttacatatacctattatcacagaatcaaatg2700
cttttgagtttgaaccagcattagctgatgtaaaaggtaagagcctaaaagataggagaa2760
aaaaagcattagagagaataagtcctgaaaatatattcaaggataaaaacaatagaataa2820
aagaatttgctaaaggtatatcagcatatagtggtgctaatttaactgatggcgattttg2880
atggtgcaaaagaagaattagatcatataatacctcgttcacataaaaaatacggtactc2940
taaatgatgaagcaaatctaatttgtgtaactcgtggtgataataaaaataaaggtaata3000
gaattttctgcctacgtgatcttgcagataactataaactaaaacagtttgagacaactg3060
atgatttagaaattgaaaagaagatagctgatacaatctgggatgctaacaagaaagatt3120
ttaaatttggtaattatcgtagttttattaacctaacaccacaagagcagaaagcatttc3180
gtcacgcgctatttctggctgatgaaaatcctatcaaacaagcagtcataagagcgataa3240
ataatcgtaatcgtacatttgtaaatggcactcaacgctattttgcagaagtactggcaa3300
acaatatctatctaagggctaaaaaagaaaatctaaatacagataaaatttcatttgatt3360
attttggtattccaactataggtaatggtagaggtattgctgaaatccgtcaactttatg3420
aaaaagttgatagtgatatacaagcttatgcaaaaggtgataaacctcaagctagctact3480
ctcacctaatagatgcgatgctggctttttgtattgctgctgatgaacacagaaatgatg3540
gaagtataggtctagaaatcgataaaaattatagtttatatccattagataaaaatacag3600
gagaagtctttaccaaagatatttttagtcaaattaaaattactgataatgagtttagcg3660
ataaaaaattagtaagaaaaaaagctatagagggctttaacacgcatagacaaatgacta3720
gagatggcatttatgcagaaaattacctaccaatactaatccataaagaactaaatgaag3780
ttagaaaaggctatacttggaaaaatagtgaagaaataaaaatattcaaaggtaaaaagt3840
acgatatacaacaattgaataaccttgtgtattgtctaaaatttgtagataaacctatat3900
ctatagatatacaaattagtaccttagaagagttaagaaatatattaacaacaaataata3960
tagctgctacagcagaatactattatataaatctaaaaacccaaaaattacatgagtatt4020
atatcgaaaactataatactgccttaggttataaaaaatacagtaaagaaatggagtttt4080
tgagaagcttagcttatcgtagcgaaagggtaaaaattaaatcaatagatgatgtaaagc4140
aggttttggataaggatagtaactttatcatcggtaagattactttaccatttaaaaaag4200
agtggcaaagactatatcgtgagtggcaaaatacaactatcaaagatgattatgagtttt4260
taaaatcattctttaatgttaaaagtattactaagttgcataaaaaagttagaaaagatt4320
tctctttacctatttctacaaatgaaggtaaattcctggtcaaaagaaaaacatgggata4380
acaattttatctatcagatattaaatgattctgattctagagcagacggaacaaagccat4440
ttattccagcttttgacatttctaaaaatgaaatagtcgaagccataattgattcattta4500
catcaaaaaatattttttggctgcctaaaaatatagaattacaaaaggtggataataaaa4560
acatttttgctatagatactagtaaatggttcgaagtagaaacacctagtgatcttagag4620
acattggaatagcaacaattcaatacaagatagataataattctcgccctaaagtcagag4680
ttaaacttgattatgttatcgatgatgatagtaagataaattattttatgaatcattctt4740
tattaaaatcaagatatcctgacaaagttttagaaattttaaaacaatcaactattatag4800
aatttgaaagttcaggttttaataaaactatcaaagaaatgcttggtatgaaattagcag4860
gtatttataatgaaacatctaataattag4889
<210>3
<211>78
<212>rna
<213>人工序列
<220>
<223>合成构建体
<220>
<221>尚未归类的特征
<222>(6)..(6)
<223>其中n是任何核苷酸。
<220>
<221>尚未归类的特征
<222>(45)..(46)
<223>其中n是任何核苷酸。
<400>3
guugunuagauuauuugguauguacuuguguuaguuuaaaguagnncuagaaaauucacu60
uuuagaccuacuuauuuu78
<210>4
<211>111
<212>rna
<213>人工序列
<220>
<223>合成构建体
<400>4
guaccaaauaauuaaugcucuguaaucauuuaaaaguauuuugaacggaccucuguuuga60
cacgucugaauaacuaaaaagcaaaaauuugccaccuaaguggcuuuuuuu111
<210>5
<211>13
<212>rna
<213>人工序列
<220>
<223>合成构建体
<400>5
guaccaaauaauu13
<210>6
<211>21
<212>prt
<213>人工序列
<220>
<223>合成构建体
<400>6
metasnasnargthralaargarghisglnargargglyileasparg
151015
lysglnleuvallys
20
<210>7
<211>35
<212>prt
<213>人工序列
<220>
<223>合成构建体
<400>7
lysasnilevalaspaspasntrpglnasnilelysglnvalleuser
