具有中和活性的抗新冠病毒兔重组单克隆抗体及应用的制作方法
1.本发明属于生物技术中的抗体制备领域,特别是涉及具有中和活性的抗新冠病毒sars-cov2-s-rbd兔重组单克隆抗体及其制备方法和应用。
背景技术:
2.新型冠状病毒2019-ncov又称sars-cov-2,人感染该病毒后主要出现呼吸道症状、发热、咳嗽以及呼吸困难等,严重者可引起肺炎、严重急性呼吸综合征、肾衰竭甚至死亡。尽管疫苗接种是预防新型冠状病毒最为有效的方法,但是目前还没有批准上市的疫苗产品,而对于已感染的病人,目前也没有特异性的治疗药物。不过,已有大量证据证明康复患者的血浆可以有效的治疗新冠患者,其中有效的成分是血浆中含有高浓度的中和抗体。因此,开发中和活性的抗体可能是治疗新型冠状病毒的一个有效的候选措施。
3.新型冠状病毒属于单股正链rna病毒,病毒粒子表面外包着脂肪膜,膜表面主要有三种糖蛋白,其中刺突糖蛋白(spike蛋白)有两个亚基即s1和s2,其中s1亚基上有一个受体ace2蛋白的结合位点(rbd),介导宿主细胞的融合。目前对该位点的中和活性抗体开发的文献已经有很多,一部分是通过康复病毒血液中分离特异性b细胞获得全人源的单克隆抗体,一部分是通过噬菌体展示技术获得人源化的单克隆抗体,但是还没有真正上市的产品。例如,中国专利申请cn2020101393840中涉及sars-cov-2抑制剂及其应用,其是从患者恢复期外周血中的pbmc细胞,通过噬菌体展示技术制备了对sars-cov-2病毒有较强的中和作用的抗体。cn2020108392266等专利申请也均通过患者恢复期外周血中的pbmc细胞制备单克隆抗体。
4.由于单一中和性抗体的阻断效果有限,在研究中发现通过鸡尾酒方式将多个中和活性的抗体联用,可以获得更佳的抗病毒的效果。由于兔抗体的亲和力相对其他来源的抗体要高,而且抗体表位覆盖面要广,因此开发针对新冠病毒rbd蛋白兔源的具有中和活性的单克隆抗体可为治疗性中和抗体的开发提供更多的选择。
技术实现要素:
5.本发明目的在于提供具有中和活性的抗新冠病毒sars-cov2-s-rbd兔重组单克隆抗体,以阻断ace2蛋白和rbd蛋白的作用,选自以下兔重组单克隆抗体的一种或多种:命名为2b6的兔重组单克隆抗体:其重链互补决定区cdr1、cdr2、cdr3的氨基酸序列分别为seq id no:1、seq id no:2、seq id no:3所示的氨基酸序列;且轻链互补决定区cdr1、cdr2、cdr3的氨基酸序列分别为seq id no:4,seq id no:5、seq id no:6所示的氨基酸序列;命名为2h5的兔重组单克隆抗体:其重链互补决定区cdr1、cdr2、cdr3的氨基酸序列分别为seq id no:10、seq id no:11、seq id no:12所示的氨基酸序列,且轻链互补决定区cdr1、cdr2、cdr3的氨基酸序列分别为seq id no:13、seq id no:14、seq id no:15所示的氨基酸序列;
命名为4b1的兔重组单克隆抗体,其重链互补决定区cdr1、cdr2、cdr3的氨基酸序列分别为seq idno:19、seq id no:20、seq id no:21所示的氨基酸序列,且轻链互补决定区cdr1、cdr2、cdr3的氨基酸序列分别为seq id no:22、seq id no:23、seq id no:24所示的氨基酸序列;命名为6b10的兔重组单克隆抗体,其重链互补决定区cdr1、cdr2、cdr3的氨基酸序列分别为seq id no:28、seq id no:29、seq id no:30所示的氨基酸序列,且轻链互补决定区cdr1、cdr2、cdr3的氨基酸序列分别为seq id no:31、seq id no:32、seq id no:33所示的氨基酸序列;命名为9d5的兔重组单克隆抗体,其重链互补决定区cdr1、cdr2、cdr3的氨基酸序列分别为seq id no:37、seq id