杜梨耐盐基因PbHSF3A及其在植物耐盐遗传改良中的应用
杜梨耐盐基因pbhsf3a及其在植物耐盐遗传改良中的应用
技术领域
1.本发明涉及基因工程技术领域,特别是涉及一种杜梨耐盐基因pbhsf3a及其在植物耐盐遗传改良中的应用。
背景技术:
2.梨是当今世界广泛栽种的果树经济作物,因为其独特的营养风味和口感,以及经济价值而受到广大人民群众的喜爱。据统计,梨是全球主要栽培的经济果树树种之一,但其极易受不同区域间复杂的地理气候等不利条件的影响,如土壤瘠薄缺水、冻害,以及沿海地区的高度盐碱化等。因此,选育抗逆性强、综合性状优良的新品种已经成为梨产业发展最关键的因素。
3.在实际的栽培生产时常常会受到干旱、涝害、盐碱地等非生物胁迫的危害。盐碱土在很大程度上会影响到果树的正常生长,而梨树也不例外,盐碱土中不断增加的盐分导致可耕地转化为非生产性荒地,盐碱地由于盐分影响植物体中的矿质离子稳态、水分平衡、固氮和光合能力等一系列生理生化特性的累积作用,导致植物产量大面积的减少。因此,选育耐盐功能强的梨栽培品种已然迫在眉睫,其可有效地减少盐碱土对梨产业造成的危害。
技术实现要素:
4.本发明的目的是提供一种杜梨耐盐基因pbhsf3a及其在植物耐盐遗传改良中的应用,以解决上述现有技术存在的问题,探索盐胁迫对梨种质资源的影响,并从形态指标以及测定生理生化指标来评定多种梨种质资源的耐盐性程度,从而选出耐盐性程度更高的梨品种,并为研究梨耐盐分子机理等提供理论依据。
5.为实现上述目的,本发明提供了如下方案:
6.本发明提供一种杜梨耐盐蛋白pbhsf3a,该基因是从杜梨中分离、克隆出的一个热激蛋白,将其命名为pbhsf3a,其氨基酸序列如seq id no.2所示:
7.mspkdkshpkspptsaefdpesiglsglsefrlevsepllgsqpipsftspvmefeafssvnplgafdftekvsiptssmggggaedvvvppqpleclqgtlvppflsktfdlvedpsldsiiswgsggrsfvvwdpsefsrfilprnfkhnnfssfvrqlntyivskvqfficmkvdgltlssgfrkvdtdkwefaneafqkgkrhllkriqrrkspqslqvgsftgpsaeagksgvegeietlqkersmlmqevvelqqqqrgtvdrmkvvnqrlqsaeqrqkqmvsflakmlqnpafvarlqqktgqkdrgssrvrrkfvkhqqhelsksdsdmegqivkyqpawrnqiisstapdsnpvpfeqsphypsqvttgklgldaestafqfvdaaldelaitqgflqtpeqegegassmvtedpffkgksvlspqqeanpkhyvsfeedllkdrtfpelfspgmesmikqediwsmdfdisagmstsmnellgnpvnydvpevgvtgelldvwdidplqeagglginkwpahesafdepqsqgvqsaskttdp。
8.本发明还提供编码上述的杜梨耐盐蛋白pbhsf3a的杜梨耐盐基因pbhsf3a。
9.进一步地,所述的杜梨耐盐基因pbhsf3a,其核苷酸序列如seq id no.1所示:
10.atgagcccaaaagacaagagccacccaaagtctccacccacttcagctgaattcgacccggaatcaattgggttatcggggttatcggaatttcgactcgaagtgtcggaaccgttactgggttctcagccgattccttcgttc
acttctcctgtaatggaattcgaagccttctcttctgtaaacccattgggtgcttttgacttcactgaaaaagtttcaattccaacgtcgtctatgggcggcggtggtgcggaggacgtcgttgttccgccgcagccactggaatgtctgcagggcactctggtgcctccatttctgtccaagactttcgatttggtcgaggacccctcgctggattcgatcatatcgtggggctccggcggccgcagcttcgtggtgtgggacccatcggagttttccagattcatcttaccgaggaacttcaagcacaacaatttctcaagttttgtgcggcagcttaatacttatattgtaagcaaggttcagttcttcatatgcatgaaagttgatggcctcacattgtcttctggttttcgcaaggttgacacagataagtgggagtttgcgaatgaagcttttcagaagggcaaaagacatttgttgaagaggatccagaggcgcaagtcacctcagtctctgcaagttgggagctttactgggccttctgcggaagcagggaagtctggagtcgaaggtgagatagagacattgcagaaagagaggagtatgttaatgcaggaggttgttgaattgcagcagcagcagcggggcacagttgaccgtatgaaggtagtgaatcagaggcttcagtctgcggagcagagacagaagcagatggtttctttcttggccaagatgcttcagaacccggcattcgtggcccgacttcaacaaaagactggacagaaagacagaggctcttcaagggtgaggaggaaatttgttaagcatcagcaacatgaactcagtaagtcagattcagatatggaaggccagattgtgaagtaccagcctgcttggagaaaccaaatcatatcctctacagcgccggattcaaatccagttccttttgaacaatctcctcattatccttcacaagttacgacaggaaaactgggtttggatgctgaaagcacggcattccaatttgtggacgctgcactagatgaattagcaatcacacaaggatttcttcaaacaccagagcaagaaggcgaaggggcatcaagcatggtaactgaagatccttttttcaaagggaagagtgttctaagcccacaacaagaggctaatcccaagcattatgtctctttcgaggaagatttgttgaaggacaggacttttccagaactgttttctccagggatggagagcatgattaaacaagaagacatatggagcatggattttgatatcagtgccggtatgtcaacttctatgaatgagttattgggtaatccagtcaactatgatgtgccagaggtgggagtgaccggtgaattgttagatgtctgggatatcgatcccctgcaagaggcaggagggttgggaatcaataagtggccagcccatgaatctgcatttgatgagcctcagagtcaaggtgtccagtctgcatctaaaactactgatccgtag。
11.本发明还提供一种植物超表达载体,含有上述的杜梨耐盐基因pbhsf3a。
12.本发明还提供一种上述的杜梨耐盐基因pbhsf3a在提高植物盐胁迫耐受性中的应用。
13.利用本领域的常规方式,将pbhsf3a基因在植株中进行超表达,可获得耐盐的转基因植株;沉默植株中pbhsf3a基因的表达,则可获得耐盐能力降低的转基因植株。
14.进一步地,所述植物选自杜梨或拟南芥。
15.本发明还提供一种上述的杜梨耐盐基因pbhsf3a在控制植物耐盐性能中的应用,其特征在于,将所述杜梨耐盐基因pbhsf3a在植物中超表达或沉默表达。
16.从杜梨中分离克隆出pbhsf3a基因,将该基因在拟南芥中超表达,获得的转基因植物耐盐性明显提高,将该基因在杜梨中沉默掉之后,沉默植株耐盐性明显降低。
17.本发明还提供一种提高植物盐胁迫耐受性的方法,利用上述的植物超表达载体,采用农杆菌介导的遗传转化方法将杜梨耐盐基因pbhsf3a导入植株中。
18.本发明公开了以下技术效果:
19.本发明通过分析在盐处理前后pbhsf3a转基因株系的表型及相关生理指标,结果表明:相对于非转pbhsf3a基因株系,pbhsf3a转基因株系具有明显的优势,例如,发芽率、叶绿素含量、fv/fm、钾离子含量及抗超氧阴离子含量较高,电导率、钠离子含量、过氧化氢含量及mda含量较低,而pbhsf3a瞬时沉默株系则与之相反,表明pbhsf3a基因是一个潜在的耐盐育种基因,可用于植物耐盐遗传改良。
附图说明
20.为了更清楚地说明本发明实施例或现有技术中的技术方案,下面将对实施例中所需要使用的附图作简单地介绍,显而易见地,下面描述中的附图仅仅是本发明的一些实施例,对于本领域普通技术人员来讲,在不付出创造性劳动的前提下,还可以根据这些附图获得其他的附图。
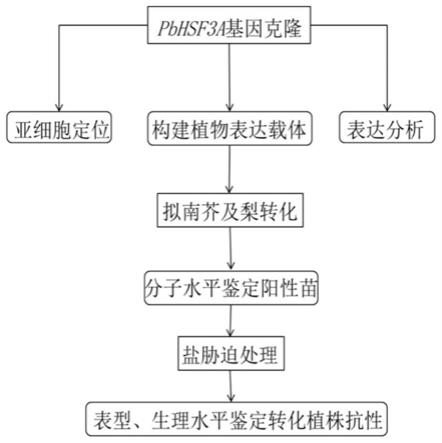
21.图1是本发明的技术流程图;
22.图2是本发明的热激蛋白pbhsf3a在非生物胁迫下的表达示意图;其中,a为盐处理下热激蛋白pbhsf3a的转录相对表达量;b为脱水处理下热激蛋白pbhsf3a转录水平的相对表达量;c为低温胁迫下热激蛋白pbhsf3a转录水平的相对表达量;
23.图3是本发明pbhsf3a基因亚细胞定位示意图;
24.图4是本发明pbhsf3a基因转基因拟南芥鉴定及相对表达量分析示意图;其中a为转基因阳性幼苗检测结果;m:dl2000的marker;
‑