151015
alalyshisglnleuhisileproileilethrgluserasnalaphe
202530
glupheglu
35
<210>8
<211>31
<212>prt
<213>人工序列
<220>
<223>合成构建体
<400>8
alalysglyasplysproglnalasertyrserhisleuileaspala
151015
metleualaphecysilealaalaaspgluhisargasnaspgly
202530
<210>9
<211>4887
<212>prt
<213>智人
<400>9
atgaactttaagatcctccctattgccatcgacctgggcgtgaagaacaccggcgtgttt60
agcgccttttaccagaagggcaccagcctggagagactggataataagaacggcaaggtg120
tatgagctcagcaaggacagctataccctgctcatgaataacaggaccgctagaaggcac180
caaagaagaggcatcgacagaaagcagctggtcaagagactgttcaaactgatttggaca240
gagcaactgaacctggagtgggataaggacacccagcaggctatctccttcctcttcaac300
aggagaggcttcagcttcattaccgacggctactcccctgagtatctgaacattgtcccc360
gaacaggtcaaggccatcctgatggacatctttgacgactacaacggagaggatgatctc420
gactcctatctgaagctggctaccgaacaggaaagcaagatttccgagatctacaacaag480
ctcatgcaaaagattctggaattcaagctcatgaagctgtgtaccgatatcaaggacgac540
aaggtcagcaccaaaaccctcaaagaaatcaccagctatgaatttgagctgctggccgat600
tacctggctaattacagcgagagcctgaagacccagaagttcagctataccgataagcaa660
ggcaatctcaaggagctgagctactatcaccatgacaagtacaatattcaggagtttctg720
aagaggcatgctaccatcaatgataggatcctcgacacactgctcaccgatgacctggat780
atctggaactttaactttgagaaattcgactttgataagaatgaagaaaagctgcaaaat840
caggaagacaaggatcacattcaggctcacctgcaccacttcgtcttcgccgtcaacaag900
atcaagagcgaaatggcttccggaggcaggcacaggagccagtacttccaggaaatcacc960
aacgtcctggacgagaacaaccaccaggaaggctacctcaagaatttctgtgagaacctg1020
cacaacaagaaatatagcaacctgtccgtgaaaaacctcgtcaacctcatcggcaacctg1080
agcaatctggagctgaagcccctgaggaagtacttcaacgacaagattcatgccaaggct1140
gaccactgggacgagcagaagttcacagagacatactgtcactggatcctgggagaatgg1200
agggtgggcgtcaaagaccaggacaaaaaagatggagctaagtacagctacaaagatctg1260
tgtaatgagctcaaacagaaggtgacaaaagccggactggtggacttcctgctggagctg1320
gatccctgcaggacaattcccccctatctcgacaacaataacaggaagcctcccaagtgc1380
caaagcctcatcctcaaccccaagttcctcgacaatcagtatcccaattggcagcagtac1440
ctgcaagaactgaaaaaactgcaaagcattcaaaactacctcgattccttcgagaccgac1500
ctcaaagtcctcaaaagcagcaaggaccaaccctacttcgtcgaatacaagagcagcaac1560
cagcagatcgcctccggacagagagactacaaagacctcgacgccaggattctgcaattc1620
atcttcgacagagtcaaggcttccgacgaactgctgctgaatgaaatctattttcaagct1680
aaaaagctcaagcagaaagccagcagcgaactcgaaaaactggagtcctccaagaaactc1740
gacgaggtgattgccaatagccaactcagccagatcctgaagagccagcatacaaatggc1800
atcttcgagcaaggcacatttctgcatctggtgtgtaaatactacaaacaaagacagagg1860
gctagggacagcagactctatatcatgcccgagtacagatacgataaaaaactgcataaa1920
tacaacaacaccggcaggtttgacgacgataaccaactgctcacctactgcaaccacaag1980
cctaggcaaaaaaggtatcagctgctgaacgacctggctggagtgctccaagtctcccct2040
aatttcctcaaggataaaattggatccgacgatgacctcttcatctccaagtggctggtc2100
gagcacatcagaggcttcaagaaggcctgcgaagattccctgaaaatccagaaggacaac2160
aggggactcctgaatcataagattaatatcgctagaaataccaagggcaaatgcgagaag2220
gagatcttcaacctgatctgcaaaatcgaaggctccgaggataagaaaggcaactataag2280
catggcctggcttatgagctcggagtgctcctgttcggagagcccaatgaggcctccaag2340
cctgaatttgacaggaagatcaagaagtttaatagcatctactccttcgcccagatccaa2400
caaatcgccttcgctgaaaggaagggcaacgctaacacctgcgccgtgtgcagcgctgat2460
aatgctcacaggatgcagcagatcaagatcacagaacccgtggaagacaataaagacaag2520
atcatcctcagcgctaaggctcagagactgcccgctattcctacaagaatcgtggacgga2580
gccgtcaagaaaatggccaccatcctggccaaaaacatcgtggatgataattggcaaaat2640