no:38、seq id no:39所示的氨基酸序列,且轻链互补决定区cdr1、cdr2、cdr3的氨基酸序列分别为seq id no:40、seq id no:41及seq id no:42所示的氨基酸序列;在上述技术方案的基础上,命名为2b6的兔重组单克隆抗体,其重链可变区序列为seq id no:7所示的氨基酸序列,且轻链可变区序列为seq id no:8所示的氨基酸序列;命名为2h5的兔重组单克隆抗体,其重链可变区序列为seq id no:16所示的氨基酸序列,且轻链可变区序列为seq id no:17所示的氨基酸序列;命名为4b1的兔重组单克隆抗体,其重链可变区序列为seq id no:25所示的氨基酸序列,且轻链可变区序列为seq id no:26所示的氨基酸序列;命名为6b10的兔重组单克隆抗体,其重链可变区序列为seq id no:34所示的氨基酸序列,且轻链可变区序列为seq id no:35所示的氨基酸序列;命名为9d5的兔重组单克隆抗体,其重链可变区序列为seq id no:43所示的氨基酸序列,且轻链可变区序列为seq id no:44所示的氨基酸序列;在上述技术方案的基础上,命名为2b6的兔重组单克隆抗体,其scfv序列为seq id no:9所示的氨基酸序列;命名为2h5的兔重组单克隆抗体,其scfv序列为seq id no:18所示的氨基酸序列;命名为4b1的兔重组单克隆抗体,其scfv序列为seq id no:27所示的氨基酸序列;命名为6b10的兔重组单克隆抗体,其scfv序列为seq id no:36所示的氨基酸序列;命名为9d5的兔重组单克隆抗体,其scfv序列为seq id no:45所示的氨基酸序列;在上述技术方案的基础上,兔重组单克隆抗体的轻链恒定区为κ链,重链恒定区为igg型。
6.本发明还提供一种核酸分子,其包含能够编码具有中和活性的抗新冠病毒sars-cov2-s-rbd兔重组单克隆抗体的重链互补决定区或轻链互补决定区的核酸序列。
7.本发明还提供一种载体,其含有上述核酸分子。
8.本发明还提供一种宿主细胞,该宿主细胞含有上述具有中和活性的抗新冠病毒sars-cov2-s-rbd兔重组单克隆抗体、上述核酸分子或上述载体。
9.本发明还提供一种偶联物,含有上述抗体。
10.本发明还提供一种药物组合物,含有主成分和辅成分,其中:主成分上述具有中和活性的抗新冠病毒sars-cov2-s-rbd兔重组单克隆抗体、上述核酸分子、上述载体、上述宿
主细胞、上述偶联物中的一种或多种,辅成分选自药学上可接受的载体或赋形剂,以及任选的其它生物活性物质。
11.本发明还提供上述具有中和活性的抗新冠病毒sars-cov2-s-rbd兔重组单克隆抗体、上述酸分子上述载体、上述宿主细胞、上述偶联物在制备治疗疾病的药物或检测试剂中的应用。
12.本发明还提供一种试剂盒,该试剂盒包含上述具有中和活性的抗新冠病毒sars-cov2-s-rbd兔重组单克隆抗体。
13.与现有技术相比,本发明具有以下优点和有益效果:本发明利用真核表达的新冠病毒rbd蛋白免疫新西兰大白兔,采用 b细胞克隆技术获得了具有中和活性的抗新冠病毒sars-cov2-s-rbd兔重组单克隆抗体,重要的是该兔重组单克隆抗体亲和力高,具有良好的特异性,同时可以阻断ace2蛋白和rbd蛋白的作用。另外,利用免疫的兔b细胞制备中和活性抗体,不存在伦理问题、感染风险、病毒外泄等,制作成本较低。本技术的兔单克隆抗体可以弥补市场上具有中和活性的抗新冠病毒抗体的诊断与治疗相关的应用。由于单一中和性抗体的阻断效果有限,本发明制备了sars-cov2-s-rbd兔重组单克隆抗体,可以通过鸡尾酒方式与其他来源的多个中和抗体联用,获得更佳的抗病毒的效果。本技术所制备的兔抗体的亲和力相对其他来源的抗体要高,而且抗体表位覆盖面要广。同时,后续我们可以将兔源单克隆进行人源化改造,可用于后续开发治疗新冠病毒感染的中和性抗体或试剂。
14.附图说明