:阴性对照wt植株;+:质粒阳性对照;1
‑
4:转基因阳性拟南芥植株;b为超表达pbhsf3a基因拟南芥植株pcr扩增结果;line1和line2为转基因拟南芥的凝胶电泳条带,wt为未转化拟南芥植株;
25.图5是本发明pbhsf3a转基因拟南芥的抗性分析;其中,a是90mm nacl浓度下野生型植株(wt)与转基因植株(oe5、oe8)的种子萌发情况;b是90mm nacl浓度下拟南芥种子萌发率;c是90mm nacl浓度下野生型植株(wt)与转基因植株(oe5、oe8)的根长表型;d是90mm nacl浓度下拟南芥植株的根长;e是90mm nacl浓度下拟南芥植株的电导率测定;
26.图6是本发明pbhsf3a转基因拟南芥耐盐性分析;其中,a
‑
b是30d正常生长盐胁迫处理前的野生型株系(wt)和转基因株系(oe5和oe8)的表型(a)和盐胁迫处理10d后的表型(b);c是90mm nacl浓度胁迫10d后拟南芥的fv/fm;d是盐胁迫后拟南芥的叶绿素含量图;e是盐胁迫后拟南芥的叶绿素表型图;f是90mm nacl浓度胁迫10d后拟南芥的电导率;g是90mm nacl浓度胁迫10d后拟南芥的mda含量;
27.图7是本发明pbhsf3a转基因拟南芥盐胁迫后na
+
/k
+
的测定;其中,a是盐胁迫10d后野生型株系与转基因株系中钠离子的含量;b是盐胁迫10d后野生型株系与转基因株系中钾离子的含量;c是盐胁迫10d后野生型株系与转基因株系中钠钾比值;
28.图8是本发明pbhsf3a转基因拟南芥盐胁迫后活性氧分析;其中,a
‑
b是盐胁迫10d后dab染色,检测h2o2的含量(a)和盐胁迫10d后nbt染色,检测anti o2‑
的含量(b);c
‑
d是分别与a和b对应,表示盐胁迫10d后h2o2的含量测定(c)和盐胁迫10d后anti o2‑
的含量测定(d);
29.图9是本发明pbhsf3a基因沉默杜梨盐胁迫下的抗性鉴定;其中,a
‑
b是35d正常生长杜梨pbhsf3a基因沉默株系盐胁迫处理前表型(a)和盐胁迫处理18d后的表型(b);c是pbhsf3a基因沉默株系盐胁迫后的fv/fm;d是盐胁迫后pbhsf3a基因沉默株系的叶绿素含量图;e是盐胁迫后pbhsf3a基因沉默株系的叶绿素表型图;f是盐胁迫18d后的电导率;g是盐胁迫18d后的mda含量;
30.图10是本发明pbhsf3a基因沉默杜梨盐胁迫后na
+
/k
+
的测定;其中,a是盐胁迫18d后野生型株系与pbhsf3a基因沉默株系中钠离子的含量;b是盐胁迫18d后野生型株系与pbhsf3a基因沉默株系中钾离子的含量;c是盐胁迫18d后野生型株系与pbhsf3a基因沉默株系中钠钾比值。
具体实施方式
31.现详细说明本发明的多种示例性实施方式,该详细说明不应认为是对本发明的限制,而应理解为是对本发明的某些方面、特性和实施方案的更详细的描述。
32.应理解本发明中所述的术语仅仅是为描述特别的实施方式,并非用于限制本发明。另外,对于本发明中的数值范围,应理解为还具体公开了该范围的上限和下限之间的每个中间值。在任何陈述值或陈述范围内的中间值以及任何其他陈述值或在所述范围内的中间值之间的每个较小的范围也包括在本发明内。这些较小范围的上限和下限可独立地包括或排除在范围内。
33.除非另有说明,否则本文使用的所有技术和科学术语具有本发明所述领域的常规技术人员通常理解的相同含义。虽然本发明仅描述了优选的方法和材料,但是在本发明的实施或测试中也可以使用与本文所述相似或等同的任何方法和材料。本说明书中提到的所有文献通过引用并入,用以公开和描述与所述文献相关的方法和/或材料。在与任何并入的文献冲突时,以本说明书的内容为准。
34.在不背离本发明的范围或精神的情况下,可对本发明说明书的具体实施方式做多种改进和变化,这对本领域技术人员而言是显而易见的。由本发明的说明书得到的其他实施方式对技术人员而言是显而易见得的。本发明说明书和实施例仅是示例性的。
35.关于本文中所使用的“包含”、“包括”、“具有”、“含有”等等,均为开放性的用语,即意指包含但不限于。
36.实施例1
37.pbhsf3a基因克隆及超表达载体构建
38.1、rna提取
39.研究材料杜梨种植在南京农业大学梨工程中心,其苗龄为45d。挑选生长势健壮的杜梨幼苗,随机称取0.1g样品,迅速用液氮进行速冻。rna的提取采用索来宝公司的总rna提取试剂盒,具体方法如下:
40.(1)样品处理:取新鲜或
‑
80℃冻存0.1g组织在液氮中研磨,把粉末加入到1ml裂解液中混匀,得到匀浆样品;
41.(2)将处理后的样品在室温放置5min,使得核酸蛋白复合物完全分离;
42.(3)向室温放置后的匀浆样品中加0.2ml氯仿,盖好管盖,剧烈振荡15秒,室温放置3
‑
5min,得到混悬液;