attaaacaggtcctgtccgccaagcaccagctccacattcccatcatcaccgagtccaat2700
gctttcgagttcgaacccgccctggctgacgtgaaaggcaaatccctcaaggacagaaga2760
aagaaggccctggagagaatttcccctgagaacatctttaaggacaaaaataacagaatt2820
aaagagtttgctaagggaatttccgcctacagcggcgccaatctgacagatggcgacttc2880
gatggcgctaaagaagagctcgaccacatcattcccagaagccacaagaagtatggaacc2940
ctcaacgatgaggccaacctcatctgcgtcaccaggggcgacaataaaaataaaggcaat3000
aggatcttctgtctgagagacctggccgataactacaaactgaaacagttcgaaaccacc3060
gacgacctggagattgagaagaaaatcgccgacaccatctgggacgctaataaaaaagac3120
tttaagttcggaaactacaggagcttcattaacctgacaccccaggaacagaaagccttt3180
aggcatgccctctttctggccgatgagaaccctatcaagcaagccgtcatcagggccatc3240
aacaacaggaataggaccttcgtcaatggcacccagaggtactttgccgaggtgctggcc3300
aataacatctatctcagggctaaaaaggagaatctcaatacagacaaaatctcctttgac3360
tattttggaatccctaccatcggaaatggcaggggaatcgctgagattagacagctgtac3420
gagaaagtcgacagcgatatccaagcctacgccaagggagataagcctcaggcttcctat3480
agccacctcatcgacgctatgctggccttttgcatcgccgccgacgagcacagaaatgat3540
ggctccatcggactggaaatcgacaagaattacagcctctaccccctcgacaaaaacaca3600
ggagaggtgttcacaaaagatattttcagccagattaagattacagacaacgaatttagc3660
gataagaaactggtgagaaagaaagctatcgagggatttaatacccataggcaaatgacc3720
agggacggcatttacgctgagaactatctccccatcctcatccacaaggaactgaacgaa3780
gtcagaaaaggatatacctggaaaaatagcgaggaaattaagattttcaaaggaaaaaag3840
tatgacatccagcagctcaacaacctcgtgtattgcctcaagttcgtggacaagcccatt3900
tccatcgacatccagatcagcacactggaagagctgaggaatatcctgaccacaaataac3960
attgccgctaccgctgagtattattacattaatctcaaaacacagaaactgcatgaatat4020
tacatcgagaactacaataccgccctgggctataagaagtattccaaggaaatggagttc4080
ctcaggtccctcgcctataggagcgagagggtgaagattaagagcatcgacgatgtcaag4140
caggtgctggacaaggatagcaacttcattattggaaaaatcacactcccctttaagaag4200
gagtggcagaggctgtacagggagtggcaaaacaccacaatcaaggacgattacgagttc4260
ctgaagagcttctttaacgtgaagagcattacaaagctgcacaagaaggtcaggaaagac4320
ttcagcctccccattagcaccaacgagggaaagttcctggtgaagaggaagacctgggac4380
aacaacttcatctaccagatcctcaatgactccgacagcagggccgacggcacaaagccc4440
tttatccctgccttcgacatcagcaagaacgaaatcgtggaggccatcatcgattccttt4500
accagcaaaaacattttctggctgcccaaaaatattgaactccagaaggtcgacaacaaa4560
aacatctttgctatcgacacatccaaatggtttgaagtcgagacaccttccgacctgagg4620
gatatcggaattgccaccattcaatataagatcgacaataatagcaggcctaaagtgagg4680
gtcaaactcgactacgtgatcgacgacgacagcaagatcaactacttcatgaaccacagc4740
ctgctgaagtccaggtatcccgacaaggtcctcgaaatcctcaagcagagcaccatcatt4800
gaatttgagtccagcggattcaacaagacaatcaaagagatgctgggcatgaaactcgcc4860
ggcatctataacgagaccagcaataac4887
<210>10
<211>63
<212>rna
<213>智人
<400>10
uaucaggcaguaccacaagcucguaauuaauaaaccaugaaaguaugguuuauuagauug60
uug63
<210>11
<211>44
<212>rna
<213>智人
<400>11
cucguaauuaauaaaccaugaaaguaugguuuauuagauuguug44
<210>12
<211>87
<212>rna
<213>智人
<400>12
guaucaggcaguaccacaagcucguaauuaauaaaccaugaaaguaugguuuauuagauu60
guugaaggcuaguccguuaucaacuug87
<210>13
<211>46
<212>rna
<213>人工序列
<220>
<223>合成构建体
<220>
<221>尚未归类的特征
<222>(1)..(1)
<223>其中n是任何核苷酸
<220>
<221>尚未归类的特征
<222>(46)..(46)
<223>其中n是任何核苷酸
<400>13
ncucguaauuaauaaaccaugaaaguaugguuuauuagauuguugn46
<210>14
<211>14
<212>rna
<213>人工序列
<220>
<223>合成构建体
<220>
<221>尚未归类的特征
<222>(14)..(14)
<223>其中n是任何核苷酸
<400>14
guaccaaauaauun14
- 还没有人留言评论。精彩留言会获得点赞!