15.为了更清楚地说明本发明实施例中的技术方案,下面将对实施例描述中所需要使用的附图作简单地介绍,显而易见地,下面描述中的附图仅仅是本发明的一些实施例,对于本领域普通技术人员来讲,在不付出创造性劳动的前提下,还可以根据这些附图获得其他的附图。
16.图1显示了rbd蛋白sds-page鉴定图。
17.图2 显示了抗体可变区pcr扩增结果。
18.图3 显示了抗体表达纯化sds-page鉴定图。
19.图4 显示了抗体elisa效价测定结果。
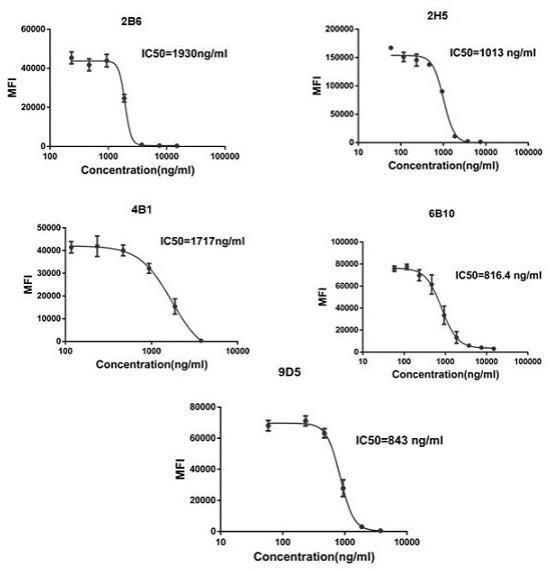
20.图5显示了竞争facs检测rbd兔重组单克隆抗体中和效价ic50。
具体实施方式
21.为使本发明实施例的目的、技术方案和优点更加清楚,下面将结合本发明实施例中的附图,对本发明实施例中的技术方案进行清楚、完整地描述,显然,所描述的实施例是本发明的一部分实施例,而不是全部的实施例。基于本发明中的实施例,本领域普通技术人员在没有做出创造性劳动的前提下所获得的所有其他实施例,都属于本发明保护的范围。
22.以下对本发明做进一步描述:在本发明中,除非另有说明,否则本文中使用的科学和技术名词具有本领域技术人员所通常理解的含义。并且,本文中所用的蛋白质和核酸化学、分子生物学、细胞和组织培养、微生物学、免疫学相关术语和实验室操作步骤均为相应
领域内广泛使用的术语和常规步骤。同时,为了更好地理解本发明,下面提供相关术语的定义和解释。
23.这里提到的术语“抗体”包括完整抗体及其任何抗原结合片段 (即“抗原结合部分”)或单链。“抗体”是指包含通过二硫键互相连接在一起的至少两条重(h)链和两条轻(l)链的糖蛋白,或其抗原结合部分。每条重链由重链可变区和重链恒定区组成。本发明中涉及的蛋白或其片段可以是天然纯化的产物,或是化学合成的产物,或使用重组技术从真核宿主(例如哺乳动物细胞)中产生。本发明所用原料及试剂均可由市场购得。
24.下面结合实施例,进一步阐述本发明的技术方案。
25.实施例:(1)rbd蛋白的制备:为了利于纯化,本发明构建了新冠病毒rbd fc融合标签蛋白真核表达质粒(ptt5-rbd-fc),转染293f细胞5天后,收集细胞培养基上清,利用proteina树脂进行纯化,并对蛋白的浓度进行鉴定。通过sds-page方式对所获得的rbd蛋白质进行分析,结果如图1所示,获得约为51.3kda的蛋白质即为rbd蛋白质。
26.(2)免疫动物外周血单个核细胞(即pbmc细胞)的获取:选择新西兰大白兔作为免疫动物。第一次免疫时将250μg rbd蛋白与等体积的完全弗氏佐剂进行乳化,对新西兰大白兔皮下背部多点进行注射,21天后进行第二次免疫,将120μg的rbd蛋白与等体积的不完全弗氏佐剂进行乳化,对新西兰大白兔背部皮下注射,21天后进行第三次免疫,即将120μg的rbd蛋白与等体积的不完全弗氏佐剂进行乳化,对新西兰大白兔背部皮下注射。第三次免疫后间隔一周,无菌采集外周血。
27.(3)rbd特异性b淋巴细胞的获取:用淋巴细胞分离液将所采集的外周血中的pbmc细胞分离出来;按照免疫磁珠操作说明,将rbd蛋白偶联到磁珠上;将rbd蛋白偶联的磁珠与分离出的pbmc细胞共同室温孵育50min后的混合物放入磁力架中,5min后磁珠全部沉入底部,弃掉上清,加入无菌的pbs洗涤细胞,重复洗细胞3次,最后分离得到的细胞即为rbd特异性的b淋巴细胞。
28.(4)b淋巴细胞的鉴定:将分离获得的b淋巴细胞进行一定倍数的稀释后置入96孔细胞培养板,加入含10%胎牛血清(fbs)和2μg/ml的人il2的1640培养基,37℃ 5% co2条件下培养6天。收集培养基上清进行抗体的鉴定。