43.(4)将混悬液在2到8℃、12000rpm的条件下离心10min,rna主要在上层无色的水相中,把水相转移到新管中,不要吸到沉淀,得到上清液;
44.(5)吸附柱前处理:在吸附柱中加入500μl洗柱液,室温放置2min,2到8℃,12000rpm离心2min,弃废液;
45.(6)第4步收集的上清液中加入200μl无水乙醇混匀,加入吸附柱并静置2min,12000rpm离心2min,弃废液;
46.(7)向吸附柱中加入600μl漂洗液(使用前请先检查是否已加入无水乙醇),12000rpm离心2min,弃废液;
47.(8)向吸附柱中加入600μl漂洗液,2
‑
8℃,12000rpm离心2min,弃废液;
48.(9)12000rpm离心2min,弃掉收集管,将吸附柱置于室温放置数分钟将吸附柱中残
余的漂洗液去除;
49.(10)将吸附柱放入新管中,向膜中央滴加50
‑
100μl的rnase free ddh2o,室温放置5min,12000rpm室温离心2min,即得到杜梨rna。
50.将提取到的杜梨rna立即于
‑
80℃超低温冰箱中保存以备用。取1
‑
2μl杜梨rna用于琼脂糖凝胶电泳,用nano
‑
drop仪进行检测,检测其浓度为300ng/μl。
51.2、基因扩增
52.取1μg杜梨rna经1u的dnase i 37℃处理30min后立即放入冰上,加入1μl 50mm edta,65℃水浴10min后立即置于冰上。cdna第一链的合成参照toyobo反转录试剂盒操作手册进行,将提取得到的杜梨rna反转录为cdna。所得的第一链cdna用于pbhsf3a基因的扩增。所述pcr扩增体系为:1μl模板cdna、5μl pcr buffer、1μl dntp mix(10mmol/l)、正向引物和反向引物各1μl、0.5μl taq dna聚合酶(5u)和10.5μl无核酶水。
53.正向引物:atgagcccaaaagacaagagcc
54.反向引物:cggatcagtagttttagatgcagac。
55.pcr按以下程序完成:94℃预变性3min;94℃变性30s,58℃退火60s,72℃延伸60s,35个循环,循环完成后72℃延伸10min。
56.3、超表达载体构建
57.凝胶回收选用axyprep dna试剂盒对扩增产物进行回收。然后把纯化的dna片段和中间载体(peasy)进行连接,采用热击法将连接产物转化到大肠杆菌感受态dh5α中,采用北京全式金生物技术有限公司生产的大肠感受态,以目的基因序列引物进行pcr验证并测序(由南京诺唯赞生物科技集团完成)。超表达载体双酶切体系见表1,连接体系见表2。
58.表1双酶切体系
[0059][0060]
表2连接体系
[0061][0062]
将测序结果正确的大肠杆菌,用axyprep质粒dna小量提取试剂(axygen,usa)盒提取质粒,质粒命名为pmv
‑
pbhsf3a。将构建好的测序正确的pmv
‑
pbhsf3a重组载体采用冻融法转入农杆菌感受态细胞(gv3101)备用。
[0063]
实施例2
[0064]
不同逆境条件处理下pbhsf3a基因的qrt
‑
pcr分析
[0065]
为了分析杜梨中pbhsf3a基因对盐(图2中的a)、脱水(图2中的b)、低温(图2中的c)的响应模式,使用real
‑
time pcr技术对pbhsf3a基因的表达模式进行分析。
[0066]
采用试剂盒法提取rna(rna提取方法参照实施例1)。cdna第一链的合成参照北京全式金生物技术有限公司的toyobo反转录试剂盒的操作手册进行。
[0067]
定量pcr检测时,在20μl的反应体系中有:10μl 2
×
mix,0.1μl cdna,5μl引物(以ubiqutin为内参引物,长度为208bp),4.9μl水。
[0068]
定量pcr的程序如表3所示。
[0069]
表3定量pcr程序
[0070][0071]
处理过程如下:
[0072]
从基质中挖出完整的杜梨幼苗,用蒸馏水将根部清洗干净,使用洁净的滤纸吸干多余水分,以保持幼苗根的完整性,将其放入装有蒸馏水的洁净培养瓶中,置于25℃培养室中预培养24小时。将预培养后的杜梨幼苗分别进行脱水、低温、盐处理。每个因素处理时,最少选用20株梨苗,采集叶片的方法为随机混合采样,采完样品后,标记,并快速放入液氮中进行速冻,所采样品均保存在
‑
80℃超低温冰箱。
[0073]
脱水处理:将杜梨幼苗根部水吸干后,置于洁净的滤纸上分别进行脱水处理,取样时间分别为0h、1.5h、6h、9h、12h、24h;
[0074]
低温处理:将预培养后的杜梨材料同组培瓶一起直接放入4℃培养室内进行低温处理,取样时间分别是0h、3h、6h、24h、48h、72h;
[0075]
盐处理:将预培养后的杜梨幼苗放入200mm/l的氯化钠溶液中,取样时间分别为0h、2h、8h、12h、36h、48h;盐采用南京于寿德生物科技有限公司购买的固体氯化钠颗粒,规格:ar500g,相对分子量为58.44。
[0076]
为确定pbhsf3a基因在逆境胁迫中的应答程度,选用荧光定量pcr检测其表达情况。如图2中的a所示,nacl处理植株时,表达量在8h迅速增高至峰值,12h后迅速下降,说明pbhsf3a基因对盐胁迫响应强烈;脱水处理时(图2中的b),表达量在6h时开始升高,9h时达到最高,后逐渐下降;4℃低温处理植株时(图2中的c),表达量在3h时最低,后逐渐上升;
[0077]
可以看出,本发明所述的pbhsf3a基因可以在多种非生物胁迫中发挥作用,对于提高植物抗旱能力、抗盐能力有显著影响,本发明主要探讨该基因在盐胁迫环境中的反应。
[0078]
实施例3
[0079]
pbhsf3a基因亚细胞定位
[0080]
1、亚细胞定位载体的构建
[0081]
根据pbhsf3a基因的核苷酸序列,在基因序列的首尾分别加入两个酶切位点,即sali和bamhi。以测序结果准确无误的目的基因提取质粒为模板,用加入酶切位点的引物对其进行pcr扩增。pcr产物经1%琼脂糖凝胶电泳检测后,将目的条带用凝胶试剂盒回收。回收纯化后的扩增片段并将其克隆到pmd19
‑
t载体中,再转化到大肠杆菌感受态dh5α中。检测转化后的菌液,将检测结果呈阳性的菌液送去测序,提取测序结果准确无误的菌液和质粒。用sali和bamhi将上述两者进行双酶切,将酶切过后的产物pbhsf3a基因和载体分别进行纯化回收。两者于16℃连接过夜,再转化至大肠杆菌感受态dh5α中备用。
[0082]
2、洋葱转化法进行亚细胞定位
[0083]
(1)挑取含pbhsf3a
‑
gfp、gfp和p19农杆菌单克隆于lb培养基中进行活化,置于28℃,220rpm培养箱中培养过夜;
[0084]
(2)取过夜培养物10ul转接到5ml lb
‑
mes培养基中,加2ul 100mm乙酰丁香酮,于28℃,220rpm培养箱中培养16
‑
18h;
[0085]
(3)收集菌液,用10mm mgcl2重悬pbhsf3a
‑
gfp、gfp和p19,od
600
值为0.8时,将目的菌液与p19等体积混合;
[0086]
(4)以500:1的比例在混好的菌液中加入乙酰丁香酮,于室温避光放置3
‑
4h;
[0087]
(5)选择生长状态良好的洋葱,切取第二三层的内表皮置于混好的菌液中侵染10min左右后平铺于ms培养基中,避光放置2
‑
3天后观察验证结果。
[0088]
为了确定pbhsf3a基因的亚细胞定位,以gfp作为对照,细胞质和细胞核中均有表达,而pbhsf3a
‑
gfp只在细胞核中表达,表明pbhsf3a基因的亚细胞定位在细胞核上(图3)。
[0089]
实施例4
[0090]
拟南芥的遗传转化及阳性鉴定
[0091]
1、拟南芥的遗传转化
[0092]
选取30
‑
40天苗龄,生长健壮的拟南芥进行遗传转化,转化前剪掉已开花序。具体操作方法如下:
[0093]
(1)在含抗性的lb液体培养基中对农杆菌pbhsf3a
‑
gfp菌株活化培养,od
600
达0.8
‑
1.0时600rpm离心5min收集菌液;
[0094]
(2)用两倍于菌液体积的1/2ms溶液重悬菌体后加入表面活性剂silwet l
‑
77即得侵染液;
[0095]
(3)将已经剪去花序的盆栽拟南芥倒置于放有侵染液的高压泵中,等压强升至0.6pm,关闭阀门,高压侵染5min;
[0096]
(4)将侵染结束后的拟南芥于暗处放置24h后转入正常的生长环境。
[0097]
2、转基因植株的筛选及鉴定
[0098]
经高压稳定转化法侵染拟南芥之后将得到t0代种子,将t0代种子置于30℃培养箱中干燥至少一周之后又经过4℃低温春化4天打破休眠,消毒后,种于含有潮霉素筛选抗性的培养基中以待观察。具体步骤如下:称量0.1g拟南芥种子置于1.5ml离心管内,于超净工作台内用70%酒精浸泡并吸打种子1min,之后用双蒸水清洗3
‑
5次;用10%次氯酸钠充分振荡消毒5min,弃次氯酸钠溶液,双蒸水清洗4
‑
5次,最后将种子平铺在含50mg/l潮霉素的ms
固体培养基上并时常观察其生长情况,在培养基中可正常生长的拟南芥苗即为转基因阳性苗。待培养皿中的抗性苗长大后可转移到基质营养土中进行下一步验证。
[0099]
称取基质中生长了15d左右的拟南芥叶片0.1g放入2ml离心管中并加入2
‑
3颗钢珠,于放有液氮的盒子内充分振荡叶片至粉末后分别加入500μl ctab溶液(65℃预热),65℃恒温水浴30min,期间上下颠倒5
‑