29.a)间接elisa鉴定包被浓度为1μg/ml的rbd蛋白,100μl/孔,4℃孵育16h;次日弃掉包被液后用含1%牛血清白蛋白(bsa)的pbs进行封闭,150μl/孔,37℃孵育1h。弃掉封闭液,加入b细胞上清,50μl/孔,37℃孵育1h。弃掉b细胞上清,用含0.5wt.%的tween-20的pbs洗板5次,2min/次,最后加入稀释5000倍的羊抗兔igg-hrp二抗,37℃孵育1h。弃掉二抗,用磷酸盐吐温缓冲液(pbst)洗板5次,2min/次。弃掉洗涤液,拍干,加入底物进行显色。结果见表1,标记为2b6,2h5,4b1,6b10,9d5的孔对应的b淋巴细胞结果为阳性。
30.表1. elisa测试结果
ꢀ
(5)抗体基因的克隆对经鉴定为阳性的b细胞进行收集,利用常规的rna提取方法提取rna后,反转录为cdna,利用抗体重链的基因引物即:上游引物5
’‑
cagtcgctggaggagtccgg-3’和下游引物5
’‑
ccattggtgagggtgcccgag-3’,抗体轻链基因的引物即:上游引物5
’‑
gacattgtgatgacccagac-3’和下游引物5
’‑
ccacctcggtcccttcgccg-3’,对抗体重轻链基因进行扩增,其扩增条件为:94℃ 3min进行变性,(95℃ 1min,56℃ 30s,72℃1min)进行30个循环反应,最后72℃延伸10min(pcr扩增结果见图2)。利用dna胶纯化回收试剂盒将pcr产物进行回收。将兔重组单克隆抗体的重轻链基因克隆至表达载体后进行转化,利用菌落pcr验证单菌落,将阳性菌落进行基因测序,即可获得特异性抗体的基因序列。通过标记为2b6,2h5,4b1,6b10,9d5的b淋巴细胞最终所获得的特异性抗体的基因序列及其氨基酸序列见表2所示。
31.表2. 特异性抗体的基因序列及其氨基酸序列(6)rbd兔重组单克隆抗体的生产和鉴定将抗体的重轻链基因的表达质粒共转染入293细胞,37℃ 5% co2培养条件下培养72h,收集细胞上清,利用proteina树脂进行抗体的纯化,纯化后定的抗体进行sds-page染色,结果见图3。
32.纯化后的抗体进行功能鉴定,具体如下:
a) elisa反应包被浓度为1μg/ml的rbd蛋白,100μl/孔,4℃孵育16h;次日弃掉包被液后用含1%bsa的pbs进行封闭,150μl/孔,37℃孵育1h。弃掉封闭液,加入不同倍比稀释的抗体,50μl/孔,37℃孵育1h。弃掉抗体,用含0.5%的tween-20的pbs洗板5次,2min/次,最后加入稀释5000倍的羊抗兔igg-hrp二抗,37℃孵育1h。弃掉二抗,用pbst洗板5次,2min/次。弃掉洗涤液,拍干,加入底物进行显色。实验结果见图4,证明2b6,2h5,4b1,6b10,9d5这5株抗体均能与rbd蛋白反应。
33.b) 基于流式的中和活性测定方法用293细胞转染ace2蛋白的质粒进行鉴定。将不同浓度的抗体与rbd-mfc蛋白于37℃中反应1h后,将上清混合物与处理的转染ace2质粒的293细胞进行反应,,4℃孵育30min。(细胞的处理方式为收集细胞于离心管中,无菌的pbs洗细胞两次,采用fc受体封闭液进行细胞表面fc受体的封闭,4℃孵育30min。离心收集细胞,用含0.5wt.%bsa的pbs洗细胞两次,加入b细胞上清,4℃孵育30min)。再用含0.5wt.%bsa的pbs洗细胞两次,最后加入羊抗鼠igg-488荧光二抗,4℃孵育30min。用含0.5wt.%bsa的pbs洗细胞两次后,用200μl 含0.5wt.%bsa的pbs重悬细胞,流式机器上机测试。实验结果见图5,证明2b6,2h5,4b1,6b10,9d5这5株抗体均能阻断rbd蛋白与ace2蛋白的结合。
34.通过以上实验证明所获得的5株抗rbd蛋白的兔重组单克隆抗体能够特异性的识别rbd蛋白,同时能够阻断rbd蛋白与ace2蛋白的结合。
35.以上所述,仅为本发明较佳的具体实施方式,但本发明的保护范围并不局限于此,任何熟悉本技术领域的技术人员在本发明揭露的技术范围内,可轻易想到的变化或替换,都应涵盖在本发明的保护范围内。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1