8次;水浴后待样品冷却至室温后于通风橱内加入500μl三氯甲烷,混匀后12000rpm室温离心10min;之后将上清转移至新的1.5ml离心管中,加入等体积的异丙醇溶液,12000rpm室温离心10min;离心后倒掉上清,切勿将管底白色絮状沉淀掉出,加1ml 75%无水乙醇清洗2次,重悬沉淀,12000rpm离心1min;倒掉上清后待酒精挥发完全后加入50μl双蒸水并置于65℃恒温烘箱30min,待沉淀完全溶解后做胶检测。
[0100]
所提dna与基因特异性引物进行pcr扩增程序,来鉴定阳性苗。反应程序及体系分别依照表4和表5所示。进行检测时,若出现预期大小的片段,且其扩增片段亮度比野生型拟南芥片段高的株系,则为阳性植株。引物设计如下:
[0101]
正向引物f:5
’‑
atgagcccaaaagacaagagcc
‑3’
[0102]
反向引物r:5
’‑
cggatcagtagttttagatgcagac
‑3’
[0103]
载体引物r:5
’‑
cgtcgtccttgaagaagatg
‑3’
[0104]
表4 pcr反应程序
[0105][0106]
表5 pcr反应体系
[0107][0108]
用农杆菌介导方法将pbhsf3a基因稳定转化到拟南芥内,将转化的t0代拟南芥种子经消毒后种在含有终浓度为20mg/l的潮霉素抗性的ms培养基中进行下一步筛选,其中抗性苗长势良好且生长速度快,而非抗性苗则生长缓慢且逐渐黄化并陆续死亡,待阳性苗长长至一定大小后将其移入基质营养土中进行培养。采集t0代拟南芥叶片和野生型拟南芥叶片提取dna并进行阳性鉴定,最终检测到4株阳性植株(图4中的a)。
[0109]
拟南芥的超表达系鉴定:将dna鉴定得到的阳性拟南芥苗进行rna提取和反转录,采用rt
‑
pcr进行超表达分析,以未经处理的野生型拟南芥cdna为阴性对照,用基因特异性引物进行扩增,用拟南芥actin作为内参。扩增结束后用2%琼脂糖凝胶电泳,通过查看目的条带的亮度来判断基因在转化苗中表达量的高低,条带越亮则说明pbhsf3a基因表达量越高(图4中的b)。本实验选择亮度高的,也就是表达量高的line1和line2这两个超表达植物株系(oe5,oe8)作为后期实验的母本拟南芥植株。继续培养超表达系植株并进行收种、烘干。
[0110]
将t1代拟南芥种子经春化作用打破休眠后,消毒并种于含有潮霉素抗生素的ms培养基中以待观察。
[0111]
实施例5
[0112]
pbhsf3a耐盐性分析及相应指标测定
[0113]
1、pbhsf3a转基因植株的抗性分析
[0114]
为验证超表达pbhsf3a基因拟南芥与盐胁迫之间的关系,将烘干后的转基因系(oe5和oe8)与野生型(wt)种子经消毒后用移液枪分别点播于80mm、90mm、100mm、110mm及125mm五种不同浓度的nacl培养基中,期间认真观察并记录种子萌发率。当nacl浓度为90mm时,超表达系萌发的种子要普遍多于野生型萌发的种子(图5中的a),且呈1.5倍的关系(图5中的b),即转基因系种子萌发率约是野生型种子萌发率的1.5倍,初步确定90mm nacl浓度为转基因株系盐胁迫的最适浓度。
[0115]
将转基因系与野生型种子以同样的方法经过消毒后播种于ms固体培养基中,待8
‑
10d后,于超净工作台内挑取几乎一致的拟南芥平铺于五种不同浓度的nacl方形培养基中,并做好标记。经观察统计,当nacl培养基浓度为90mm时转基因株系拟南芥生长速度稍快于野生型拟南芥,且较其他浓度表型差异更为明显(图5中c和d)。结合上述种子萌发率实验,更进一步确定了pbhsf3a转基因株系盐胁迫的最适浓度。
[0116]
确定最适盐浓度后,测定经盐胁迫处理后的转基因和野生型拟南芥的电导率。转基因株系(oe5和oe8)的相对电导率要明显低于野生型株系(图5中的e),说明pbhsf3a转基因株系经盐胁迫处理后受到的损害更小,也即转基因株系的耐盐性要高于野生型株系。
[0117]
2、pbhsf3a转基因植株的耐盐性分析
[0118]
将ms培养基中长至2
‑
4片叶的拟南芥幼苗移入基质营养土中进行培养,生长至30
‑
40d时,停止浇水,具体天数根据拟南芥生长状态来决定,3
‑
5d后改用75mm、90mm、100mm、110mm及125mm五种不同浓度的nacl溶液分组浇灌,且每个浓度设置三个重复进行盐处理10d后观察其表型,并分别测定其细胞中丙二醛含量、叶绿素含量、过氧化氢含量、抗超氧阴离子含量及钠钾离子含量,从而分析其细胞中活性氧的残留量。
[0119]
mda含量的测定:(1)使用南京建成公司的试剂盒,操作步骤如表6所示。操作完成后,于涡旋仪震荡混匀,在95℃以上沸水浴20min,反应完成后,取出,用流水冲洗冷却至室温,准确吸取各管0.25ml反应液转移至新的96孔酶标板中,使用酶标仪在520nm下测定各孔吸光度值(计算时要减去空板读数)。(2)mda含量(nmol/g)=(测定od值
‑
空白od值)/(标准od值
‑
空白od值*标准品浓度(10nmol/ml)/样本浓度(g/ml),注:样本浓度=植物组织重(g)/所加提取液的量(ml)。
[0120]
h2o2含量的测定:(1)使用南京建成公司的试剂盒,操作步骤如表7所示。盖上盖,涡
旋仪震荡混匀,于室温,3000
‑
3500rpm,离心5min,准确吸取0.20ml各管反应液,用移液枪准确地加入到新的96孔酶标板中,使用酶标仪于波长405nm,光径1cm处,测定各孔吸光度值。(2)组织中h2o2含量(mmol/gprot=(测定od值
‑
空白od值)/(标准od值
‑
空白od值*标准品浓度(163mmol/l)/待测样本蛋白浓度(gprot/l)。
[0121]
抗超氧阴离子自由基活性的测定:(1)使用南京建成公司的试剂盒,操作步骤如表8所示。操作完成后用涡旋仪震荡混匀,于室温下静置反应10min,用移液枪吸取200μl各管反应液,将其加入到洁净的96孔酶标板中,于波长550nm处,用酶标仪测定各孔吸光度值。(2)组织中抗超氧阴离子活力单位(u/gprot)=(对照od值
‑
测定od值)/(对照od值
‑
标准od值)*标准品浓度(0.15mg/ml)*1000ml/待测样本蛋白浓度(gprot/l)。
[0122]
na
+
/k
+
的测定:将野生型和转基因植株地上及地下部分放入牛皮袋中密封,并置于65℃恒温烘箱中烘3
‑
5天至恒重;称取烘好的干样0.05g于研钵充分研磨后放入10ml离心管中,加入2ml浓盐酸并于室温下浸提3天;浸提结束后,吸取上清至新的10ml离心管中,用4倍双蒸水进行稀释,每个处理重复3组;采用火焰光度计测定上述浸提液中的钠离子和钾离子,测定时需要配置钠钾混合标准曲线,钾离子和钠离子的母液浓度为5mg/ml。
[0123]
表6 mda含量的测定操作步骤
[0124][0125][0126]
表7 h2o2含量的测定操作步骤
[0127][0128]
表8抗o2‑
活力测定操作步骤
[0129][0130]
转基因株系及野生型拟南芥苗处理前生长状态基本一致,此时每盆拟南芥的光合速率基本一致(图6中的a)。经10天盐胁迫处理之后,每组处理苗均呈现不同程度的萎蔫干枯及叶片黄化现象,当盐浓度为90mm时,野生型拟南芥与转基因拟南芥黄化现象差异最明显,此时光合效率在野生型和转基因拟南芥之间的差异也最明显(图6中的b),转基因拟南芥的叶片生长状态明显好于野生型。与野生型相比,转基因株系的光合效率(fv/fm)呈明显的上升趋势(图6中的c);通过测定盐胁迫10d之后各株系拟南芥的叶绿素后发现,两个转基因株系拟南芥的叶绿素均呈深绿色且差别不大(图6中的e),但叶绿素含量明显高于呈黄绿色的野生型拟南芥(图6中的d)。野生型株系(wt)的电导率更高,且约为转基因株系(oe5和oe8)的1.5倍,说明经盐胁迫后,转基因株系的膜透性增大程度较野生型低,也即转基因株系的膜损伤程度更低(图6中的f);通过测定丙二醛含量发现,转基因株系的丙二醛含量呈明显的降低趋势,约为野生型株系丙二醛含量的三分之一(图6中的g),说明转基因型拟南芥的膜脂过氧化程度更低,细胞膜受到盐胁迫的的损伤更小,通过分析可以得出pbhsf3a转基因株系增强了植株的光合速率,很大程度上提高了植株的耐盐性。
[0131]
3、转基因植株盐胁迫后na
+
/k
+
的测定
[0132]
拟南芥经盐胁迫处理10d之后,转基因株系(oe5和oe8)的钠离子含量与野生型株系(wt)相比相比呈明显的下降趋势,且野生型拟南芥中钠离子含量约为转基因株系的1.4倍左右(图7中的a);而与野生型拟南芥相比,两转基因株系拟南芥的钾离子含量呈上升趋势,且在数据方面转基因型拟南芥的钾离子含量约为野生型的1.5倍(图7中的b);将经盐处理之后拟南芥体内钠钾离子含量相比之后可以发现,转基因拟南芥体内的钠钾比明显低于野生型拟南芥(图7中的c)。由上述实验结果中可以得出,在盐胁迫条件下,pbhsf3a转基因植株为了维持体内钾离子含量的稳态,通过排出体内多余的钠离子,进而提高了植株的耐盐性,尽量维持植株的正常生长。
[0133]
4、转基因植株盐胁迫后活性氧分析
[0134]
对pbhsf3a转基因拟南芥进行二氨基联苯胺(dab)和硝基蓝四唑(nbt)染色,用以表现植株盐胁迫后的活性氧(h2o2和o2‑
)的剩余情况。由于植株活性氧的含量根其氧化胁迫
密切相关,因此植株体内活性氧的清除有利于其更好地生长。
[0135]
将苗龄为30d左右的超表达pbhsf3a基因拟南芥(oe5和oe8)与野生型拟南芥经盐胁迫10d后,选取大小基本一致的叶片放入经磷酸缓冲液溶解的dab(图8中的a)和nbt(图8中的b)染液中进行脱色处理。实验结果表明超表达pbhsf3a基因拟南芥叶片的颜色较野生型拟南芥叶片浅;为了验证dab和nbt染色结果的可靠性,我们采用试剂盒法来测定经盐处理10d的转基因株系(oe5和oe8)与野生型(wt)拟南芥中过氧化氢含量及抗超氧阴离子含量。实验结果表明,两转基因株系拟南芥的过氧化氢含量较野生型拟南芥的低(图8中的c),而抗超氧阴离子含量则明显高于野生型处理苗(图8中的d)。由以上实验结果可以看出,在盐胁迫条件下pbhsf3a转基因拟南芥比野生型拟南芥能更及时消除其体内有毒的氧化物,使得植株受到盐胁迫的损伤更低,从而提高了植株的耐盐性。
[0136]
实施例6
[0137]
pbhsf3a基因沉默杜梨盐胁迫下的抗性鉴定及相应指标测定
[0138]
1、瞬时转化转录因子pbhsf3a的杜梨株系的制备,具体方法如下:选取在人工气候室中生长5周左右的杜梨(pyrus ussuriensis)用于农杆菌侵染。
[0139]
(1)于lb培养基(50mg/l卡那霉素+100mg/l利福平+50mg/l庆大霉素)划线培养带有目的质粒的gv3101农杆菌,挑取农杆菌克隆于5ml lb培养基中28℃培养过夜。
[0140]
(2)测定农杆菌菌液od
600
值之后,3000rpm离心10min收集菌液,弃上清。用乙酰丁香酮(acetosyringone)溶液[10mm mes(ph 5.6)+10mm mgcl2+200um acetosyringone]悬浮,调整至od
600
为0.8
‑
1.0,室温静置3h。
[0141]
(3)将含有目的质粒的农杆菌用于杜梨叶片的注射。
[0142]
(4)用1ml注射器(去掉针头),在梨叶片背面注射,每株杜梨苗选择三片叶子注射即可,标记好注射过的叶片。
[0143]
(5)将注射过的梨植株在室温下避光2
‑
3d后放回人工气候室培养,观察瞬时转化的结果。
[0144]
2、pbhsf3a基因沉默杜梨盐胁迫下的抗性分析
[0145]
为了进一步验证pbhsf3a基因具有正调控植物生长的作用,采用经过病毒诱导的基因沉默技术(vigs),以此抑制杜梨中的pbhsf3a的表达,从而减弱植株的耐盐功能。
[0146]
在进行盐胁迫之前,通过荧光叶绿素表型图可以看出每盆杜梨苗生长状态并无明显差别(图9中的a);经18d盐胁迫处理之后,将pbhsf3a基因沉默掉之后的杜梨苗的生长状态明显差于对照组(wt),也即ptrv
‑
pbhsf3a植株受到的盐胁迫损伤更大(图9中的b)。与对照组(wt)相比,ptrv
‑
pbhsf3a植株的原始光能转化率(fv/fm)稍低(图9中的c),且通过提取每株处理杜梨苗的叶绿素发现,ptrv
‑
pbhsf3a植株(ptrv
‑
1和ptrv
‑
2)的含量均低于wt的叶绿素含量(图9中的d),且ptrv
‑
pbhsf3a植株得叶绿素呈明显的黄绿色,而对照组(wt)呈深绿色(图9中的e)。通过测定盐胁迫后杜梨苗的电导率可以看出,ptrv
‑
2组杜梨苗的电导率最高,约为80%,ptrv
‑
1组杜梨苗的次之,而对照组(wt)处理杜梨苗的电导率最低,约为30%(图9中的f);可以看出,经盐胁迫的条件下,将pbhsf3a基因沉默掉的杜梨苗的膜透性增大,植株受到的损伤更严重。通过测定经盐胁迫18d之后处理杜梨苗的丙二醛含量,可以发现,两组ptrv
‑
pbhsf3a植株的丙二醛含量更高(图9中的g),也即将pbhsf3a基因沉默之后的植株较野生型而言对盐胁迫环境更敏感,受到的损伤也更大。
[0147]
3、pbhsf3a基因沉默杜梨盐胁迫后na
+
/k
+
的测定
[0148]
测定将pbhsf3a基因沉默掉并经盐胁迫处理18d后杜梨体内钠钾离子含量的变化情况。对照组(wt)杜梨苗的钠离子含量最低,ptrv
‑
2组稍高,而ptrv
‑
1组杜梨苗的钠离子含量最高(图10中的a);经18d盐处理后,将pbhsf3a基因沉默掉的杜梨苗体内的钾离子含量呈下降趋势,且ptrv
‑
1组杜梨苗的钾离子含量最低(图10中的b);通过比较三组杜梨苗体内钠钾含量的比值可以看出,ptrv
‑
pbhsf3a杜梨苗(ptrv
‑
1和ptrv
‑
2)的钠钾比含量明显高于对照组(wt)植株(图10中的c)。由上述实验结果可以说明,将pbhsf3a基因沉默掉之后的杜梨苗与野生型杜梨苗在同一盐胁迫环境中对盐更敏感,表现出更低的耐盐性。
[0149]
由上述实施例可知,本发明提供的pbhsf3a基因参与了植物非生物胁迫中盐胁迫的过程,并起着重要的正调控作用。
[0150]
以上所述的实施例仅是对本发明的优选方式进行描述,并非对本发明的范围进行限定,在不脱离本发明设计精神的前提下,本领域普通技术人员对本发明的技术方案做出的各种变形和改进,均应落入本发明权利要求书确定的保护范围内。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1